

Summary
The cellular E3 ubiquitin ligase E6AP (UBE3A) interacts with the cancer associated HPV E6 oncoproteins, where together with the viral E6 oncoprotein it binds and targets the degradation of the p53 tumor suppressor. We find that the HPV-11 E6 protein also associates with E6AP in vivo, and thereby can target the degradation of an E6-associated protein. Mutation of an E6-binding LXXLL peptide motif on E6AP eliminated the association, revealing a common mode of interaction between high and low-risk E6 proteins and E6AP. E6AP was required for the in vivo degradation of DLG1 by both HVP-18 E6 and a chimeric HPV-11 E6. The common functional interaction of both cancer-associated and non-cancer-associated E6 proteins with E6AP establishes a common mechanism for E6 proteins trophic to mucosal squamous epithelium.
Keywords: ubiquitin, proteasome, cervical cancer, respiratory papillomatosis, RRP, genital warts
Introduction
Papillomaviruses are causative agents of benign tumors of cutaneous and mucosal squamous epithelia. A subset of papillomaviruses induce mucosal epithelial tumors that may develop into epithelial malignancies; these types are termed “high-risk” HPV types (prototypes are HPV-16, 18, and 31). Conversely, HPV types that produce only benign mucosal epithelial papillomas are termed “low-risk” types (typified by HPV types 6 and 11) (reviewed in (Munger et al., 2004)). However, low-risk papillomaviruses can produce debilitating benign anogenital or respiratory tumors, accounting for their medical importance (Sinal and Woods, 2005). While considerable effort has established a diverse set of functions for high-risk E6 proteins, little is known about the functions of the low-risk E6 proteins. However, HPV-11 E6 (11E6) is essential for maintenance of the viral DNA at normal copy numbers, and a set of mutants in E6 which results in reduced episomal copy number has been described (Oh, Longworth, and Laimins, 2004).
E6 oncoproteins are small zinc finger proteins that fold to create a peptide binding pocket whose structure has been recently modeled (Nomine et al., 2006). The high risk HPV E6 oncoproteins and E6 from bovine papillomavirus (BE6) interact with cellular proteins by binding to an 8 amino acid peptide displayed on the target protein (XLXXLLXX, abbreviated LXXLL here, with limited substitution of leucine by other hydrophobic amino acids) (Chen et al., 1998; Elston, Napthine, and Doorbar, 1998; Vande Pol, Brown, and Turner, 1998); this interaction is required for cellular transformation by BE6 (Bohl et al., 2000; Vande Pol, Brown, and Turner, 1998). HPV-16 E6 (16E6) interacts with a LXXLL peptide sequence found on the cellular E3 ubiquitin ligase E6AP (termed LQELL in E6AP), and together with E6AP binds to the p53 tumor suppressor protein resulting in ubiquitin-mediated degradation of p53 by the proteosome (Huibregtse, Scheffner, and Howley, 1991; Huibregtse, Scheffner, and Howley, 1993a; Huibregtse, Scheffner, and Howley, 1993b; Werness, Levine, and Howley, 1990). The efficient in vivo degradation of p53 by high-risk E6 requires both E6AP and the E3 ubiquitin ligase activity of E6AP (Cooper et al., 2003), and LXXLL peptides can compete the degradation of p53 in vitro and in vivo (Sterlinko Grm et al., 2004). High-risk E6 proteins also target the degradation of other cellular proteins, particularly cellular PDZ proteins. These PDZ proteins bind the carboxy terminal five amino acids of high risk E6 proteins that is a ligand for PDZ domains (Kiyono et al., 1997; Lee, Weiss, and Javier, 1997). While the cellular PDZ protein scribble has been found to associate with E6AP together with E6 (Nakagawa and Huibregtse, 2000), some evidence has been presented that E6AP does not play a role in the degradation of the DLG1 PDZ domain protein by 18E6 (Pim et al., 2000), but as yet an alternative ubiquitin ligase has not been identified. How the low-risk E6 proteins interact with cellular proteins has not been defined, and binding of low-risk E6 proteins to LXXLL peptides has not been demonstrated (Zanier et al., 2005). However, low-risk E6, like high-risk E6, has the ability to target the degradation of proteins in vitro, as fusion of a PDZ ligand to the carboxy terminus of HPV-11 E6 (11E6) confers upon 11E6 the ability to degrade DLG1 in vitro, similar to high-risk 18E6 (Pim, Thomas, and Banks, 2002). But the ubiquitin ligase by which 11E6 might target proteins for degradation has not been identified.
Results
We noted that E6AP interacted with both 11E6 and 16E6 in yeast two-hybrid assays if the ubiquitin ligase activity of E6AP was inactivated by mutation (in Fig. 1A, E6AP-C843A is mutated in ubiquitin ligase activity). We have previously shown that co-expression of E6 and wild-type E6AP in this yeast system results in loss of detectable E6AP protein while the expression of the ubiquitin-ligase defective E6AP-C843A is retained (Cooper et al., 2003). In the context of the full E6AP protein, 16E6 interacted with both E6AP-C843A containing the intact LQELL motif and the E6AP-C843A-LS point mutant (LQELL mutated to LQELS), but not doubly mutant E6AP-C843A-AS (LQELL mutated to LQEAS) binding site (Fig. 1A). When the E6 binding site of E6AP was displayed as isolated peptides fused to a transactivator, 16E6 would only interact with LQELL but not LQELS (Fig. 1B). 11E6 was more selective in its interaction with E6AP, interacting well with E6AP-C843A (LQELL), weakly with E6AP-C843A-LQELS but not with E6AP-C843A-LQEAS (Fig. 1A). But unlike 16E6, 11E6 did not detectably interact with LQELL or LQELS displayed as peptides outside the context of the full E6AP-C843A protein (Fig. 1B).
Figure 1. 11E6 interacts with E6AP in a yeast two-hybrid assay.
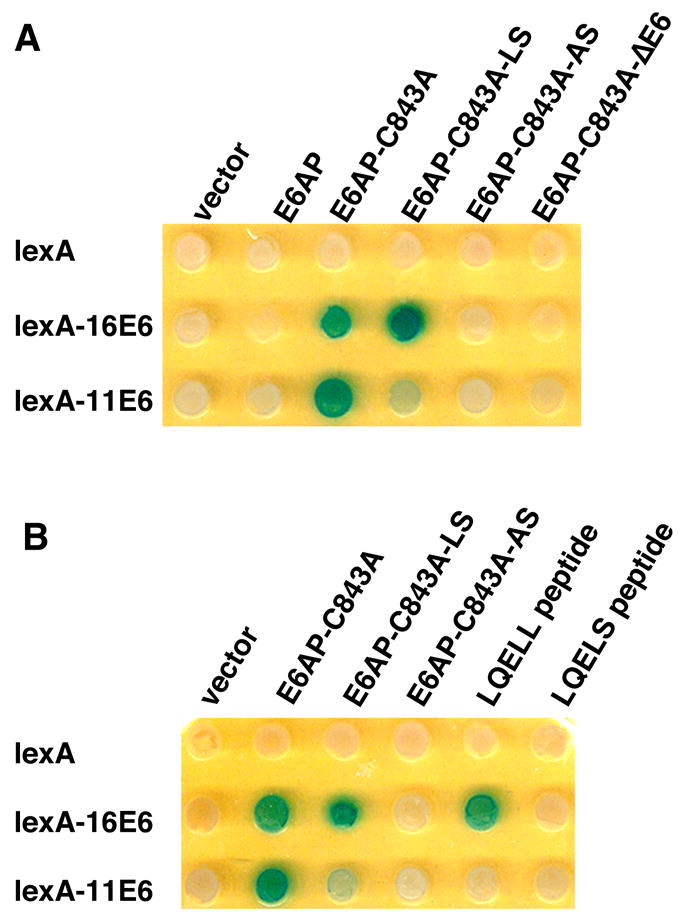
E6AP fusions to the B42 transactivator expressed in the strain YPH500 were introduced by mating to the lexA responsive lacZ and His3 reporter strain TAT7 containing either LexA or LexA fused to 11E6 or 16E6 as previously described (Cooper et al., 2003; Vande Pol, Brown, and Turner, 1998). Diploid yeast were selected on glucose plates by auxotrophic markers and then patched to galactose-XGAL plates to induce expression of the B42 transactivator prey fusion proteins. A. 11E6 binds the same region of E6AP as 16E6. B42 transactivator domain fusions to E6AP, E6AP-C843A (mutated in ubiquitin ligase activity), and E6AP-C843A further mutated in the 16E6-binding site (aa. 409-LQELL-413) LQELS or LQEAS are indicated. E6AP-C843A-ΔE6 contains an in-frame deletion of amino acids 391–408 deleting the entire E6 binding site. Galactose-XGAL plates test for interaction with the indicated lexA fusions with blue color indicating in vivo interaction. B. Interaction of 16E6 but not 11E6 with the isolated E6 binding site of E6AP-C843A. The peptides ELTLQELLGEERR (termed LQELL peptide) or ELTLQELSGEERR (termed LQELS) fused to the HA epitope and the B42 transactivator were tested on Galactose XGAL plates for two-hybrid interaction with the indicated LexA fusions.
To confirm the yeast two-hybrid interactions, E6AP, 16E6 and 11E6 were co-expressed in vivo, immunoprecipitated with antibodies directed to the E6 epitope tags, and analyzed by immuno-blot. 11E6 and 16E6 each associated with E6AP by co-immunoprecipitation (Fig. 2A). Reciprocal co-immune precipitation using the EE epitope tag on E6AP gave the same result (Fig. 2B). Mutation of the ubiquitin ligase activity of E6AP (E6AP-C843A) enhanced the yield of the co-immune precipitated products. Mutation of the LQELL E6 binding motif of E6AP to LQELS caused a partial reduction, and further mutation to LQEAS caused a full loss of co-immune precipitation for both 11E6 and 16E6 (Figs. 2A and B). The association of untagged 11E6 with E6AP was also observed by transient co-expression of the two proteins, purification of E6AP and visualization of both proteins by silver stained gels (see supplemental data S1).
Figure 2. Association of 11E6 and 16E6 with E6AP in mammalian cells. A. Immune precipitation of FLAG-tagged 11E6 and 16E6 co-precipitates E6AP.
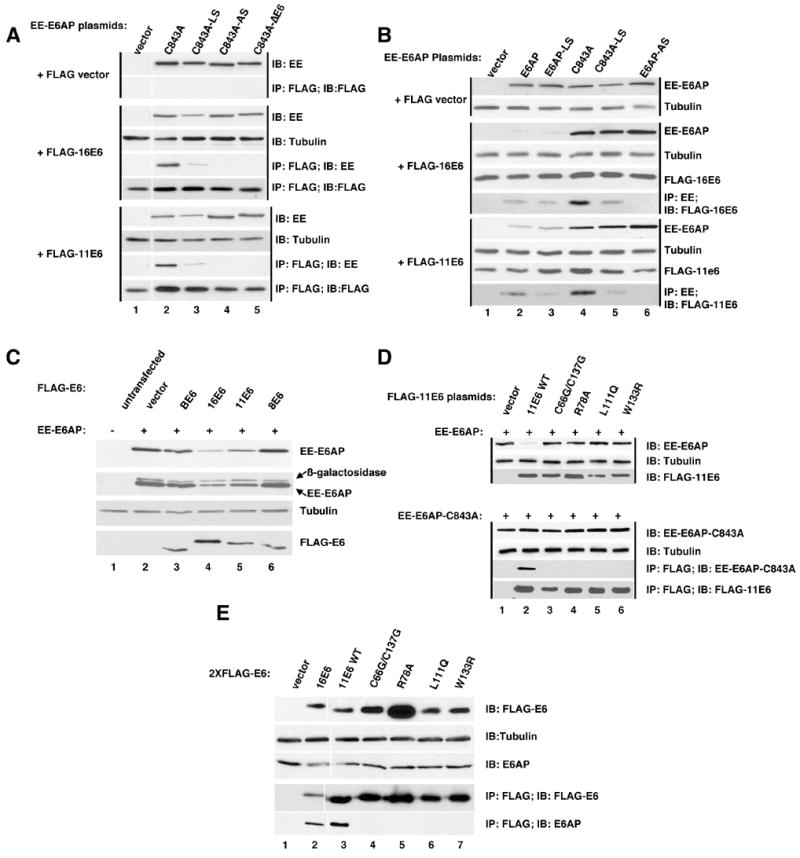
The indicated plasmids were transiently overexpressed in CV-1 cells by vaccinia pTM1 transfection, protein equalized NP40 soluble lysates prepared and either analyzed by immunoblot with the indicated antibodies or immune precipitated with antibodies directed to indicated epitope tags (Flag or EE) as previously described (Cooper et al., 2003). Black vertical lines group samples transfected with either FLAG vector, FLAG-16E6, or FLAG-11E6. A white vertical line between lanes 1 and 2 indicates the position of a lane excised from the figure. B. Immune precipitation of E6AP co-precipitates 11E6 and 16E6. Performed as in A. Black vertical lines group samples transfected with either FLAG vector, FLAG-16E6, or FLAG-11E6. C. Reduction of E6AP expression by co-expression of 11E6 and 16E6. The indicated plasmids were co-expressed in CV-1 cells as in parts A and B with a constant amount of beta-galactosidase included as an internal expression control. Protein normalized samples from NP40 lysed cells were probed with antibodies first for the EE tag on E6AP, then beta-galactosidase, then cellular tubulin as a loading control and finally FLAG-E6. D. 11E6 mutants defective for HPV-11 plasmid maintenance fail to interact with E6AP. In two parallel sets of transfections either EE-E6AP or EE-E6AP-C843A (mutated in ubiquitin ligase activity)were co-transfected with 11E6 wild-type (WT) or the indicated 11E6 mutants. Mutant C66G/C137G are in the zinc binding motif, L111Q and W133R are in conserved buried residues and R78A was undetermined in location in a recent structural model of E6 proteins (Nomine et al., 2006). Cells transfected with EE-E6AP were harvested in SDS lysis buffer and analyzed for expression in the upper three panels. Cells transfected with EE-E6AP-C843A were lysed in NP40 lysis buffer and protein content equalized, with a portion analyzed for EE-E6AP-C843A and tubulin expression in the lysate and the remainder immune precipitated with antibody to FLAG. Immunoblots were performed with the indicated antibodies. E. 11E6 and 16E6 associate with E6AP when stably expressed in C33A cells. C33A cells were retrovirally transduced with 2X-FLAG-tagged vector, 16E6, 11E6 or the indicated 11E6 mutants. 5% of clarified and protein equalized NP40 lysates from 6×107 pooled drug resistant cells 10 passages after completion of drug selection were analyzed by immunoblot for the expression of FLAG-E6 (top panel), tubulin (second panel) and E6AP (third panel from top). The remaining lysate was immune precipitated with rabbit polyclonal antibody to FLAG epitope and washed precipitates were analyzed for the expression of FLAG-E6 and associated E6AP using mouse monoclonal antibodies to FLAG and E6AP.
16E6 co-expression with E6AP has been shown to reduce expression levels of E6AP (Kao et al., 2000). We observed this with 11E6 as well, and it required an intact LQELL motif on E6AP, as mutation of LQELL to LQELS caused a slight restoration of E6AP levels and further mutation to LQEAS a complete restoration of co-expressed E6AP levels (Fig. 2B). Mucosal papillomavirus E6 types 11 and 16 were able upon co-expression to decrease levels of E6AP while E6 proteins from BPV-1 and cutaneous HPV-8 did not (Figs. 2B and 2C). Expression of co-transfected beta-galactosidase was not similarly affected by the expression of E6 proteins (Fig. 2C).
11E6 is necessary for the maintenance of normal copy number of episomal HPV-11 genomes in transfected keratinocytes; that study also characterized a set of 11E6 mutants that each failed to support normal plasmid copy number (Oh, Longworth, and Laimins, 2004). In order to determine if one or more of these 11E6 mutants were defective for association with E6AP or failed to reduce co-expressed E6AP levels, epitope tagged 11E6 and the 11E6 mutants were co-expressed with E6AP-C843A (mutated in ubiquitin ligase activity) and analyzed for association by immune precipitation with antibodies directed against the E6 tags as illustrated in Fig 2D. 11E6 mutants that fail to support HPV-11 episomal copy number failed to co-precipitate with E6AP-C843A (Fig. 2D lower panel), and failed to induce the loss of co-expressed E6AP after transient expression (Fig 2D upper panel). Experiments 2A through D all utilized transient overexpression. In order to determine if 11E6 was associated with endogenous E6AP under stable expression conditions, 2X-FLAG epitope-tagged 16E6, 11E6, and the 11E6 mutants used in Fig. 2D were introduced into HPV-negative C33A cervical cancer cells by retroviral transduction, pooled drug resistant colonies were grown to confluency, and passaged in 1:5 dilutions ten times. Protein-equalized NP40 cell lysates from 6 × 107 cells of each cell line were immune precipitated with antibody to FLAG and analyzed for associated endogenous E6AP. Both FLAG-tagged 16E6 and 11E6 were found to be associated with E6AP while none of the 11E6 mutants that are defective for stable HPV-11 plasmid maintenance were associated with E6AP (Fig. 2E).
E6AP may interact with 11E6 by sensitive immunoblot assays, yet not be a major interacting cellular protein with 11E6. Further, the ability of 16E6 and 11E6 to induce the loss of E6AP when co-expressed might distort the association pattern of E6 with cellular proteins isolated from in vivo co-expression. In order to determine if E6AP is a principle interaction target with 11E6, a tandem affinity purification tag (chitin binding domain fused to a TEV protease site and FLAG epitope tag) was fused to the amino-terminus of either 16E6, 16E6_Y79N (the Y79N mutant is decreased for interaction with E6AP (Cooper et al., 2003)), or 11E6 and expressed in CV1 cells. CBD-TEV-FLAG-E6 fusions in the clarified soluble cell fraction were bound to chitin beads which were recovered by centrifugation but were not washed with detergent containing buffers, and those beads were then added to excess clarified lysate from HaCat cells. After incubation, the chitin beads were recovered, washed with NP40-containing lysis buffer and E6 and any associated proteins were released from the beads by TEV protease cleavage followed by affinity capture with FLAG antibody beads and elution by FLAG peptide. A silver stained gel shows 16E6 and associated products with identifications determined by mass spectrometry (Fig. 3A). Candidate E6AP bands (silver stained bands between 90 to 100 kDa) were excised and the proteins identified by mass spectrometry. A distinct 16E6-associated band at 100 kDa was identified as E6AP that was not retained by 16E6_Y79N, demonstrating that by this assay, E6AP is a major interacting protein with 16E6, as expected, and that the Y79N mutation that is defective for association with E6AP lost association in this assay. A parallel binding assay using 11E6 and 1E6 fusions demonstrated E6AP as a major associated protein for 11E6 but a co-migrating band was not observed with 1E6 (Fig. 3B). This demonstrates that E6AP is preferentially captured by 11E6 from unfractionated cell lysate in a similar fashion as by 16E6.
Figure 3. E6AP association with 16E6 and 11E6 in cellular lysates.
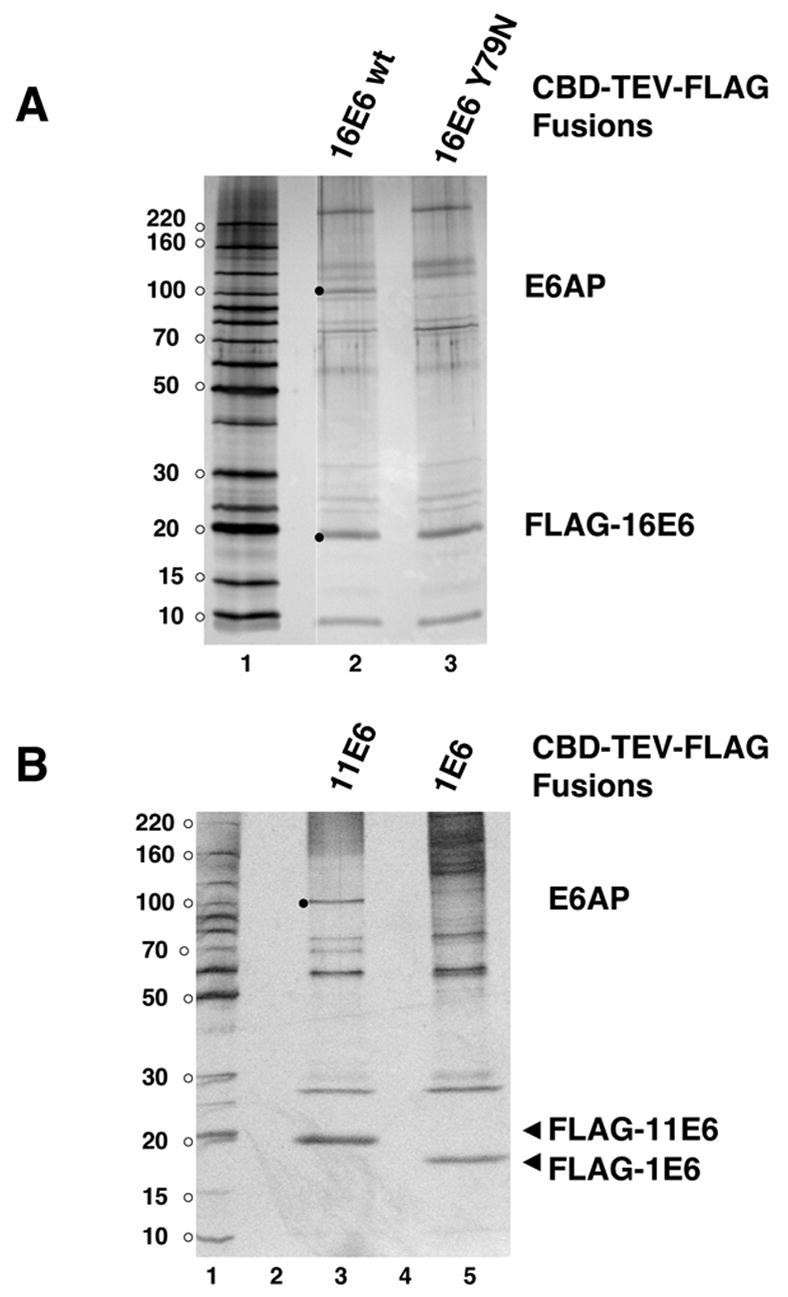
CBD-TEV-FLAG fusions to 16E6, 16E6_Y79N, 11E6, and 1E6 were produced in CV1 cells by vaccinia virus transduction, bound to chitin beads and combined with clarified HaCat cell lysate as described in the methods. FLAG-E6 and associated proteins were released from chitin beads by TEV protease cleavage, and then purified on FLAG antibody beads, washed extensively, eluted by FLAG peptide. A. Association of E6AP with 16E6. Black dots indicate silver-stained bands excised from a 4–20% gradient polyacrylamide gel for protein identification by in-gel trypsinization and LC-MS identification of tryptic peptides. Thirty-one unique peptides in the indicated 100 kDa band in the 16E6 lane were derived from E6AP, while no E6AP peptides were identified in the corresponding gel slice for 16E6_Y79N. Molecular weight markers (lane 1) contain 100 ng protein per band. B. Association of E6AP with 11E6. In vitro binding and in-gel trypsinization performed as in A. The indicated 100 kDa band in lane 3 contained 60 unique peptides derived from E6AP.
E6AP was discovered as an essential cellular co-factor for the targeted degradation of p53 by 16E6 (Huibregtse, Scheffner, and Howley, 1991; Scheffner et al., 1990). Studies have documented that 11E6 fails to associate with (Werness, Levine, and Howley, 1990) or target the degradation of p53 (Scheffner et al., 1990), and as yet cellular proteins that are clearly targeted for degradation by wild-type 11E6 have not yet been identified. However, it was shown that 11E6 could target the degradation of DLG1 in vitro if a seven amino acid PDZ ligand derived from HPV-18 E6 was fused to the carboxy-terminus of 11E6 (this 11E6 mutant is termed here 11.18E6) (Pim, Thomas, and Banks, 2002). In order to determine if E6AP can provide the ubiquitin ligase activity in that assay, E6AP null fibroblasts were co-transfected with EE-tagged DLG1, various E6 genes (FLAG-tagged 11E6, PDZ-ligand-tagged 11E6 (ll.18E6) or 18E6), and wild-type E6AP or E6AP-C843A (Fig. 4). Wild-type 11E6 did not significantly alter the expression levels of DLG1 regardless of co-expressed E6AP or the presence of proteasome inhibitor MG132. In contrast, 18E6 reduced the expression levels of DLG1 only in the presence of co-expressed E6AP (and not E6AP-C843A), and this was largely abrogated by the addition of proteasome inhibitor MG132. This indicates that in fibroblasts, the degradation of DLG1 by 18E6 is dependent upon both E6AP and the proteasome. 11E6 with a PDZ ligand from 18E6 fused to the carboxy-terminus of 11E6 performed similar to 18E6 in that the loss of DLG1 expression was only observed with E6AP (and not with E6AP-C843A) and was reversed by proteasome inhibition (Fig. 4). Expression levels of transfected E6AP were low in this assay and difficult to detect by immunoblot. While both full length FLAG-18E6 as well as a presumed spliced and truncated FLAG-18E6* product were observed, only FLAG-11E6 migrating at the expected full-length size was observed.
Figure 4. E6AP and proteasome-dependent degradation of a synthetic substrate by 11E6.
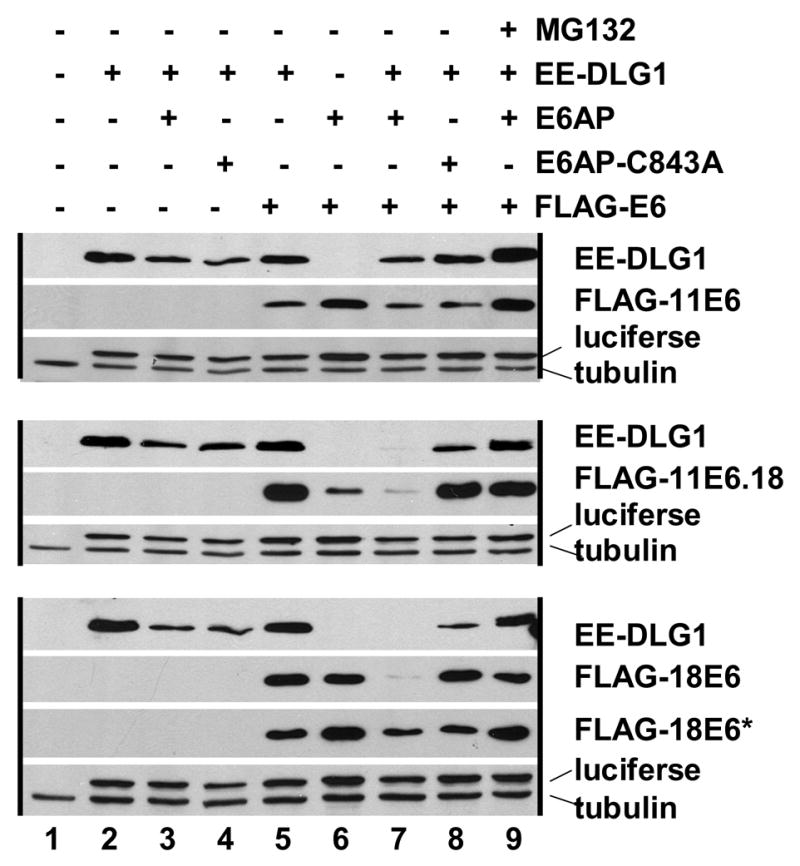
E6AP null mouse embryo fibroblasts were transfected with the indicated plasmids. SDS lysed cells were analyzed by immunoblot 24 hrs later. Black vertical lines group together samples transfected either with FLAG vector, FLAG-11E6, or FLAG-11.18E6 (11E6 with a c-terminal fusion to the 7 amino acid PDZ ligand of 18E6). FLAG-18E6 transfected cells produced two species corresponding to the expected 20 kDa full-length FLAG-18E6 and a smaller indicated band that is presumed to result from an internally spliced 18E6 product termed 18E6*. A constant amount of luciferase was included in the transfection mixture in lanes 2–9 as a transfection efficiency control.
Discussion
Our experiments demonstrate that 11E6, like 16E6, associates with E6AP. The association depends upon an intact LXXLL motif on E6AP (E6AP aa. 408-TLQELLGE-415), and this association can result in the reduction of co-expressed E6AP protein levels and degradation of an 11E6 associated protein (the synthetic interaction of DLG1 with 11.18E6). Bacterially expressed and purified 16E6 directly interacts with LQELL peptides, but this has not been demonstrated in this study, so the possibility remains that an additional cellular protein could function as an adapter, coupling 11E6 to the LQELL motif of E6AP indirectly.
Despite the similarity between 11E6 and 16E6 in interaction with E6AP, several differences were noted. One difference between 11E6 and 16E6 was seen in the ability of 16E6 to interact with the isolated LQELL peptide in yeast, which was not observed with 11E6. In further experiments not shown here, we have observed that 16E6 overexpressed in one cell lysate interacts with E6AP readily when mixed with E6AP expressed in a different cell lysate, but that 11E6 associated less efficiently under these conditions, and was much more efficiently associated with E6AP when co-expressed with E6AP. Further, GST or CBD fusions to 11E6 prepared in bacteria and purified and washed in the presence of non-ionic detergents failed to interact with E6AP at all (our unpublished observations). This may account for the failure of previous studies to identify E6AP or its LQELL peptide as a principle interacting protein with low-risk E6 proteins (Elbel et al., 1997; Zanier et al., 2005).
Previous studies had demonstrated that the expression of 16E6 reduced co-expressed E6AP (Talis, Huibregtse, and Howley, 1998). We observed the same result for 11E6 (Figs. 2B, 2C, 2D, and 4). In addition, mutants of 11E6 that in a previous study failed to support the stable plasmid replication of HPV-11 when expressed in the full context of the HPV genome also in our studies failed to either associate with E6AP or target the degradation of E6AP, despite similar or greater levels of expression (Figs. 2D and E). This supports the biological significance of the association of 11E6 with E6AP.
E6AP participates in the targeted degradation of both p53 and cellular PDZ proteins when associated with high-risk E6 proteins such as 16E6 or 18E6. Using a chimera of 11E6 with the PDZ ligand of 18E6, we observed the targeted degradation of DLG1 in an E6AP and proteasome dependent manner, similar to a parallel experiment using 18E6. Thus, 11E6 association with E6AP resulted in the proteasome-dependent degradation of an additional associated protein. Our result with 18E6 is in contrast to earlier studies using in vitro degradation assays of DLG1 by 18E6, in which depletion of E6AP from reticulocyte lysate by immune precipitation, or inhibition of degradation by various peptide competitors ablated the degradation of p53 but not DLG1. This result prompted the authors to postulate the presence of an additional ubiquitin ligase (not E6AP) that when associated with 18E6 targeted the degradation of DLG1(Grm and Banks, 2004; Sterlinko Grm et al., 2004). Given the differing sensitivity of substrates to in vitro degradation, antibody-mediated depletion studies may have uncertainties if residual E6AP is present. Our genetic results do not completely eliminate the possibility of an alternative ubiquitin ligase to E6AP that is present in reticulocyte lysate but not evident in fibroblasts. However, our results demonstrate that E6AP can provide a ubiquitin ligase activity to target the degradation of DLG1 by 18E6, and that E6AP null fibroblasts do not express sufficient amounts of an alternative ubiquitin ligase to be apparent in this assay.
We demonstrated that E6AP is a principle binding partner of both 11E6 and 16E6 in vitro (Fig. 3). This resulted from competitive binding of total cell lysate to the E6 proteins. Given that E6AP is not an abundant protein, this argues that E6AP is a principle associated protein for both high and low-risk E6 proteins.
This study has not yet identified authentic cellular substrates, if any, that are targeted for degradation by 11E6 together with E6AP, which is the topic of ongoing investigations. It may be that such substrates were not present in the soluble HaCat cell lysate used in Fig. 3. Alternatively, formation of a ternary complex between 11E6, E6AP and a putative cellular protein degradation target may be an active process, unlike with 16E6 where interaction with E6AP and p53 occurs in vitro (Huibregtse, Scheffner, and Howley, 1993b). It is also possible that the 11E6-E6AP complex has a primary role besides the targeted degradation of particular cellular proteins. It is important to note that although we have shown that 11E6 together with E6AP have the capacity to target an associated protein for degradation, there is no evidence that this occurs in the viral life cycle. The concept that 11E6 targets an additional cellular protein for degradation, like 16E6, is at this point an attractive assumption for which there is as yet no clear evidence. It has been proposed that E6AP acts as a transcription factor coactivator for steroid receptors (Nawaz et al., 1999), and association with 11E6 might modulate the role of E6AP as a coactivator. For its proposed role as a steroid receptor family co-activator, the ubiquitin ligase activity was found to be dispensable, indicating E6AP has functions that are not dependent upon the ubiquitin ligase activity, and that the ubiquitin ligase defective mutant used in this study (E6AP-C843A) is not globally defective (Nawaz et al., 1999). Independent investigations of E6AP as a transcription co-activator of the telomerase promoter have not as yet defined the role of the ubiquitin ligase function of E6AP for E6 transactivation of the telomerase promoter (Liu et al., 2005). A recent study has also demonstrated RNAi-mediated knockdown of either high-risk E6 or E6AP in HPV positive cervical cancer cell lines had very similar effects upon the transcriptional profile of the cells, indicating that alterations in transcription by high-risk E6 proteins are predominantly mediated by E6AP in some way (Kelley et al., 2005).
The common association of E6 proteins from both high-risk and low-risk mucosal papillomavirus types with E6AP is a significant if not surprising observation. Given the prevalence and morbidity associated with mucosal papillomavirus infections, modulation of either E6AP function or the E6-E6AP interaction is an attractive target for anti-papillomavirus therapeutics for both cancerous and non-cancerous tumors. Recent experiments have demonstrated that RNAi mediated decrease of E6AP expression in cervical cancer cell lines results in growth suppression and elevation of p53 expression (Hengstermann et al., 2005). Further studies of the association of low-risk E6 proteins with E6AP may reveal a possible role of E6AP in the full virus life cycle.
Materials and Methods
Cells and tissue culture
CV-1 and HaCat cells were maintained in DMEM media supplemented with 10% newborn calf serum, glutamine and antibiotics. E6AP null mouse fibroblasts are derived from primary mouse embryo fibroblasts deleted in both Ube3a (E6AP) genes and are spontaneously immortalized (Jiang et al., 1998). Ube3A null fibroblasts were transfected by Lipofectamine 2000 (Invitrogen) in accordance with the manufacturer’s instructions, and transient Vaccinia virus expression of proteins was performed in CV-1 cells as described (Elroy-Stein, Fuerst, and Moss, 1989).
Plasmids
Human E6AP and E6AP-C843A-(mutated in ubiquitin ligase activity) cDNAs were provided by John Huibregtse (University of Texas, Austin) and are numbered in accordance to Genbank Q05086. E6AP mutants in the E6 binding motif were created by oligonucleotide directed mutagenesis and were fully sequenced and were without polymerase-generated errors. 11E6 with a fusion to the PDZ domain of 18E6 was recreated as described (Pim, Thomas, and Banks, 2002). Epitope tagged plasmids used in mammalian transfection were created in pcDNA3, and retroviral transductions were performed as previously described (Bohl et al., 2000).
Yeast Two Hybrid Assay
The TAT7 strain of yeast was a gift of R. Sternglanz and contains lexA responsive lacZ and His3 reporter genes, and is MATa and is leu2, ade2, and trp1. YPH 500 is mat alpha, ura 3-52, ly2-801, ade2-101, trp1-Δ63, his3-Δ200, leu2Δ1, and is used to introduce plasmids by mating with TAT7. Empty expression plasmids containing appropriate selection markers were introduced into mating strains so that for any particular experiment all selected yeast contained the same number of plasmid types and expressed the same auxotrophic phenotypes. Yeast mating, selection and transfection were as previously described (Vande Pol, Brown, and Turner, 1998). The yeast prey plasmids expressing E6AP, E6AP-C843A fused to the B42 transactivation domain and prey plasmids expressing the E6 binding motifs of E6AP (the isolated E6AP E6 binding motif (amino acids 407–418, LQELL)), or mutated peptide from E6AP (amino acids 407–418, LQELS) been previously described (Cooper et al., 2003), and are separated from the B42 transactivation domain by the hemeagglutinin HA epitope.
Tandem Affinity Purification
The CBD-TEV-FLAG purification cassette (chitin binding domain in frame fused to a TEV (tobacco etch virus protease) cleavage site followed by a FLAG antibody epitope tag) was fused in frame to either 16E6, 16E6_Y79N 11E6, or 1E6 was expressed in confluent monkey CV1 cells by lipofection and T7 polymerase directed expression using the T7-expresssing vaccinia virus Vtf-7 (Elroy-Stein, Fuerst, and Moss, 1989). 24 hrs. after infection, media was removed, cells washed three times with ice cold PBS, and lysed on ice with 0.5X NP40 lysis buffer (1X NP40 lysis buffer contains150mM NaCl; 50mM Tris pH 7.5; 50mM NaF; 5mM NaPPi; 1% IGEPAL; 0.01% phenylmethylsulfonyl flouride; 5 mM EDTA; 1mM sodium vanadate; 1ug/ml leupeptin/aprotinin). Lysates were centrifuged at 15000 × G at 4 °C for 10 minutes. Clarified lysates from approximately 1 × 107 CV1 cells were incubated with 25 ul chitin beads for 30 min. with rocking at 4 °C. The chitin beads were recovered by centrifugation and the cell lysate was remove and replaced by cell lysate from 2×108 HaCat cells lysed in 5 mls 0.5X NP40 lysis buffer, and incubated with rotation at 4 °C for 1 hr. The beads were recovered by centrifugation, and washed 4 times with 1X NP40 lysis buffer and twice by TEV protease cleavage buffer, followed by cleavage of E6 from the beads with 1U TEV according to the manufacturer’s instructions (Invitrogen). The beads were discarded and the supernatant for the TEV cleavage reaction was applied to 20 ug FLAG M2 antibody covalently immobilized on agarose beads for 1 hr at 4 °C. The FLAG beads were washed three times with NP40 lysis buffer, twice with 0.25X NP40 lysis buffer and FLAG-E6 with associated proteins were eluted by three rounds of elution with 2 ug FLAG peptide in 25 ul 0.25X NP40 buffer. Eluates were combined, frozen, lyophilized, and resuspended in SDS PAGE sample buffer and applied to 4–20% Novex polyacrylamide gels. Gels were stained with silver to visualize protein bands. The gel bands were subsequently cut from the gel and destained. The proteins were reduced, alkylated and digested with trypsin in the gel. The peptides formed in the digestion were extracted, concentrated, and characterized by capillary column LC-tandem MS. Database searches were performed using the program SEQUEST.
Western Blot Analysis
0.5X NP40 Cell lysates were equalized for protein content as determined with a commercial kit (Bio-Rad) before electrophoresis; equalized proteins boiled in complete SDS sample buffer were resolved by SDS-PAGE electrophoresis and transferred to PVDF membranes and ponceau stained. Antibody sources: epitope tags were obtained from Sigma Chemicals (M2 FLAG and rabbit polyclonal against the FLAG epitope), Covance (rabbit polyclonal anti-EE epitope), beta-galactosidase (Promega, monoclonal), E6AP (monoclonal, BD Biosystems) and tubulin (Sigma mouse monoclonal). Immune precipitations with FLAG antibody used FLAG-M2 covalently coupled to agarose beads (Sigma).
Acknowledgments
This research was supported by Public Health Service CA-80931 to S.V. from the National Cancer Institute, and by institutional support for the Department of Pathology Collaborative Mass Spectrometry Facility at the University of Virginia.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bohl J, Das K, Dasgupta B, Vande Pol SB. Competitive binding to a charged leucine motif represses transformation by a papillomavirus E6 oncoprotein. Virology. 2000;271(1):163–170. doi: 10.1006/viro.2000.0316. [DOI] [PubMed] [Google Scholar]
- Chen JJ, Hong Y, Rustamzadeh E, Baleja JD, Androphy EJ. Identification of an alpha helical motif sufficient for association with papillomavirus E6. J Biol Chem. 1998;273(22):13537–44. doi: 10.1074/jbc.273.22.13537. [DOI] [PubMed] [Google Scholar]
- Cooper B, Schneider S, Bohl J, Jiang Y, Beaudet A, Vande Pol S. Requirement of E6AP and the features of human papillomavirus E6 necessary to support degradation of p53. Virology. 2003;306(1):87–99. doi: 10.1016/s0042-6822(02)00012-0. [DOI] [PubMed] [Google Scholar]
- Elbel M, Carl S, Spaderna S, Iftner T. A comparative analysis of the interactions of the E6 proteins from cutaneous and genital papillomaviruses with p53 and E6AP in correlation to their transforming potential. Virology. 1997;239(1):132–49. doi: 10.1006/viro.1997.8860. [DOI] [PubMed] [Google Scholar]
- Elroy-Stein O, Fuerst TR, Moss B. Cap-independent translation of mRNA conferred by encephalomyocarditis virus 5’ sequence improves the performance of the vaccinia virus/bacteriophage T7 hybrid expression system. Proc Natl Acad Sci U S A. 1989;86(16):6126–6130. doi: 10.1073/pnas.86.16.6126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elston RC, Napthine S, Doorbar J. The identification of a conserved binding motif within human papillomavirus type 16 E6 binding peptides, E6AP and E6BP. J Gen Virol. 1998;79(Pt 2):371–4. doi: 10.1099/0022-1317-79-2-371. [DOI] [PubMed] [Google Scholar]
- Grm HS, Banks L. Degradation of hDlg and MAGIs by human papillomavirus E6 is E6-AP-independent. J Gen Virol. 2004;85(Pt 10):2815–9. doi: 10.1099/vir.0.80035-0. [DOI] [PubMed] [Google Scholar]
- Hengstermann A, D’Silva MA, Kuballa P, Butz K, Hoppe-Seyler F, Scheffner M. Growth suppression induced by downregulation of E6-AP expression in human papillomavirus-positive cancer cell lines depends on p53. J Virol. 2005;79(14):9296–300. doi: 10.1128/JVI.79.14.9296-9300.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huibregtse JM, Scheffner M, Howley PM. A cellular protein mediates association of p53 with the E6 oncoprotein of human papillomavirus types 16 or 18. Embo J. 1991;10(13):4129–35. doi: 10.1002/j.1460-2075.1991.tb04990.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huibregtse JM, Scheffner M, Howley PM. Cloning and expression of the cDNA for E6-AP, a protein that mediates the interaction of the human papillomavirus E6 oncoprotein with p53. Mol Cell Biol. 1993a;13(2):775–784. doi: 10.1128/mcb.13.2.775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huibregtse JM, Scheffner M, Howley PM. Localization of the E6-AP regions that direct human papillomavirus E6 binding, association with p53, and ubiquitination of associated proteins. Mol Cell Biol. 1993b;13(8):4918–27. doi: 10.1128/mcb.13.8.4918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang YH, Armstrong D, Albrecht U, Atkins CM, Noebels JL, Eichele G, Sweatt JD, Beaudet AL. Mutation of the Angelman ubiquitin ligase in mice causes increased cytoplasmic p53 and deficits of contextual learning and long-term potentiation [see comments] Neuron. 1998;21(4):799–811. doi: 10.1016/s0896-6273(00)80596-6. [DOI] [PubMed] [Google Scholar]
- Kao WH, Beaudenon SL, Talis AL, Huibregtse JM, Howley PM. Human papillomavirus type 16 E6 induces self-ubiquitination of the E6AP ubiquitin-protein ligase. J Virol. 2000;74(14):6408–17. doi: 10.1128/jvi.74.14.6408-6417.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley ML, Keiger KE, Lee CJ, Huibregtse JM. The global transcriptional effects of the human papillomavirus E6 protein in cervical carcinoma cell lines are mediated by the E6AP ubiquitin ligase. J Virol. 2005;79(6):3737–47. doi: 10.1128/JVI.79.6.3737-3747.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiyono T, Hiraiwa A, Fujita M, Hayashi Y, Akiyama T, Ishibashi M. Binding of high-risk human papillomavirus E6 oncoproteins to the human homologue of the Drosophila discs large tumor suppressor protein. Proc Natl Acad Sci U S A. 1997;94(21):11612–6. doi: 10.1073/pnas.94.21.11612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SS, Weiss RS, Javier RT. Binding of human virus oncoproteins to hDlg/SAP97, a mammalian homolog of the Drosophila discs large tumor suppressor protein. Proc Natl Acad Sci U S A. 1997;94(13):6670–5. doi: 10.1073/pnas.94.13.6670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Yuan H, Fu B, Disbrow GL, Apolinario T, Tomaic V, Kelley ML, Baker CC, Huibregtse J, Schlegel R. The E6AP ubiquitin ligase is required for transactivation of the hTERT promoter by the human papillomavirus E6 oncoprotein. J Biol Chem. 2005;280(11):10807–16. doi: 10.1074/jbc.M410343200. [DOI] [PubMed] [Google Scholar]
- Munger K, Baldwin A, Edwards KM, Hayakawa H, Nguyen CL, Owens M, Grace M, Huh K. Mechanisms of human papillomavirus-induced oncogenesis. J Virol. 2004;78(21):11451–60. doi: 10.1128/JVI.78.21.11451-11460.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakagawa S, Huibregtse JM. Human scribble (Vartul) is targeted for ubiquitin-mediated degradation by the high-risk papillomavirus E6 proteins and the E6AP ubiquitin-protein ligase. Mol Cell Biol. 2000;20(21):8244–53. doi: 10.1128/mcb.20.21.8244-8253.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawaz Z, Lonard DM, Smith CL, Lev-Lehman E, Tsai SY, Tsai MJ, O’Malley BW. The Angelman syndrome-associated protein, E6-AP, is a coactivator for the nuclear hormone receptor superfamily. Mol Cell Biol. 1999;19(2):1182–9. doi: 10.1128/mcb.19.2.1182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomine Y, Masson M, Charbonnier S, Zanier K, Ristriani T, Deryckere F, Sibler AP, Desplancq D, Atkinson RA, Weiss E, Orfanoudakis G, Kieffer B, Trave G. Structural and functional analysis of E6 oncoprotein: insights in the molecular pathways of human papillomavirus-mediated pathogenesis. Mol Cell. 2006;21(5):665–78. doi: 10.1016/j.molcel.2006.01.024. [DOI] [PubMed] [Google Scholar]
- Oh ST, Longworth MS, Laimins LA. Roles of the E6 and E7 proteins in the life cycle of low-risk human papillomavirus type 11. J Virol. 2004;78(5):2620–6. doi: 10.1128/JVI.78.5.2620-2626.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pim D, Thomas M, Banks L. Chimaeric HPV E6 proteins allow dissection of the proteolytic pathways regulating different E6 cellular target proteins. Oncogene. 2002;21(53):8140–8. doi: 10.1038/sj.onc.1206026. [DOI] [PubMed] [Google Scholar]
- Pim D, Thomas M, Javier R, Gardiol D, Banks L. HPV E6 targeted degradation of the discs large protein: evidence for the involvement of a novel ubiquitin ligase. Oncogene. 2000;19(6):719–25. doi: 10.1038/sj.onc.1203374. [DOI] [PubMed] [Google Scholar]
- Scheffner M, Werness BA, Huibregtse JM, Levine AJ, Howley PM. The E6 oncoprotein encoded by human papillomavirus types 16 and 18 promotes the degradation of p53. Cell. 1990;63(6):1129–36. doi: 10.1016/0092-8674(90)90409-8. [DOI] [PubMed] [Google Scholar]
- Sinal SH, Woods CR. Human papillomavirus infections of the genital and respiratory tracts in young children. Semin Pediatr Infect Dis. 2005;16(4):306–16. doi: 10.1053/j.spid.2005.06.010. [DOI] [PubMed] [Google Scholar]
- Sterlinko Grm H, Weber M, Elston R, McIntosh P, Griffin H, Banks L, Doorbar J. Inhibition of E6-induced degradation of its cellular substrates by novel blocking peptides. J Mol Biol. 2004;335(4):971–85. doi: 10.1016/j.jmb.2003.10.079. [DOI] [PubMed] [Google Scholar]
- Talis AL, Huibregtse JM, Howley PM. The role of E6AP in the regulation of p53 protein levels in human papillomavirus (HPV)-positive and HPV-negative cells. J Biol Chem. 1998;273(11):6439–45. doi: 10.1074/jbc.273.11.6439. [DOI] [PubMed] [Google Scholar]
- Vande Pol SB, Brown MC, Turner CE. Association of Bovine Papillomavirus Type 1 E6 oncoprotein with the focal adhesion protein paxillin through a conserved protein interaction motif. Oncogene. 1998;16(1):43–52. doi: 10.1038/sj.onc.1201504. [DOI] [PubMed] [Google Scholar]
- Werness BA, Levine AJ, Howley PM. Association of human papillomavirus types 16 and 18 E6 proteins with p53. Science. 1990;248(76):76–9. doi: 10.1126/science.2157286. [DOI] [PubMed] [Google Scholar]
- Zanier K, Charbonnier S, Baltzinger M, Nomine Y, Altschuh D, Trave G. Kinetic analysis of the interactions of human papillomavirus E6 oncoproteins with the ubiquitin ligase E6AP using surface plasmon resonance. J Mol Biol. 2005;349(2):401–12. doi: 10.1016/j.jmb.2005.03.071. [DOI] [PubMed] [Google Scholar]