

Abstract
Males of the moth Cosmosoma myrodora (Arctiidae) acquire pyrrolizidine alkaloid by feeding on the excrescent fluids of certain plants (for instance, Eupatorium capillifolium). They incorporate the alkaloid systemically and as a result are protected against spiders. The males have a pair of abdominal pouches, densely packed with fine cuticular filaments, which in alkaloid-fed males are alkaloid laden. The males discharge the filaments on the female in bursts during courtship, embellishing her with alkaloid as a result. The topical investiture protects the female against spiders. Alkaloid-free filaments, from alkaloid-deprived males, convey no such protection. The males also transmit alkaloid to the female by seminal infusion. The systemic alkaloid thus received, which itself may contribute to the female's defense against spiders, is bestowed in part by the female on the eggs. Although paternal contribution to egg defense had previously been demonstrated for several arctiid moths, protective nuptial festooning of a female by its mate, such as is practiced by C. myrodora, appears to be without parallel among insects.
Keywords: pyrrolizidine alkaloid, paternal investment, predation, Arctiidae, Cosmosoma myrodora
Here we describe the behavior of a moth in which acquisition of pyrrolizidine alkaloid (henceforth, for simplicity, sometimes called alkaloid) plays a crucial role in defense. We demonstrate that the alkaloid is initially procured by the male from plants, and that the male derives protection against spiders (Nephila clavipes) as a result. We demonstrate further that the male allocates some of the acquired alkaloid to a cottony mass of cuticular filaments that he keeps tucked away in the abdomen and that he discharges on the female in bursts during courtship. We show that the alkaloid-laden filaments thus bestowed by the males protect the female against N. clavipes, whereas alkaloid-free filaments from control males that had no access to alkaloid provide no such protection. We show further that the female receives additional alkaloid from the male through seminal infusion, an endowment that can itself contribute to her defense and that she shares with the eggs. The moth, Cosmosoma myrodora, known as the scarlet-bodied wasp moth, is a stunningly beautiful member of the family Arctiidae (Fig. 1A) (1).
Figure 1.

(A) Cosmosoma myrodora (male) on its larval foodplant (Mikania scandens). (B) Courting Cosmosoma in wind tunnel; the male (Left) has just ejected flocculent, which is visible as a diffuse misty cloud. (C) Ventral view of male; the flap that covers the abdominal pouches is the area between the white markings at base of abdomen. (D) Same, with flocculent being teased out with forceps. (Bar = 1 cm.)
The study was undertaken at a field site, the Archbold Biological Station, Lake Placid, Highlands County, FL, and in our laboratories.
Henceforth we refer to the moth by its generic designation. For the male's mass of filaments, we adopt the term flocculent.
Materials and Methods
Cosmosoma.
Field-collected moths were obtained at the Archbold Station by baiting with air-dried roots of Eupatorium capillifolium (Asteraceae), a plant that we found to contain pyrrolizidine alkaloids (see below). We had previously noted Cosmosoma to be attracted to such bait. The roots were hung in clusters, about 1.5 m above the ground, and were kept moistened. They were inspected intermittently throughout the night for visiting Cosmosoma that had alighted on them. Additional Cosmosoma were captured at a mercury vapor light trap. Offspring of field-collected females were raised in our Wake Forest laboratories on their natural larval foodplant, Mikania scandens (Asteraceae), providing the bulk of individuals on which our observations were made.
Flocculent.
The pouches in the male that contain the flocculent are two invaginations of the ventral surface of the abdomen (spanning sternites II–IV), ordinarily concealed by a flap (Fig. 1C). The flocculent can be readily removed by teasing with forceps (Fig. 1D). For scanning electron microscopy, portions of flocculent were air dried and gold coated.
Plant Analyses.
Air-dried powdered plant material [M. scandens leaves
(50 g) or E. capillifolium roots (80 g)] was extracted in a
Soxhlet apparatus with methanol for 24 h. After filtration, the
solvent was removed in vacuo, and the residue was
partitioned between dichloromethane and 0.5 N aqueous HCl. The aqueous
phase was separated, basified by the addition of concentrated aqueous
NH3, and extracted with dichloromethane. After
evaporation and filtration, the extract was analyzed by HPLC-MS by
using a Hewlett–Packard 1090 II pump linked to a Micromass
(Manchester, U.K.) Quattro I mass spectrometer, operated in positive
ion electrospray mode with a C18 reversed-phase
column (Metachem 250 × 4.6 mm Inertsil 5 μ ODS-3; solvent
water/acetonitrile, 9:1). For analyses of the plant's N-oxide
content, the aqueous phase was acidified with 2 N HCl and treated with
Zn dust for 24 h at 25°C. After filtration and adjusting to pH 9
by addition of concentrated aqueous NH3, the
solution was extracted repeatedly with a mixture of dichloromethane and
ethanol (9:1). Evaporation yielded 55 mg (E. capillifolium)
and 3 mg (M. scandens) of yellowish oils, which were
analyzed by HPLC-MS as described above. The E. capillifolium
extract was then chromatographed over a short silica column (length 5
cm) by using a mixture of methanol and dichloromethane (1:6) containing
2% aqueous NH3 as a solvent, which yielded 29.5
mg of a 1:3 mixture of two diastereomeric pyrrolizidine alkaloids,
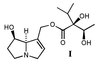
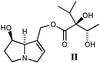
identified by NMR spectroscopy as intermedine (I) and
lycopsamine (II), respectively (ref. 2;
Schemes 1 and 2).
Qualitative Alkaloid Analyses.
Flocculent (teased from the abdominal pouches of live males) and air-dried bodies of Cosmosoma males and females were extracted with dichloromethane for 12 h at 25°C. After filtration, the extracts were analyzed by GC by using a Hewlett–Packard 5890 gas chromatograph equipped with a J & W (J & W Scientific, Folsom, CA) DB5-coated capillary column and by HPLC by using a Hewlett–Packard 1090 Series II instrument equipped with a diode array detector and a C18 reversed-phase column (C-18 BDS Hypersil column, 250 × 4.6 mm, 5-μm particle size) eluted at 1 ml/min with a phosphate buffer/acetonitrile mixture (92:8).
Quantitative Alkaloid Analyses.
Cosmosoma adults were extracted overnight with 500 μl of a phosphate buffer solution (2.7 g potassium dihydrogenphosphate/2 ml triethylamine/0.4 ml trifluoroacetic acid in 4 liters of water; pH adjusted to 4.0 with phosphoric acid). After centrifugation, the residue was reextracted with additional buffer solution (200 μl). The combined extracts were filtered (0.45-μm filter membrane) and analyzed by HPLC by using the HPLC system described above. Egg and flocculent samples were extracted similarly by using 200 μl of phosphate buffer solution. The alkaloid riddelline served as the internal standard. The calibration curves used in the analyses (r2 0.9993 and 0.9924 for adults and eggs, respectively) were constructed by using either monocrotaline and monocrotaline N-oxide or the 1:3 mixture of intermedine and lycopsamine isolated from the roots of E. capillifolium. Unless otherwise specified, the analytical alkaloid values reported herein represent the sum of both the free-base and N-oxide content.
Presentation of Alkaloidal Offerings.
Individual laboratory-raised Cosmosoma males were presented
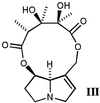
with a crystalline offering of either monocrotaline
(III) (500, 750, or 2,500 μg; N-oxide form), a
pyrrolizidine alkaloid available to us in quantity, or of the 1:3
intermedine/lycopsamine mixture (500 or 750 μg) extracted from
E. capillifolium. The samples were delivered in ethanolic
solution onto depression slides and after evaporation of the solvent
were enriched by addition of 50 μl of a 5% sucrose solution. A slide
was presented to a male for an hour a day on each of the 5 days (or
fewer days when so specified) after adult emergence (a fresh 50 μl of
the sugar solution was added daily to the sample). Males readily fed on
these samples (Scheme 3).
Mating.
Cosmosoma males and females were confined overnight in pairs in cages (32-liter Styrofoam containers covered with tulle). Copulation typically was initiated at dusk and lasted up to 9 h. Some of the matings were obtained by pairing prospective mates in the wind tunnel.
Courtship.
This behavior was videotaped (JVC BR9000U, Yokahama, video recorder) in a laboratory wind tunnel (60 × 60 × 150 cm; 25 cm/s wind speed). The video camera (GBC CCD-500E) was prefocused on the stationary “calling” (that is, pheromone-emitting) female. Illumination (<5 lux) was provided by red-filtered fluorescent lights.
Predation Tests.
These were carried out with the orb-weaving spider, N. clavipes in accord with a protocol previously used with other moths (3). Individual Cosmosoma were flipped from vials into the web of an individual spider, where they became entangled, and a record was kept of whether the spider ate the moth or freed it by cutting it from the web.
Experimental Application of Flocculent.
Individual virgin females (1–2 days old) were placed in 1-oz plastic containers (creamer cups) into which had been introduced flocculent (approximately half the amount present in the pouches) from a 4-day-old virgin male. The females invariably fluttered on being confined, and in the process stirred up the flocculent and coated themselves with filaments. Experimental females received flocculent from males that had fed on an alkaloid sample (500 μg of the intermedine/lycopsamine mixture presented at age of 2–3 days). Control females received flocculent from males that had no access to alkaloid. All females were presented to N. clavipes within minutes after being treated.
Statistics.
Data are expressed as mean ± standard error.
Results
Absence of Alkaloid in Larval Foodplant.
Samples of leaves of M. scandens that were analyzed failed to reveal presence of pyrrolizidine alkaloids. Such alkaloids are therefore available to Cosmosoma only as adults from an exogenous source.
Presence of Alkaloids in E. capillifolium.
The isolation of the two alkaloids, intermedine and lycopsamine, from the roots of E. capillifolium came as no surprise. Structurally related alkaloids had been reported previously from other species of Eupatorium (for example, E. fortunei and E. cannabium) (4, 5). In E. capillifolium roots, the two alkaloids occurred mostly (over 90%) in the form of their N-oxides.
Intermedine, lycopsamine, and their respective N-oxides make up about 95% of the alkaloid content of E. capillifolium roots. In addition, smaller amounts of four structurally related pyrrolizidine alkaloids are present, two of which to our knowledge have not been described from elsewhere in nature (F.C.S., unpublished results).
Acquisition of Alkaloid from E. capillifolium.
The sample of 54 Cosmosoma that were collected at the E. capillifolium baits included only one female. Alighted males actively probed the root surface with their proboscis, indicating that they were feeding, or attempting to feed, from excrescent material.
Captured males that were analyzed (n = 6) proved to contain intermedine and lycopsamine, the two principal pyrrolizidine alkaloids in E. capillifolium, in the ratio of 1:3. Whereas in the plant the alkaloids occurred predominantly in the N-oxide form, they occurred in both oxidation states in Cosmosoma. Furthermore, the ratio of N-oxide to free base of the two alkaloids varied greatly from moth to moth (on average, the ratio was 1:1). Cosmosoma can evidently convert pyrrolizidine alkaloids metabolically from one oxidation state to the other, as has been shown to be the case for another pyrrolizidine alkaloid-ingesting moth (6).
Most of the males attracted to E. capillifolium were in pristine condition, indicating that they might have been on their first visit to the plant. But occasional older males of faded appearance were also noted.
Fate of Ingested Alkaloid.
Six males that fed on monocrotaline N-oxide were checked for alkaloid content by HPLC. The bodies were analyzed separately from the flocculent, which was teased out of the abdominal pouches with forceps. The bodies of these moths contained 127 ± 36 μg alkaloid per male (61% N-oxide), whereas the flocculent contained 13 ± 3 μg alkaloid per male (79% free base). Given that the flocculent amounts to only 0.5% (330 ± 60 μg) of the total mass of the moth (67 ± 7 mg), this means that the alkaloid was about 20 times more concentrated in the flocculent than in the moth's body. The flocculent also contained traces of linoleic acid and linolenic acid, as indicated by GC and NMR spectroscopy.
Courtship.
In the wind tunnel, males flew upwind toward the stationary female, indicating that, as is so often the case with moths (7), the female is the source of a sex-attractant pheromone. On arrival beside a female, the male did not at once alight but continued fluttering while remaining at close range. Abruptly then, and while still on the wing, it discharged the flocculent, enshrouding the female in what had the appearance of an explosively delivered aerial net. This filamentous flotsam did not all at once close on the female but diffused toward her slowly, like a drifting cloud of mist (Fig. 1B). Up to three consecutive emissions were delivered by individual males, as a consequence of which the females were left conspicuously “decorated.” Filaments clung to all her body parts and were particularly noticeable on the antennae (Fig. 2B). After delivery of the flocculent, the males usually attempted promptly to copulate.
Figure 2.

(A) Isolated filaments from flocculent. (B) Portion of antenna of female Cosmosoma, beset with flocculent filaments. (C and D) Enlarged view of filament (stereo pair). (E) Portion of inner lining of abdominal pouch of male, showing the sockets in which the filaments are presumably initially implanted. (Bars = A, 10 μm; B, 100 μm; E, 1 μm.)
Not all males released flocculent on encountering the female. Of 45 males that were tested (and that were presented beforehand with 0–2,500 μg monocrotaline N-oxide), 28 delivered flocculent, and 17 failed to do so. The former group was significantly more successful in securing copulations (57%) than the latter group (23.5%) (P < 0.05, χ2 test).
The Flocculent.
When we prodded open the ventral pouches of a male, the flocculent, which is ordinarily densely packed within, tended to bulge out. Further prodding may cause clusters of filaments to break loose and to float away on air. The filaments tend to stick to foreign objects, but they readily separate from one another. To effect their mass expulsion, the male presumably needs do no more than partially evert the pouches by compressing them from within.
Anatomically, the filaments may represent modified scales. Indeed, the cuticular membrane lining the pouches bears pores (Fig. 2E) that may be the openings of the sockets in which the filaments were originally implanted.
The filaments are elaborately structured (Fig. 2 A–D). Flattened rather than cylindrical, and highly sculpted, they seem to be built for lightness, flexibility, and strength. The sculpting also ensures that a vast surface is provided for dissemination of the alkaloid.
Nuptial Transfer of Alkaloids.
Females that mated with alkaloid-fed males acquired alkaloid systemically from their partners. We analyzed females (n = 8) that as virgins had mated with males that had fed on the intermedine/lycopsamine mixture (750 μg). We found all to contain alkaloid but in amounts that differed depending on whether they had laid eggs. Those that had not oviposited (n = 5) contained 310 ± 74 μg alkaloid, whereas those that oviposited (n = 3) contained only 82 ± 15 μg alkaloid. Not only had the females received a nuptial gift of alkaloid, presumably by seminal infusion, but it seemed that they had passed some of the chemical onto the eggs. We also analyzed the male partners (n = 8) of these females after mating and found them to contain 369 ± 49 μg alkaloid. The males could therefore be expected to have contained on the order of 680 μg of alkaloid before mating, almost the full amount offered to them on the depression slide.
Alkaloid Transfer to Eggs.
We analyzed three egg clusters from three separate females that each had been mated as virgins to males that had fed on alkaloid (750 μg, intermedine/lycopsamine mixture). The average alkaloid content per egg for the 3 clusters was: 1.92 μg (32 eggs), 0.36 μg (50 eggs), and 1.29 μg (65 eggs). An egg cluster from a control female that had been mated to an alkaloid-deprived male proved to be alkaloid free.
Predation Tests: Males.
Males (all virgin) that we offered to N. clavipes fared differently depending on whether they had imbibed alkaloid (Fig. 3). Seven males that had fed on individual monocrotaline N-oxide samples (500 μg) were all cut loose by the spiders. They were freed live and uninjured. In sharp contrast, five of seven males that had had no access to alkaloid were eaten.
Figure 3.

Fate of Cosmosoma, offered to N. clavipes spiders. Males: (−) alkaloid free; (+) alkaloid fed. Females: (V) virgin, alkaloid free; (M−) mated with alkaloid-free male; (M+) mated with alkaloid-fed male; (VF−) virgin, festooned with flocculent of alkaloid-free male; (VF+) virgin, festooned with flocculent of alkaloid-fed male. Statistics: *, P < 0.05; **, P < 0.01 (Fisher Exact Probability).
Predation Tests: Females.
Females, too, fared differently vis à vis N. clavipes, depending on whether they contained alkaloid (Fig. 3). Seven females mated as virgins to alkaloid-bearing males (males that had fed on 500 μg of the intermedine/lycopsamine mixture) were all cut from the web by the spiders, without sustaining injury. Five of six females that had mated with alkaloid-free males were eaten. Five of seven virgin females were eaten as well.
Predation Tests: Flocculent-Treated Females.
Eight virgin females that were experimentally coated with flocculent from males that had no access to alkaloid were eaten by N. clavipes. In contrast, over half (5 of 9) of the virgin females that were coated with flocculent from males that had fed on 500 μg of the intermedine/lycopsamine mixture were set free, uninjured by the spiders (Fig. 3).
Discussion
The defensive role of pyrrolizidine alkaloids in the life of Cosmosoma seems established, certainly vis à vis spiders. The adult male, after acquiring alkaloid, was rejected by N. clavipes, as was the female after receipt of alkaloid from her mate. The eggs too can be expected to be protected by their alkaloidal endowment. The amount of alkaloid they contain, on the order of 1 μg per egg, was shown to be effective in egg defense against ants, coccinellid beetles, and chrysopid larvae in another arctiid moth, Utetheisa ornatrix (8–10). There is also no question that N. clavipes is a natural threat to Cosmosoma. In central Florida, where we made our observations, the spider and moth coexist, particularly at the edge of lakes and streams. We even noted on one occasion in a N. clavipes web, at the margins of a perforation where the spider appeared to have cut out a moth, traces of crimson scales that we thought could only have come from a Cosmosoma.
Most remarkable about the behavior of Cosmosoma is the flocculent delivery system of the male. That the flocculent is laden with alkaloid and the female so richly embellished with its filaments indicated strongly that the flocculent might serve for defense of the female, which proved to be the case. The alkaloid that the female receives systemically by seminal infusion from the male might, of course, also contribute to the female's defense (just as the alkaloid sequestered from plants and stored systemically by the male protects the male), but the flocculent is evidently protective in its own right: virgin females, devoid of systemic alkaloid and endowed experimentally with no more than the flocculent of an alkaloid-laden male, derived substantial protection from the endowment.
The flocculent has the potential also of being of immediate benefit to the female. One could imagine the female being protected by her topical coating from the very outset to the end of copulation, a period of hours during which she is quiescent and at risk from any number of predators, including ants and lycosid spiders. Whether by the time the female uncouples from the male she is already doubly protected by the alkaloid coating and her internal alkaloid load depends on how quickly the systemic alkaloid is distributed through her body. There is evidence from another arctiid (U. ornatrix) that alkaloid received seminally from the male is deployed defensively with such speed that by the end of mating the female is already fully protected by the gift and rejected on the basis of mere contact by spiders (11).
We do not know how long the topical filamentous shroud persists on an embellished female Cosmosoma, nor how often the investiture might be renewed through remating. We suspect that the bulk of the filaments sticking to a female might be lost in relatively quick order after the female takes flight, but that a small remnant might suffice to impart a significant level of distastefulness. Spiders appear to be highly sensitive to pyrrolizidine alkaloids (3, 12) and, given how potently the individual filaments are laced with alkaloid in Cosmosoma, it could be that even a sparse topical sprinkling of filaments suffices to deter spiders.
We are also ignorant of how often male Cosmosoma mate in nature and of whether they ever exhaust their flocculent in the process of courting. For that matter, we are ignorant of whether males visit alkaloid-containing plants as a matter of routine to replenish the systemic alkaloid they give up at mating, or whether they procure alkaloid only once at the beginning of adult existence. The finding that the male Comosoma spotted at our plant baits included individuals of worn appearance suggests that multiple visitation might be the norm. The female, in turn, if she mates multiply, could reap benefits quite aside from the periodic restoration of her defensive cloak. Multiple spermatophores would provide her with a diversity of sperm, with multiple loads of nutrient, and with the capacity to restore at intervals the quantity of systemic alkaloid she relinquishes to the eggs.
A question that remains intriguing is whether the flocculent, or more specifically its contained alkaloid, has signal value in courtship. The alkaloid could, for instance, provide a measure of the male's “worth,” in the sense that it could be an indicator of the male's alkaloid content and therefore indirectly of the amount of alkaloid the male holds in store for seminal transmission to the female. On the basis of such advertisement, the female could exercise mate choice. That in our wind tunnel experiments the females exercised preference for males that expelled flocculent might be taken to be suggestive in this regard. However, additional data we have from such experiments (13) indicate females do not discriminate between flocculent-expelling males on the basis of whether their flocculent was alkaloid laden. Moreover, this lack of discrimination holds even when the alkaloid fed to the males is the intermedine/lycopsamine mixture (rather than monocrotaline, as in our present wind-tunnel tests) (W.E.C., unpublished results). It appears therefore that the females either ignore differences in flocculent alkaloid content or are unable to perceive the difference because of the extremely low volatility of the alkaloid. Interestingly, in two other lepidopterans, U. ornatrix and Danaus gillippus, in which alkaloid content of the male is assessed by the female in courtship, the assessment is based not on the alkaloid itself but on volatile pheromonal derivatives (hydroxydanaidal, danaidone) of the alkaloid produced by the male in proportion to its alkaloid content (3, 14, 15).
One could, of course, envision an alternative communicative strategy for Cosmosoma in which the female accepts most males, as long as they are able to expel flocculent but subsequently uses sperm only from males rich in alkaloid. By such strategy, the female could be exercising sperm selection rather than mate choice and could benefit both phenotypically (she could garner at least some resource with each mating, in the form of alkaloid, nutrient, or both) and genetically (she could be selecting for males adept at alkaloid sequestration, a trait that could be heritable). In U. ornatrix there is suggestive evidence that the female exercises such postcopulatory sperm selection (16).
An intriguing possibility is that the male, by festooning its mating partner, might prevent that partner from remating. It would certainly be advantageous for males to guard against sperm competition by labeling their mates in this fashion, just as it would be advantageous for males to discriminate against labeled females.
Cosmosoma is not alone among Arctiidae in possessing male abdominal pouches laden with flocculent. Such structures have been noted in a number of species of Cosmosoma, all members of the tribe Euchromiini, in which the pouches may even have evolved more than once (17). No function has been demonstrated for the pouches, although an interesting observation suggests they may at times be used for defense. Even slight manual disturbance is said to result in forceful expulsion of flocculent in some species (18). We never noted our Cosmosoma to respond in such fashion.
Nor is Cosmosoma alone among arctiids in being attracted to pyrrolizidine alkaloids or pyrrolizidine alkaloid-containing plants (19, 20). Alkaloid sequestration, for protective purposes as in Cosmosoma, is therefore likely to occur in other species as well.
One open question is whether predators other than spiders, especially vertebrates, are also deterred by the acquired alkaloids of Cosmosoma. The relationship of Cosmosoma to vertebrates might be particularly interesting, given that the moth is so distinctly aposematic. One wonders whether vertebrates might be intrinsically predisposed to avoid Cosmosoma, or whether they might discriminate against the moth only as a consequence of experience with alkaloid-laden individuals. It would be interesting to know, in this context, what fraction of the Cosmosoma population is alkaloid laden at any one time. Cosmosoma could be illustrative of a classical case of automimicry (21), where the alkaloid-laden individuals are the “models” and their alkaloid-free counterparts the “mimics.” One would predict that the fraction of alkaloid-laden individuals in nature should be high. Males, one would expect, for both defensive and reproductive reasons, should give high priority to alkaloid sequestration, whereas females, for the same reasons, should give high priority to mating.
Some uncertainty remains regarding the actual plants visited by Cosmosoma in nature. E. capillifolium, almost certainly, is one of them, as is perhaps one of its congeners, E. mohrii. But there could be other species as well. In the Florida region where we studied Cosmosoma, the moth is usually not abundant, and the chance of spotting males during actual alkaloid imbibition is slight.
Acknowledgments
We thank Scott Smedley (Cornell) and Jeff Slotten (Gainesville, FL) for providing live Cosmosoma, Athula Attygalle (Cornell) for help with MS, Rob Davidson (Wake Forest) and Maria Eisner (Cornell) for taking the electronmicrographs, Melinda McElveen (Wake Forest) and Jess Hines (Wake Forest) for carrying out some of the baiting experiments, Nickolay Hristov (Wake Forest) for taking the photo in Fig. 1B, the personnel of the Archbold Biological Station for much help and hospitality, and Kern Reeve (Cornell) and Carmen Rossini (Universidad de la República, Montevideo) for comments on the manuscript. The work was supported in part by National Institutes of Health Grants AI02908 and GM53830.
Footnotes
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.260503797.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.260503797
References
- 1.LeConte J L. The Complete Writings of Thomas Say on the Entomology of North America. Vol. 1. New York: Baillier; 1859. p. 42. , Plate 19. [Google Scholar]
- 2.Roitman J N. Australian J Chem. 1983;36:769–778. [Google Scholar]
- 3.Eisner T, Meinwald J. In: Pheromone Biochemistry. Prestwich G D, Blomquist G J, editors. Orlando: Academic; 1987. pp. 251–269. [Google Scholar]
- 4.Liu K, Roder E, Chen H L, Xiu X J. Phytochemistry. 1992;31:2573–2574. [Google Scholar]
- 5.Pederson E. Phytochemistry. 1975;14:2086–2087. [Google Scholar]
- 6.Ehmke A, Witte L, Biller A, Hartmann T. Z Naturforsch. 1990;45c:1185–1192. [Google Scholar]
- 7.Cardé R T. In: Chemical Ecology of Insects. Bell W J, Cardé R T, editors. London: Chapman & Hall; 1984. pp. 111–124. [Google Scholar]
- 8.Dussourd D E, Ubik K, Harvis C, Resch J, Meinwald J, Eisner T. Proc Natl Acad Sci USA. 1988;85:5992–5996. doi: 10.1073/pnas.85.16.5992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hare J F, Eisner T. Oecologia. 1993;96:9–18. doi: 10.1007/BF00318024. [DOI] [PubMed] [Google Scholar]
- 10.Eisner T, Eisner M, Rossini C, Iyengar V K, Roach B L, Benedikt E, Meinwald J. Proc Natl Acad Sci USA. 2000;97:1634–1639. doi: 10.1073/pnas.030532797. . (First Published February 4, 2000; 10.1073/pnas.030532797) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.González A, Rossini C, Eisner M, Eisner T. Proc Natl Acad Sci USA. 1999;96:5570–5574. doi: 10.1073/pnas.96.10.5570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Eisner T, Eisner M. Psyche. 1991;98:111–118. [Google Scholar]
- 13.Boada R. M.S. thesis. Wake Forest University, Winston-Salem, NC: Department of Biology; 1997. [Google Scholar]
- 14.Eisner T, Meinwald J. Proc Natl Acad Sci USA. 1995;92:50–55. doi: 10.1073/pnas.92.1.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Conner W E, Eisner T, Vander Meer R K, Guerrero A, Meinwald J. Behav Ecol Sociobiol. 1981;9:227–235. [Google Scholar]
- 16.LaMunyon C W, Eisner T. Proc Natl Acad Sci USA. 1994;91:7081–7084. doi: 10.1073/pnas.91.15.7081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Weller S J, Simmons R B, Boada R, Conner W E. Ann Ent Soc Am. 2000;93:921–928. [Google Scholar]
- 18.Blest A D. Zoologica. 1964;49:161–181. [Google Scholar]
- 19.Pliske T. Environ Entomol. 1975;4:455–473. [Google Scholar]
- 20.Boppré M. J Chem Ecol. 1990;16:165–185. doi: 10.1007/BF01021277. [DOI] [PubMed] [Google Scholar]
- 21.Wickler W. Mimicry in Plants and Animals. London: World University Library; 1968. [Google Scholar]