

Abstract
Plasma levels of nitrite ions have been used as an index of nitric oxide synthase (NOS) activity in vivo. Recent data suggest that nitrite is a potential intravascular repository for nitric oxide (NO), bioactivated by a nitrite reductase activity of deoxyhemoglobin. The precise levels and compartmentalization of nitrite within blood and erythrocytes have not been determined. Nitrite levels in whole blood and erythrocytes were determined using reductive chemiluminescence in conjunction with a ferricyanide-based hemoglobin oxidation assay to prevent nitrite destruction. This method yields sensitive and linear measurements of whole blood nitrite over 24 hours at room temperature. Nitrite levels measured in plasma, erythrocytes, and whole blood from 15 healthy volunteers were 121 plus or minus 9, 288 plus or minus 47, and 176 plus or minus 17 nM, indicating a surprisingly high concentration of nitrite within erythrocytes. The majority of nitrite in erythrocytes is located in the cytosol unbound to proteins. In humans, we found a significant artery-to-vein gradient of nitrite in whole blood and erythrocytes. Shear stress and acetylcholine-mediated stimulation of endothelial NOS significantly increased venous nitrite levels. These studies suggest a dynamic intravascular NO metabolism in which endothelial NOS-derived NO is stabilized as nitrite, transported by erythrocytes, and consumed during arterial-to-venous transit. (Blood. 2005;106:734-739)
Introduction
Nitric oxide (NO) is a gas that is continuously synthesized in endothelial cells and executes multiple functions that maintain vascular homeostasis. In the vascular system NO is synthesized by the type III isoform of NO synthase (endothelial NOS [eNOS]).1,2 When NO is released from the endothelium, it may diffuse abluminally into smooth muscle cells causing vasodilation; when released luminally into the bloodstream NO reacts with intraerythrocytic oxyhemoglobin to form nitrate, and a portion of the remaining NO is oxidized to nitrite.3,4 We have recently shown that nitrite has the potential to be a major intravascular NO storage molecule in humans that is capable of transducing NO bioactivity distal to its site of formation.5
Plasma nitrite has been described as an index of eNOS activity in the regional6 and systemic circulation in humans and various mammals.7 Despite the growing appreciation of an important potential role for nitrite in physiology and as a disease marker, the actual circulating levels of nitrite in humans have been difficult to measure, owing to the relative instability of nitrite in blood, as well as contaminating nitrite in clinical blood collection tubes and laboratory ware. This has resulted in reported levels ranging from undetectable8 to 20 μM.9 A recent report identified some of the analytical problems of measuring nitrite in plasma, potentially explaining the wide range of reported levels.7 In that study plasma nitrite was determined with 3 independent analytical methods and rapid sample preparations in 7 mammalian species, strongly suggesting that in vivo plasma nitrite concentrations are below 500 nM in mammals. The short half-life of nitrite in whole blood, demanding rapid separation of plasma from erythrocytes, is a considerable obstacle for plasma nitrite determination in clinical practice, and perhaps more importantly, the measure of plasma nitrite ignores the potential importance of erythrocytes in nitrite/NO metabolism.
Nitrite, like NO, reacts with erythrocytic oxyhemoglobin to form nitrate and methemoglobin.10 However, nitrite will react with deoxyhemoglobin to form NO and methemoglobin,11 a reaction that we have proposed contributes to erythrocyte-dependent hypoxic vasodilation.5,12 Such a model of dynamic intravascular blood flow regulation suggests a rapid reaction of nitrite with intraerythrocytic hemoglobin.5,13 We, therefore, hypothesized that the erythrocyte might be an additional reservoir for nitrite.
To determine whole blood and erythrocytic nitrite concentration, we developed a ferricyanide-based hemoglobin oxidation “stabilization-solution,” designed to limit ex vivo nitrite oxidation by oxyhemoglobin and reduction by deoxyhemoglobin, that otherwise occurs after blood collection. Using this methodology, we studied the compartmentalization of nitrite in human plasma and erythrocytes and characterized mechanisms governing nitrite formation and consumption in the human circulation.
Materials and methods
Human volunteers
Blood for human in vitro studies was drawn from healthy volunteers enrolled in an institutional review board (IRB)-approved human subjects blood collection protocol. To evaluate whole blood nitrite as an index of regional NOS activity, 8 healthy volunteers (4 women and 4 men; age range, 22-31 years) were enrolled in an IRB-approved forearm blood flow study. Exclusion criteria were the presence of cardiovascular risk factors, liver disease, renal failure, or acute and chronic infections (including HIV). Volunteers fasted overnight before studies began. All participants signed informed written consent, per the Declaration of Helsinki. Human protocols were approved by the Scientific and Institutional Review Boards of the Intramural Division of the National Heart Lung and Blood Institute, NIH. Animal experiments have been approved by the Animal Care and Use Committee of the NIDDK.
Ferricyanide-based hemoglobin oxidation assay
To measure nitrite in whole blood, we mixed whole blood with a nitrite preservation solution containing 0.8 M ferricyanide (an oxidizing agent), 10 mM N-ethylmaleimide (NEM; a thiol alkylating agent), and 1% NP-40 (a cytolytic agent to provide access of the ferricyanide and NEM to the red cell contents) in a 5:1 dilution (vol:vol; whole blood-stop solution) immediately on blood collection. Preliminary studies showed this ratio to be optimal for consistent results (data not shown). Following blood addition to the preservation solution, the sample was deproteinated with methanol (vol: vol; 1:1) and spun down at 15 000g for 2 minutes. Then, 200 μL supernatant was injected into a solution of acidified tri-iodide, purging with helium in-line with a gas-phase chemiluminescence NO analyzer (Sievers, Boulder, CO). The tri-iodide solution was prepared by mixing 2 g potassium iodide, 1.3 g iodine dissolved in 40 mL water with 140 mL acetic acid. Then, 6 to 8 mL of this solution was placed in the purge vessel into which samples were injected. This solution will reduce nitrite, iron-nitrosylheme and S-nitrosothiols to NO gas, which is detected by the NO analyzer.14-16 In additional experiments, plasma and erythrocytes were immediately separated by centrifugation at 2000g for 1 minute. Erythrocytes were treated with the ferricyanide-based preservation solution as described for whole blood. The nitrite levels in plasma, red cells, and whole blood in 15 healthy human volunteers were then determined using the reductive tri-iodide-based chemiluminescence assay with and without a 2-minute pretreatment with a specific scavenger of nitrite (10% acid sulfanilamide).17 Sensitivity, linearity, recovery, and reproducibility were determined by first adding known amounts of sodium nitrite into whole blood, then transferring this specimen into nitrite preservation solution. Stability of nitrite in the stabilization solution at different concentrations and temperatures and over time was evaluated. Cytosol and membrane fractions have been isolated from erythrocytes using modified standard protocols.18 Whole blood was collected into ice cold phosphate-buffered saline (PBS; 1:5 dilution) and then exposed to 100% carbon monoxide for 5 minutes. Then erythrocytes were pelleted and resuspended to hematocrit of 50% in PBS. To 4 mL of the erythrocyte solution 32 mL of 5 mM of dibasic sodium phosphate at pH 8 was added. Lysed erythrocytes were spun at 22 000g for 40 minutes, the supernatant discarded, then resuspended with 12 mL sodium phosphate solution, and then spun at 22 000g for 20 minutes. Then the erythrocyte pellet was resuspended in PBS to a final volume of 2.5 mL and injected into tri-iodide. In control experiments erythrocytic nitrite remained stable over a period of 2 hours when equilibrated with carbon monoxide. The cytosol of erythrocytes was obtained by lysing erythrocytes in 10 mM ferricyanide in MilliPore (CQ-Gard, Bedford, MA) water and subsequent spinning of the lysis solution for 20 minutes at 22 000g. The supernatant was injected directly and after passing it through a Sephadex G25 sizing column. The eluate was injected into tri-iodide to determine the portion of nitrite bound on anionic binding sites of hemoglobin.
To compare whole blood nitrite levels from humans to often used laboratory animals, we studied the whole blood nitrite levels of 25 C57BL/6 mice under basal conditions and 20 mice treated with L-NG-nitro-L-arginine methyl ester (L-NAME; 100 mg/kg) intraperitoneally. Whole blood was drawn out of the retro-orbital vein from these mice. We measured whole blood nitrite 2, 4, 6, and 24 hours after intraperitoneal injection of L-NAME in 5 mice at each time point.
Whole blood nitrite as an index of acute changes of NOS activity in humans
Arterial and venous angio catheters were placed in the brachial arteries and antecubital veins of 8 healthy volunteers. Basal forearm blood flow was measured by venous occlusion strain-gauge plethysmography, as previously described.19 Arterial and venous blood was drawn for the measurement of basal whole blood and erythrocyte nitrite levels. Blood for whole blood nitrite determinations was immediately treated at the bedside with nitrite preservation solution. Erythrocytes were separated after venisection by spinning whole blood at 2000g for 1 minute and were then frozen in dry ice. After basal measurements, acetylcholine (ACh) was infused at 7.5 μg/min for 3 minutes and every 30 seconds during ACh infusion venous blood was drawn and immediately mixed with nitrite preservation solution for whole blood nitrite measurements. Whole blood samples were kept on ice until measured by tri-iodide chemiluminescence on the same day. In additional experiments, a blood pressure cuff was placed around the arm and inflated to 250 mm Hg, circulatory stasis was maintained for 5 minutes, and then the blood pressure cuff was released. The release of the blood pressure cuff triggers a flow-mediated vasodilation of the brachial artery, which is nearly exclusively dependent on endothelial NOS.20 Before and 1 minute after ischemia forearm blood flow was measured via venous occlusion plethysmography and venous blood was drawn for nitrite measurements.
Statistical analysis
Data processing was performed with the different modules of the software package SPSS (Release 11.0, SPSS, Chicago, IL) and Prism (Graph Pad Version 4). Data are given as the means plus or minus SEM and significance accepted with P less than .05.
Results
Ferricyanide-based hemoglobin oxidation assay preserves nitrite in blood
Erythrocytes and whole blood were treated in a 5:1 ratio with nitrite preservation solution as described in “Materials and methods.” Injection of 200 μL plasma, erythrocytes, and whole blood solutions into tri-iodide produced NO signals from 5 to 20 mV (Figure 1A). When samples were pretreated with acid sulfanilamide, no chemiluminescence signal was detectable, indicating specificity of the assay for nitrite. During sample processing of erythrocytes and whole blood, samples were diluted by 2.5 with nitrite preservation solution and methanol; for Figure 1A plasma was diluted with PBS by 2.5 for visual comparison of NO peak height from erythrocytes and whole blood. The assay was sensitive to 1 pmol nitrite added to whole blood with 200 μL sample injections (signal-to-noise ratio, 3:1). The assay was linear over the desired concentration range of 0 to 100 μM (Figure 1B; r2 = 0.998, P < .001) and the recovery was 97% plus or minus 5% (n = 20). The measured coefficient of variation was 5%. There was no detectable difference between samples that were measured directly and those subjected to freeze/thaw and deproteinated with methanol (98% ± 6%, n = 20; Figure 1C). Nitrite additions to a final concentration of 1 μM in whole blood revealed nitrite stability over 24 hours at room temperature after addition of the nitrite preservation solution (Figure 1D). The half-life of nitrite in whole blood without nitrite preservation solution was 10.2 minutes at room temperature (Figure 1D insert), a result consistent with the previously published half-life.21
Figure 1.

Validation of assay based on ferricyanide to stabilize nitrite in whole blood. (A) Original registration of reductive tri-iodide-based chemiluminescence after injection of 200 μL plasma, treated erythrocytes, and whole blood with or without addition of acid sulfanilamide. Loss of signal after sulfanilamide indicates specificity of the assay for nitrite. (B) Recovery and linearity of nitrite spiked in whole blood to which nitrite preservation solution was added. (C) Freeze-thaw and sample processing does not influence nitrite concentration in whole blood. Open circles represent measurements after freezing, storage at -80°C for 4 days, thawing, and deproteination. With an injection volume of 200 μL, 1 pmol nitrite could be recovered at a signal-to-noise ratio of 3:1. (D) Stability of nitrite in whole blood. Spiking of nitrite to a final concentration of 1 μM and measurement of nitrite concentration of whole blood up to 24 hours after nitrite addition. Samples were either treated with nitrite preservation solution (○) or saline (□) after addition of nitrite. The inset illustrates the disappearance of nitrite in whole blood with or without nitrite preservation solution. Samples were stored at room temperature during incubation.
Levels of nitrite in whole blood and in erythrocytes
Using this validated assay, we found that the highest concentration of nitrite in blood could be recovered in the erythrocyte fraction (n = 15; healthy human volunteers; Figures 1A and 2A). The nitrite levels measured in plasma, directly measured in red blood cells (RBCs) and whole blood, and calculated for whole blood (based on measurements in plasma and RBC fractions and corrected for hematocrit) of 15 healthy volunteers were 121 plus or minus 9, 288 plus or minus 47, 176 plus or minus 17, and 184 plus or minus 22 nM, respectively. Whole blood nitrite levels were similar at 8 am and 12 am in studied individuals (8 am, 181 ± 29 nM; 12 am, 176 ± 17 nM; n = 8). Whole blood nitrite levels were similar when drawn on 3 consecutive days with a coefficient of variation of less than 5% between measurements (n = 3). The agreement between measured and calculated whole blood nitrite levels indicates the internal consistency between these methods, an important prerequisite in developing a clinically useful assay. The half-life of nitrite at room temperature in the erythrocyte pellet was similar to that in whole blood, whereas nitrite levels in plasma and water did not change measurably over 30 minutes (data not shown).
Figure 2.

Nitrite concentration in whole human blood and its components. (A) Distribution of nitrite in the various compartments of whole blood (WB). Measured whole blood concentration treated with nitrite preservation solution equals the calculated whole blood nitrite concentration from plasma and RBC nitrite after the allowance for hematocrit. (B) Location of nitrite within the erythrocyte. Error bars show standard error of the mean.
To investigate where nitrite was located in the erythrocyte, the cytosol was separated from the erythrocyte membrane. Virtually all nitrite was located in the cytosol, 94% plus or minus 4% of the entire erythrocyte fraction. To test whether nitrite is bound to proteins in the cytosol of erythrocytes, we passed the cytosol through a G25 column. This treatment of the sample eliminated the nitrite signal almost completely (Figure 2B).
As a comparison to humans we measured the basal and NOS-inhibited whole blood nitrite levels in mice. The whole blood nitrite levels of C57BL/6 mice was 650 plus or minus 29 nM and decreased to 556 plus or minus 26, 455 plus or minus 11, 421 plus or minus 18, and 551 plus or minus 30 nM at 2, 4, 6, and 24 hours after intraperitoneal L-NAME injection (P = .002).
Changes of whole blood levels in the human forearm with modulation of NOS activity
To test the hypothesis that stimulation of endogenous eNOS increases whole blood nitrite concentration, we studied human forearm blood flow and accompanying biochemical changes during the infusion of the endothelium-dependent vasodilator, ACh, and following shear stress-mediated NOS activation induced by reactive hyperemia. Infusion of ACh into the brachial artery and ischemia-induced flow-mediated dilation increased forearm blood flow from 3.6 plus or minus 0.5 to 18.2 plus or minus 3.6 mL/min/100 g tissue for ACh (n = 8; P < .001) and from 3.2 plus or minus 0.3 to 39.5 plus or minus 1.6 mL/min/100 g tissue 1 minute after induced shear stress (Figure 3A). NOS stimulation by ACh or shear stress increased venous whole blood nitrite concentration (ACh, from 143 ± 9 to 246 ± 22 nM; shear, from 143 ± 7 to 218 ± 18 nM 1 minute after ischemia; n = 4).
Figure 3.

Changes in nitrite concentration during blood flow through the human forearm circulation. (A) Infusion of acetylcholine (ACh) (7.5 μg/min) intra-arterially in the human forearm and ischemia-induced flow-mediated dilation causes a 4- to 10-fold increase in forearm blood flow. (B) The stimulation reverses the normal small uptake of nitrite to instead cause a release of nitrite into the circulation, a result consistent with NO-to-nitrite conversion during forearm circulatory passage. Horizontal bars represent mean values. (C) Comparison of nitrite formation under basal conditions (saline infusion) and NOS stimulation via ACh or shear stress. (D) Time profile of nitrite responses in whole venous blood during continuing ACh infusion (representative results). Error bars show standard error of the mean.
Consistent with artery-to-vein gradient in plasma,14 we found an artery-to-vein gradient of nitrite in whole blood and erythrocytes across the human forearm circulation (whole blood: artery, 176 ± 10 to vein, 143 ± 7 nM; P = .003; Figure 3B). A similar artery-to-vein gradient was observed in the erythrocyte fraction (23 ± 8 nM; P = .04; n = 6). When nitrite extraction was calculated using the Fick principle, we observed a net extraction of nitrite from the circulation of 135 plus or minus 40 pmol/min/100 g tissue. Infusion of ACh or shear stress resulted in a net nitrite formation across the forearm circulation (from basal: -135 ± 40 to 1690 ± 330 with ACh stimulation or 2761 pmol/min/100 mL tissue with shear stress; P = .001; Figure 3C). The increase in nitrite levels in whole blood was transient during ACh stimulation of endogenous NO synthesis. When blood was drawn from the effluent vein under ACh infusion, whole blood nitrite concentration increased to a maximum 90 seconds after the infusion began and then decreased to near basal levels as the infusion continued (Figure 3D). Lauer et al6 have described a similar phenomenon for plasma nitrite levels.
Discussion
We have established an assay for whole blood nitrite based on the use of ferricyanide to oxidize erythrocytic hemoglobin to methemoglobin and thus limit hemoglobin-dependent redox reactions with nitrite. We demonstrate that this reaction terminates hemoglobin-nitrite reactions, which are responsible for the relatively rapid consumption of nitrite after blood is drawn from the body. This assay allows for room temperature collection of blood at the bedside for the determination of actual in vivo nitrite levels in erythrocytes and whole blood. Using this assay, we found that (1) the basal whole blood nitrite levels in healthy resting humans are about 200 nM, (2) nitrite in erythrocytes constitutes about two thirds of intravascular nitrite at concentrations approaching 300 nM, and (3) there is a whole blood and erythrocyte nitrite artery-to-vein gradient across the human forearm circulation in healthy human volunteers, consistent with a dynamic oxygen-dependent nitrite metabolism.
Whole blood nitrite increases by various stimulators of NOS activity
NO reacts with hemoglobin in a nearly diffusion-limited reaction to form nitrate and methemoglobin and with oxygen to form various oxidation products.22 This results in an exceptionally short half-life of NO in blood and creates considerable difficulties in the direct measurement of the blood levels of the NO radical.23 However, compelling evidence indicates that one of the oxidative products of NO in blood, nitrite, reflects a constant proportion of the total endothelial NO synthesis. This thesis is principally supported by the observation that 90% of circulating plasma nitrite is derived from NOS activity in humans and other mammals.24 In addition, plasma nitrite levels are reduced up to 70% in eNOS knock-out mice as compared to control mice7,25 and Lauer et al6 demonstrated that plasma nitrite levels reflect acute changes in regional NOS activity. A valid index of NOS activity would yield insights into NO metabolism both in vitro and in vivo and may also serve as a surrogate of endothelial dysfunction in patients with atherosclerosis and other diseases exhibiting a change in NO bioavailability.26,27
Plasma nitrite has been used as a surrogate for intravascular nitrite concentration for several reasons. First, nitrite is taken up quickly into erythrocytes from plasma and reacts with hemoglobin to form nitrate. Only a portion of intravascular nitrate is derived from NOS-dependent pathways; thus, once nitrite is converted to nitrate it is not an index of acute changes in NOS activity using currently available analytical methodologies. Second, proteins interfere with most nitrite assays7 and by separating erythrocytes from plasma many proteins are eliminated from the sample. Plasma nitrite is widely accepted as an index of NOS activity in vivo; however, the need for rapid blood processing after venisection creates a considerable obstacle for plasma nitrite determination in clinical practice. Moreover, changes in intraerythrocytic nitrite cannot be evaluated.
To overcome these analytical problems, we developed a ferricyanide-based hemoglobin oxidation assay that stabilizes nitrite in lysed whole blood and the erythrocyte. The principle of the method is that ferricyanide reacts with hemoglobin to form methemoglobin (Fe-II-hemoglobin + ferricyanide → Fe-III-hemoglobin + ferrocyanide).28 Methemoglobin, unlike hemoglobin, does not react with nitrite to form nitrate nor does it reduce nitrite to NO.29 Because the reaction of hemoglobin with nitrite represents the major mechanism for nitrite degradation, the addition of ferricyanide stabilizes nitrite in whole blood. We find that measurement of nitrite in whole blood using this method is sensitive, linear, and reproducible. We recently validated the reductive chemiluminescence against 2 other independent (flow injection analysis combined with the Griess reagent and high-performance liquid chromatography [HPLC]) assays for nitrite (r2 = 0.9).7 Reaction of whole blood with preservation solution stabilizes nitrite for 24 hours at room temperature. Moreover, freezing at -80°C, storage over 4 days at -80°C, and freeze-thawing samples does not alter nitrite concentration measurably in the presence of ferricyanide. Using this methodology, we have demonstrated that whole blood nitrite reflects increases of NOS activity stimulated by ACh infusion and shear stress in vivo. Whole blood nitrite measurements complement the analysis of plasma nitrite as an index of NOS activity. It facilitates the analysis of intravascular nitrite concentrations so that it can be readily used in clinical practice and for the first time integrates the measurement of nitrite in erythrocytes, a compartment that has been neglected so far in the assessment of intravascular NO/nitrite fluxes.
Erythrocytes contain the majority of intravascular nitrite in whole blood
We have recently shown that nitrite has the potential to be a major intravascular NO storage molecule and that hemoglobin acts as a nitrite reductase contributing to RBC-dependent hypoxic vasodilation.5,12 Such a model requires a reaction of nitrite with intraerythrocytic hemoglobin.5,13 We, thus, hypothesized that erythrocytes would serve as a reservoir for intravascular nitrite. In 15 healthy individuals we could show that two thirds of intravascular nitrite is located in erythrocytes and the majority of nitrite in erythrocytes is located in the cytosol. In these individuals we found that the levels of nitrite in the erythrocyte approached 300 nM, confirming a recent report by Bryan et al.30 We found an artery-to-vein gradient for whole blood and erythrocyte nitrite in humans consistent with nitrite use as a source of bioactive NO in the microcirculation.14 On deoxygenation of erythrocytes resident nitrite may be catalytically reduced to produce NO, putatively providing a mechanism for matching local blood flow to oxygen demand.5
Higher concentrations of nitrite in the erythrocyte than in plasma are surprising in the presence of 20 mM heme. At first sight it may be expected that nitrite enters erythrocytes via the anion exchanger (AE-1). However, incubation of erythrocytes with 4,4′di-isothiocyanatostilbene-2,2′-disulfonic acid (DIDS), an inhibitor of AE-1, does not change the uptake rate of nitrite from plasma (data not shown), corroborating previous reports in carp, pigs, and humans.21,31 If nitrite would follow the Gibbs-Donnan equilibrium like bicarbonate or chloride, the ratio of erythrocyte nitrite concentration to plasma nitrite concentration would be expected to be 0.7. The apparent departure from this points either toward a more complicated mechanism of nitrite uptake from plasma31 or a constant formation of nitrite within the erythrocyte. These results suggest a dynamic control of nitrite decay/nitrite reduction to NO and nitrite uptake/formation within the erythrocyte.
There are several eNOS-independent influences on blood nitrite such as the amount of nitrate in diet32 and systemic inflammatory processes.26 Dietary nitrate is converted to nitrite in the oral cavity by commensal bacteria containing nitrate reductase. Nitrite can then be absorbed via the gastrointestinal tract and increases time dependently plasma nitrite concentration after a nitrate challenge. To design clinical studies measuring plasma or erythrocyte nitrite, one has to bear in mind the possible influence factors on intravascular nitrite concentrations.
In Figure 4 we present a summary of current knowledge about biochemical pathways of intravascular NO metabolism. As noted, NO produced in endothelial cells by the action of eNOS, which diffuses luminally, undergoes diverse oxidative/reductive reactions to produce the multiple valence states of NO, symbolized as NxOy.22 These states react to a small, but potentially biologically important extent, to produce species such as RXNO,33 RSNO,34 and nitrated lipids,35 which have been shown to have biologic signaling activity.
Figure 4.
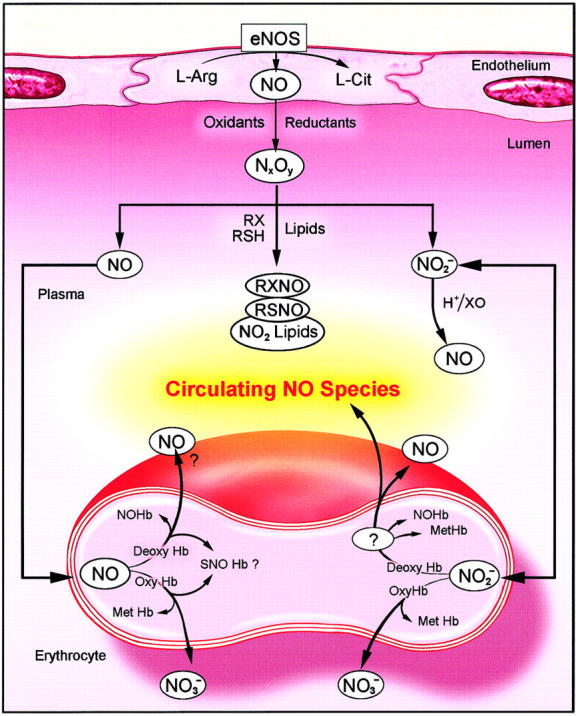
A model of intravascular metabolism of NO. NO produced by eNOS may diffuse into the vascular lumen as well as the underlying smooth muscle. The majority of this NO enters the erythrocyte and reacts with oxyhemoglobin (Oxy Hb) to form nitrate (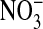 ); a minor portion may escape the hemoglobin scavenger and react with plasma constituents to form nitros(yl)ated species (RXNO, nitrosothiols: RSNO), nitrated lipids (NO2 lipids), and nitrite (
); a minor portion may escape the hemoglobin scavenger and react with plasma constituents to form nitros(yl)ated species (RXNO, nitrosothiols: RSNO), nitrated lipids (NO2 lipids), and nitrite (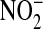 ). Each of these species is capable of transducing NO bioactivity far from its location of formation. Nitrite may diffuse into the erythrocytes where it appears in a higher concentration than in plasma. In the erythrocyte nitrite reacts with deoxyhemoglobin (Deoxy Hb) to form nitric oxide and methemoglobin (Met Hb) and other NO adducts. NO can then either diffuse out of the erythrocyte directly or via an intermediate NO metabolite. The question mark circled in white refers to the possibility of an intermediate during nitrite bioactivation. NO Hb indicates iron nitrosylhemoglobin; SNO Hb, nitrosohemoglobin; L-Arg, L-arginine; L-Cit, L-citrulline; NxOy: higher N oxides.
). Each of these species is capable of transducing NO bioactivity far from its location of formation. Nitrite may diffuse into the erythrocytes where it appears in a higher concentration than in plasma. In the erythrocyte nitrite reacts with deoxyhemoglobin (Deoxy Hb) to form nitric oxide and methemoglobin (Met Hb) and other NO adducts. NO can then either diffuse out of the erythrocyte directly or via an intermediate NO metabolite. The question mark circled in white refers to the possibility of an intermediate during nitrite bioactivation. NO Hb indicates iron nitrosylhemoglobin; SNO Hb, nitrosohemoglobin; L-Arg, L-arginine; L-Cit, L-citrulline; NxOy: higher N oxides.
NO, which has not yet undergone these reactions, is believed to be largely destroyed by reaction with trace amounts of plasma hemoglobin36 or to diffuse into erythrocytes to react with oxyhemoglobin to produce nitrate and methemoglobin. NO, which reacts with deoxyhemoglobin, produces primarily iron-nitrosyl-hemoglobin (NO-Hb). Intraerythrocytic NO-Hb may decay under certain conditions to release NO from the erythrocyte.37 An additional species, S-nitroso-hemoglobin (SNO-Hb) has also been postulated to be a biologic source of NO,38 but its quantities, stability, and biologic role remain uncertain.39-41
The present study highlights the nitrite/erythrocyte pathways that are shown on the right of Figure 4. Nitrite ions produced in plasma from oxidation of intravascular NO can diffuse reversibly into erythrocytes and react with hemoglobin and undergo oxidative and reductive reactions. Similar to NO, intraerythrocytic nitrite, which constitutes the bulk of intravascular nitrite, reacts with oxyhemoglobin to produce methemoglobin and nitrate.10 The reaction of nitrite with deoxyhemoglobin produces either NO or other NO intermediates that are bioactive.5,12,13 (In addition it is to be noted that intravascular reduction of nitrite may occur at low pH by disproportionation and may also be catalyzed by the enzyme xanthine oxido-reductase.42) These erythrocytic reactions, based on erythrocytic nitrite fluxes and metabolism, are emerging as important modulators of intravascular NO bioactivity, especially under physiologic hypoxia.
Besides nitrite, erythrocytes contain several vasodilators among which are nitros(yl)ated proteins,38 nitrated lipids,43 and adenosine triphosphate (ATP).44 Erythrocytes modulate the concentration of endothelin-145 and angiotensin II.46 The long-perceived dogma that erythrocytes merely terminate NO bioactivity, limiting its action to the diffusion radius, should be modified to suggest that erythrocytes participate in the regulation of vascular homeostasis, which includes control of leukocyte adhesion and platelet aggregation as well as vascular tone.
The development of a means to measure nitrite in whole blood and erythrocytes, together with the characterization of erythrocytic nitrite levels in humans, has important implications for the study of intravascular NO species. First of all, it lays the basis for the establishment of new markers of endothelial dysfunction and, secondly, it makes the study of intravascular nitrite fluxes across the red blood cell membrane and the kinetics of intraerythrocytic chemical reactions more feasible. The latter will help to clarify the role of NO, nitrite, and other related species in physiologic erythrocyte-dependent hypoxic vasodilation.
Note added in proof. A recent methodological paper by Rogers et al47 reports total human red cell NO species of about 550 nM, consistent with our determination of about 160 nM iron-nitrosylhemoglobin and 55 nM S-nitroso-hemoglobin,14 and, in this report, 290 nM nitrite ions. The 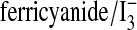 assay used in this and previous publications5,12,14,15,19,36 prevents NO rebinding to heme and yields linear and sensitive measurements with high signal to noise for human biological samples (Figure 1A).
assay used in this and previous publications5,12,14,15,19,36 prevents NO rebinding to heme and yields linear and sensitive measurements with high signal to noise for human biological samples (Figure 1A).
Acknowledgments
The artistic work of Mrs Lydia Kibiuk is greatly acknowledged.
Prepublished online as Blood First Edition Paper, March 17, 2005; DOI 10.1182/blood-2005-02-0567.
An Inside Blood analysis of this article appears in the front of this issue.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 U.S.C. section 1734.
References
- 1.Moncada S, Palmer RMJ, Higgs EA. Nitric oxide, biology pathophysiology and pharmacology. Pharmacol Rev. 1991;43: 109-142. [PubMed] [Google Scholar]
- 2.Ignarro LJ, Napoli C. Novel features of nitric oxide, endothelial nitric oxide synthase, and atherosclerosis. Curr Atheroscler Rep. 2004;6: 281-287. [DOI] [PubMed] [Google Scholar]
- 3.Kelm M. Nitric oxide metabolism and breakdown. Biochim Biophys Acta. 1999;1411: 273-289. [DOI] [PubMed] [Google Scholar]
- 4.Wang X, Tanus-Santos JE, Reiter CD, et al. Biological activity of nitric oxide in the plasmatic compartment. Proc Natl Acad Sci U S A. 2004;101: 11477-11482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cosby K, Partovi KS, Crawford JH, et al. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med. 2003;9: 1498-1505. [DOI] [PubMed] [Google Scholar]
- 6.Lauer T, Preik M, Rassaf T, et al. Plasma nitrite rather than nitrate reflects regional endothelial nitric oxide synthase activity but lacks intrinsic vasodilator action. Proc Natl Acad Sci U S A. 2001;98: 12814-12819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kleinbongard P, Dejam A, Lauer T, et al. Plasma nitrite reflects constitutive nitric oxide synthase activity in mammals. Free Radic Biol Med. 2003;35: 790-796. [DOI] [PubMed] [Google Scholar]
- 8.Meulemans A, Delsenne F. Measurement of nitrite and nitrate levels in biological samples by capillary electrophoresis. J Chromatogr B Biomed Appl. 1994;660: 401-404. [DOI] [PubMed] [Google Scholar]
- 9.Gorenflo M, Zheng C, Poge A, et al. Metabolites of the L-arginine-NO pathway in patients with left-to-right shunt. Clin Lab. 2001;47: 441-447. [PubMed] [Google Scholar]
- 10.Kosaka H, Tyuma I. Mechanism of autocatalytic oxidation of oxyhemoglobin by nitrite. Environ Health Perspect. 1987;73: 147-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Doyle MP, Pickering RA, DeWeert TM, Hoekstra JW, Pater D. Kinetics and mechanism of the oxidation of human deoxyhemoglobin by nitrites. J Biol Chem. 1981;256: 12393-12398. [PubMed] [Google Scholar]
- 12.Hunter CJ, Dejam A, Blood AB, et al. Inhaled nebulized nitrite is a hypoxia-sensitive NO-dependent selective pulmonary vasodilator. Nat Med. 2004;10: 1122-1127. [DOI] [PubMed] [Google Scholar]
- 13.Nagababu E, Ramasamy S, Albernethy R, Rifkind M. Active nitric oxide produced in the red cell under hypoxic conditions by deoxyhemoglobin-mediated nitrite reduction. J Biol Chem. 2003;278: 46349-46356. [DOI] [PubMed] [Google Scholar]
- 14.Gladwin MT, Shelhamer JH, Schechter AN, et al. Role of circulating nitrite and S-nitrosohemoglobin in the regulation of regional blood flow in humans. Proc Natl Acad Sci U S A. 2000;97: 11482-11487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gladwin MT, Wang X, Reiter CD, et al. S-nitrosohemoglobin is unstable in the reductive red cell environment and lacks O2/NO-linked allosteric function. J Biol Chem. 2002;277: 27818-27828. [DOI] [PubMed] [Google Scholar]
- 16.Feelisch M, Rassaf T, Mnaimneh S, et al. Concomitant S-, N-, and heme-nitros(yl)ation in biological tissues and fluids: implications for the fate of NO in vivo. FASEB J. 2002;16: 1775-1785. [DOI] [PubMed] [Google Scholar]
- 17.Marley R, Patel RP, Orie N, Caeser E, Darley-Usmar V, Moore K. Formation of nanomolar concentrations of S-nitroso-albumin in human plasma by nitric oxide. Free Radic Biol Med. 2001;31: 688-696. [DOI] [PubMed] [Google Scholar]
- 18.Schwoch G, Passow H. Preparation and properties of human erythrocyte ghosts. Mol Cell Biochem. 1973;2: 197-218. [DOI] [PubMed] [Google Scholar]
- 19.Gladwin MT, Schechter AN, Ognibene FP, et al. Divergent nitric oxide bioavailability in men and women with sickle cell disease. Circulation. 2003;107: 271-278. [DOI] [PubMed] [Google Scholar]
- 20.Kelm M. Flow-mediated dilatation in human circulation: diagnostic and therapeutic aspects. Am J Physiol Heart Circ Physiol. 2002;282: H1-H5. [DOI] [PubMed] [Google Scholar]
- 21.May JM, Qu Z-C, Xia L, Cobb CE. Nitrite uptake and metabolism and oxidant stress in human erythrocytes. Am J Physiol Cell Physiol. 2000;279: C1946-C1954. [DOI] [PubMed] [Google Scholar]
- 22.Espey MG, Miranda KM, Thomas DD, et al. A chemical perspective on the interplay between NO, reactive oxygen species, and reactive nitrogen oxide species. Ann N Y Acad Sci. 2002;962: 195-206. [DOI] [PubMed] [Google Scholar]
- 23.Vallance P, Patton S, Bhagat K, et al. Direct measurement of nitric oxide in human beings. Lancet. 1995;345: 153-154. [DOI] [PubMed] [Google Scholar]
- 24.Rhodes PM, Leone AM, Francis PL, Struthers AD, Moncada S. The L-arginine:nitric oxide pathway is the major source of plasma nitrite in fasted humans. Biochem Biophys Res Commun. 1995;209: 590-596. [DOI] [PubMed] [Google Scholar]
- 25.Goedecke A, Decking UKM, Ding Z, et al. Coronary hemodynamics in endothelial NO synthase knockout mice. Circ Res. 1998;82: 186-194. [DOI] [PubMed] [Google Scholar]
- 26.Crawford JH, Chacko BK, Pruitt HM, Piknova B, Hogg N, Patel RP. Transduction of NO-bioactivity by the red blood cell in sepsis: novel mechanisms of vasodilation during acute inflammatory disease. Blood. 2004;104: 1375-1382. [DOI] [PubMed] [Google Scholar]
- 27.Dejam A, Hunter CJ, Schechter AN, Gladwin MT. Emerging role of nitrite in human biology. Blood Cells Mol Dis. 2004;32: 423-429. [DOI] [PubMed] [Google Scholar]
- 28.Perrella M, Shrager RI, Ripamonti M, Manfredi G, Berger RL, Rossi-Bernardi L. Mechanism of the oxidation reaction of deoxyhemoglobin as studied by isolation of the intermediates suggests tertiary structure dependent cooperativity. Biochemistry. 1993;32: 5233-5238. [DOI] [PubMed] [Google Scholar]
- 29.Wanat A, Gdula-Argasinska J, Rutkowska-Zbik D, Witko M, Stochel G, van Eldik R. Nitrite binding to metmyoglobin and methemoglobin in comparison to nitric oxide binding. J Biol Inorg Chem. 2002;7: 165-176. [DOI] [PubMed] [Google Scholar]
- 30.Bryan NS, Rassaf T, Rodriguez J, Feelisch M. Bound NO in human red blood cells: fact or artifact? Nitric Oxide. 2004;10: 221-228. [DOI] [PubMed] [Google Scholar]
- 31.Jensen FB. Nitrite disrupts multiple physiological functions in aquatic animals. Comp Biochem Physiol A Mol Integr Physiol. 2003;135: 9-24. [DOI] [PubMed] [Google Scholar]
- 32.Lundberg JO, Weitzberg E, Cole JA, Benjamin N. Nitrate, bacteria and human health. Nat Rev Microbiol. 2004;2: 593-602. [DOI] [PubMed] [Google Scholar]
- 33.Rassaf T, Bryan NS, Kelm M, Feelisch M. Concomitant presence of N-nitroso and S-nitroso proteins in human plasma. Free Radic Biol Med. 2002;33: 1590-1596. [DOI] [PubMed] [Google Scholar]
- 34.Stamler JS, Simon DI, Osborne JA, et al. S-Nitrosylation of proteins with nitric oxide: synthesis and characterization of biologically active compounds. Proc Natl Acad Sci U S A. 1992;89: 444-448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lim DG, Sweeney S, Bloodsworth A, et al. Nitrolinoleate, a nitric oxide-derived mediator of cell function: synthesis, characterization, and vasomotor activity. Proc Natl Acad Sci U S A. 2002;99: 15941-15946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Reiter CD, Wang X, Tanus-Santos JE, et al. Cell-free hemoglobin limits nitric oxide bioavailability in sickle-cell disease. Nat Med. 2002;8: 1383-1389. [DOI] [PubMed] [Google Scholar]
- 37.Herold S. The outer-sphere oxidation of nitrosyliron(II)hemoglobin by peroxynitrite leads to the release of nitrogen monoxide. Inorg Chem. 2004;43: 3783-3785. [DOI] [PubMed] [Google Scholar]
- 38.Jia L, Bonaventura C, Bonaventura J, Stamler JS. S-nitrosohaemoglobin: a dynamic activity of blood involved in vascular control. Nature. 1996;380: 221-226. [DOI] [PubMed] [Google Scholar]
- 39.Rassaf T, Bryan NS, Maloney RE, et al. NO adducts in mammalian red blood cells: too much or too little? Nat Med. 2003;9: 481-482. [DOI] [PubMed] [Google Scholar]
- 40.Gladwin MT, Lancaster JR Jr, Freeman BA, Schechter AN. Nitric oxide's reactions with hemoglobin: a view through the SNO-storm. Nat Med. 2003;9: 496-500. [DOI] [PubMed] [Google Scholar]
- 41.Dejam A, Kleinbongard P, Rassaf T, et al. Thiols enhance NO formation from nitrate photolysis. Free Radic Biol Med. 2003;35: 1551-1559. [DOI] [PubMed] [Google Scholar]
- 42.Zweier JL, Samouilov A, Kuppusamy P. Non-enzymatic nitric oxide synthesis in biological systems. Biochim Biophys Acta. 1999;1411: 250-262. [DOI] [PubMed] [Google Scholar]
- 43.Baker PR, Schopfer FJ, Sweeney S, Freeman BA. Red cell membrane and plasma linoleic acid nitration products: synthesis, clinical identification, and quantitation. Proc Natl Acad Sci U S A. 2004;101: 11577-11582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jagger JE, Bateman RM, Ellsworth ML, Ellis CG. Role of erythrocyte in regulating local O2 delivery mediated by hemoglobin oxygenation. Am J Physiol Heart Circ Physiol. 2001;280: H2833-2839. [DOI] [PubMed] [Google Scholar]
- 45.Tippler B, Herbst C, Simmet T. Evidence for the formation of endothelin by lysed red blood cells from endogenous precursor. Eur J Pharmacol. 1994;271: 131-139. [DOI] [PubMed] [Google Scholar]
- 46.Moore AF, Gurchinoff S, Brashear W, Bumpus FM, Chang R, Khairallah PA. Angiotensinase activity in red blood cell membranes and intact adrenal cells. Res Commun Chem Pathol Pharmacol. 1977;18: 697-707. [PubMed] [Google Scholar]
- 47.Rogers SC, Khalatbari A, Gapper PW, Frenneaux MP, James PE. Detection of human haemoglobin-bound nitric oxide. J Biol Chem. Prepublished on May 6, 2005, as DOI 10.1074/jbc.M501179200. [DOI] [PubMed]