

Abstract
The molecular mechanisms that regulate the formation of the lymphatic vascular system remain poorly characterized. Whereas studies in embryonic stem (ES) cells have provided major new insights into the mechanisms of blood vessel formation, the development of lymphatic endothelium has not been previously observed. We established embryoid bodies (EBs) from murine ES cells in the presence or absence of lymphangiogenic growth factors. We found that lymphatic endothelial cells develop at day 18 after EB formation. These cells express CD31 and the lymphatic lineage markers Prox-1 and Lyve-1, but not the vascular marker MECA-32, and they frequently sprout from preexisting blood vessels. Lymphatic vessel formation was potently promoted by VEGF-A and VEGF-C but not by bFGF. Our results reveal, for the first time, that ES cells can differentiate into lymphatic endothelial cells, and they identify the EB assay as a powerful new tool to dissect the molecular mechanisms that control lymphatic vessel formation.
Introduction
The lymphatic vascular system plays a pivotal role in mediating tissue fluid homeostasis and cancer metastasis, but the molecular mechanisms that regulate its formation remain poorly characterized. Recent studies in mice deficient in the lymphatic-specific transcription factor Prox-11 have confirmed the hypothesis originally proposed by Florence Sabin more than 100 years ago, which is that in mammals the lymphatic vascular system develops by endothelial cell sprouting from embryonic veins.2,3 Prox-1 plays a major role in the induction of lymphatic cell fate in early lymphatic progenitors that bud from the cardinal veins1 and in the lymphatic reprogramming of differentiated vascular endothelial cells.4-6 Vascular endothelial growth factor-C (VEGF-C) is necessary for the further embryonic development of the lymphatic vascular system.7 Although a number of additional mediators, such as angiopoietin-2,8 neuropilin-2,9 and podoplanin,10 have recently been shown to contribute to lymphatic vessel formation, research into the mechanisms of lymphatic vessel development has been hampered by the frequent embryonic or perinatal lethality observed in the different mutant mouse models.7,10,11
Similar obstacles were encountered in the investigation of blood vessel development. However, it has been found that embryonic stem (ES) cells can differentiate into vascular endothelial cells in monolayer cultures and within the 3-dimensional structure of ES cell-derived embryoid bodies (EBs).12-14 Consecutively, studies in normal and mutant ES cells have provided new insights into the molecular mechanisms of blood vessel formation. In contrast, the development of lymphatic endothelium has not been observed thus far in ES or EB cultures observed for up to 14 days after their formation.15
We hypothesized that, as in normal embryonic development, a longer time period might be needed for the induction of lymphatic endothelial differentiation than for vascular differentiation in EBs and that the formation of lymphatic endothelium might be stimulated by the addition of lymphangiogenic growth factors such as VEGF-C16 and VEGF-A.17,18 We established EBs from murine ES cells and investigated expression of the lymphatic lineage markers Prox-1 and LYVE-1, a hyaluronan receptor, for up to 4 weeks after the initiation of EB formation, in the presence or absence of lymphangiogenic growth factors.
Study design
Cell culture and establishment of EBs
Murine R1 ES cells19 (passages 15-25), 129/Sv-derived, were cultured on mitotically inactivated primary mouse embryonic fibroblasts20 in Dulbecco modified Eagle medium (Gibco, Eggenstein, Germany) supplemented with 15% fetal bovine serum (FBS; Gibco), 1 M HEPES buffer (Sigma, St. Louis, MO), 100 nM sodium pyruvate (Sigma), 0.12% monothioglycerol (Sigma), and 1000 IU/mL recombinant leukemia inhibitory factor (LIF; Chemicon International, Temecula, CA).21 Embryonic fibroblasts and LIF were removed and ES cells were transferred to suspension culture for EB formation, as described.12 After 3 days, EBs of the same size were transferred into 12-well dishes (BD Biosciences, Franklin Lakes, NJ) and were cultured for 14 days without LIF. Then 20 ng/mL recombinant human VEGF-A 165 (R&D Systems, Minneapolis, MN), 200 ng/mL VEGF-C (R&D Systems), or 20 ng/mL basic fibroblast growth factor (bFGF; R&D Systems) was added or not added for 4 or 7 days to media containing 5% or 15% FBS. EBs were fixed in cold 100% methanol at days 14, 18, 21, and 27.
Immunofluorescence and quantitative analyses of vessel development
EBs were stained using antibodies against mouse LYVE-1 (AngioBio, Del Mar, CA), CD31, MECA-32 (BD Biosciences), Prox-1 (Covance, Berkeley, CA), and corresponding secondary antibodies labeled with Alexa-Fluor 488 or 594 (Molecular Probes, Eugene, OR). Cell nuclei were counterstained with 20 g/mL Hoechst bisbenzimide (Sigma). Specimens were examined with a Zeiss Axiovert 200 M microscope, and images were captured with a Zeiss AxioCam-MRm (Zeiss, Oberkochen, Germany), equipped with a Plan Neo 20× dry objective with an aperture of 0.40 (Zeiss) or a Plan Fluotar 40×/1.0 NA oil objective (Zeiss). EBs were also assessed with a Leica-DM-IRBE microscope and a Leica-TCS-4D confocal system (Leica, Wetzlar, Germany). The lymphatic vascular area was determined in EBs stained for CD31 and Prox-1 at 200× magnification, and mean and SEM were calculated for each group. For the relative blood vessel area, 3 fields with the highest vascular densities were evaluated per EB at 50× magnification. Digital images were captured with a Zeiss AxioCam-MRm, and quantitative image analysis was performed using IP-Lab software (Scanalytics, Fairfax, VA). Statistical analysis was performed using the 2-sided unpaired Student t test.
Results and discussion
Development of lymphatic endothelial cells in EBs
Differential immunostains of EBs for the panvascular marker CD31 and for the lymphatic-specific transcription factor Prox-1 or the lymphatic-specific hyaluronan receptor LYVE-1 revealed numerous CD31+/Prox-1-/LYVE-1- vascular endothelial cells 14 days after the initiation of EB formation, whereas no CD31+/Prox-1+/LYVE-1+ lymphatic endothelial cells were observed (data not shown), in agreement with previous studies.15 Beginning at day 18, several vascular structures consisting of CD31+/Prox-1+ lymphatic endothelial cells were detected, with a typical nuclear expression of Prox-1 (Figure 1A-F; day 27). Lymphatic endothelial cells also expressed LYVE-1 (Figure 1G-I; day 21), further confirming their lymphatic lineage differentiation. In contrast, Prox-1+ lymphatic endothelial cells did not express MECA-32, a blood vascular marker (Figure 1M-O; day 21). Occasionally, we found CD31+/LYVE-1+/Prox-1- vascular structures, similar to endothelial cells of the murine cardinal veins at embryonic day 10.5 (E10.5), when budding of Prox-1+ lymphatic progenitors is initiated. Likely, these cells represent vascular endothelial cells at the stage of lymphatic competence.22
Figure 1.
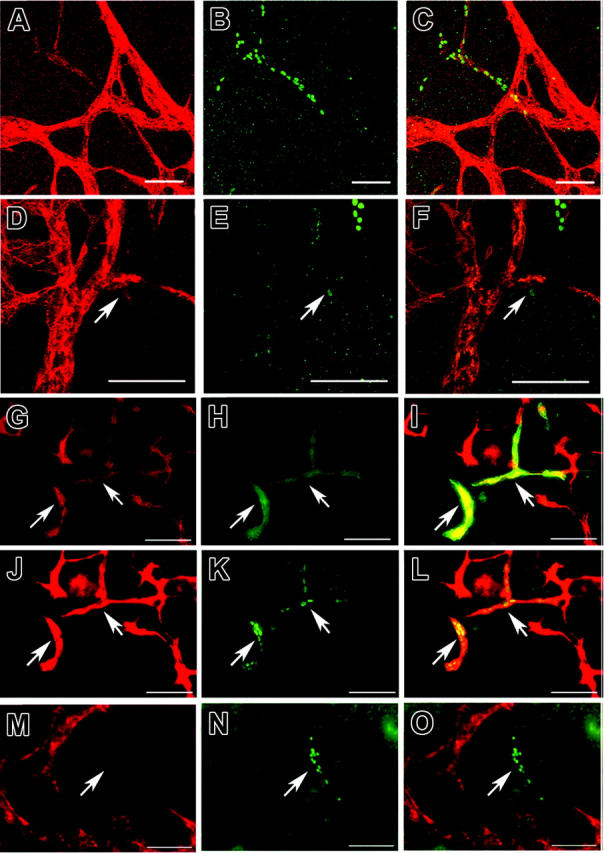
Development of lymphatic endothelial cells in embryoid bodies.. Double-immunofluorescence stains of 27-day-old (A-F) and 21-day-old (G-O) EBs, cultured in the absence of VEGF-A or VEGF-C, for CD31-revealed (red; A,D) and Prox-1-revealed (green; B,E) CD31+ blood vessels and CD31+/Prox-1+ lymphatic vessels (C,F: merged images). Digital sectioning by laser confocal microscopy (D-F) revealed sprouting of lymphatic vessels (arrows) from preexisting blood vessels. Differential immunofluorescence stains for CD31 (red; G,J) and LYVE-1 (green; H) revealed that the CD31+/LYVE-1+ lymphatic vessels also expressed Prox-1 (green; K, arrows). (I,L) Merged images. Blood vessels expressed the vascular-specific marker MECA-32 (red; M), whereas Prox-1+ lymphatic vessels (green; N, arrow) were MECA-32-. (O) Merged image. (J-L) Scale bars, 100 μm (A-L), 50 μm (M-O).
Lymphatic endothelial cells were predominantly found within the central areas of EBs, in the vicinity of areas of blood vessel formation. Frequently, lymphatic endothelial cells appeared to be connected to vascular structures, as evaluated by routine fluorescence microscopy. Digital sectioning with confocal laser microscopy confirmed that CD31+/Prox-1+ lymphatic endothelial cells were often connected to CD31+/Prox-1- vascular structures (Figure 1D-F), indicating that they were budding from the primitive vascular networks; this was followed by sprouting, similar to the early development of lymphatic progenitors during mouse embryogenesis.23 We also observed lymphatic endothelial cells that were not connected to vascular structures, indicating that they either developed independently from lymphatic progenitors or that they had migrated away from their vascular area of origin. Future time-lapse studies of ES cells that express green fluorescence protein under the control of lymphatic lineage-specific promoters such as Prox-1 or LYVE-1 will be needed to further establish the detailed sequence of events during lymphatic vessel formation in EBs.
VEGF-A and VEGF-C promote lymphatic endothelial cell formation in EBs
Previously, VEGF-A, VEGF-C, and bFGF were found to induce lymphatic vessel growth in adult mice.16-18,24 Adding VEGF-A or VEGF-C at day 14 of EB development, for 4 days, potently promoted lymphatic vessel formation, as evaluated by the average number of and the average area covered by lymphatic vascular structures per EB (Figure 2A-B). In contrast, bFGF had no effect (Figure 2A-B), whereas all factors promoted blood vessel growth (data not shown), in accordance with previous results.15,25 After 7 days of treatment, the effect of VEGF-C was more potent than that of VEGF-A (average lymphatic area, VEGF-C, +415%, vs VEGF-A, +115%; average number of lymphatic vascular structures, VEGF-C, +111%, vs VEGF-A, +42%). Future studies are needed to evaluate the relative contributions of VEGFR-2 compared with VEGFR-3 to these effects.
Figure 2.

VEGF-A and VEGF-C, but not bFGF, promote the formation of lymphatic vascular structures in embryoid bodies. EBs were cultured for 14 days in 15% FBS-containing media. At day 14, VEGF-A (20 ng/mL; n = 6), VEGF-C (200 ng/mL; n = 7), or bFGF (20 ng/mL; n = 5) were added or not added (controls, n = 8) for 4 days. EBs were stained for CD31 and Prox-1. (A) Average area occupied by lymphatic vessels per EB. (B) Average number of lymphatic vessels per EB. Data represent mean ± SEM. *P < .05; **P < .01.
Taken together, our results reveal, for the first time, that mouse ES cells can differentiate into lymphatic endothelial cells and that this differentiation is promoted by VEGF-A and VEGF-C. They also identify the EB assay as a simple and powerful new in vitro tool to dissect the molecular mechanisms that control lymphatic vessel formation.
Acknowledgments
We thank L. Nguyen and L. Janes for expert technical assistance.
Prepublished online as. Blood First Edition Paper, September 29, 2005; DOI 10.1182/blood-2005-08-3400.
R.L. designed and performed the research and wrote the paper. F.N. performed the research and analyzed the data. L.L. performed the research and analyzed the data. M.D. designed the research, analyzed the data, and wrote the paper.
Supported by National Institutes of Health grants CA69184, CA86410, and CA92644 (M.D.); American Cancer Society Research Project grant 99-23901 (M.D.); Swiss National Fund grant 3100A0-108207 (M.D.); Fonds für wissenschaftliche Förderung grant S9408-B11 (M.D.); and the Deutsche Krebshilfe (R.L.).
An Inside. Blood analysis of this article appears at the front of this issue.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 U.S.C. section 1734.
References
- 1.Wigle JT, Harvey N, Detmar M, et al. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. EMBO J. 2002;21: 1505-1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sabin F. On the origin of the lymphatic system from the veins and the development of the lymph hearts and thorarcic duct in the pig. Am J Anat. 1902;1: 367-391. [Google Scholar]
- 3.Sabin F. On the development of the superficial lymphatics in the skin of the pig. Am J Anat. 1904;3: 183-195. [Google Scholar]
- 4.Hong YK, Shin JW, Detmar M. Development of the lymphatic vascular system: a mystery unravels. Dev Dyn. 2004;231: 462-473. [DOI] [PubMed] [Google Scholar]
- 5.Hong YK, Foreman K, Shin JW, et al. Lymphatic reprogramming of blood vascular endothelium by Kaposi sarcoma-associated herpesvirus. Nat Genet. 2004;36: 683-685. [DOI] [PubMed] [Google Scholar]
- 6.Petrova TV, Makinen T, Makela TP, et al. Lymphatic endothelial reprogramming of vascular endothelial cells by the Prox-1 homeobox transcription factor. EMBO J. 2002;21: 4593-4599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Karkkainen MJ, Haiko P, Sainio K, et al. Vascular endothelial growth factor C is required for sprouting of the first lymphatic vessels from embryonic veins. Nat Immunol. 2004;5: 74-80. [DOI] [PubMed] [Google Scholar]
- 8.Gale NW, Thurston G, Hackett SF, et al. Angiopoietin-2 is required for postnatal angiogenesis and lymphatic patterning, and only the latter role is rescued by angiopoietin-1. Dev Cell. 2002;3: 411-423. [DOI] [PubMed] [Google Scholar]
- 9.Yuan L, Moyon D, Pardanaud L, et al. Abnormal lymphatic vessel development in neuropilin 2 mutant mice. Development. 2002;129: 4797-4806. [DOI] [PubMed] [Google Scholar]
- 10.Schacht V, Ramirez MI, Hong YK, et al. T1α/podoplanin deficiency disrupts normal lymphatic vasculature formation and causes lymphedema. EMBO J. 2003;22: 3546-3556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dumont DJ, Jussila L, Taipale J, et al. Cardiovascular failure in mouse embryos deficient in VEGF receptor-3. Science. 1998;282: 946-949. [DOI] [PubMed] [Google Scholar]
- 12.Risau W, Sariola H, Zerwes HG, et al. Vasculogenesis and angiogenesis in embryonic-stem-cell-derived embryoid bodies. Development. 1988;102: 471-478. [DOI] [PubMed] [Google Scholar]
- 13.Vittet D, Prandini MH, Berthier R, et al. Embryonic stem cells differentiate in vitro to endothelial cells through successive maturation steps. Blood. 1996;88: 3424-3431. [PubMed] [Google Scholar]
- 14.Wang R, Clark R, Bautch VL. Embryonic stem cell-derived cystic embryoid bodies form vascular channels: an in vitro model of blood vessel development. Development. 1992;114: 303-316. [DOI] [PubMed] [Google Scholar]
- 15.Nilsson I, Rolny C, Wu Y, et al. Vascular endothelial growth factor receptor-3 in hypoxia-induced vascular development. FASEB J. 2004;18: 1507-1515. [DOI] [PubMed] [Google Scholar]
- 16.Jussila L, Alitalo K. Vascular growth factors and lymphangiogenesis. Physiol Rev. 2002;82: 673-700. [DOI] [PubMed] [Google Scholar]
- 17.Hirakawa S, Kodama S, Kunstfeld R, Kajiya K, Brown LF, Detmar M. VEGF-A induces tumor and sentinel lymph node lymphangiogenesis and promotes lymphatic metastasis. J Exp Med. 2005;201: 1089-1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hong YK, Lange-Asschenfeldt B, Velasco P, et al. VEGF-A promotes tissue repair-associated lymphatic vessel formation via VEGFR-2 and the α1β1 and α2β1 integrins. FASEB J. 2004;18: 1111-1113. [DOI] [PubMed] [Google Scholar]
- 19.Nagy A, Rossant J, Nagy R, Abramow-Newerly W, Roder JC. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proc Natl Acad Sci U S A. 1993;90: 8424-8428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wobus AM, Holzhausen H, Jakel P, Schoneich J. Characterization of a pluripotent stem cell line derived from a mouse embryo. Exp Cell Res. 1984;152: 212-219. [DOI] [PubMed] [Google Scholar]
- 21.Williams RL, Hilton DJ, Pease S, et al. Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature. 1988;336: 684-687. [DOI] [PubMed] [Google Scholar]
- 22.Oliver G. Lymphatic vasculature development. Nat Rev Immunol. 2004;4: 35-45. [DOI] [PubMed] [Google Scholar]
- 23.Wigle JT, Oliver G. Prox1 function is required for the development of the murine lymphatic system. Cell. 1999;98: 769-778. [DOI] [PubMed] [Google Scholar]
- 24.Kaipainen A, Korhonen J, Mustonen T, et al. Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc Natl Acad Sci U S A. 1995;92: 3566-3570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Magnusson PU, Ronca R, Dell'Era P, et al. Fibroblast growth factor receptor-1 expression is required for hematopoietic but not endothelial cell development. Arterioscler Thromb Vasc Biol. 2005;25: 944-949. [DOI] [PubMed] [Google Scholar]