

Abstract
An assay based on target cells infected with green fluorescent protein labeled murine cytomegalovirus (GFP-MCMV) and dual color flow cytometry for detecting antibody to MCMV is described. After optimizing conditions for this technique, kinetics of anti-MCMV IgG antibody response were tested in susceptible (BALB/c) and resistant (C57BL/6) mouse strains following primary MCMV infection. Previously published antibody kinetics were confirmed in susceptible mice, with peak IgG response seen ~8 weeks after primary infection, decreasing by 20 weeks after infection. In contrast, MCMV resistant C57BL/6 mice showed significantly lower IgG antibody responses than susceptible mice. Although several techniques have been previously described to detect murine antibody responses to MCMV, including nuclear anti-complement immunofluorescence, viral immunoblotting, complement fixation, indirect immunofluorescence, indirect hemagglutination, and enzyme-liked immunosorbent assay techniques, these techniques are all time consuming and laborious. The technique presented is a simple time efficient alternative to detect previous MCMV antibody responses in experimentally infected mice.
1. INTRODUCTION
Cytomegalovirus (CMV) is a ubiquitous herpes family virus that commonly infects humans. Following a mild self-limited primary infection, CMV subsequently establishes latency wherein infectious virus is undetectable in host tissues until some stimulus causes reactivation. Episodes of reactivation are known to be pathogenic in immunosuppressed populations, such as AIDS patients or transplant recipients (Simmons et al., 1977; Spector et al., 1998). Recent studies in previously immunocompetent critically ill patients have also suggested pathogenicity(Cook et al., 2003; Cook et al., 1998; Curtsinger et al., 1989; Cushing et al., 1993; Heininger et al., 2001; Heininger et al., 2000; Jaber et al., 2005; Papazian et al., 1996). Because of its pathogenic implications, the mechanism by which CMV reactivation occurs has received considerable attention.
Reactivation studies are problematic in human hosts because of obvious ethical limitations, and consequently in-vivo studies of CMV reactivation have required development and use of animal models. Fortunately, murine CMV (MCMV) infection has been well characterized, and is similar to human CMV (Collins et al., 1993; Henson et al., 1966). In susceptible mouse strains, intra-peritoneal inoculation of MCMV causes acute infection, with subsequent development of latency in host tissues (Balthesen et al., 1993; Collins et al., 1993; Gonczol et al., 1985; Kurz et al., 1997; Kurz et al., 1999). MCMV can then be reactivated from latency in-vivo by a variety of stimuli (Bevan et al., 1996; Cook et al., 2002; Furrarah and Sweet, 1994; Gonczol et al., 1985). This model thus affords a unique opportunity to study reactivation of CMV and its pathologic consequences.
For practical reasons, we have become interested in monitoring humoral responses to infection with MCMV as a method to confirm successful primary infection. Viral latency generally requires 4–6 months after primary infection to develop, after which animals are subjected to experimental regimens to study reactivation. These experimental regimens are often quite complex, and thus it is desirable to insure adequacy of primary infection in individual animals prior to embarking on reactivation experiments.
In mice experimentally infected with MCMV, serum evaluation provides an easy avenue to confirm successful primary infection. Numerous techniques have been described that detect MCMV-specific antibody responses in mouse sera following infection with MCMV. These include nuclear anti-complement immunofluorescence, viral immunoblotting, complement fixation, indirect immunofluorescence, indirect hemagglutination, and enzyme-liked immunosorbent assay (ELISA) techniques (Anderson et al., 1983; Anderson et al., 1986; Castellano et al., 1977; Classen et al., 1987; Farrell and Shellam, 1989; Kettering et al., 1977; Lussier et al., 1987; Selgrade et al., 1983). We have found that use of these techniques, which each have their specific strengths and weaknesses, can be time consuming and tedious. Our group has previously described flow cytometry based antibody detection assays for allo-antibody following allogeneic transplantation in clinical as well as experimental settings (Pelletier et al., 2002; VanBuskirk et al., 1998), and felt that development of a similar technique to confirm antibody response to infections with MCMV would be useful.
The biggest obstacle to using flow cytometry to detect MCMV antibody has been identification of infected target cells. Flow cytometry detection of infected target cells has previously required labeled antibodies to MCMV specific antigen. Labeled polyclonal or MCMV specific monoclonal antibodies have been described, but both bind antigenic sites on infected cell surfaces. This binding would potentially compete with anti-MCMV antibody in sera from infected animals, confounding detection of antibodies in sera. This conundrum has prevented development of a simple flow cytometry based technique to detect MCMV specific antibodies. Recently, development of green fluorescent protein labeled MCMV (GFP-MCMV) has made identification of infected cells by flow cytometry a simple matter.
The present investigation describes a technique by which MCMV antibody responses to infection can be confirmed utilizing dual color flow cytometry. Briefly, target cells infected with GFP-MCMV are utilized to detect MCMV specific antibody from sera of mice previously infected with MCMV. Described are factors that may influence measurement of antibody responses utilizing this technique. Finally, kinetics of MCMV antibody responses are described for two mouse strains that are known to be susceptible or resistant to infection.
2. Methods
2.1 Animals and virus
Female BALB/c mice (Harlan, Indianapolis IN) 6–8 weeks of age were used in this study. Purified Smith strain (VR-194/1981) MCMV was obtained from ATCC (Rockville, MD). Primary MCMV infection was achieved by intra-peritoneal (i.p.) injection of 5 × 104 PFU of Smith strain MCMV or 1 × 105 PFU of MCMV deletion mutant for m157 protein, referred to as Δm157 MCMV (kind gift Dr. U. Koszinowski (Bubic et al., 2004)). Sera from infected mice were acquired 2, 4, 8, and 20 weeks after infection. Sera were obtained via retro-orbital puncture under inhalational anesthesia. Sera were stored immediately at −80°C. All mice were housed in a large animal facility, isolated from other mice, adhering to the Guide for the Care and Use of Laboratory Animals prepared by the National Research Council (NIH Publication No. 86–23, revised 1985) following a protocol approval by our Institutional Review Board.
For flow cytometry studies, we utilized MCMV strain RVG102 (a kind gift of Drs Henry and Hamilton). This virus is Smith strain MCMV with the gene for green fluorescent protein inserted in the IE2 position and is designated GFP-MCMV. This virus has been shown to express GFP during acute infection and reactivation, and the growth characteristics of this virus in vitro and in vivo are similar to that of wild type virus (Henry et al., 2000).
2.2 Target cells
Murine embryo fibroblast NIH/3T3 cells (CRL-1658) were obtained from ATCC (Manassas, VA). Cells were grown to confluence in T-175mm flasks using Dulbecco’s Modified Eagle Medium containing 10% normal calf serum (Gibco, Carlsbad, CA). Cells were dislodged from flasks, rinsing with cold phosphate buffered saline, followed by trypsinization, then split 1:5 and allowed to grow to confluence. One day after subculture, cells were infected with purified GFP-MCMV at multiplicity of infection of 1. Prior to use in the MCMV-reactive antibody detection assay, cells were confirmed to be optimally infected by detection of GFP in at least 50% of cells by flow cytometric analysis using an EPICS XL flow cytometer (Beckman Coulter, Miami, FL). Cells usually required 2–3 days to reach this level of infection.
2.3 Flow cytometry
Presence of MCMV-reactive IgG1 antibody was determined by the ability of sera to bind with GFP-MCMV infected 3T3 fibroblasts. Infected target cells were washed 3x in PBS-FACS (PBS, 5% FCS) and adjusted to 0.5 million per sample tube. Experimental sera were then added at a 1:20 dilution in PBS-FACS consistent with previous investigations (Classen et al., 1987). Following one hour incubation on ice, target cells were washed 3x in PBS-FACS. R-phycoerythrein (PE)-conjugated rat anti-mouse IgG1 (BD Biosciences, San Jose, CA) was then added at a 1:100 dilution in PBS-FACS. Anti-murine IgG1 antibody was chosen because preliminary studies using non-type specific anti-murine IgG antibody showed higher levels of non-specific binding to MCMV infected cells (7–8%, data not shown). Following 1 hour incubation on ice in the dark, target cells were washed 3x in PBS-FACS and then fixed in 0.5 ml of PBS-FACS containing 1% formalin. Binding of anti-mouse IgG1 was detected by flow cytometry using an EPICS XL flow cytometer (Beckman Coulter, Miami, FL). Results were derived from analysis of 5000 gated events of GFP-MCMV infected NIH/3T3 fibroblasts and are shown as the percentage of GFP-MCMV infected NIH/3T3 fibroblasts that bound detectable MCMV-reactive antibody. Controls for each flow cytometry study included infected target cells alone, infected cells with secondary antibody only, and infected cells with naïve BALB/c or C57BL/6 sera as applicable.
2.4 ELISA
MCMV virions were purified from infected cell supernatants using low speed centrifugation (1500 rpm, 10 min) to clarify, then high speed centrifugation at 20,000 rpm for 90 min after layering on 15% sucrose/PBS cushion in SW28 (Beckman) tubes. Supernatants and sucrose cushions were then aspirated, and virus pellets re-suspended in PBS and quantified. Purified virus (0.25 μg/100 μL/well) was added to 96-well plates (Nunc MaxiSorp) and incubated overnight at 4°C. For ELISA assays, plates were washed with PBS-T and blocked with 5% non-fat milk/PBS for 1 hour at 37°C. After washing, diluted primary antisera (100 μL) were added to wells at concentrations of 1:20, 1:40, 1:80, 1:160, 1:320, 1:640, and 1:1320, then incubated at room temperature for 2 hours. Wells were washed with PBS-T, secondary antibody (goat anti-mouse IgG1-AP conjugate) was added at a dilution of 1:1000 to each well, and plates were incubated at room temperature for 1 hour. After extensive PBS washing, AP substrate (Zymogen) was added to each well. After approximately 20 minutes, absorption of each well at 405 nm was determined using a Wallace Victor 3 plate reader (Perkin Elmer).
2.5 Statistical Analyses
Statistical analyses utilized two tailed Students t-test. P-values < 0.05 were considered significant for all testing. Means are expressed as mean + standard error. Statistical software used was Graphpad Prism (Version 4.03, GraphPad Software, San Diego CA).
3. Results
3.1 Detection of fibroblasts infected with GFP-MCMV
Fibroblasts that were not infected or that were infected with GFP-MCMV (Henry et al., 2000) were analyzed by flow cytometry. Results are shown in figure 1A&B respectively, with infected cells showing a significant shift along the GFP axis (figure 1B). For this particular example, the percent of infected cells measured ~83%. Of interest, there are two populations seen in figure 1B that are distinct from non-infected cells. The smaller peak was thought to represent cells infected with nascent virus produced in these cultures, cells infected with virus that fail to express GFP, or possibly non infected cells in contact with infected cells or other factors in the culture that affect forward and side scatter. Further analysis of this small population showed that ~60% of these cells bind MCMV antibody (not shown), and thus this population likely represents cells at relatively early stages of infection or exposed to recombinants that fail to express GFP. In either case, we conclude that cells infected with GFP-MCMV can be detected by flow cytometry
Figure 1. Detection of cells infected with GFP-MCMV by flow cytometry.
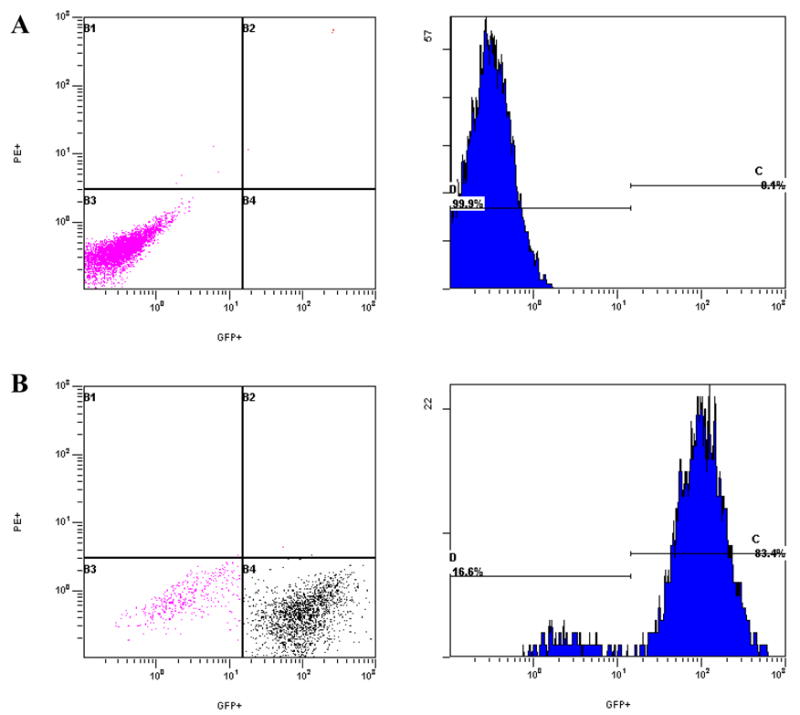
Histograms and dot plots illustrate green fluorescent protein (GFP) detection in uninfected fibroblasts and fibroblasts infected with GFP-labeled murine cytomegalovirus (GFP-MCMV). A. Uninfected NIH-3T3 fibroblasts. B. GFP-MCMV-infected NIH-3T3 fibroblasts.
3.2 Detection of MCMV Antibody
Because infected cells are known to express CMV antigens on their surface (Holtappels et al., 2002; Loh et al., 1991; Pereira et al., 1982; Stinski et al., 1979; Sullivan-Tailyour and Garnett, 1986; Tripathy et al., 2006), infected cells were expected to serve as suitable targets for MCMV antibody in sera from infected mice. This required exposure of target cells to MCMV reactive sera, and identification of antigen bound anti-CMV antibodies using PE-labeled mouse anti-IgG1 secondary antibody. This is conceptually illustrated in figure 2. Thus cells which co-localize both GFP and PE were considered positive for MCMV antibody binding.
Figure 2. Schematic of flow cytometry technique to measure murine antibody responses to MCMV.
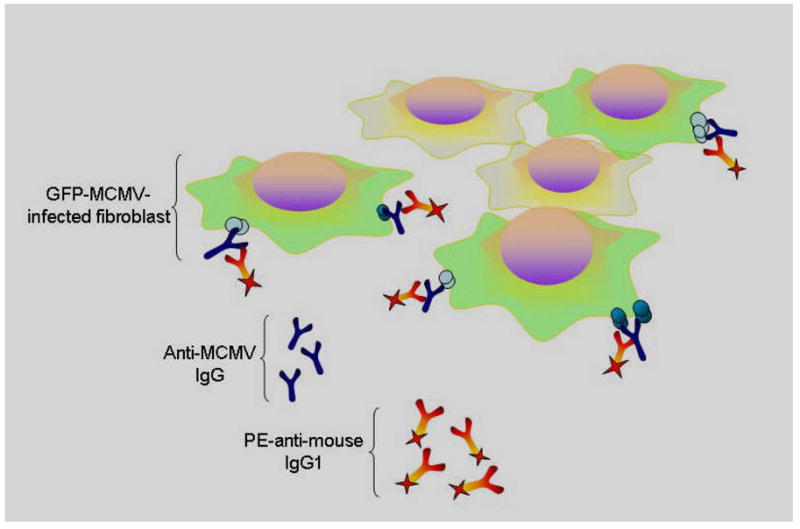
Cells infected with green fluorescent protein labeled murine cytomegalovirus (GFP-MCMV), shown as green above, are detectable by flow cytometry. These GFP-MCMV infected target cells are incubated with sera from previously infected mice, which contains anti-MCMV antibody, along with phycoerythrein labeled anti-murine IgG1 antibody. Cells that co-localize these fluorophores are detectable by dual color flow cytometry, providing a method to detect development anti-MCMV antibodies in mice.
To test this hypothesis, 3T3 cells infected with GFP-MCMV were exposed to PE labeled anti-IgG1 antibody and evaluated by flow cytometry as a negative control. As shown in figure 3A, there was little non-specific secondary antibody binding to target cells. Target cells were then incubated with sera from BALB/c mice previously infected with MCMV (5×104 pfu i.p.), and then with PE labeled murine anti-igG1 antibody. Infected cells binding antibody were identified by dual color, and ~95% of infected cells (GFP+) bound antibody for these positive controls (figure 3B). Infected target cells alone, infected target cells with secondary antibody alone, and infected target cells exposed to naïve BALB/c or C57BL/6 sera and secondary antibody consistently showed very low binding (Table 1). In addition, non-infected cells tested with MCMV reactive or naïve sera and PE-labeled secondary, or PE-labeled secondary alone showed <1% binding (data not shown). Low-level binding of naïve sera to infected cells is important because it demonstrates that antibody in CMV infected sera is not binding non-specifically to virally-encoded Fc-receptor proteins (Crnkovic-Mertens et al., 1998; Thale et al., 1994). Thus we conclude that anti-CMV IgG1 antibody response can be measured using this technique.
Figure 3. Dual color flow cytometry to detect anti-MCMV antibodies.
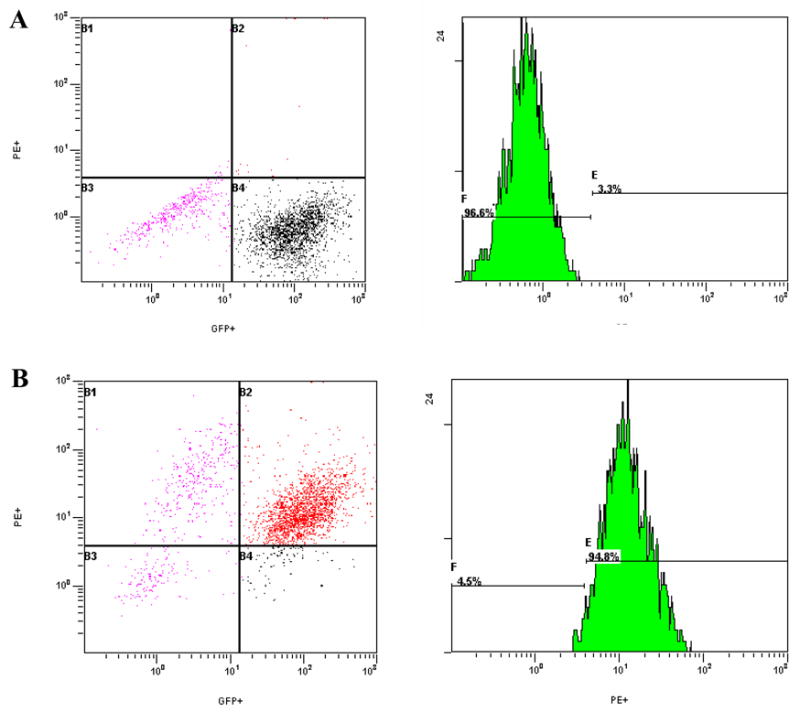
Shown are dot plots and histograms comparing reactivity of green fluorescent labeled murine cytomegalovirus (GFP-MCMV) infected target cells with naïve mouse sera (A) or sera from mice previously infected with Smith strain MCMV (B). Target cells infected with GFP-MCMV were first incubated with sera from uninfected or MCMV-infected mice, rinsed, and then incubated with PE-labeled antibody to murine IgG1.
Table 1. Specificity of flow cytometry assay to measure anti-MCMV antibodies in mouse sera.
| n | Mean target cell binding | Std. Error | |
|---|---|---|---|
| MCMV-infected BALB/c sera | 31 | 93.48 | 0.90 |
| Naive BALB/c sera | 39 | 2.72 | 0.24 |
| Naive BL6 sera | 19 | 3.11 | 0.43 |
| Secondary Ab1 | 18 | 2.00 | 0.31 |
| Cells only2 | 18 | 1.67 | 0.32 |
GFP-MCMV-infected NIH 3T3 cells incubated only with PE-labeled secondary antibody only.
GFP-MCMV-infected NIH 3T3 cells only
3.3 Effect of percent infected targets on antibody detection
In the course of our studies, we discovered variability with time in percentages of target cells expressing GFP after infection. It was unclear whether overall percentage of infected cells would influence detection of anti-CMV antibody. Typically, one day after infection, 15–25% of cells were GFP-positive, and by two days after infection, 80–95% were GFP-positive. It is conceivable that at different times after infection, differential surface protein expression may occur, and that this might influence antibody binding and detection.
To test this possibility, cells were exposed to GFP-MCMV and analyzed by flow cytometry at 24 and 48 hrs pi for antibody binding. At 24 hr after infection 19% of the target cells were positive for GFP, and this number rose to 98% at 48 hr after infection. Positive control sera binding was not significantly different at either time point (figure 4). Although it takes longer to perform the flow analyses when a lower percentage of cells are infected, we conclude that percentage of cells expressing GFP does not influence measurement of antibody binding to target cells.
Figure 4. Target cells analyzed 24 or 48 hr after infection show identical antibody binding profiles.
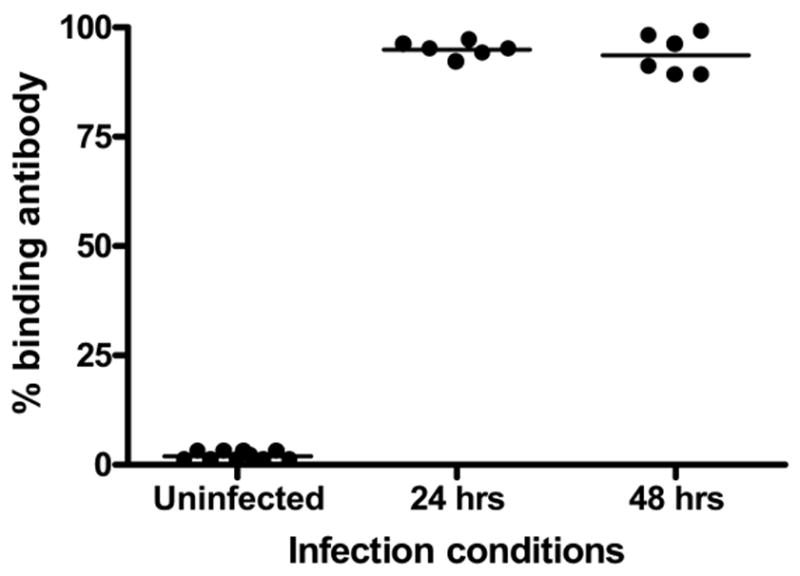
NIH-3T3 cells exposed to green fluorescent protein (GFP) labeled murine cytomegalovirus (MCMV) for 24 or 48 hours were evaluated as targets for antibody binding using MCMV-reactive standard sera and phycoerythrein (PE)-labeled anti-murine IgG1 secondary. Antibody binding is expressed as percentage of GFP positive cells which co-localized PE.
3.4 Optimum number of target cells determination
We next evaluated the optimal number of cells for each flow cytometry analysis. All initial studies were performed utilizing 1×106 target cells for each analysis. We therefore tested percent for binding of positive control sera to 1×106, 5×105, and 2×105 target cells. There were no significant differences in binding between 1×106 and 5×105 target cells, but there was slightly but significantly lower percent binding seen when 2×105 target cells were used (data not shown, students t-test p<0.05). We conclude that 5×105 target cells are optimal for each serum sample evaluation.
3.5 Technique sensitivity & Specificity
To determine sensitivity of this method, serial dilutions of positive control sera were performed. Initial studies were performed with a 1:20 dilution of sera for each analysis, thus subsequent dilutions included 1:40, 1:80, 1:160, 1:320, 1:640, and 1:1320. We could consistently detect antibody binding to target cells for all dilutions up to 1:160 (Figure 5A). More dilute sera samples (1:320 and beyond) were indistinguishable from negative controls. ELISA analysis allowed detection at dilutions up to 1:320, becoming indistinguishable from negative controls beyond that (Figure 5B). These results are similar to ELISA results from other investigators (Shanley et al., 1981). A dilution of 1:20 for positive controls consistently gave 93–98% target cell binding and was thus chosen for all subsequent studies.
Figure 5. Comparison of flow cytometry and ELISA techniques.
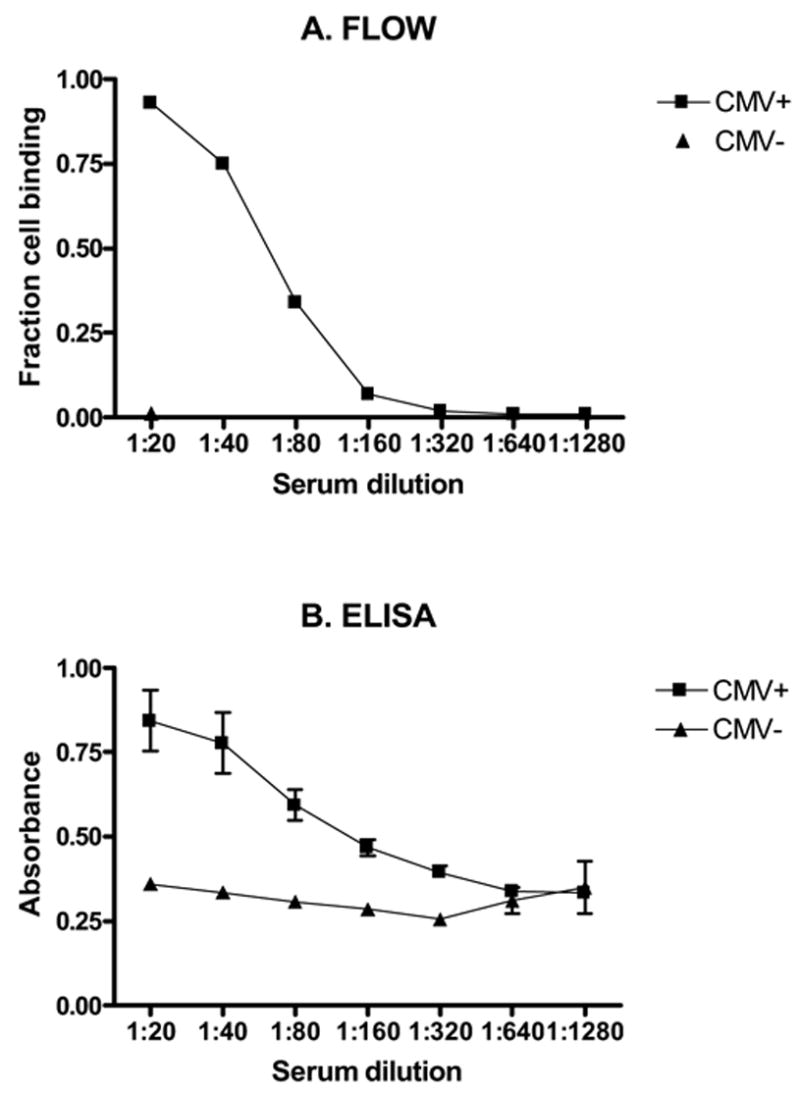
Sensitivity of flow cytometry (A) and enzyme linked immunosorbent assays (ELISA) (B) techniques were compared using serially diluted sera from BALB/c mice previously infected with murine cytomegalovirus (MCMV). Flow cytometry allowed detection of MCMV antibody to a dilution of 1:160. ELISA allowed detection of antibody to a dilution of 1:320. Results represent means of three runs for each dilution.
Statistical sensitivity and specificity were also estimated for this flow cytometry technique. Based upon our previous work (VanBuskirk et al., 1998), and because our negative controls had very tight distributions, we considered < 5% cell binding to be background (negative), and >5% cell binding to be positive. Sera from all previously infected mice tested positive for target cell binding (80/80), making sensitivity for the technique 100% (95% confidence intervals 94–100%). Similarly, all naïve sera tested were negative for target cell binding (58/58), giving a specificity of 100% (95% confidence interval 92–100%).
3.6 Kinetics of antibody response
To test the technique experimentally, cohorts of n=10 BALB/c or C57BL/6 mice were infected with 5104 pfu of MCMV, and blood was obtained at serial time points of 2, 4, 8, and 20 weeks after primary infection. Development of anti-CMV antibody responses over time is illustrated in figure 6. BALB/c mice develop detectable anti-CMV antibody responses by 2 weeks pi, increasing in magnitude through 8 weeks after infection (figure 6). At this time, percentages of target cells positive for antibody binding (96%) were comparable to control sera (98% data not shown). This is followed by a slight decrease in antibody titer by week 20, which has been described by other authors (Classen et al., 1987; Lussier et al., 1987).
Figure 6. Comparison of MCMV IgG1 antibody responses to infection in different mouse strains.
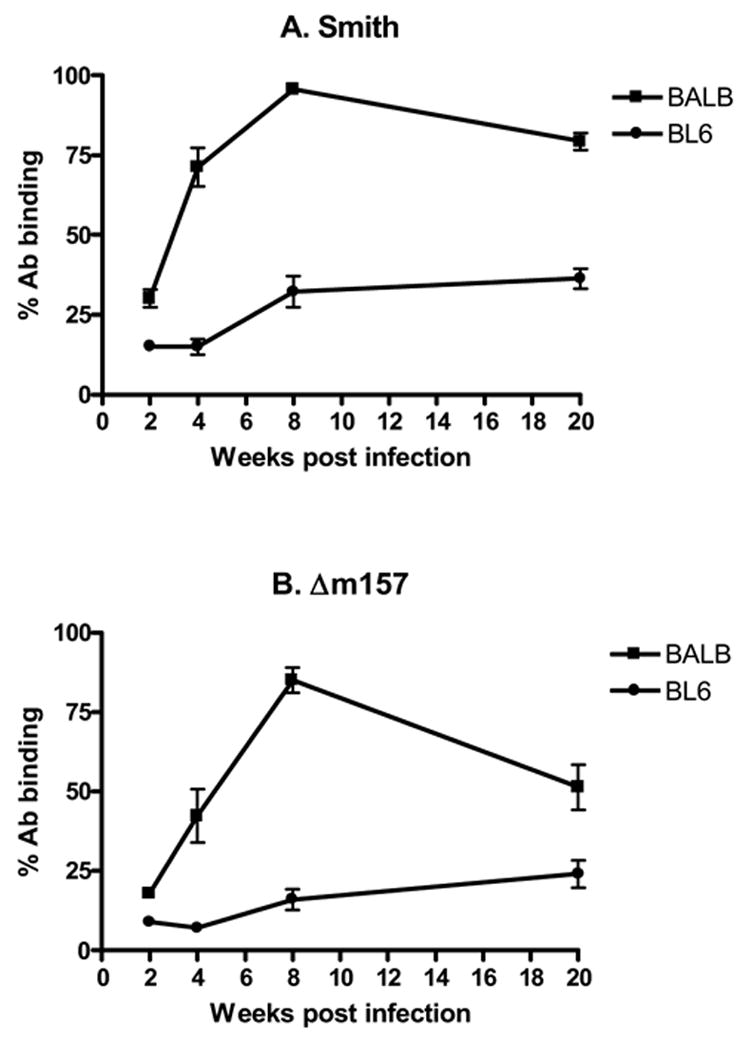
Mice were injected with A. 5 × 104 PFU of Smith strain murine cytomegalovirus (MCMV), or B. 1 × 105 m157 deletion mutant MCMV (Δm157). Sera were collected and analyzed 2, 4, 8, and 20 weeks after infection for MCMV specific antibody by flow cytometry. A. MCMV antibody responses to Smith MCMV infection were significantly higher in BALB/c mice than in C57BL/6 mice at all time points (all p<0.0001). B. Similarly, BALB/c mice infected with Δm157 MCMV had significantly higher antibody responses than C57BL/6 mice at all time points (all p<0.003), despite having equal tissue viral titers (not shown). Antibody binding is expressed as percentage of GFP infected target cells that co-localized anti-MCMV antibody. Each data point represents results from n=10 mice.
Interestingly, C57BL/6 mice had attenuated MCMV IgG1 antibody responses compared with BALB/c mice, with significantly lower antibody binding at each time point (figure 6A). Because C57BL/6 mice are MCMV resistant, we postulated that lower viral titers might explain this lower antibody response. C57BL/6 resistance to MCMV has been recently localized to MCMV gene m157(Arase et al., 2002; Brown et al., 2001; Bubic et al., 2004; Smith et al., 2002; Voigt et al., 2003). Thus to test this hypothesis, cohorts of C57BL/6 and BALB/c mice were infected with Δm157 MCMV, and sera were studied 2, 4, 8, and 20 weeks after infection for MCMV antibodies. Concomitantly infected mice (cohorts of 5) were evaluated 4 weeks after infection by plaque assays, and confirmed equal virus titers in both mouse strains (data not shown). Despite equal viral titers in both mouse strains, C57BL/6 mice still developed significantly lower antibody responses than BALB/c mice at all time points studied (Figure 6B). We thus conclude that this technique will detect antibody responses in either mouse strain for at least 5 months after primary infection, and that C57BL/6 mice develop lower IgG1 antibody responses to MCMV infection.
4. DISCUSSION
These data describe a relatively quick and easy method to confirm anti-MCMV antibody responses in murine sera. Advantages of the technique include rapid results and ease of performance, as well as flexibility in running small or large numbers of samples with equal ease. We typically utilize the technique to confirm previous infection with MCMV for our animal studies. Mice that demonstrate low MCMV antibody titers 8–12 weeks after infection are subsequently excluded from labor intensive reactivation studies.
MCMV humoral responses have been fairly well characterized by previous investigations. Polyclonal antibody responses to multiple viral proteins occur following MCMV infection, with production of virus specific antibodies of both IgG and IgM classes (Araullo-Cruz et al., 1978; Farrell and Shellam, 1989; Karupiah et al., 1998; Lawson et al., 1988; Selgrade et al., 1983). Adoptive transfer of anti-CMV antibody has been shown to be protective during acute infection (Araullo-Cruz et al., 1978; Farrell and Shellam, 1991; Lawson et al., 1988; Shanley et al., 1981), but does not prevent development of latent infection (Shanley et al., 1981). Although preformed antibody does help protect from lethal primary infection, it is not required for viral control in non-lethal models, but does appear to be important in viral control after reactivation from latency (Jonjic et al., 1994).
Previous kinetic studies have shown that development of an MCMV specific IgG response begins 5–10 days after primary infection, and peaks ~45–90 days after primary infection (Araullo-Cruz et al., 1978; Classen et al., 1987; Gonczol et al., 1985; Lawson et al., 1988; Selgrade et al., 1983). Data from our study corroborate these previous findings, with BALB/c mice developing demonstrable IgG1 antibody by 2 weeks after infection, increasing to maximum by 8 weeks then beginning to decline by 20 weeks after infection (figure 6A). C57BL/6 mice showed a somewhat different pattern, with significantly slower IgG1 antibody kinetics than BALB/c mice after infection (figure 6A). The only published antibody responses to MCMV in C57BL/6 mice that we are aware of have been short term studies, up to ~45 days pi (Jonjic et al., 1994), and there appear to be no comparative data for BALB/c and C57BL/6 mice in the literature using any technique. It thus appears that C57BL6 mice may have a less prominent antibody response to MCMV infection.
This diminished antibody response in C57BL/6 mice is consistent with previous studies showing that other MCMV resistant mouse strains develop less antiviral antibody than susceptible mice (Lawson et al., 1988). C57BL/6 mice exhibit lower viral titers in tissues than BALB/c mice after primary infection (Chalmer et al., 1977; Grundy et al., 1981; Scalzo et al., 1990), and thus lower anti-viral antibody responses to wild type virus could simply reflect a lower viral antigen load. To eliminate this possibility, we utilized a second strain of MCMV with deletion of gene m157. Despite equal viral titers for this Δm157 virus in BALB/c and C57BL/6 mice, C567BL6 mice had lower IgG1 antibody responses than BALB/c mice (Figure 6B). These results suggest that development of humoral responses in C57BL/6 mice after MCMV infection is not simply an issue of viral load, at least for the IgG1 subclass. C57BL/6 mice develop strong IgG1 antibody responses to other antigens (VanBuskirk et al., 1998), so generic impairment of antibody formation unique to C57BL/6 mice cannot explain the lower responses to MCMV. Cytokine responses are known to influence antibody isotype switching (reviewed in (Stavnezer, 1996)), so it is possible that this observed difference between BALB/c and C57BL/6 IgG1 antibody responses may be a consequence of different innate cytokine responses to MCMV infection. The influence of cytokines and isotype switching in this model system is a subject of ongoing study in our group. For now, this decreased IgG1 antibody response in C57BL/6 mice remains somewhat of a curiosity.
Previous investigators have argued that if humoral responses are important for viral control, then higher antibody titers and earlier kinetics would be characteristic in that mouse strain (Lawson et al., 1988). Conversely, if humoral antiviral responses were not important to a particular strain, then antibody levels might be lower and slower to develop. If these hypotheses are true, then one could conclude from our results that humoral responses are important in viral control during acute infection in BALB/c mice. Passive immunization studies have shown reduction in mortality when a lethal dose of MCMV was given (Araullo-Cruz et al., 1978; Farrell and Shellam, 1991; Lawson et al., 1988; Shanley et al., 1981), but there are no data that we are aware of directly studying the importance of antibody responses in viral control in the BALB/c strain. Work by Jonic et al using B-cell deficient mice clearly supports the hypothesis that humoral responses are dispensable during primary MCMV infection (Jonjic et al., 1994), but B-cell knockout mice used in these studies had a C57BL/6 background. C57BL/6 mice have well described alternative pathways to control MCMV (Dokun et al., 2001; Rodriguez et al., 2004; Scalzo et al., 1990), and therefore extrapolating these results to BALB/c mice might be inappropriate. It is therefore possible that antibody responses in BALB/c mice are more important in viral control than in C57BL/6 mice. This area of research needs further exploration before definitive conclusions can be drawn.
Previously the most sensitive described technique to detect MCMV antibody response after infection has been ELISA. Our ELISA results are consistent with those published by others(Shanley et al., 1981), and suggest that ELISA should remain the “gold standard” for absolute sensitivity. Unfortunately, there is some controversy with this technique possibly being overly sensitive. One previous investigation using ELISA suggested an incidence of MCMV infection as high as 55% in animals acquired from multiple vendors (Anderson et al., 1986). In contrast, investigations published using a similar ELISA technique during that same time period suggested that this incidence was overestimated (Classen et al., 1987). In addition to this potential over-sensitivity, we have been frustrated with the laborious nature of ELISA, which often requires 5–10 days in reagent preparation. In contrast, for those with ready access to flow cytometry, our described technique requires only 1–2 days to infect cells. Because our aim was to develop a less time consuming technique to screen previously infected animals for successful experimental infection, we are satisfied that the sensitivity achieved using flow cytometry suits that purpose.
Specificity of this technique was not completely studied; therefore we recommend that this technique should not stand alone as a surveillance tool for MCMV infection. It is unknown whether other viral antibodies will cross react with MCMV infected target cells to give false positives. Should investigators be concomitantly utilizing other viruses, particularly herpes family viruses, cross reactivity studies should be performed before relying upon this method. Based upon previously published work using ELISA or other previously mentioned techniques, it is unlikely that significant cross reactivity will occur with MCMV infected cells. Nevertheless, absent definitive experimental data, we recommend that our technique should not be used alone as a definitive diagnostic test for MCMV. Accordingly, all of our animals undergo confirmation of previous infection/reactivation studies utilizing sensitive PCR techniques at conclusion of reactivation experiments.
In summary, an assay based on target cells infected with GFP-MCMV and dual color flow cytometry for detecting antibody to MCMV is described. This technique is less labor intensive and more efficient than previously described molecular techniques. It is sensitive and specific enough to be utilized for screening animals experimentally infected with MCMV, and provides a simple method to confirm successful primary infection.
Acknowledgments
The authors would like to thank Kelly Nye and Jonathan Campbell for their technical assistance in flow cytometry.
Footnotes
This work was supported by NIH grant R01 AI053094.
Bibliography
- Anderson CA, Murphy JC, Fox JG. Evaluation of murine cytomegalovirus antibody detection by serological techniques. Journal of Clinical Microbiology. 1983;18:753–8. doi: 10.1128/jcm.18.4.753-758.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson CA, Murphy JC, Fox JG. Surveillance of mice for antibodies to murine cytomegalovirus. J Clin Microbiol. 1986;23:1152–1154. doi: 10.1128/jcm.23.6.1152-1154.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arase H, Mocarski ES, Campbell AE, Hill AB, Lanier LL. Direct Recognition of Cytomegalovirus by Activating and Inhibitory NK Cell Receptors. Science. 2002;296:1323–1326. doi: 10.1126/science.1070884. [DOI] [PubMed] [Google Scholar]
- Araullo-Cruz TP, Ho M, Armstrong JA. Protective effect of early serum from mice after cytomegalovirus infection. Infect Immun. 1978;21:840–842. doi: 10.1128/iai.21.3.840-842.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balthesen M, Messerle M, Reddehase MJ. Lungs are a major organ site of cytomegalovirus latency and recurrence. Journal of Virology. 1993;67:5360–6. doi: 10.1128/jvi.67.9.5360-5366.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevan IS, Sammons CC, Sweet C. Investigation of murine cytomegalovirus latency and reactivation in mice using viral mutants and the polymerase chain reaction. Journal of Medical Virology. 1996;48:308–20. doi: 10.1002/(SICI)1096-9071(199604)48:4<308::AID-JMV3>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- Brown MG, Dokun AO, Heusel JW, Smith HRC, Beckman DL, Blattenberger EA, Dubbelde CE, Stone LR, Scalzo AA, Yokoyama WM. Vital Involvement of a Natural Killer Cell Activation Receptor in Resistance to Viral Infection. Science. 2001;292:934–937. doi: 10.1126/science.1060042. [DOI] [PubMed] [Google Scholar]
- Bubic I, Wagner M, Krmpoti A, Saulig T, Kim S, Yokoyama WM, Jonji S, Koszinowski UH. Gain of Virulence Caused by Loss of a Gene in Murine Cytomegalovirus. J Virol. 2004;78:7536–7544. doi: 10.1128/JVI.78.14.7536-7544.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castellano GA, Hazzard GT, Madden DL, Sever JL. Comparison of the enzyme-linked immunosorbent assay and the indirect hemagglutination test for detection of antibody to cytomegalovirus. Journal of Infectious Diseases. 1977;136(Suppl):S337–40. doi: 10.1093/infdis/136.supplement_2.s337. [DOI] [PubMed] [Google Scholar]
- Chalmer JE, Mackenzie JS, Stanley NF. Resistance to murine cytomegalovirus linked to the major histocompatibility complex of the mouse. Journal of General Virology. 1977;37:107–14. doi: 10.1099/0022-1317-37-1-107. [DOI] [PubMed] [Google Scholar]
- Classen DC, Morningstar JM, Shanley JD. Detection of antibody to murine cytomegalovirus by enzyme-linked immunosorbent and indirect immunofluorescence assays. Journal of Clinical Microbiology. 1987;25:600–4. doi: 10.1128/jcm.25.4.600-604.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins T, Pomeroy C, Jordan MC. Detection of latent cytomegalovirus DNA in diverse organs of mice. Journal of Infectious Diseases. 1993;168:725–9. doi: 10.1093/infdis/168.3.725. [DOI] [PubMed] [Google Scholar]
- Cook C, Zhang X, McGuinness B, Lahm M, Sedmak D, Ferguson R. Intra-abdominal Bacterial Infection Reactivates Latent Pulmonary Cytomegalovirus in Immunocompetent Mice. Journal of Infectious Diseases. 2002;185:1395–1400. doi: 10.1086/340508. [DOI] [PubMed] [Google Scholar]
- Cook CH, Martin LC, Yenchar JK, Lahm MC, McGuinness B, Davies EA, Ferguson RM. Occult herpes family viral infections are endemic in critically ill surgical patients. Critical Care Medicine. 2003;31:1923–9. doi: 10.1097/01.CCM.0000070222.11325.C4. [DOI] [PubMed] [Google Scholar]
- Cook CH, Yenchar JK, Kraner TO, Davies EA, Ferguson RM. Occult herpes family viruses may increase mortality in critically ill surgical patients. American Journal of Surgery. 1998;176:357–60. doi: 10.1016/s0002-9610(98)00205-0. [DOI] [PubMed] [Google Scholar]
- Crnkovic-Mertens I, Messerle M, Milotic I, Szepan U, Kucic N, Krmpotic A, Jonjic S, Koszinowski UH. Virus attenuation after deletion of the cytomegalovirus Fc receptor gene is not due to antibody control. Journal of Virology. 1998;72:1377–82. doi: 10.1128/jvi.72.2.1377-1382.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtsinger LJ, Cheadle WG, Hershman MJ, Cost K, Polk J, Hiram C. Association of cytomegalovirus infection with increased morbidity is independent of transfusion. The American Journal of Surgery. 1989;158:606–611. doi: 10.1016/0002-9610(89)90204-3. [DOI] [PubMed] [Google Scholar]
- Cushing D, Elliot S, Caplan E, Batlas S, Sridhara R, Wade J. Herpes Simplex Virus and Cytomegalovirus Excretion Associated with Increased Ventilator Days in Trauma Patients. Journal of Trauma. 1993;25:161. [Google Scholar]
- Dokun AO, Kim S, Smith HR, Kang HS, Chu DT, Yokoyama WM. Specific and nonspecific NK cell activation during virus infection. Nature Immunology. 2001;2:951–6. doi: 10.1038/ni714. [DOI] [PubMed] [Google Scholar]
- Farrell HE, Shellam GR. Immunoblot analysis of the antibody response to murine cytomegalovirus in genetically resistant and susceptible mice. Journal of General Virology. 1989;70:2573–86. doi: 10.1099/0022-1317-70-10-2573. [DOI] [PubMed] [Google Scholar]
- Farrell HE, Shellam GR. Protection against murine cytomegalovirus infection by passive transfer of neutralizing and non-neutralizing monoclonal antibodies. J Gen Virol. 1991;72:149–156. doi: 10.1099/0022-1317-72-1-149. [DOI] [PubMed] [Google Scholar]
- Furrarah AM, Sweet C. Studies of the pathogenesis of wild-type virus and six temperature-sensitive mutants of mouse cytomegalovirus. Journal of Medical Virology. 1994;43:317–30. doi: 10.1002/jmv.1890430402. [DOI] [PubMed] [Google Scholar]
- Gonczol E, Danczig E, Boldogh I, Toth T, Vaczi L. In vivo model for the acute, latent and reactivated phases of cytomegalovirus infection. Acta Microbiologica Hungarica. 1985;32:39–47. [PubMed] [Google Scholar]
- Grundy JE, Mackenzie JS, Stanley NF. Influence of H-2 and non-H-2 genes on resistance to murine cytomegalovirus infection. Infect Immun. 1981;32:277–286. doi: 10.1128/iai.32.1.277-286.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heininger A, Jahn G, Engel C, Notheisen T, Unertl K, Hamprecht K. Human cytomegalovirus infections in nonimmunosuppressed critically ill patients. Critical Care Medicine. 2001;29:541–7. doi: 10.1097/00003246-200103000-00012. [DOI] [PubMed] [Google Scholar]
- Heininger A, Vogel U, Aepinus C, Hamprecht K. Disseminated fatal human cytomegalovirus disease after severe trauma. Critical Care Medicine. 2000;28:563–6. doi: 10.1097/00003246-200002000-00046. [DOI] [PubMed] [Google Scholar]
- Henry SC, Schmader K, Brown TT, Miller SE, Howell DN, Daley GG, Hamilton JD. Enhanced green fluorescent protein as a marker for localizing murine cytomegalovirus in acute and latent infection. Journal of Virological Methods. 2000;89:61–73. doi: 10.1016/s0166-0934(00)00202-0. [DOI] [PubMed] [Google Scholar]
- Henson D, Smith RD, Gehrke J. Non-fatal mouse cytomegalovirus hepatitis. Combined morphologic, virologic and immunologic observations. American Journal of Pathology. 1966;49:871–88. [PMC free article] [PubMed] [Google Scholar]
- Holtappels R, Grzimek NKA, Simon CO, Thomas D, Dreis D, Reddehase MJ. Processing and Presentation of Murine Cytomegalovirus pORFm164-Derived Peptide in Fibroblasts in the Face of All Viral Immunosubversive Early Gene Functions. J Virol. 2002;76:6044–6053. doi: 10.1128/JVI.76.12.6044-6053.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaber S, Chanques G, Borry J, Souche B, Verdier R, Perrigault PF, Eledjam JJ. Cytomegalovirus Infection in Critically Ill Patients: Associated Factors and Consequences. 10.1378/chest.127.1.233 Chest. 2005;127:233–241. doi: 10.1378/chest.127.1.233. [DOI] [PubMed] [Google Scholar]
- Jonjic S, Pavic I, Polic B, Crnkovic I, Lucin P, Koszinowski UH. Antibodies are not essential for the resolution of primary cytomegalovirus infection but limit dissemination of recurrent virus. J Exp Med. 1994;179:1713–1717. doi: 10.1084/jem.179.5.1713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karupiah G, Sacks TE, Klinman DM, Fredrickson TN, Hartley JW, Chen JH, Morse HC., 3rd Murine cytomegalovirus infection-induced polyclonal B cell activation is independent of CD4+ T cells and CD40. Virology. 1998;240:12–26. doi: 10.1006/viro.1997.8900. [DOI] [PubMed] [Google Scholar]
- Kettering JD, Schmidt NJ, Gallo D, Lennette EH. Anti-complement immunofluorescence test for antibodies to human cytomegalovirus. J Clin Microbiol. 1977;6:627–632. doi: 10.1128/jcm.6.6.627-632.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurz S, Steffens HP, Mayer A, Harris JR, Reddehase MJ. Latency versus persistence or intermittent recurrences: evidence for a latent state of murine cytomegalovirus in the lungs. Journal of Virology. 1997;71:2980–7. doi: 10.1128/jvi.71.4.2980-2987.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurz SK, Rapp M, Steffens HP, Grzimek NK, Schmalz S, Reddehase MJ. Focal transcriptional activity of murine cytomegalovirus during latency in the lungs. Journal of Virology. 1999;73:482–94. doi: 10.1128/jvi.73.1.482-494.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawson CM, Grundy JE, Shellam GR. Antibody responses to murine cytomegalovirus in genetically resistant and susceptible strains of mice. Journal of General Virology. 1988;69:1987–98. doi: 10.1099/0022-1317-69-8-1987. [DOI] [PubMed] [Google Scholar]
- Loh LC, Balachandran N, Britt WJ. Characterization of a membrane-associated phosphoprotein of murine cytomegalovirus (pp50) and its immunological cross-reactivity with a human cytomegalovirus protein. Virology. 1991;183:181–194. doi: 10.1016/0042-6822(91)90131-t. [DOI] [PubMed] [Google Scholar]
- Lussier G, Guenette D, Descoteaux JP. Comparison of serological tests for the detection of antibody to natural and experimental murine cytomegalovirus. Canadian Journal of Veterinary Research. 1987;51:249–52. [PMC free article] [PubMed] [Google Scholar]
- Papazian L, Fraisse A, Garbe L, Zandotti C, Thomas P, Saux P, Pierrin G, Gouin F. Cytomegalovirus. An unexpected cause of ventilator-associated pneumonia. Anesthesiology. 1996;84:280–7. doi: 10.1097/00000542-199602000-00005. [DOI] [PubMed] [Google Scholar]
- Pelletier RP, Hennessy PK, Adams PW, VanBuskirk AM, Ferguson RM, Orosz CG. Clinical significance of MHC-reactive alloantibodies that develop after kidney or kidney-pancreas transplantation. American Journal of Transplantation. 2002;2:134–41. doi: 10.1034/j.1600-6143.2002.020204.x. [DOI] [PubMed] [Google Scholar]
- Pereira L, Hoffman M, Gallo D, Cremer N. Monoclonal antibodies to human cytomegalovirus: three surface membrane proteins with unique immunological and electrophoretic properties specify cross-reactive determinants. Infect Immun. 1982;36:924–932. doi: 10.1128/iai.36.3.924-932.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez M, Sabastian P, Clark P, Brown MG. Cmv1-Independent Antiviral Role of NK Cells Revealed in Murine Cytomegalovirus-Infected New Zealand White Mice. J Immunol. 2004;173:6312–6318. doi: 10.4049/jimmunol.173.10.6312. [DOI] [PubMed] [Google Scholar]
- Scalzo AA, Fitzgerald NA, Simmons A, La Vista AB, Shellam GR. Cmv-1, a genetic locus that controls murine cytomegalovirus replication in the spleen. Journal of Experimental Medicine. 1990;171:1469–83. doi: 10.1084/jem.171.5.1469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selgrade MK, Huang YS, Graham JA, Huang CH, Hu PC. Humoral antibody response to individual viral proteins after murine cytomegalovirus infection. Journal of Immunology. 1983;131:3032–5. [PubMed] [Google Scholar]
- Shanley JD, Jordan MC, Stevens JG. Modification by adoptive humoral immunity of murine cytomegalovirus infection. Journal of Infectious Diseases. 1981;143:231–7. doi: 10.1093/infdis/143.2.231. [DOI] [PubMed] [Google Scholar]
- Simmons RL, Matas AJ, Rattazzi LC, Balfour HH, Jr, Howard JR, Najarian JS. Clinical characteristics of the lethal cytomegalovirus infection following renal transplantation. Surgery. 1977;82:537–46. [PubMed] [Google Scholar]
- Smith HRC, Heusel JW, Mehta IK, Kim S, Dorner BG, Naidenko OV, Iizuka K, Furukawa H, Beckman DL, Pingel JT, Scalzo AA, Fremont DH, Yokoyama WM. Recognition of a virus-encoded ligand by a natural killer cell activation receptor. PNAS. 2002;99:8826–8831. doi: 10.1073/pnas.092258599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spector SA, Wong R, Hsia K, Pilcher M, Stempien MJ. Plasma Cytomegalovirus (CMV) DNA Load Predicts CMV Disease and Survival in AIDS Patients. J Clin Invest. 1998;101:497–502. doi: 10.1172/JCI1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stavnezer J. Antibody class switching. Advances in Immunology. 1996;61:79–146. doi: 10.1016/s0065-2776(08)60866-4. [DOI] [PubMed] [Google Scholar]
- Stinski MF, Mocarski ES, Thomsen DR, Urbanowski ML. Membrane glycoproteins and antigens induced by human cytomegalovirus. Journal of General Virology. 1979;43:119–29. doi: 10.1099/0022-1317-43-1-119. [DOI] [PubMed] [Google Scholar]
- Sullivan-Tailyour G, Garnett HM. Plasma membrane proteins and glycoproteins induced by human cytomegalovirus infection of human embryonic fibroblasts. Journal of General Virology. 1986;67:515–26. doi: 10.1099/0022-1317-67-3-515. [DOI] [PubMed] [Google Scholar]
- Thale R, Lucin P, Schneider K, Eggers M, Koszinowski UH. Identification and expression of a murine cytomegalovirus early gene coding for an Fc receptor. J Virol. 1994;68:7757–7765. doi: 10.1128/jvi.68.12.7757-7765.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tripathy SK, Smith HRC, Holroyd EA, Pingel JT, Yokoyama WM. Expression of m157, a Murine Cytomegalovirus-Encoded Putative Major Histocompatibility Class I (MHC-I)-Like Protein, Is Independent of Viral Regulation of Host MHC-I. J Virol. 2006;80:545–550. doi: 10.1128/JVI.80.1.545-550.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- VanBuskirk AM, Wakely ME, Sirak JH, Orosz CG. Patterns of allosensitization in allograft recipients: long-term cardiac allograft acceptance is associated with active alloantibody production in conjunction with active inhibition of alloreactive delayed-type hypersensitivity. Transplantation. 1998;65:1115–23. doi: 10.1097/00007890-199804270-00017. [DOI] [PubMed] [Google Scholar]
- Voigt V, Forbes CA, Tonkin JN, Degli-Esposti MA, Smith HRC, Yokoyama WM, Scalzo AA. Murine cytomegalovirus m157 mutation and variation leads to immune evasion of natural killer cells. PNAS. 2003;100:13483–13488. doi: 10.1073/pnas.2233572100. [DOI] [PMC free article] [PubMed] [Google Scholar]