

Abstract
PURPOSE
Provide a reproducible method for culturing confluent monolayers of hfRPE cells that exhibit morphology, physiology, polarity, and protein expression patterns similar to native tissue.
METHODS
Human fetal eyes were dissected on arrival, and RPE cell sheets were mechanically separated from the choroid and cultured in a specifically designed medium comprised entirely of commercially available components. Physiology experiments were performed with previously described techniques. Standard techniques were used for immunohistochemistry, electron microscopy, and cytokine measurement by ELISA.
RESULTS
Confluent monolayers of RPE cell cultures exhibited epithelial morphology and heavy pigmentation, and electron microscopy showed extensive apical membrane microvilli. The junctional complexes were identified with immunofluorescence labeling of various tight junction proteins. The mean transepithelial potential (TEP) was 2.6 ± 0.8 mV, apical positive, and the mean transepithelial resistance (RT) was 501 ± 138 Ω· cm2 (mean ± SD; n = 35). Addition of 100 μM adenosine triphosphate (ATP) to the apical bath increased net fluid absorption from 13.6 ± 2.6 to 18.8 ± 4.6 μL · cm−2 per hour (mean ± SD; n = 4). In other experiments, VEGF was mainly secreted into the basal bath (n = 10), whereas PEDF was mainly secreted into the apical bath (n = 10).
CONCLUSIONS
A new cell culture procedure has been developed that produces confluent primary hfRPE cultures with morphological and physiological characteristics of the native tissue. Epithelial polarity and function of these easily reproducible primary cultures closely resemble previously studied native human fetal and bovine RPE–choroid explants.
Multiple physiological functions of the RPE serve to protect the health and integrity of the distal retina and the extracellular space outside Bruch's membrane. Disease processes, such as age-related macular degeneration (AMD), that develop within the RPE–distal retinal complex can lead to significant vision loss.1 2 3 4 5 6 7 8 These functions determine the RPE's ability to control and respond to varying levels of light, vitamin A uptake and delivery, outer segment phagocytosis, oxygen consumption, nutrition and waste product exchange between the subretinal space (SRS), and choroidal blood supply.
Cell culture models can play an important role in the study of the physiology and pathophysiology of native tissue.9 10 11 A strength of this approach is the ability to isolate specific epithelial functions and the set of involved proteins. This ability is particularly important for work in human cell culture, for which the access to fresh native tissue is limited. In some cases, it is possible to compare the results obtained using cell culture to similar experiments using native tissue in vitro.12 A weakness of this system is that the full spectrum of signals from retina and choroid are not available for interactions with the RPE. However, it is possible in this system to mimic some of the interactions that occur in vivo.13 14 15 16
Previously, we developed a preparation of native human fetal RPE and identified some of the plasma membrane and intracellular mechanisms that regulate intracellular pH, [Ca2+], and transepithelial ion and solute-linked fluid transport.12 17 18 19 20 21 22 Because of the limited availability of these tissues, we have developed a set of standard procedures for culturing confluent monolayers of human fetal RPE cells. One goal was to find a set of well- defined commercially available components that could be used to produce stable and reproducible RPE cultures. Our initial requirements were that the cell cultures form single monolayers that resemble native tissue as closely as possible in morphology (uniform hexagonal arrays of cells, similar in size and shape), confluence, uniform pigmentation, transepithelial potential (TEP), and transepithelial resistance (RT >100 Ω· cm2). The minimum electrical resistance requirements have been determined in experiments on native bovine and human RPE.19 21 23 24
In the present experiments we further characterized these cultures using electron microscopy (EM) and immunohistochemistry to identify cell structures and localize apical and basolateral membrane and intercellular junctional complex proteins. ELISAs were used to measure cytokine levels and confirm the polarity of secretion of selected cytokines. Intracellular microelectrodes were used to characterize receptor-mediated second-messenger pathways and their downstream electrophysiological properties at the apical and basolateral membranes. The capacitance probe technique was used to measure net transepithelial fluid transport. This model system has been used to help identify some of the mitochondrial pathways that mediate oxidative stress and the protective effects of lipoic acid.22
METHODS
Human Fetal Tissue
The research followed the tenets of the Declaration of Helsinki and the NIH institutional review board. Fetal eyes were obtained by an independent procurer, Advanced Bioscience Resources (Alameda, CA), from random donors at 16 to 18 weeks of gestation, placed in tissue-delivery medium–containing tubes and packed on ice, and delivered by an overnight priority delivery service. All tissues were used less than 26 hours after enucleation.
Solutions for Physiology Experiments and Cell Culture Medium Preparation
The human fetal eye transportation medium (Table 1) is prepared using 5 mM KCl, 0.8 mM MgCl2, 113.4 mM NaCl, 26.3 mM NaHCO3, 1 mM NaH2PO4, 3 mM Na2HPO4, 5 mM taurine, 5.6 mM glucose, 1.8 mM CaCl2 after adjusting pH to 7.4 by using CO2. Additional components include 20 mL/L MEM amino acid 50x (Sigma-Aldrich, St. Louis, MO), 10 mL/L MEM vitamin solution 100x (Sigma-Aldrich), 0.11g/L pyruvic acid sodium (cell-culture tested; Sigma-Aldrich), 10 mL/L penicillin-streptomycin 100x (Invitrogen, Carlsbad, CA), 10 mL/L glutamax-I, 200 nM-100x (Invitrogen). Final transportation medium was filtered, aliquoted into 15 mL centrifuge tubes, and stored in 4°C up to 2 months.
Table 1.
Human Fetal Eye Transportation Medium Components
| Name | Sigma-Aldrich | Invitrogen | Concentration | Storage |
|---|---|---|---|---|
| KCl | P-5405 | 5 mM | Room | |
| MgCl2 | M-8266 | 0.8 mM | Desiccator | |
| NaCl | S-5886 | 113.4 mM | Room | |
| NaHCO3 | S-6297 | 26.3 mM | Room | |
| NaH2PO4 | S-5011 | 1 mM | Room | |
| Na2HPO4 | S-5136 | 3 mM | Room | |
| Taurine | T-0625 | 5 mM | Room | |
| Glucose | G-6152 | 5.6 mM | Room | |
| CaCl2 | 383147 | 1.8 mM | Desiccator | |
| MEM amino acids | M-5550 | 20 mL/L | +4°C | |
| MEM vitamin solution | M-6895 | 10 mL/L | −20°C | |
| Pyruvic acid sodium | P-4562 | 0.11 g/L | +4°C | |
| Penicillin-streptomycin | 15140-122 | 10 mL/L | −20°C | |
| GlutaMAX I | 35050-061 | 10 mL/L | −20°C |
MEM-α modified medium (Sigma-Aldrich) was used as the base medium to prepare 5% and 15% serum-containing media for culturing RPE cells (RPE medium; Table 2). Fetal bovine serum was used in media preparation (Atlanta Biologicals, Norcross, GA), screened for toxicity, and heat inactivated. Cell culture medium also contained: N1 supplement (Sigma-Aldrich) 1:100 mL/mL, glutamine-penicillin-streptomycin (Sigma-Aldrich) 1:100 mL/mL, and nonessential amino acid solution (Sigma-Aldrich) 1:100 mL/mL. In addition, hydrocortisone (20 μg/L), taurine (250 mg/L), and triiodo-thyronin (0.013 μg/L) (THT) were dissolved in PBS to a final concentration of 1:500 (mL/mL) and aliquots stored at −80°C until added to the RPE medium. The composition of this medium was arrived at by using a variety of basic media (e.g., DMEM 12, DMEM, MEM) which were evaluated for their ability to promote hexagonal RPE morphology and pigment density and to generate epithelial electrical potential and resistance (EVOM) consistent with that of native tissue. In addition to this basic medium, we included several components and combinations of components from previous studies of human RPE.19 25 Other components (e.g., taurine, serum, combinations of antibiotics) and growth factors (e.g., bFGF, epidermal growth factor [EGF], brain-derived neurotrophic factor [BDNF], and ciliary neurotrophic factor [CNTF]) were also tested for their ability to maintain longterm viability in in vitro physiology experiments.26 These media were also tested with a variety of substrate coatings such as collagen, laminin, fibronectin, gelatin, and extracellular matrix. The substrate that produced the best morphology and physiology was human extracellular matrix from human placenta (followed by fibronectin). Ringer solutions for the physiology experiments contained the following (in mM): 120 NaCl, 5 KCl, 23 NaHCO3, 1 MgCl2, 1.8 CaCl2, 2.0 taurine, 1 glutathione, and 10 glucose. This solution was bubbled continuously with 5% CO2-10% O2-85% N2, to a stable pH of ∼7.4 and an osmolarity of 295 ± 5 mOsM.
Table 2.
Human Fetal RPE Medium Components for Preparation of ∼500 mL Medium
| Name | Sigma | Amount | Storage |
|---|---|---|---|
| MEM, αmodification | M-4526 | 500 mL | +4°C |
| N1 supplement | N-6530 | 5 mL | +4°C |
| Glutamine-penicillin-streptomycin | G-1146 | 5 mL | −20°C |
| Non essential amino acids | M-7145 | 5 mL | +4°C |
| THT* | −80°C | ||
| Taurine | T-0625 | 125 mg | |
| Hydrocortisone | H-0396 | 10 μg | |
| Triiodo-thyronin | T-5516 | 0.0065 μg | |
| Fetal bovine serum, heat inactivated*,† | 5% or 15% | −80°C |
THT is made by dissolving taurine-hydrocortisone-triiodo-thyronin in 1–1.5 mL PBS before making the medium. Multiple aliquots are made and stored at −80°C to simplify culturing preparation of the culture medium.
Fetal bovine serum is not obtained from Sigma-Aldrich.
Cell Culture
On receipt, intact globes were rinsed in antibiotic–antimycotic solution (diluted to 10x; cat. no. 15240-096; Invitrogen) plus gentamicin (1 mg/mL) for 3 to 5 minutes (Fig. 1). Antibiotics were rinsed off twice with medium or PBS. After the anterior portion of the eye was removed, the posterior poles were incubated in dispase-I solution (2 U/mL, cat. no. 1284908; Roche Diagnostics, Indianapolis, IN) in 5% serum containing medium for 30 minutes in 37°C-5% CO2. After dispase treatment, the posterior poles were transferred to a 5% serum-containing RPE medium in Petri dishes with silicon padding (Sylgard 184; Dow Corning, Midland, MI) and dissected into quadrants, and the retina was gently removed with forceps. Single-cell RPE layers were peeled off in small sheets and collected in 5% serum-containing RPE medium. The cells were then washed (centrifugation at 100g), separated from their syncytium by gently pipetting them, with or without trypsin treatment, and then put into Primaria flasks (example: cat. no. 08-772-45; Fisher Scientific, Pittsburgh, PA) with a 15% serum-containing RPE medium. This medium was replaced after 1 day with 5% serum-containing RPE medium, and subsequent changes were made every 3 to 4 days. After 3 to 4 weeks, the cells became confluent and uniformly pigmented. They were then trypsinized in 0.25% trypsin for 10 to 15 minutes, resuspended in 15% serum-containing RPE cell culture medium, and seeded onto clear cell culture inserts at 200 × 103 per well (Transwell; Corning Costar, Corning, NY), using 12-mm diameter inserts, 0.4-μm pores, polyester membranes (example: cat. no. 07-200-161, Fisher Scientific). Before seeding, the wells were coated with human extracellular matrix (10 μg in 150 μL HBSS per well, cat. no. 354237; BD Biosciences, Franklin Lakes, NJ) and cured with UV light in the hood for 2 hours. In some cases, the trypsinization procedure was repeated for a second time, to collect the cells that did not detach after the first trypsinization.
FIGURE 1.

The main steps in the preparation of hfRPE cell cultures.
The same protocol excluding coating with ECM was used to culture cells on the flasks. Cells were used in experiments when they had a total tissue resistance of >200 Ω· cm2 and were uniformly pigmented (Fig. 2).
FIGURE 2.

Microphotograph of cultured RPE cells grown on inserts for 4 weeks, taken with an inverted microscope. As in native tissues, these cells formed a single monolayer, were well pigmented, and were arranged in a regular hexagonal mosaic. Magnification, x20.
Electrophysiology
Calomel electrodes in series with Ringer solutions and agar bridges were used to measure the TEP, and an intracellular microelectrode, referenced to either the apical (A) or basal (B) bath, was used to measure the membrane potentials, VA and VB, where TEP = VB – VA. As previously described,27 conventional microelectrodes were made from borosilicate glass tubing of 0.5 mm inner diameter and 1 mm outer diameter, with a filament (Sutter Instrument Co., Novato, CA) and were back-filled with 150 mM KCl, and had resistances of 80 to 200 MΩ.
The total RT and the ratio of the apical-to-basolateral membrane resistance (RA/RB) were obtained by passing 2- to 4-μA current pulses across the tissue and measuring the resultant changes in TEP, VA, and VB. RT is the resultant change in TEP divided by the current amplitude, and the RA/RB ratio is the absolute value of the change in VA divided by the change in VB (RA/RB = iΔVA/iΔVB). The current-induced voltage deflections were digitally subtracted from the records for clarity. In these experiments, the black bar indicates a solution change (e.g., see Fig. 7) in the manifold outside the recording chamber. In some cases, the response onset was variably delayed because of “dead space” in the fluid delivery system and because of thickness variations in the unstirred layer at the cell membrane.
FIGURE 7.

The effect of changing apical bath potassium on TEP, RT, VA, VB, and RA/RB. Reduction of apical [K]o from 5 to 2 mM (solid bar) increased TEP (top) by ≈1.8 mV and RT by ≈18 Ω· cm2 (solid line and open diamonds, respectively). Concomitantly, both membranes rapidly hyperpolarized (bottom, solid lines), and RA/RB increased by threefold to 0.3 (bottom, open triangles). Representative of nine similar recordings.
Fluid Transport
Transepithelial water flow measurements were made with a refined capacitance probe technique in a modified Üssing chamber apparatus (Tripathi S, et al. IOVS 2004;45:ARVO E-Abstract 1092).27 28 A capacitance probe (MTI Instruments Inc., Albany, NY) tracked changes in aqueous level as water moved across the RPE by measuring changes in the dielectric thickness between the aqueous phase and the probe. Viability of the tissue was ascertained by recording TEP and RT measured by injecting known bipolar currents via Ag/AgCl pellet electrodes. The current method for measuring transepithelial water flow (Jv) of epithelia, used in the current study, combines and improves on several features of earlier studies (Tripathi S, et al. IOVS 2004;45:ARVO E-Abstract 1092).27 28 29
Electron Microscopy
Tissues were fixed in 4% glutaraldehyde-buffered solution at room temperature for 2 hours and rinsed three times for 5 minutes in PBS. These tissues were then treated with 1% ice-cold osmium tetroxide in PBS solution for 1 hour. After osmication, tissues were rinsed in PBS and processed through a battery of ethanol dehydration steps (three washes in 50%, 70%, 85%, and 100% ethanol, respectively) for plastic embedding. A routine transmission electron microscopy protocol was used to obtain the RPE micrographs.30 31
Immunohistochemistry
Immunofluorescence
Primary antibodies were labeled with a commercial antibody labeling technology (Zenon; Invitrogen Corp.), according to the manufacturer's instructions. Cultures growing on cell culture inserts (Transwell; Corning Costar) were placed on ice and washed three times with cold PBS. The cells were fixed for 30 minutes in 4% formaldehyde-PBS, washed three times with PBS, and permeabilized for 30 minutes with 0.2% Triton X-100-PBS. The cells were washed three times with PBS, blocked with signal enhancer (Image-IT FX; Invitrogen) for 30 minutes at room temperature, washed three times with PBS, stained with labeled primary antibody at a 1:100 dilution in PBS, washed three times with PBS, and treated with 4% formaldehyde-PBS for an additional 15 minutes at room temperature. The cells were then mounted on glass slides with antifade reagent with 4′,6′-diamino-2-phenylindole (DAPI; Prolong Gold; Invitrogen) and imaged with a microscope (Axioplan 2 with Axiovision 3.4 software with ApoTome; Carl Zeiss Meditec, Inc., Dublin, CA).
Western Blot Analysis
Four-month-old cultures were rinsed with cold PBS, harvested with a cell scraper, and pelleted at 4°C. The pellet was lysed with RIPA buffer (Sigma-Aldrich, St. Louis, MO) supplemented with complete protease inhibitors (Roche) for 20 minutes on ice with periodic vortexing. Cell lysate was centrifuged at 14,000g for 10 minutes, and the supernatant was collected. Extracted protein was quantitated with a BCA protein assay (Pierce Chemical, Rockford, IL). Twenty micrograms of total protein was loaded into each lane of a 4% to 12% polyacrylamide gradient gel (NuPAGE gel; Invitrogen) under reducing conditions and transferred to nitrocellulose membranes. Membranes were probed with antibodies and developed with enhanced chemiluminescence (Supersignal Pico ECL; Pierce Chemical, Rockford, IL) and imaged with a gel documentation system (Autochemie; UVP, Upland, CA).
ELISAs for Human VEGF, PEDF
VEGF and PEDF levels were measured with samples collected from the apical and basal sides of the same insert (n = 9). For VEGF, all steps were performed at room temperature, and each step was followed by three washes with 400 μL/well of washing buffer (0.05% Tween-20 in PBS, pH 7.4). A 96-well EIA plate (Corning Costar) was coated with 100 μL/well of 0.4 μg/mL polyclonal goat anti-human VEGF antibody (R&D Systems, Minneapolis, MN) overnight in PBS (pH 7.4). The plate was blocked by adding 300 μL/well of PBS containing 1% BSA, 5% sucrose, and 0.05% NaN3 for 1 hour. A set of seven-point VEGF standards (10–1000 pg/well; R&D Systems) and samples (cell culture medium) in 100 μL diluent (20 nM Tris-HCl, 150 mM NaCl, 0.1% BSA, and 0.05% Tween-20, pH 7.3) were added in triplicate and incubated for 2 hours. The plate was then incubated with 100 μL/well of 0.2 μg/mL biotinylated goat anti-human VEGF antibody (R&D Systems) in diluent for 2 hours. Streptavidin-alkaline phosphatase (100 μL/well, 1:2000 in diluent; Life Technologies, Gaithersburg, MD) was added to the plate for 30 minutes, followed by its substrate, p-nitrophenyl phosphate (pNPP; 1 mg/mL in 0.2 M Tris buffer, pH 9.8). Optical densities were obtained within 1 to 4 hours with a microplate reader at 405 nm with wavelength correction at 570 nm, and data were analyzed on computer (Excel; Microsoft, Redmond, WA).
The PEDF ELISA was performed according to kit instructions (Chemicon, Temecula, CA). Samples were treated with 8 M urea for 1 hour on ice, loaded on a precoated plate after addition of 1:100 diluent, and incubated for 1 hour. Biotinylated mouse anti-human PEDF was added for 1 hour. Detection with Streptavidin-horseradish peroxidase and substrates was similar to the VEGF ELISA described earlier.
RESULTS
Figure 3 illustrates the growth of a typical cell culture (P0) seeded on a flask (Primaria; Corning Costar). The first few attached cells were seen 4 hours after seeding and most live cells were attached in the first 24 hours. Figure 3A shows that the initial growth of the culture occurred in the form of small islands, consisting of 10 to 20 RPE cells. Single cells were also present (not shown), but they grew at a much slower rate than the RPE islands. Pigment density was observed to decrease over the first few divisions, suggesting that the cells share the original complement of pigment from the native tissue. As the cells continued to divide, pigment density increases, indicating the de nova synthesis of pigment (Fig. 3B). The newly dividing cells retained their epithelioid morphology. As the cell culture matured, cell shape and pigment density became more uniform (Fig. 3D). After 30 days in culture, pigment density was comparable to the native tissue.
FIGURE 3.

Light microscope images of RPE cultured in a flask at various times: (A) 1, (B) 5, (C) 12, and (D) 14 days.
The morphology of the cells was evaluated using electron microscopy (EM) of plastic embedded hfRPE cultures. Fig. 4A shows an EM micrograph of RPE cells cultured on a plastic semiporous membrane. The cells grew in a single monolayer with well-developed apical membrane processes and junctional complexes between the cells (Fig. 4D). As in native tissue they had basally located nuclei and contained pigment granules that congregated close to the apical membrane. Figures 4B and 4C show the basal side of RPE cells (x50,000) with clearly visible basal infolding. In some cells (Fig. 4C), we observed structures close to the basal membrane that can be designated basilar deposits (term suggested by reviewer). These resemble other types of sub-RPE basal deposits that consist of diffuse heterogeneous material mainly formed from periodically spaced collagen fibers in adult eyes.32 33 34 In hfRPE cultures, these deposits were observed in 2-and 4- month-old, but not in 1-month-old, monolayers. There were one to five such deposits per cell, and each deposit contained three to five bands with 50 to 60 nm periodicity.
FIGURE 4.
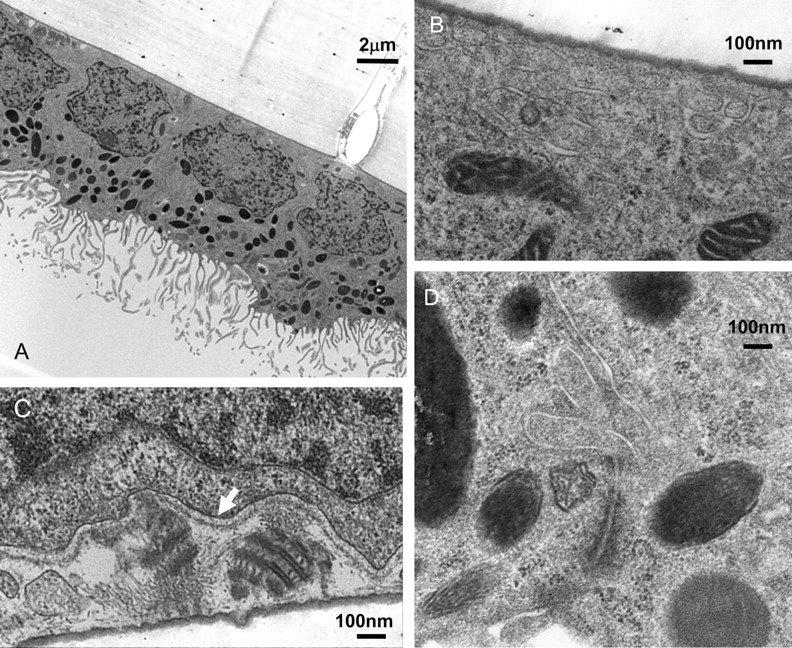
(A) Electron micrograph of RPE cells on cell culture inserts. Each cell exhibited large apical processes, pigmentation was primarily located on the apical side, and nuclei were located close to the basal membrane. Cells appear to be growing in a single monolayer with well-pronounced tight-junctional complexes. (B, C) Basal side of RPE cell image. (C) Many RPE cells contain laminar-deposit–like structures apparently secreted through the RPE basolateral membrane (arrow). (D) Tight-junctional complex. Magnification: (A) x5,000; (B–D) x50,000.
Immunohistochemistry and Western Blot Analysis
RPE cell cultures were tested for various tight junction, visual cycle, membrane, and cytoskeletal markers, such as occludin (Fig. 5A), claudin-19 and -10 (Figs. 5B 5C), CRALBP (Fig. 5D), RPE65 (Fig. 5E) , cytokeratin (Fig. 5F), bestrophin (Fig. 5G), NaK-ATPase (Fig. 5H), and ezrin (Fig. 5I).
FIGURE 5.
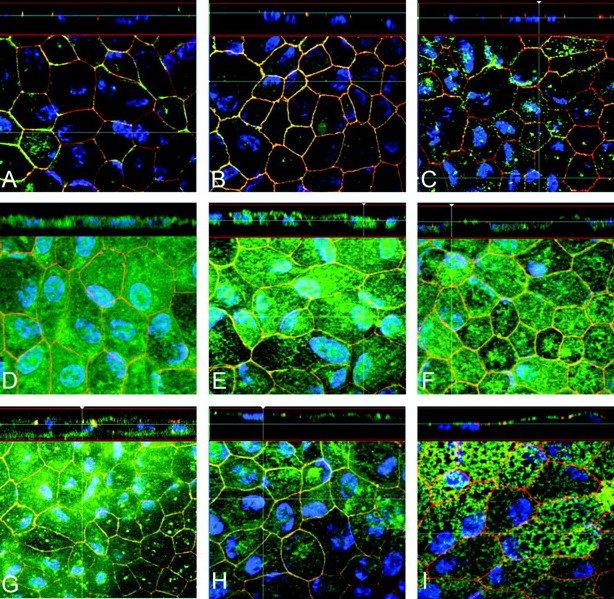
Representative immunohistochemistry staining of hfRPE cultures grown on cell culture inserts. Bottom of each panel is an en face view of a cell culture monolayer shown in a maximum-intensity projection through the z-axis. Top of each panel is a cross section through the z-plane of multiple optical slices. Blue: DAPI-stained nuclei; red: ZO1-stained nuclei. Green: occludin (A), claudin-19 (B), claudin-10 (C), CRALBP (D), RPE65 (E), cytokeratin (F), bestrophin (G), NaK-ATPase (H), and ezrin (I).
Occludin and ZO-1 are two examples of epithelial integral tight junction proteins that are part of the junctional complex. ZO-1 expression occurs first, during the early stages of confluence, upon cell–cell contact and can be used to help localize the tight junctions. Claudin (and occludin) appears later and is more closely related to epithelial barrier function and polarity. The molecular composition of the tight junctions was clarified by the identification of occludin by Furuse et al.35 and claudin-1 and -2.36 Recent microarray data in hfRPE indicate that claudin-19 and -10 are most abundant in the hfRPE preparation (Wang F, et al. IOVS 2006;47:ARVO E-Abstract 2855).
Ezrin is a cytoskeleton protein that has been identified exclusively in RPE cell apical processes.37 38 Apical localization of ezrin in hfRPE cultures has helped to confirm RPE polarization and the presence of apical processes, also seen in EM (Fig. 4A). CRALBP (Fig. 5D) and RPE65 (Fig. 5E) , two proteins critical for the visual cycle, were also identified.
The membrane localizations of NaK-ATPase (apical) and bestrophin (basolateral) indicate normal RPE polarity. The functional localization of the NaK-ATPase to the apical membrane has been demonstrated13 19 23 39 40 and corroborated by immunohistochemistry on native and cultured RPE tissues.41 42 43 44 Immunohistochemistry results were confirmed by Western blot. Figure 6 shows representative Western blot analysis identifying the six proteins (RPE65, NaK-ATPase, ezrin, cytokeratin, rbCRALB, and bestrophin).
FIGURE 6.

Western blot identifying the six proteins in native hfRPE cultured cells. Cells were cultured in Primaria flasks.
Electrophysiology
We also quantified several key functional properties of these hfRPE cell cultures. Confluent monolayers on cell migration filters were mounted in modified Üssing chambers and used to analyze single-cell and epithelial physiological responses using standard microelectrode intracellular recording techniques, as described in the Methods section. In 35 experiments, the TEP was 2.6 ± 0.8 mV (mean ± SD) and the total monolayer resistance (RT) was 501 ± 138 Ω· cm2. In 12 intracellular recording experiments, the apical membrane potential (VA) was −52.3 ± 5.4 mV and the basolateral membrane potential (VB) was −49.8 ± 3.7, whereas the ratio of apical-to-basolateral membrane resistance (RA/RB) was 0.25 ± 0.2.
The experiments summarized in Figure 7 illustrate typical electrophysiological responses to lowering potassium concentration ([K+]o) on the apical side from 5 to 2 mM (solid bar). This [K+]o decrease mimics what occurs in vivo in the subretinal space after the transition from dark to light.45 In nine experiments, this apical [K+]o decrease (Fig. 7, bottom) hyperpolarized the apical membrane by 10.7 ± 2.5 mV and the basolateral membrane by 9.2 ± 3.2 mV (mean ± SD). As illustrated in Figure 7 , top, the mean TEP transiently increased and then returned toward baseline. In nine cultures, the mean increase was 1.8 ± 0.7 mV. In 2 mM [K+]o, both RT (top) and RA/RB (bottom) increased. The mean increase was 18.3 ± 9.1 Ω· cm2 and 0.28 ± 0.07, respectively, consistent with an increase in apical membrane resistance probably mediated by the apical membrane K channels.
This conclusion was corroborated by the data illustrated in Figure 8 which shows that apical Ba2+, a specific K channel blocker, depolarized VA by ≈10 mV and decreased TEP by ≈1.5 mV. Concomitantly, RT and RA/RB were increased by ≈20 Ω· cm2 and 0.2, respectively. All these changes are consistent with the closure of apical membrane K channels. Lowering apical [K+]o in the presence of Ba2+ failed to produce further membrane voltage or resistance changes (Fig. 7), as previously observed in native tissue.19 46 47
FIGURE 8.

The effect of changing apical [K]o from 5 to 2 mM (solid bar) on TEP, VA, VB, and RA/RB in the presence of 1 nM barium in the apical bath (hatched bar). Addition of apical Ba2+ depolarized both membranes (bottom and top traces denoted as in Fig. 7), decreased TEP and increased RT. RA/RB increased twofold, from 0.2 to 0.4. Apical Ba2+ blocked all the [K]o-induced electrical changes. Representative of four similar recordings.
Figure 9 illustrates typical hfRPE electrical responses to a [K+]o increase in the basal bath of these cultured RPE cells. In this experiment, we were able to obtain intracellular recordings along with the changes in TEP and RT. Increasing basal [K+]o from 5 to 50 mM depolarized VB by ≈21 mV (bottom) as reflected by the large increase in TEP (4 mV; top). At the same time, the RA/RB ratio increased by ≈0.5, and RT initially decreased by ≈20 Ω· cm2 and then increased, probably due to [K+]o-induced cell swelling, as observed in native bovine RPE.48 The initial changes in resistance were probably caused by a [K+]o-induced conductance increase in basolateral membrane K channels. In four experiments, this 10-fold increase in basal [K+]o increased TEP and RT by 3.2 ± 0.9 mV and 33.7 ± 9.5 Ω· cm2, respectively.
FIGURE 9.

The effect of changing basal bath [K]o. Bottom and top traces defined as in Figure 7. Increasing basal [K]o from 5 to 50 mM (solid bar) depolarized VA and VB, increased TEP (≈3 mV). RT transiently decreased and then increased (≈19 Ω· cm2). RA/RB increased twofold, from 0.2 to 0.9. Representative of three similar recordings.
In native tissues, it has been shown that the RPE apical membrane contains α1-adrenergic and P2Y2 purinoceptors that can be activated by epinephrine in the nanomolar range and ATP/UTP in the micromolar range. This activation has been shown to increase basolateral membrane Cl conductance and to increase cell Ca2+ and net fluid transport across the RPE (Figs. 10 11 12).21 27 28 49 50 Figure 10 shows the epinephrine-induced intracellular responses obtained from a monolayer of cultured hfRPE cells. Apical epinephrine depolarized VB by ≈8 mV (bottom) and increased TEP by ≈0.4 mV (top); therefore the depolarization was slightly greater at the basolateral membrane. At the same time, RT decreased and the RA/RB ratio increased by ≈0.15, consistent with an increase in basolateral membrane conductance. All these electrical changes were also observed in native RPE tissue (bovine and human). In four experiments, epinephrine depolarized VB by 13.3 ± 8.9 mV and increased TEP by 0.7 ± 0.7 mV. At the same time, total tissue resistance RT decreased by 25.3 ± 10.9 Ω· cm2 and the RA/RB ratio increased by 0.16 ± 0.03.
FIGURE 10.

Apical epinephrine-induced electrical responses. Bottom and top traces defined as in Figure 7. Epinephrine (10 nM) was perfused into the apical bath (solid bar) and depolarized VA and VB and increased TEP. RA/RB almost doubled from 0.2 to 0.35 and RT decreased by ≈ 10 Ω· cm2. This is representative of four similar recordings.
FIGURE 11.

Apical ATP-induced changes in hfRPE membrane voltage and resistance. Bottom and top traces defined as in Figure 7. Addition of ATP (100 μM) to the apical bath (solid bar) caused a rapid depolarization of both membranes and TEP initially decreased. RT decreased and RA/RB increased twofold to 0.4. Representative of 10 similar recordings.
FIGURE 12.

ATP-induced changes in TEP, RT and fluid transport. Top: JV is plotted as a function of time and net fluid absorption indicated by positive values. No JV measurements were made during the solution composition changes (period marked by “probe out”). Bottom: TEP and RT plotted as a function of time in control Ringer and in the presence of 100 μM ATP. ATP increased JV by 6 μL · cm−2 per hour, decreased RT by ≈30 Ω· cm2, and transiently decreased TEP by ≈3 mV.
In Figure 11, before the addition of ATP to the apical bath, TEP was 1 mV, RT was ≈500 Ω· cm2, VA ≈−56 mV, and RA/RB = 0.1. Addition of ATP to the apical bath caused a rapid depolarization of both membranes, and initially the apical membrane depolarized at a faster rate than the basolateral membrane, since the TEP decreased. The TEP then reversed and began to increase concomitant with the continued depolarization of VB.VA also depolarized, but at a slower rate. RT decreased by ≈40 Ω· cm2, and RA/RB increased approximately fourfold to 0.4, indicating an increase in basolateral membrane conductance, as seen in native RPE. In three experiments with microelectrodes, VA depolarized by 15 ± 3.1 mV and RA/RB increased by 0.24 ± 0.6. In 11 experiments, measuring only the changes in TEP and RT, the initial transient decrease in TEP was 1.6 ± 1.1 mV, and the simultaneous decrease in RT was 38.7 ± 14.4 Ω· cm2 (mean ± SD).
In this preparation, the TEP response did not include the initial rapid increase as observed in bovine and human native tissues after P2Y2 receptor activation. The absence of this transience may be due to a difference in the timing of ATP-induced apical or basolateral membrane voltage changes.27 50
The data summarized in Figure 12 show that activation of the apical membrane P2Y2 receptors with ATP reversibly increased net fluid transport across hfRPE in the retina to choroid direction (absorption). ATP (100 μM) increased JV by ≈8 μL · cm−2 per hour and deceased RT by ≈40 Ω· cm2. At the same time, the TEP transiently decreased by 2.5 mV and then rebounded to a new steady state below the original baseline. In four experiments, apical ATP increased net JV from13.6 ± 2.6 to 18.8 ± 4.6 μL · cm−2 per hour (mean ± SD).
Mean TEP decreased by 1.3 ± 0.9 mV (initial transient only) and RT decreased by 35.6 ± 14 Ω· cm2. These electrical changes are consistent with the ATP-induced increase in basolateral membrane Cl channel conductance observed in native bovine RPE.27
It is thought that normal fenestration of the choroidal blood supply is maintained in part by a balanced secretion of pro-(VEGF) and anti-(PEDF) angiogenic factors by RPE cells. Figure 13 summarizes the data showing that hfRPE cells grown on cell culture inserts (Transwell; Corning Costar) preferentially secreted VEGF to the choroidal side of the tissue and PEDF to the apical side. The medium from each well was used to quantify the amount of VEGF and PEDF secreted into the apical and basal baths, respectively. All samples were collected from the same plate. The mean rate of secretion of VEGF to the basal and apical baths was 14.7 ± 0.13 ng/well per 24 hours and 8.7 ± 0.17 ng/well per 24 hours, respectively (mean ± SEM; n = 9; P <0.005). In contrast, PEDF was secreted into the apical bath at a 70% greater rate than into the basal bath (661 ± 32 ng/well per 24 hours vs. 285 ± 10 ng/well per 24 hours; n = 9; P < 0.05). The magnitude of these PEDF secretion rates is more than 20 times greater than the VEGF rates. Based on these data, the ratio of these two angiogenic factors is [PEDF/VEGF]Ap = 76 and [PEDF/VEGF]Ba = 19.4, suggesting a possible contributing factor to the maintenance of fenestrated versus nonfenestrated blood vessels in the retina and choroid, respectively.
FIGURE 13.

Polarized secretion of VEGF and PEDF in cultured human fetal RPE. Media from apical and basal baths were collected separately at 8 weeks after the cells were seeded onto cell culture inserts. ELISA for VEGF and PEDF were performed in triplicate for each of the 12 wells (24 samples). VEGF protein in the basal bath was 1.7-fold higher than in the apical bath (n = 9; P < 0.005), whereas PEDF protein in the apical bath was 2.3-fold higher than in the basal bath (n = 9; P < 0.05).
DISCUSSION
In the past decade, there has been an increased interest in RPE function and its role in multiple retinal diseases.51 52 53 54 55 56 A corollary of this increased interest is the need to develop in vitro cell culture models to study a wide range of RPE physiology and pathophysiology. As in other organ systems, it has been difficult to create RPE cell lines and primary cultures with the appropriate morphologic and structural characteristics.57 58 59 60 In addition, there has been relatively little focus on the comparison of these various model systems with the physiology of native tissue.61 62 63 64 Such comparisons are a necessary requirement for a cell culture system that closely models the wide range of structural, functional, and regulatory characteristics of native mammalian or human RPE.
Native adult or fetal human RPE tissue is rarely used for in vitro RPE research because of its limited availability, its relatively long delivery time, and its limited viability once it arrives in the laboratory. In the present study, primary cell culture experiments were performed by producing a large number of physiologically similar cells from native human fetal tissue. The functional and anatomic similarity of these hfRPE cultured cell monolayers to native RPE tissue is based in part on a variety of previous physiological experiments using native human fetal, adult amphibian, and other mammalian RPE.13 14 18 19 20 21 22 27 65 66 These comparisons suggest that primary hfRPE cell cultures would be an easily accessible and reproducible model for a wide variety of in vitro experiments using native tissue.
A PubMed search (http://www.ncbi.nih.gov/entrez/ provided in the public domain by the National Center for Biotechnology Information, Bethesda, MD) indicates that primary RPE cultures have been successfully obtained from adult and fetal human, monkey, dog, and rat RPE tissues. However, by far, the most studies have been performed with a human RPE cell line, ARPE-19, spontaneously transformed using multiple trypsinizations. Over the past 10 years, this preparation has been used to study a variety of molecular, biochemical, and cellular mechanisms that determine the structure, function, and regulation of RPE cells and mediate its interactions with the distal retina.64 67 68 69 70 71 72 These studies suggest that ARPE-19 cells retain signal transduction pathways that can mediate or model a variety of native tissue functions. A further advantage of this cell line is that it provides an ostensibly unlimited supply of RPE cells.
The importance of cell culture conditions for the ARPE-19 cell line was emphasized in the original publication64 but those conditions, often ignored, could cause significant changes in the properties of the cell line. For example, in Dunn et al.,64 the CO2 level for culturing ARPE-19 is 10%, whereas many or most other groups have used the more conventional 5% CO2, which, in HCO3-buffered systems, would achieve an extracellular pH of approximately 7.4 with 23.5 mM HCO3. In DMEM, the medium normally used with ARPE-19 cells, the HCO3 levels are closer to 42 mM and require 10% CO2 to maintain a pH of 7.4. Thus, a lower CO2 could result in an abnormally high extracellular pH, which might alter a variety of RPE physiological processes13 and phenotype. The recent paper of Klimanskaya et al.,73 provides a transcriptomic analysis of ARPE-19 cells that show no message for a wide variety of human RPE proteins (e.g., bestrophin and PEDF), but this lack of expression may have partly resulted from the choice of cell culture conditions.74
The ARPE-19 preparation is a spontaneously transformed cell line that may not exhibit important properties of the native tissue, for example: (1) its active gene profile appears to be significantly different from that of the human genome73 74 75; (2) there is no clear EM evidence of apical processes and in later passages, melanization is absent; (3) RT is low72 compared with that of native tissue, suggesting abnormal junctional complexes, and there is no information about apical or basolateral membrane potentials or TEP (channels, electrogenic transporters); (4) there is no extant information or evidence about the ability of ARPE-19 monolayers to transport ions, solute, or fluid; (5) some groups have reported that it may take as long as 90 days to see some pigment development and other functional characteristics; for example, confluent monolayers do not exhibit RT above 100 Ω· cm2 even after 90 days.76
Hu and Bok25 have developed a primary cell culture of human fetal RPE cells derived from fetal eyes of 17- to 24-weeks' gestation. In contrast to the ARPE19 cell line, these cultures retain important characteristics of native adult tissue including melanization, morphology, physiology, protein expression, and trafficking.72 77 They are produced by growing human fetal RPE cells to confluence in low-Ca2+ medium, to obtain the “floaters” that form the basis of all subsequent cultures. These floaters are typically collected over a period of 10 to 30 days, frozen or seeded on cell culture inserts (Transwells; Corning Costar), maintained in Chee's essential medium replacement (CEMR),25 and used 1 to 8 months after seeding.62 It is noteworthy that CEMR contains 1% heat-inactivated calf serum and bovine retina extract (0.5% vol/vol). The number of cultures that can be produced per eye is not specified in these reports.25 62 Possible drawbacks of this preparation include the following: (1) limited number of cell cultures that can be produced per eye; (2) the use of floaters collected over several days in small batches, from one or more eyes, which may preselect for certain RPE cell subtypes and prevent elaboration of the full mosaic of epithelial proteins78 79 ; (3) a culture growing time that varies from 1 month to 2 years25 62 80 ; and (4) the use of bovine retinal extracts that vary from animal to animal in CEMR.
In the present experiments, we sought to make the methodology as widely accessible as possible by using only commercially available supplies. The present methods help ensure that the number of usable zero and first-passage cells are sufficient in quantity for a wide range of different experiments. We have determined that one human fetal eye is sufficient to seed four T25 P0 flasks and that each P0 flask can be passaged into four P1 flasks; these 16 P1 flasks combined contain 160 to 250 million cells. For comparison, the average number of RPE cells in an adult human eye is approximately 5.6 million, and in the fetal eye there are 0.8 million cells, assuming equal density (http://webvision.med.utah.edu/).
This hfRPE cell culture model displays classic epithelial polarity (Fig. 2) as demonstrated by the presence of apical and basolateral membrane-specific channels, transporters, and receptor proteins. Also present are the junctional complex proteins (Fig. 5) that undoubtedly contribute to the maintenance of cell polarity, electrophysiologically measured RT and TEP, and the in vivo differences of the chemical composition between the subretinal and choroidal extracellular spaces.81 82 83 The claudins are a growing family of proteins with at least 24 homologous isoforms. Strong evidence has now accumulated that claudins provide fundamentally important determinants of regulated paracellular permeability and charge selectivity.81 84 85 Claudin-1 and -3 have been identified in the tight junctions of rabbit, mouse, and human fetal RPE44 86 87 and have been localized in the cilia of mouse RPE.88 The developmental appearance of various claudins (for example claudin-1, -2, -5, and -12) and their correlation to changes in RT have been studied in embryonic chick RPE.89 90 Which particular set of claudins serves to help regulate RPE tight junction permeability and RPE physiology remains to be determined. Microarray data in hfRPE indicate that claudin-19 and -10 are most abundant (Wang F, et al. IOVS 2006;47:ARVO E-Abstract 2855). By regulating paracellular cation conductance, claudin-19 is thought to play a significant role in the development of polycystic kidney disease91 whereas claudin-10 has been linked to several cancers. However, the role of these and other claudins (e.g., claudin-1, -2, -3, and -9) in adult human RPE is unknown.
Intracellular microelectrode recordings (Figs. 7 8 9 10 11) measured the steady state difference in apical and basolateral membrane potentials and can be used to calculate apical and basolateral membrane and paracellular path resistances. These confluent RPE monolayers also exhibit levels of net active solute-linked fluid transport comparable to that measured in in vitro native mammalian and human fetal RPE and in the intact eye.13 27
Figure 13 shows that VEGF is preferentially secreted into the basal bath. This polarized secretion may be necessary so that RPE cells can modulate the homeostasis of the extracellular space around Bruch's membrane and at the same time modulate the density of endothelial cell fenestrations in the choroidal blood supply.92 93 PEDF is another small molecule secreted by the RPE. It is a member of a family of proteins (serpins) known to be neuroprotective and antiangiogenic.94 In contrast to VEGF, PEDF in this model is more highly secreted into the apical side of the RPE, possibly for retinal neuroprotection and partly because the RPE is responsible for preventing retinal capillaries from growing into the photoreceptor layer.95 The level of PEDF secretion reported herein is similar to that found in cultured monkey RPE.96
During EM examination of some hfRPE cells, we observed deposits close to the basal side of the epithelium (Fig. 4). The ultrastructure of these deposits is reminiscent of RPE melanin granules and suggests their possible origin. Also, these deposits are commonly found in degenerative disease or when cells have experienced stress.4 32 97 98 Further investigation is necessary to determine the nature of these deposits in hfRPE cells and their relationship to basal laminar deposits in adult AMD eyes. Conversely, the observation that these laminar deposits are also observed in adult degenerative diseases such as AMD or cancer-affected eyes suggests a possible developmental regression in diseased adult eyes. In either case, the present model may be helpful in understanding the physiological consequence of these deposits and their possible role in neurodegenerative disease.
ACKNOWLEDGEMENTS
The authors thank Anahita Jafari and Connie Zhi for expert technical help with cell culture; Chi-Chao Chan, Mary Alice Crawford, and Joe Hackett for help with histology; Shoichiro Tsukita, Kyoto University (claudin 19), Michael Redmond (LRAT, RPE 65), and John Saari (CRALBP and RPE 65) for kindly providing antibodies; and Brian Brooks, Karl Csaky, Igal Gery, Ward Peterson, and Barry Winkler for helpful advice and comments on the manuscript.
REFERENCES
- 1.Gallemore RP, Hughes BA, Miller SS. Retinal pigment epithelial transport mechanisms and their contributions to electroretinogram. Prog Retin Eye Res. 1997;16:509–566. [Google Scholar]
- 2.Roth F, Bindewald A, Holz FG. Key pathophysiologic pathways in age-related macular disease. Graefes Arch Clin Exp Ophthalmol. 2004;242:710–716. doi: 10.1007/s00417-004-0976-x. [DOI] [PubMed] [Google Scholar]
- 3.Anderson DH, Mullins RF, Hageman GS, Johnson LV. A role for local inflammation in the formation of drusen in the aging eye. Am J Ophthalmol. 2002;134:411–431. doi: 10.1016/s0002-9394(02)01624-0. [DOI] [PubMed] [Google Scholar]
- 4.Zarbin MA. Current concepts in the pathogenesis of age-related macular degeneration. Arch Ophthalmol. 2004;122:598–614. doi: 10.1001/archopht.122.4.598. [DOI] [PubMed] [Google Scholar]
- 5.Hageman GS, Anderson DH, Johnson LV, et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc Natl Acad Sci USA. 2005;102:7227–7232. doi: 10.1073/pnas.0501536102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kuksa V, Imanishi Y, Batten M, et al. Retinoid cycle in the vertebrate retina: experimental approaches and mechanisms of isomerization. Vision Res. 2003;43:2959–2981. doi: 10.1016/s0042-6989(03)00482-6. [DOI] [PubMed] [Google Scholar]
- 7.McHenry CL, Liu Y, Feng W, et al. MERTK arginine-844-cysteine in a patient with severe rod-cone dystrophy: loss of mutant protein function in transfected cells. Invest Ophthalmol Vis Sci. 2004;45:1456–1463. doi: 10.1167/iovs.03-0909. [DOI] [PubMed] [Google Scholar]
- 8.Holz FG, Pauleikhoff D, Klein R, Bird AC. Pathogenesis of lesions in late age-related macular disease. Am J Ophthalmol. 2004;137:504–510. doi: 10.1016/j.ajo.2003.11.026. [DOI] [PubMed] [Google Scholar]
- 9.Themistocleous GS, Katopodis H, Sourla A, et al. Three-dimensional type I collagen cell culture systems for the study of bone pathophysiology. In Vivo. 2004;18:687–696. [PubMed] [Google Scholar]
- 10.Mendes F, Doucet L, Hinzpeter A, et al. Immunohistochemistry of CFTR in native tissues and primary epithelial cell cultures. J Cyst Fibros. 2004;3(suppl2):37–41. doi: 10.1016/j.jcf.2004.05.008. [DOI] [PubMed] [Google Scholar]
- 11.Gudjonsson T, Villadsen R, Ronnov-Jessen L, Petersen OW. Immortalization protocols used in cell culture models of human breast morphogenesis. Cell Mol Life Sci. 2004;61:2523–2534. doi: 10.1007/s00018-004-4167-z. [DOI] [PubMed] [Google Scholar]
- 12.Liou GI, Samuel S, Matragoon S, et al. Alternative splicing of the APC gene in the neural retina and retinal pigment epithelium. Mol Vis. 2004;10:383–391. [PubMed] [Google Scholar]
- 13.Hughes BA, Gallemore R, Miller SS. Transport mechanisms in the retinal pigment epithelium. In: Marmor MF, editor. The Retinal Pigment Epithelium. Oxford University Press New York; 1998. pp. 103–134. [Google Scholar]
- 14.Gallemore R, Hughes BA, Miller SS. Light-induced responses of retinal pigment epithelium. In: Marmor MF, editor. The Retinal Pigment Epithelium. Oxford University Press New York; 1998. pp. 175–198. [Google Scholar]
- 15.Steinberg RH. Monitoring communications between photoreceptors and pigment epithelial cells: effects of “mild” systemic hypoxia. Friedenwald lecture. Invest Ophthalmol Vis Sci. 1987;28:1888–1904. [PubMed] [Google Scholar]
- 16.Steinberg RH, Linsenmeier RA, Griff ER. Three light-evoked responses of the retinal pigment epithelium. Vision Res. 1983;23:1315–1323. doi: 10.1016/0042-6989(83)90107-4. [DOI] [PubMed] [Google Scholar]
- 17.Mircheff AK, Miller SS, Farber DB, et al. Isolation and provisional identification of plasma membrane populations from cultured human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1990;31:863–878. [PubMed] [Google Scholar]
- 18.Lin H, Kenyon E, Miller SS. Na-dependent pHi regulatory mechanisms in native human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1992;33:3528–3538. [PubMed] [Google Scholar]
- 19.Quinn RH, Miller SS. Ion transport mechanisms in native human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1992;33:3513–3527. [PubMed] [Google Scholar]
- 20.la Cour M, Lin H, Kenyon E, Miller SS. Lactate transport in freshly isolated human fetal retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1994;35:434–442. [PubMed] [Google Scholar]
- 21.Quinn RH, Quong JN, Miller SS. Adrenergic receptor activated ion transport in human fetal retinal pigment epithelium. Invest Ophthalmol Vis Sci. 2001;42:255–264. [PubMed] [Google Scholar]
- 22.Voloboueva LA, Liu J, Suh JH, et al. (R)-α-lipoic acid protects retinal pigment epithelial cells from oxidative damage. Invest Ophthalmol Vis Sci. 2005;46:4302–4310. doi: 10.1167/iovs.04-1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Joseph DP, Miller SS. Apical and basal membrane ion transport mechanisms in bovine retinal pigment epithelium. J Physiol. 1991;435:439–463. doi: 10.1113/jphysiol.1991.sp018518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Steinberg RH, Miller SS, Stern WH. Initial observations on the isolated retinal pigment epithelium-choroid of the cat. Invest Ophthalmol Vis Sci. 1978;17:675–678. [PubMed] [Google Scholar]
- 25.Hu J, Bok D. A cell culture medium that supports the differentiation of human retinal pigment epithelium into functionally polarized monolayers. Mol Vis. 2001;7:14–19. [PubMed] [Google Scholar]
- 26.Peterson WM, Miller SS. Identification and functional characterization of a dual GABA/taurine transporter in the bullfrog retinal pigment epithelium. J Gen Physiol. 1995;106:1089–1122. doi: 10.1085/jgp.106.6.1089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Maminishkis A, Jalickee S, Blaug SA, et al. The P2Y(2) receptor agonist INS37217 stimulates RPE fluid transport in vitro and retinal reattachment in rat. Invest Ophthalmol Vis Sci. 2002;43:3555–3566. [PubMed] [Google Scholar]
- 28.Edelman JL, Miller SS. Epinephrine stimulates fluid absorption across bovine retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1991;32:3033–3040. [PubMed] [Google Scholar]
- 29.Jiang C, Finkbeiner WE, Widdicombe JH, et al. Altered fluid transport across airway epithelium in cystic fibrosis. Science. 1993;262:424–427. doi: 10.1126/science.8211164. [DOI] [PubMed] [Google Scholar]
- 30.Baffi J, Byrnes G, Chan CC, Csaky KG. Choroidal neovascularization in the rat induced by adenovirus mediated expression of vascular endothelial growth factor. Invest Ophthalmol Vis Sci. 2000;41:3582–3589. [PubMed] [Google Scholar]
- 31.Chan CC, Cogan DG, Bucci FS, et al. Anterior corneal dystrophy with dyscollagenosis (Reis-Bucklers type?) Cornea. 1993;12:451–460. doi: 10.1097/00003226-199309000-00013. [DOI] [PubMed] [Google Scholar]
- 32.Curcio CA, Presley JB, Millican CL, Medeiros NE. Basal deposits and drusen in eyes with age-related maculopathy: evidence for solid lipid particles. Exp Eye Res. 2005;80:761–775. doi: 10.1016/j.exer.2004.09.017. [DOI] [PubMed] [Google Scholar]
- 33.Gong H, Ruberti J, Overby D, et al. A new view of the human trabecular meshwork using quick-freeze, deep-etch electron microscopy. Exp Eye Res. 2002;75:347–358. [PubMed] [Google Scholar]
- 34.Curcio CA, Millican CL. Basal linear deposit and large drusen are specific for early age-related maculopathy. Arch Ophthalmol. 1999;117:329–339. doi: 10.1001/archopht.117.3.329. [DOI] [PubMed] [Google Scholar]
- 35.Furuse M, Hirase T, Itoh M, et al. Occludin: a novel integral membrane protein localizing at tight junctions. J Cell Biol. 1993;123:1777–1788. doi: 10.1083/jcb.123.6.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Furuse M, Fujita K, Hiiragi T, et al. Claudin-1 and -2: novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J Cell Biol. 1998;141:1539–1550. doi: 10.1083/jcb.141.7.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kivela T, Jaaskelainen J, Vaheri A, Carpen O. Ezrin, a membrane-organizing protein, as a polarization marker of the retinal pigment epithelium in vertebrates. Cell Tissue Res. 2000;301:217–223. doi: 10.1007/s004410000225. [DOI] [PubMed] [Google Scholar]
- 38.Bonilha VL, Rodriguez-Boulan E. Polarity and developmental regulation of two PDZ proteins in the retinal pigment epithelium. Invest Ophthalmol Vis Sci. 2001;42:3274–3282. [PubMed] [Google Scholar]
- 39.Miller SS, Steinberg RH. Active transport of ions across frog retinal pigment epithelium. Exp Eye Res. 1977;25:235–248. doi: 10.1016/0014-4835(77)90090-2. [DOI] [PubMed] [Google Scholar]
- 40.Miller SS, Steinberg RH. Passive ionic properties of frog retinal pigment epithelium. J Membr Biol. 1977;36:337–372. doi: 10.1007/BF01868158. [DOI] [PubMed] [Google Scholar]
- 41.Rizzolo LJ. The distribution of Na+,K(+)-ATPase in the retinal pigmented epithelium from chicken embryo is polarized in vivo but not in primary cell culture. Exp Eye Res. 1990;51:435–446. doi: 10.1016/0014-4835(90)90156-o. [DOI] [PubMed] [Google Scholar]
- 42.Hernandez EV, Hu JG, Frambach DA, Gallemore RP. Potassium conductances in cultured bovine and human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1995;36:113–122. [PubMed] [Google Scholar]
- 43.Ruiz A, Bhat SP, Bok D. Expression and synthesis of the Na,K-ATPase beta 2 subunit in human retinal pigment epithelium. Gene. 1996;176:237–242. doi: 10.1016/0378-1119(96)00258-2. [DOI] [PubMed] [Google Scholar]
- 44.Rajasekaran SA, Hu J, Gopal J, et al. Na,K-ATPase inhibition alters tight junction structure and permeability in human retinal pigment epithelial cells. Am J Physiol. 2003;284:C1497–C1507. doi: 10.1152/ajpcell.00355.2002. [DOI] [PubMed] [Google Scholar]
- 45.Steinberg RH. Interactions between the retinal pigment epithelium and the neural retina. Doc Ophthalmol. 1985;60:327–346. doi: 10.1007/BF00158922. [DOI] [PubMed] [Google Scholar]
- 46.Bialek S, Joseph DP, Miller SS. The delayed basolateral membrane hyperpolarization of the bovine retinal pigment epithelium: mechanism of generation. J Physiol. 1995;484:53–67. doi: 10.1113/jphysiol.1995.sp020647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Oakley B, 2nd, Steinberg RH, Miller SS, Nilsson SE. The in vitro frog pigment epithelial cell hyperpolarization in response to light. Invest Ophthalmol Vis Sci. 1977;16:771–774. [PubMed] [Google Scholar]
- 48.Bialek S, Miller SS. K+ and Cl− transport mechanisms in bovine pigment epithelium that could modulate subretinal space volume and composition. J Physiol. 1994;475:401–417. doi: 10.1113/jphysiol.1994.sp020081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Joseph DP, Miller SS. Alpha-1-adrenergic modulation of K and Cl transport in bovine retinal pigment epithelium. J Gen Physiol. 1992;99:263–290. doi: 10.1085/jgp.99.2.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Peterson WM, Meggyesy C, Yu K, Miller SS. Extracellular ATP activates calcium signaling, ion, and fluid transport in retinal pigment epithelium. J Neurosci. 1997;17:2324–2337. doi: 10.1523/JNEUROSCI.17-07-02324.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Strauss O. The retinal pigment epithelium in visual function. Physiol Rev. 2005;85:845–881. doi: 10.1152/physrev.00021.2004. [DOI] [PubMed] [Google Scholar]
- 52.Schneider U, Gelisken F, Inhoffen W, Bartz-Schmidt KU. Pigment epithelial detachments with retinal vascular anomalous complex in age-related macular degeneration. Ophthalmologica. 2005;219:303–308. doi: 10.1159/000086116. [DOI] [PubMed] [Google Scholar]
- 53.Emerson GG, Ghazi NG. Spontaneous rip of the retinal pigment epithelium with a macular hole in neovascular age-related macular degeneration. Am J Ophthalmol. 2005;140:316–318. doi: 10.1016/j.ajo.2005.01.028. [DOI] [PubMed] [Google Scholar]
- 54.Kasahara E, Lin LR, Ho YS, Reddy VN. SOD2 protects against oxidation-induced apoptosis in mouse retinal pigment epithelium: implications for age-related macular degeneration. Invest Ophthalmol Vis Sci. 2005;46:3426–3434. doi: 10.1167/iovs.05-0344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Schuster A, Apfelstedt-Sylla E, Pusch CM, et al. Autoimmune retinopathy with RPE hypersensitivity and ‘negative ERG’ in X-linked hyper-IgM syndrome. Ocul Immunol Inflamm. 2005;13:235–243. doi: 10.1080/09273940590928571. [DOI] [PubMed] [Google Scholar]
- 56.Cahill MT, Mruthyunjaya P, Bowes Rickman C, Toth CA. Recurrence of retinal pigment epithelial changes after macular translocation with 360 degrees peripheral retinectomy for geographic atrophy. Arch Ophthalmol. 2005;123:935–938. doi: 10.1001/archopht.123.7.935. [DOI] [PubMed] [Google Scholar]
- 57.Widdicombe JH, Sachs LA, Finkbeiner WE. Effects of growth surface on differentiation of cultures of human tracheal epithelium. In Vitro Cell Dev Biol Anim. 2003;39:51–55. doi: 10.1290/1543-706X(2003)039<0051:EOGSOD>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 58.Sachs LA, Finkbeiner WE, Widdicombe JH. Effects of media on differentiation of cultured human tracheal epithelium. In Vitro Cell Dev Biol Anim. 2003;39:56–62. doi: 10.1290/1543-706X(2003)039<0056:EOMODO>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 59.Lopez-Souza N, Avila PC, Widdicombe JH. Polarized cultures of human airway epithelium from nasal scrapings and bronchial brushings. In Vitro Cell Dev Biol Anim. 2003;39:266–269. doi: 10.1290/1543-706X(2003)039<0266:PCOHAE>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 60.Widdicombe JH, Sachs LA, Morrow JL, Finkbeiner WE. Expansion of cultures of human tracheal epithelium with maintenance of differentiated structure and function. Biotechniques. 2005;39:249–255. doi: 10.2144/05392RR02. [DOI] [PubMed] [Google Scholar]
- 61.McKay BS, Burke JM. Separation of phenotypically distinct subpopulations of cultured human retinal pigment epithelial cells. Exp Cell Res. 1994;213:85–92. doi: 10.1006/excr.1994.1176. [DOI] [PubMed] [Google Scholar]
- 62.Frambach DA, Fain GL, Farber DB, Bok D. Beta adrenergic receptors on cultured human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1990;31:1767–1772. [PubMed] [Google Scholar]
- 63.Davis AA, Bernstein PS, Bok D, et al. A human retinal pigment epithelial cell line that retains epithelial characteristics after prolonged culture. Invest Ophthalmol Vis Sci. 1995;36:955–964. [PubMed] [Google Scholar]
- 64.Dunn KC, Aotaki-Keen AE, Putkey FR, Hjelmeland LM. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp Eye Res. 1996;62:155–169. doi: 10.1006/exer.1996.0020. [DOI] [PubMed] [Google Scholar]
- 65.Blaug S, Quinn R, Quong J, et al. Retinal pigment epithelial function: a role for CFTR? Doc Ophthalmol. 2003;106:43–50. doi: 10.1023/a:1022514031645. [DOI] [PubMed] [Google Scholar]
- 66.Wang F, Rendahl KG, Manning WC, et al. AAV-mediated expression of vascular endothelial growth factor induces choroidal neovascularization in rat. Invest Ophthalmol Vis Sci. 2003;44:781–790. doi: 10.1167/iovs.02-0281. [DOI] [PubMed] [Google Scholar]
- 67.Marin-Castano ME, Csaky KG, Cousins SW. Nonlethal oxidant injury to human retinal pigment epithelium cells causes cell membrane blebbing but decreased MMP-2 activity. Invest Ophthalmol Vis Sci. 2005;46:3331–3340. doi: 10.1167/iovs.04-1224. [DOI] [PubMed] [Google Scholar]
- 68.Maidji E, Tugizov S, Jones T, et al. Accessory human cytomegalovirus glycoprotein US9 in the unique short component of the viral genome promotes cell-to-cell transmission of virus in polarized epithelial cells. J Virol. 1996;70:8402–8410. doi: 10.1128/jvi.70.12.8402-8410.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Holtkamp GM, Van Rossem M, de Vos AF, et al. Polarized secretion of IL-6 and IL-8 by human retinal pigment epithelial cells. Clin Exp Immunol. 1998;112:34–43. doi: 10.1046/j.1365-2249.1998.00560.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Finnemann SC, Bonilha VL, Marmorstein AD, Rodriguez-Boulan E. Phagocytosis of rod outer segments by retinal pigment epithelial cells requires alpha(v)beta5 integrin for binding but not for internalization. Proc Natl Acad Sci USA. 1997;94:12932–12937. doi: 10.1073/pnas.94.24.12932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Handa JT, Reiser KM, Matsunaga H, Hjelmeland LM. The advanced glycation endproduct pentosidine induces the expression of PDGF-B in human retinal pigment epithelial cells. Exp Eye Res. 1998;66:411–419. doi: 10.1006/exer.1997.0442. [DOI] [PubMed] [Google Scholar]
- 72.Deora AA, Gravotta D, Kreitzer G, et al. The basolateral targeting signal of CD147 (EMMPRIN) consists of a single leucine and is not recognized by retinal pigment epithelium. Mol Biol Cell. 2004;15:4148–4165. doi: 10.1091/mbc.E04-01-0058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Klimanskaya I, Hipp J, Rezai KA, et al. Derivation and comparative assessment of retinal pigment epithelium from human embryonic stem cells using transcriptomics. Cloning Stem Cells. 2004;6:217–245. doi: 10.1089/clo.2004.6.217. [DOI] [PubMed] [Google Scholar]
- 74.Tian J, Ishibashi K, Honda S, et al. The expression of native and cultured human retinal pigment epithelial cells grown in different culture conditions. Br J Ophthalmol. 2005;89:1510–1517. doi: 10.1136/bjo.2005.072108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Sharma RK, Orr WE, Schmitt AD, Johnson DA. A functional profile of gene expression in ARPE-19 cells. BMC Ophthalmol. 2005;5:25. doi: 10.1186/1471-2415-5-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Abe T, Sugano E, Saigo Y, Tamai M. Interleukin-1beta and barrier function of retinal pigment epithelial cells (ARPE-19): aberrant expression of junctional complex molecules. Invest Ophthalmol Vis Sci. 2003;44:4097–4104. doi: 10.1167/iovs.02-0867. [DOI] [PubMed] [Google Scholar]
- 77.Deora AA, Philp N, Hu J, et al. Mechanisms regulating tissue-specific polarity of monocarboxylate transporters and their chaperone CD147 in kidney and retinal epithelia. Proc Natl Acad Sci USA. 2005;102:16245–16250. doi: 10.1073/pnas.0504419102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Burke JM, Hjelmeland LM. Mosaicism of the retinal pigment epithelium: seeing the small picture. Mol Interv. 2005;5:241–249. doi: 10.1124/mi.5.4.7. [DOI] [PubMed] [Google Scholar]
- 79.Shlyonsky V, Goolaerts A, Van Beneden R, Sariban-Sohraby S. Differentiation of epithelial Na+ channel function: an in vitro model. J Biol Chem. 2005;280:24181–24187. doi: 10.1074/jbc.M413823200. [DOI] [PubMed] [Google Scholar]
- 80.Hu JG, Gallemore RP, Bok D, et al. Localization of NaK ATPase on cultured human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1994;35:3582–3588. [PubMed] [Google Scholar]
- 81.Van Itallie CM, Anderson JM. Claudins and epithelial paracellular transport. Annu Rev Physiol. 2006;68:403–429. doi: 10.1146/annurev.physiol.68.040104.131404. [DOI] [PubMed] [Google Scholar]
- 82.Turksen K, Troy TC. Barriers built on claudins. J Cell Sci. 2004;117:2435–2447. doi: 10.1242/jcs.01235. [DOI] [PubMed] [Google Scholar]
- 83.Johnson LG. Applications of imaging techniques to studies of epithelial tight junctions. Adv Drug Deliv Rev. 2005;57:111–121. doi: 10.1016/j.addr.2004.08.004. [DOI] [PubMed] [Google Scholar]
- 84.Schneeberger EE, Lynch RD. The tight junction: a multifunctional complex. Am J Physiol. 2004;286:C1213–C1228. doi: 10.1152/ajpcell.00558.2003. [DOI] [PubMed] [Google Scholar]
- 85.Yu AS, McCarthy KM, Francis SA, et al. Knockdown of occludin expression leads to diverse phenotypic alterations in epithelial cells. Am J Physiol. 2005;288:C1231–C1241. doi: 10.1152/ajpcell.00581.2004. [DOI] [PubMed] [Google Scholar]
- 86.Morcos Y, Hosie MJ, Bauer HC, Chan-Ling T. Immunolocalization of occludin and claudin-1 to tight junctions in intact CNS vessels of mammalian retina. J Neurocytol. 2001;30:107–123. doi: 10.1023/a:1011982906125. [DOI] [PubMed] [Google Scholar]
- 87.Xu H, Dawson R, Crane IJ, Liversidge J. Leukocyte diapedesis in vivo induces transient loss of tight junction protein at the blood-retina barrier. Invest Ophthalmol Vis Sci. 2005;46:2487–2494. doi: 10.1167/iovs.04-1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Nishiyama K, Sakaguchi H, Hu JG, et al. Claudin localization in cilia of the retinal pigment epithelium. Anat Rec. 2002;267:196–203. doi: 10.1002/ar.10102. [DOI] [PubMed] [Google Scholar]
- 89.Kojima S, Rahner C, Peng S, Rizzolo LJ. Claudin 5 is transiently expressed during the development of the retinal pigment epithelium. J Membr Biol. 2002;186:81–88. doi: 10.1007/s00232-001-0137-7. [DOI] [PubMed] [Google Scholar]
- 90.Rahner C, Fukuhara M, Peng S, et al. The apical and basal environments of the retinal pigment epithelium regulate the maturation of tight junctions during development. J Cell Sci. 2004;117:3307–3318. doi: 10.1242/jcs.01181. [DOI] [PubMed] [Google Scholar]
- 91.Lee NP, Tong MK, Leung PP, et al. Kidney claudin-19: localization in distal tubules and collecting ducts and dysregulation in polycystic renal disease. FEBS Lett. 2006;580:923–931. doi: 10.1016/j.febslet.2006.01.019. [DOI] [PubMed] [Google Scholar]
- 92.Roberts WG, Palade GE. Increased microvascular permeability and endothelial fenestration induced by vascular endothelial growth factor. J Cell Sci. 1995;108:2369–2379. doi: 10.1242/jcs.108.6.2369. [DOI] [PubMed] [Google Scholar]
- 93.Yokomori H, Oda M, Yoshimura K, et al. Vascular endothelial growth factor increases fenestral permeability in hepatic sinusoidal endothelial cells. Liver Int. 2003;23:467–475. doi: 10.1111/j.1478-3231.2003.00880.x. [DOI] [PubMed] [Google Scholar]
- 94.Bouck N. PEDF: anti-angiogenic guardian of ocular function. Trends Mol Med. 2002;8:330–334. doi: 10.1016/s1471-4914(02)02362-6. [DOI] [PubMed] [Google Scholar]
- 95.Kijlstra A, La Heij E, Hendrikse F. Immunological factors in the pathogenesis and treatment of age-related macular degeneration. Ocul Immunol Inflamm. 2005;13:3–11. doi: 10.1080/09273940590909185. [DOI] [PubMed] [Google Scholar]
- 96.Pfeffer BA, Becerra SP, Borst DE, Wong P. Expression of transthyretin and retinol binding protein mRNAs and secretion of transthyretin by cultured monkey retinal pigment epithelium. Mol Vis. 2004;10:23–30. [PubMed] [Google Scholar]
- 97.Yazama F, Kadonosono K, Itoh N, Ohno S. Role of matrix metalloproteinase-7 in angiogenesis associated with age-related macular degeneration. J Electron Microsc (Tokyo) 2002;51:127–131. doi: 10.1093/jmicro/51.2.127. [DOI] [PubMed] [Google Scholar]
- 98.Marmorstein AD, Marmorstein LY, Sakaguchi H, et al. Spectral profiling of autofluorescence associated with lipofuscin, Bruch's Membrane, and sub-RPE deposits in normal and AMD eyes. Invest Ophthalmol Vis Sci. 2002;43:2435–2441. [PubMed] [Google Scholar]