

Abstract
Three types of trivalent influenza vaccines were analysed for their in vitro stimulatory properties on immune cells from young healthy volunteers. A whole inactivated virus (WV) vaccine, a conventional subunit (c-SU) preparation and a new virosomal subunit (v-SU) vaccine were used. Blood-derived DC up-regulated MHC class II, CD54, CD80 and CD86 after exposure to WV vaccine, indicating their functional maturation, but were only moderately affected by subunit (SU) vaccines. In addition, IL-12 and tumour necrosis factor-alpha (TNF-α) secretion by DC were markedly enhanced by WV, but not by SU vaccines. The production of IL-2 and interferon-gamma (IFN-γ) by PBMC was also strongly stimulated by WV, but much less by SU vaccines, among which the v-SU vaccine was a better stimulator of IL-2 secretion. In contrast to WV vaccine both SU vaccines were powerful stimulators of PBMC proliferation. Our results suggest that the presence of influenza core components leads to the activation of DC and triggers the production of cytokines by PBMC. SU vaccines are in contrast excellent stimulators of T cell growth. A combination of WV and SU vaccines in immunization regimes might allow optimal T cell priming as well as the efficient generation and maintenance of memory cells.
Keywords: influenza, different vaccines, T cells, dendritic cells, cytokines, immune system
INTRODUCTION
Influenza continues to constitute a major cause of morbidity and mortality in high risk patient groups, such as the very young, the elderly or persons with decreased pulmonary function [1–3]. Vaccination is therefore of utmost importance. Although influenza vaccines reduce the severity of illness, efficacy is limited among certain groups [4–8]. The development of improved vaccines and vaccination strategies is clearly needed to enhance immune responsiveness following vaccination and to achieve high levels of protection in all risk groups. Although there have been numerous recent attempts to improve vaccine potency and efficacy, only a moderate degree of success has been achieved [9–11]. This is at least partly due to our limited understanding of how different influenza vaccines interact with the immune system. Although cellular immunity plays a crucial role in the control of influenza infections, influenza vaccine immunogenicity studies have traditionally focused on antibody production (for review see [7]). Limited information is available on T cell reactivity following influenza vaccination [12–14], while the effects of influenza vaccines on antigen-presenting cells (APC) are only now being investigated [15,16].
The aim of our study was to analyse the effects of three different types of influenza vaccines on cellular immunity. DC, which are the most powerful APC, and PBMC were studied. The stimulatory properties of a whole inactivated influenza virus vaccine (WV vaccine), a conventional influenza subunit preparation (c-SU vaccine), as well as a virosomal subunit vaccine (v-SU vaccine) [11] in which influenza haemagglutinins are incorporated into the surface of liposomes, were compared (Fig. 1)
Fig. 1.
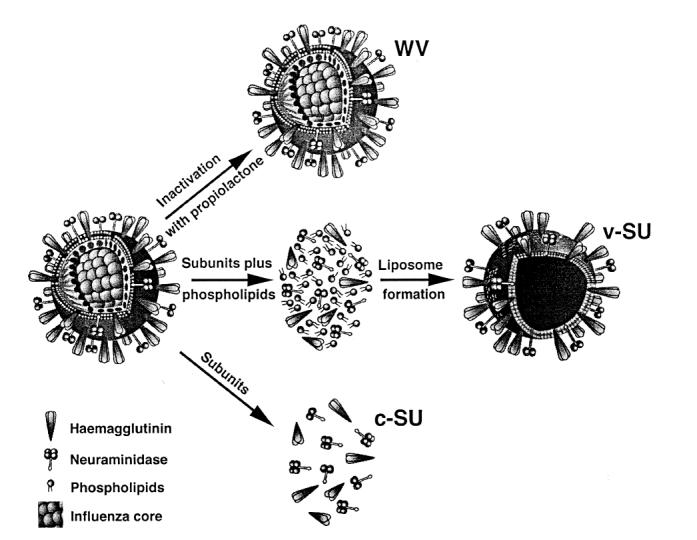
Schematic representation of the three types of influenza vaccines used. The whole virus (WV) vaccine contains intact inactivated virions, while subunit vaccines (SU) contain viral haemagglutinin and neuraminidase. In the case of virosomal SU vaccine, envelope proteins are incorporated into the surface of liposomes composed of phosphatidylcholine and phosphatidylethanolamine. v-SU, Virosomal subunit influenza vaccine; c-SU, conventional subunit influenza vaccine.
MATERIALS AND METHODS
Reagents, monoclonal antibodies and serum
IL-4, granulocyte-macrophage colony-stimulating factor (GM-CSF) and IL-2 were kindly provided by Sandoz Pharma AG (Basel, Switzerland). Mouse MoAbs directed against the following cell surface determinants were used: MHC class II (HLA-DR; An der Grub GmbH, Kaumberg, Austria; FITC-conjugated), CD44 (Monosan, Uden, The Netherlands; FITC-conjugated), CD54 (Monosan; PE-conjugated), CD19 (Dako A/S, Glostrup, Denmark; PE-conjugated), CD3 (SeraLab, Crawley Down, UK; FITC-conjugated), CD1a (Serotec, Oxford, UK; PE-conjugated), CD14 (Serotec; FITC-conjugated), CD80 (Becton Dickinson, San Jose, CA; PE-conjugated) and CD86 (Becton Dickinson; FITC-conjugated). Fetal calf serum (FCS) was purchased from SEBAK GmbH (Stuben, Austria).
Vaccines
A commercially available trivalent intact virion influenza vaccine (Inflexal-Berna; Swiss Serum & Vaccine Institute, Berne, Switzerland), a conventional subunit preparation as well as a new virosomal subunit vaccine were used [11]. Each vaccine lot was checked for its biological activity before release. All vaccines included the following three influenza strains: A/Singapore/6/86 (H1N1), A/Johannesburg/33/94 (H3N2) and B/Beijing/184/93. The concentrations of haemagglutinin (HA) and neuraminidase (NA) of each strain were identical in the three vaccine types. The average diameter of virosomes is 150 nm, similar in size to the whole influenza virus (80–120 nm).
Preparation of PBMC
PBMC were obtained from young healthy volunteers (< 30 years old, mean 26.5 ± 2.1 years, range 24–29 years; n = 21, 13 males, eight females). Cells were purified from heparinized blood by Ficoll–Paque (Pharmacia, Uppsala, Sweden) density centrifugation and washed twice in RPMI 1640 (Gibco, Grand Island, NY). For some experiments PBMC were depleted of adherent cells by a 2-h incubation at 37°C.
Preparation of PBMC for cytokine analysis
PBMC were seeded at a density of 106 cells/well in 48-well plates (Falcon, Becton Dickinson, Franklin Lakes, NJ). They were then cultured at 37°C, 5% CO2 in RPMI, 10% FCS and 1% penicillin/streptomycin (P/S; Gibco) as culture medium (CM) with or without influenza vaccines at a concentration of 1 μg/ml. In pilot experiments this dose had for all vaccine types been found to be optimally stimulatory on all cell types tested. Conditioned supernatants were removed after 48 h for IL-2 analysis and after 7 days for the determination of interferon-gamma (IFN-γ). Supernatants were centrifuged at 250 g and were stored at −20°C until assayed for the presence of cytokines. The optimal incubation periods for the determination of each cytokine following stimulation with the different vaccines had been assessed in pilot experiments.
Purification of DC
DC were prepared according to a published method [17–19]. In brief, PBMC were resuspended in CM and allowed to adhere to six-well plates (Falcon; 9 × 106 cells/well). After 2 h at 37°C, non-adherent cells were removed and adherent cells cultured in CM supplemented with 800 U GM-CSF and 1000 U IL-4 per ml. Cells were then fed every other day with fresh CM containing 800 U GM-CSF and 300 U IL-4 per ml.
Preparation of DC for surface marker and cytokine secretion analysis
After 1 week in culture, DC, which were at that time point mostly non-adherent, were removed from the plate and washed twice in RPMI. They were then counted and incubated at 106 cells per tube in CM at 37°C, 5% CO2 in tissue culture tubes (Greiner, Kremsmünster, Austria) in the absence or presence of influenza vaccine (1 μg/ml). After 24 h supernatants were harvested, centrifuged and stored at −20°C. Cells were washed and analysed by immunofluorescence staining as described below.
Immunofluorescence staining and FACScan analysis
Cells were transferred into round-bottomed tubes (105 cells/tube; Becton Dickinson, Mountain View, CA) and washed at 4°C in PBS containing 0.1% FCS. Antibodies were added and the cells left to incubate at 4°C. After 40 min the cells were washed twice in PBS. Analysis was performed on a Becton Dickinson FACScan. Five thousand scatter-gated cells were analysed in each sample. The frequency and fluorescence profiles of the cells were determined with logarithmic signal amplifiers.
Proliferation assays
PBMC were cultured for 5 days at 105 cells/well in 96-well flat-bottomed plates (Falcon) in CM with and without influenza vaccines (1 μg/ml). After the respective incubation periods cells were pulsed with 1 μCi of 3H-thymidine (Amersham, Aylesbury, UK) and 3H-thymidine incorporation was assessed by scintillation counting. All assays were carried out in triplicates. Results were expressed as ct/min − background proliferation (ct/min in the absence of stimuli).
Cytokine determinations by ELISA
Cytokine concentrations in conditioned supernatants were determined by commercially available ELISA kits: IL-2 (CYF, Immune Sciences Inc., College Park, MD); IL-4 and IFN-γ (Genzyme Corp., Cambridge, MA); IL-12 (Endogen Inc., Cambridge, MA; specific for the p75 heterodimer; sensitivity > 5 pg/ml) and tumour necrosis factor-alpha (TNF-α; Endogen; sensitivity > 5 pg/ml).
Statistical analysis
Student's t-tests for paired and unpaired data were used for statistical evaluation.
RESULTS
WV vaccine stimulates the expression of MHC class II and CD54 on DC
Immunofluorescence staining confirmed that DC expressed high levels of HLA-DR, CD54, CD44 and CD1a. Lack of CD3+, CD14+ and of CD19+ cells demonstrated the purity of the population. Incubation of DC with WV vaccine for 24 h led to a marked increase in the surface expression of MHC class II, CD54, CD80 and CD86 on the great majority of cells (Fig. 2). SU vaccines were less effective, as they were only moderately stimulatory on the expression of both molecules. CD44 was unaffected by influenza vaccines (not shown).
Fig. 2.
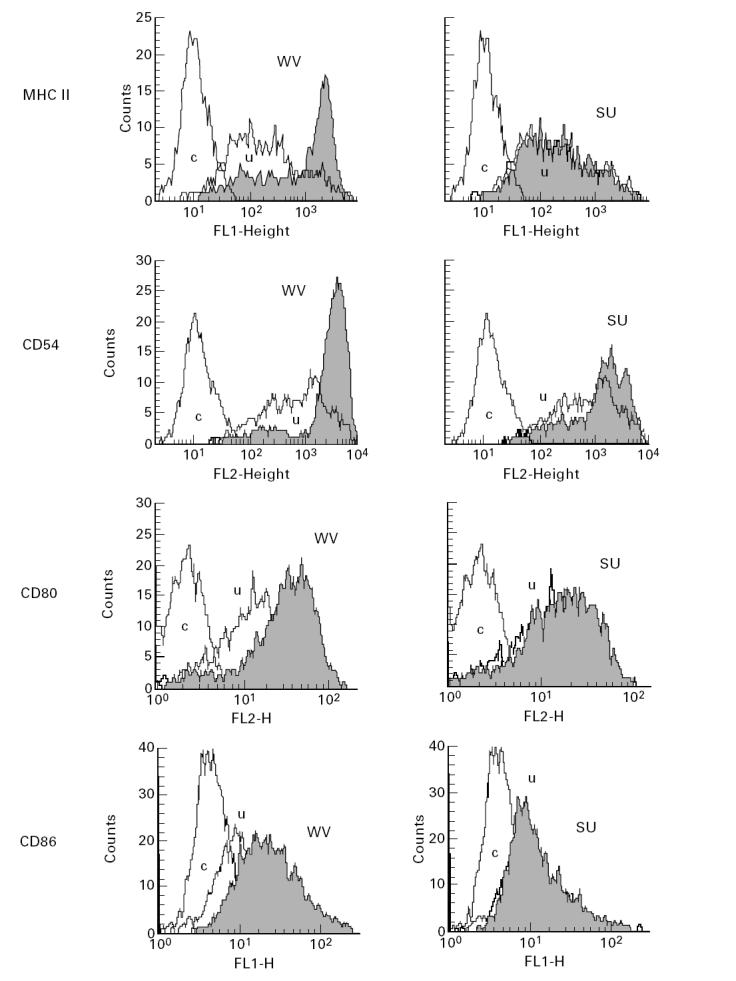
WV vaccine stimulates the expression of MHC class II, CD54, CD80 and CD86 on DC. Immunofluorescence staining of purified DC incubated for 24 h with either WV (left panels) or v-SU (right panels) vaccine. The Figure shows one of 12 identical experiments. WV vaccine is more efficient in stimulating MHC class II, CD54, CD80 and CD86 expression than subunit vaccines. Similar results were obtained when c-SU vaccine was used instead of v-SU vaccine. c, Control; u, unstimulated control.
WV vaccine enhances the secretion of IL-12 and TNF-α by DC
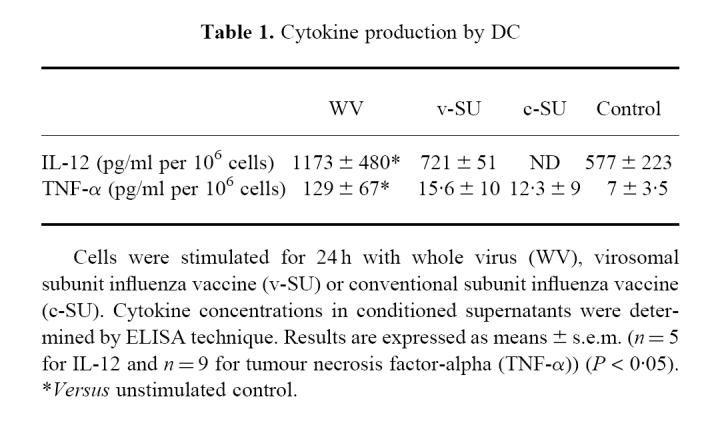
The secretion of IL-12 and TNF-α by DC was assessed by measuring the concentrations of the two cytokines in supernatants conditioned by DC in the presence or absence of influenza vaccines (Table 1). Unstimulated DC produced IL-12, but only low amounts of TNF-α. WV vaccine significantly increased the secretion of IL-12 and of TNF-α. In contrast, both SU vaccines did not significantly affect cytokine production by DC.
Table 1.
Cytokine production by DC
Influenza vaccines stimulate the secretion of IL-2 and IFN-γ by PBMC: WV has a stronger stimulatory effect than SU vaccines
We tested the effects of the different vaccines on the production of cytokines by PBMC (Fig. 3). Unstimulated PBMC produced low amounts of IFN-γ but no IL-2 and no IL-4. Incubation with influenza vaccines led to a pronounced increase in the secretion of IL-2 and IFN-γ, while IL-4 remained undetectable. WV vaccine had a more pronounced stimulatory effect than the SU preparations. v-SU vaccine was still a better stimulator of IL-2 secretion than c-SU vaccine.
Fig. 3.
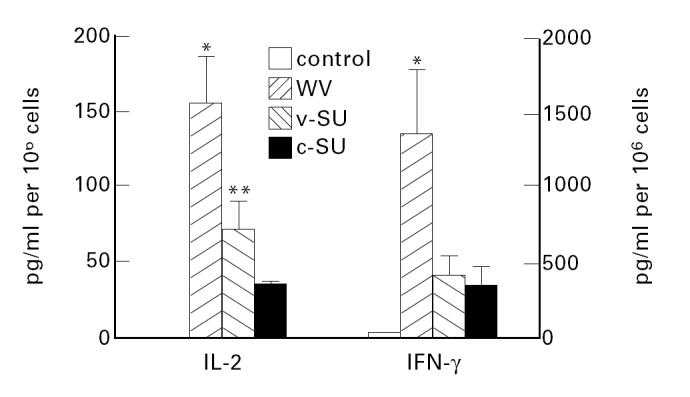
Influenza vaccines stimulate the secretion of IL-2 and IFN-γ by PBMC. WV vaccine has a stronger stimulatory effect than SU vaccines. Cells were stimulated with WV, v-SU or c-SU vaccine. Cytokine concentrations in conditioned supernatants from stimulated and unstimulated cells were determined by ELISA technique. Results are expressed as means ± s.e.m. (n = 15) *P < 0.005 versus SU vaccines; **P < 0.05 versus c-SU.
Influenza vaccines stimulate the proliferation of PBMC: SU vaccines have a stronger stimulatory effect than WV vaccine
PBMC proliferation was significantly enhanced by stimulation with influenza vaccines. Both SU vaccines were by far better stimulators than the WV vaccine, which induced only a moderate proliferation (Fig. 4). The pronounced stimulatory effect of SU vaccines on T cell proliferation was also observed when purified DC loaded with influenza vaccines were used as APC (Fig. 5).
Fig. 4.
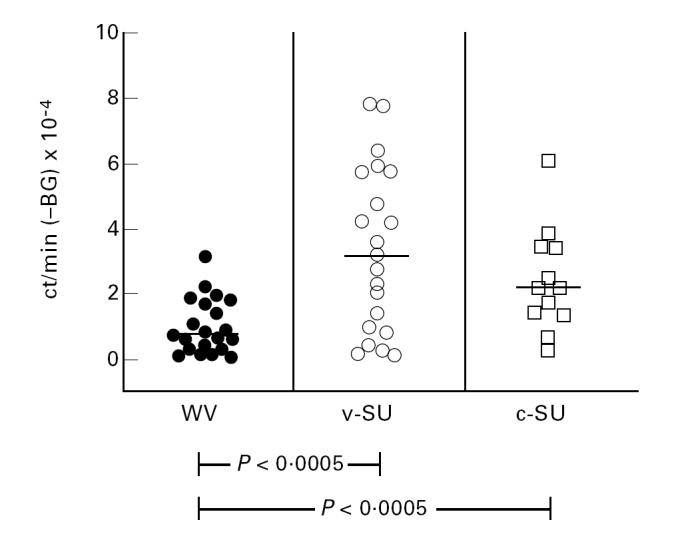
Influenza vaccines stimulate the proliferation of PBMC: SU vaccines have a stronger stimulatory effect than WV vaccine. Proliferation (3H-thymidine incorporation) of PBMC in response to stimulation with WV, v-SU or c-SU (1 μg/ml) vaccine. Results are expressed as mean ct/min − background (BG) ± s.e.m. Proliferation in the absence of antigen was considered as background proliferation.
Fig. 5.
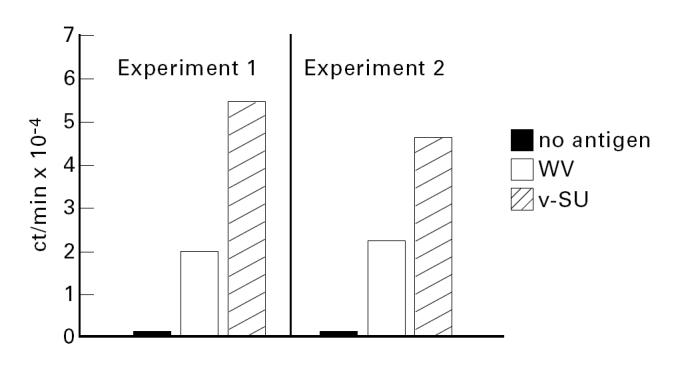
Purified DC loaded with either WV or v-SU vaccine (1 μg/24 h) stimulated the proliferation (3H-thymidine incorporation) of PBMC depleted of adherent cells. Two independent experiments are shown. Results are expressed as ct/min. Means of triplicate determinations are given. Identical results were obtained when c-SU was used instead of v-SU vaccine.
DISCUSSION
DC are highly specialized APC, which are of major importance for the initiation of immune responses [20–22]. DC progenitors from the bone marrow enter the blood and seed non-lymphoid and lymphoid tissues, where they develop into immature DC with high ability for antigen uptake and processing and yet low ability in T cell stimulation [21,23]. Locally produced inflammatory cytokines and/or the encounter of pathogens promote the maturation of DC from a processing to a presenting stage characterized by the expression of costimulatory molecules and cytokines [21,24–26]. Blood-derived DC are mostly related to the mononuclear phagocyte system and large quantities of DC can easily be generated from monocyte-enriched PBMC using GM-CSF and IL-4 [27,28]. This provides a model system to study the effects of potentially stimulatory agents on DC maturation.
Recently DC have attracted a great deal of attention as possible adjuvants for tumour vaccines [29,30]. A detailed understanding of DC activation following vaccination seems therefore a desirable goal for the rational design and development of efficient vaccines.
Relatively little information is presently available on the effects of vaccines on DC. Our present results demonstrate that WV influenza vaccine supports the maturation of blood-derived CD1a+ CD14− DC. It increases the expression of the functionally important surface molecules MHC class II, CD54, CD80 and CD86 and stimulates the secretion of IL-12 and TNF-α. The magnitude of the effects was comparable to the one induced by conventional stimulatory agents such as lipopolysaccharide (LPS) (not shown). DC thus acquire the characteristics necessary for a powerful stimulation of T cells [31]. IL-2 and IFN-γ production by PBMC were, indeed, most efficiently stimulated by WV vaccine. Recent results from our laboratory demonstrate that both cytokines were mainly produced by CD4+ T cells, which were almost exclusively activated and propagated when PBMC were stimulated with inactive influenza virus [32]. In contrast to WV vaccine, SU vaccines did not stimulate DC maturation. This was not due to decreased antigen uptake, as fluorescence-labelled SU vaccines are readily incorporated into any type of APC (unpublished observation).
Our data on the effects of WV influenza vaccine are in good agreement with a recent report on the effects of measles virus strains on DC, which also demonstrates the induction of DC surface marker expression and IL-12 production after virus treatment [33]. Whether the stimulatory effects of different viruses on DC result from a surface contact between measles/influenza virus and their respective receptors is not yet clear, but seems unlikely in view of the very weak effect of SU vaccines, which contain the surface proteins of the influenza virus necessary for receptor binding, but no core components. As measles virus as well as influenza virus belong to the group of RNA viruses, it is tempting to speculate that certain viral RNA sequences might have immunostimulatory properties similar to the short bacterial DNA sequences, which have recently been shown to be able to play an adjuvant role in immune responses [34].
It was also of interest that WV influenza vaccine was only weakly stimulatory on the proliferation of PBMC, which was most pronounced when SU vaccines were used. Diminished T cell growth in spite of high IL-2 production may indicate that APC stimulated with WV vaccine might have an inhibitory effect on T cell proliferation. This is in agreement with a recent concept according to which subpopulations of mature DC may express Fas ligand (FasL) and kill CD4+ T cells that are responding to antigen by a mechanism that involves FasL on the DC and Fas expressed by the activated T cell [35]. A similar mechanism might reduce the number of T cells responsive to WV vaccine stimulation and thus decrease the expansion of influenza-specific populations. Whether T cells activated with WV vaccine could still be rescued by consecutive stimulation with SU vaccines is not yet known. This question seems, however, worth further investigation, as it might be important for the design of vaccination strategies.
Simultaneous consecutive applications of both vaccine types could combine the advantages of both substances. WV vaccine would induce optimal antigen presentation, stimulate the production of Th1-type cytokines and allow the recruitment of CD8+ cells [16]. These qualities might be particularly advantageous in situations in which the efficient activation of naive influenza-specific T cells can be required, such as for instance in the case of antigenic shifts. SU vaccines could, on the other hand, potently trigger the expansion of preprimed helper populations and thus provide a good basis for antibody production and the maintenance of immunological memory. Incorporation of viral proteins into the surface of liposomes may hereby be of additional advantage, as increased IL-2 production was noted when the v-SU preparation was used as a stimulus. This finding is in accordance with results from in vivo studies in which aged persons immunized with v-SU vaccine had increased titres of influenza-specific antibodies when compared with those immunized with WV or c-SU vaccines [11].
In conclusion, our results may help to improve our understanding of cellular immune responses following vaccination with different types of influenza vaccine and may therefore have an impact on the design and clinical trial of more effective vaccination strategies.
Acknowledgments
We are grateful to Georg Wick for his continuous encouragement and support. We thank Nikolaus Romani for technical advice and the critical reading of the manuscript. We would also like to thank Irmgard Albert for excellent technical assistance. This study was supported by a grant from the Austrian National Bank (Grant no. 5568).
REFERENCES
- 1.Glezen WP. Serious morbidity and mortality associated with influenza epidemics. Epidemiol Rev. 1993;4:25–44. doi: 10.1093/oxfordjournals.epirev.a036250. [DOI] [PubMed] [Google Scholar]
- 2.Immunization Practices Advisory Committee. Prevention and control of influenza. Morbid Mortal Wkly Rep. 1996;45:1–24. [Google Scholar]
- 3.Couch RB, Kasel JA, Glezen WP, et al. Influenza: its control in persons and populations. J Infect Dis. 1986;153:431–40. doi: 10.1093/infdis/153.3.431. [DOI] [PubMed] [Google Scholar]
- 4.Wang EEL, Prober CG, Manson B, Corey M, Levison H. Association of respiratory viral infections with pulmonary deterioration in patients with cystic fibrosis. New Engl J Med. 1984;311:1653–8. doi: 10.1056/NEJM198412273112602. [DOI] [PubMed] [Google Scholar]
- 5.Feery BJ, Evered MG, Morrison EM. Different protection rates in various groups of volunteers given subunit influenza virus vaccine in 1976. J Infect Dis. 1979;139:237. doi: 10.1093/infdis/139.2.237. [DOI] [PubMed] [Google Scholar]
- 6.Marks MI. Respiratory viruses in cystic fibrosis. New Engl J Med. 1984;311:1695–6. doi: 10.1056/NEJM198412273112610. [DOI] [PubMed] [Google Scholar]
- 7.Beyer WEP, Palache AM, Baljet M, et al. Antibody induction by influenza vaccine in the elderly: a review of the literature. Vaccine. 1989;7:303–8. doi: 10.1016/0264-410x(89)90150-3. [DOI] [PubMed] [Google Scholar]
- 8.Wortley PM, Farizo KM. Pneumococcal and influenza vaccination levels among HIV-infected adolescents and adults receiving medical care in the United States. Adult and Adolescent Spectrum of HIV Disease Project Group. AIDS. 1994;8:941–4. doi: 10.1097/00002030-199407000-00010. [DOI] [PubMed] [Google Scholar]
- 9.Gupta RK, Siber GR. Adjuvants for human vaccines—current status, problems and future prospects. Vaccine. 1995;13:1263–76. doi: 10.1016/0264-410x(95)00011-o. [DOI] [PubMed] [Google Scholar]
- 10.Monto AS. Influenza vaccines for the elderly. New Engl J Med. 1994;31:807–8. doi: 10.1056/NEJM199409223311212. [DOI] [PubMed] [Google Scholar]
- 11.Glück R, Mischler R, Finkel B, et al. Immunogenicity of new virosome influenza vaccine in elderly people. Lancet. 1994;344:160–3. doi: 10.1016/s0140-6736(94)92758-8. [DOI] [PubMed] [Google Scholar]
- 12.Schwab R, Russo C, Weksler ME. Altered major histocompatibility complex-restricted antigen recognition by T cells from elderly humans. Eur J Immunol. 1992;22:2989–93. doi: 10.1002/eji.1830221134. [DOI] [PubMed] [Google Scholar]
- 13.McElhaney JE, Meneilly GS, Beattie BL, et al. The effect of influenza vaccination on IL-2 production in healthy elderly: implications for current vaccination practices. J Gerontol. 1992;47:3–8. doi: 10.1093/geronj/47.1.m3. [DOI] [PubMed] [Google Scholar]
- 14.Huang YP, Pechere JC, Michel M, et al. In vivo T cell activation, in vitro defective IL-2 secretion, and response to influenza vaccination in elderly women. J Immunol. 1992;148:515–22. [PubMed] [Google Scholar]
- 15.Bhardwaj N, Seder RA, Reddy A, et al. IL-12 in conjunction with dendritic cells enhances antiviral CD8+ CTL response in vitro. J Clin Invest. 1996;98:715–22. doi: 10.1172/JCI118843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bender A, Bui LK, Feldman MV, et al. Inactivated influenza virus, when presented on dendritic cells, elicits human CD8+ cytolytic T cell responses. J Exp Med. 1995;182:1663–71. doi: 10.1084/jem.182.6.1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin-4 and downregulated by tumor necrosis factor-α. J Exp Med. 1994;179:1109–18. doi: 10.1084/jem.179.4.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Romani N, Gruner S, Brang D, et al. Proliferating of dendritic cell progenitors in human blood. J Exp Med. 1994;180:83–93. doi: 10.1084/jem.180.1.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Steger MM, Maczek C, Grubeck-Loebenstein B. Morphologically and functionally intact dendritic cells can be derived from the peripheral blood of aged individuals. Clin Exp Immunol. 1996;105:544–50. doi: 10.1046/j.1365-2249.1996.d01-790.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schuler G, Thurner B, Romani N. Dendritic cells: from ignored cells to major players in T-cell-mediated immunity. Int Arch Allergy Immunol. 1997;112:317–22. doi: 10.1159/000237474. [DOI] [PubMed] [Google Scholar]
- 21.Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991;9:271–96. doi: 10.1146/annurev.iy.09.040191.001415. [DOI] [PubMed] [Google Scholar]
- 22.Romani N, Schuler G. The immunologic properties of epidermal Langerhans cells as a part of the dendritic cell system. Springer Semin Immunol. 1992;13:265–79. doi: 10.1007/BF00200527. [DOI] [PubMed] [Google Scholar]
- 23.McWilliam AS, Nelson D, Thomas JA, et al. Rapid dendritic cell recruitment is a hallmark of the acute inflammatory response at mucosal surfaces. J Exp Med. 1994;179:1331–6. doi: 10.1084/jem.179.4.1331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Caux C, Liu YJ, Banchereau J. Recent advances in the study of dendritic cells and follicular dendritic cells. Immunol Today. 1995;16:2–4. doi: 10.1016/0167-5699(95)80061-1. [DOI] [PubMed] [Google Scholar]
- 25.MacPherson GG, Jenkins CD, Stein MJ, et al. Endotoxin-mediated dendritic cell release from the intestine. Characterization of released dendritic cells and TNF dependence. J Immunol. 1995;154:1317–22. [PubMed] [Google Scholar]
- 26.Sallusto F, Cella M, Danieli C, et al. Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartment: downregulation by cytokines and bacterial products. J Exp Med. 1995;182:389–400. doi: 10.1084/jem.182.2.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Peters JH, Gieseler R, Thiele B, et al. Dendritic cells: from ontogenetic orphans to myelomonocytic descendants. Immunol Today. 1996;17:273–8. doi: 10.1016/0167-5699(96)80544-5. [DOI] [PubMed] [Google Scholar]
- 28.Zhou LJ, Tedder TF. CD14+ blood monocytes can differentiate into functionally mature CD83+ dendritic cells. Proc Natl Acad Sci USA. 1996;93:2588–92. doi: 10.1073/pnas.93.6.2588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hsu FJ, Benike C, Fagnoni F, et al. Vaccination of patients with B-cell lymphoma using autologous antigen-pulsed dendritic cells. Nat Med. 1996;2:52–58. doi: 10.1038/nm0196-52. [DOI] [PubMed] [Google Scholar]
- 30.Young JW, Inaba K. Dendritic cells as adjuvants for class I major histocompatibility complex-restricted antitumor immunity. J Exp Med. 1996;183:7–11. doi: 10.1084/jem.183.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cella M, Sallusto F, Lanzavecchia A. Origin, maturation and antigen presenting function of dendritic cells. Curr Opin Immunol. 1997;9:10–16. doi: 10.1016/s0952-7915(97)80153-7. [DOI] [PubMed] [Google Scholar]
- 32.Saurwein-Teissl M. 1998. The immune response to influenza vaccines: studies in the elderly and the young. [Thesis] [Google Scholar]
- 33.Schnorr JJ, Xanthakos S, Keikavoussi P, et al. Induction of maturation of human blood dendritic cell precursors by measles virus is associated with immunosuppression. Proc Natl Acad Sci USA. 1997;94:5326–31. doi: 10.1073/pnas.94.10.5326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Roman M, Martin-Orozco E, Goodman JS, et al. Immunostimulatory DNA sequences function as T helper-1-promoting adjuvants. Nat Med. 1997;3:849–54. doi: 10.1038/nm0897-849. [DOI] [PubMed] [Google Scholar]
- 35.Süss G, Shortman K. A subclass of dendritic cells kills CD4 T cells via Fas/Fas-ligand-induced apoptosis. J Exp Med. 1996;183:1789–96. doi: 10.1084/jem.183.4.1789. [DOI] [PMC free article] [PubMed] [Google Scholar]