

Abstract
Individuals with cerebellar lesions are impaired in the timing of repetitive movements that involve the concatenation of discrete events such as tapping a finger. In contrast, these individuals perform comparably to controls when producing continuous repetitive movements. Based on this, we have proposed that the cerebellum plays a key role in event timing—the representation of the temporal relationship between salient events related to the movement (e.g., flexion onset or contact with a response surface). In the current study, we used fMRI to examine cerebellar activity during discrete and continuous rhythmic movements. Participants produced rhythmic movements with the index finger either making smooth, continuous transitions between flexion and extension or with a pause inserted before each flexion phase making the movement discrete. Lateral regions in lobule VI, ipsilateral to the moving hand were activated in a similar manner for both conditions. However, activation in the superior vermis was significantly greater when the movements were discrete compared to when the movements were continuous. This pattern was not evident in cortical regions within the field of view, including M1 and SMA. The results are consistent with the hypothesis that subregions of the cerebellum are selectively engaged during tasks involving event timing.
Patients with cerebellar damage exhibit increased temporal variability in the production of timed movements (Ivry and Keele, 1989; Spencer et al., 2003). A corresponding deficit is observed on duration discrimination tasks (Ivry and Keele, 1989; Mangels et al., 1998; Nichelli et al., 1996) suggesting a critical role for the cerebellum in the representation of temporal information. Imaging studies of production (Kawashima et al., 2000; Lutz et al., 2000) and perception (Jueptner et al., 1995; Lewis and Miall, 2003a; Smith et al., 2003) tasks have provided converging evidence for this hypothesis.
A recent series of studies points to an important constraint on the functional domain of cerebellar timing. Patients with cerebellar damage exhibit increased temporal variability when producing repetitive movements that entail discrete events which demarcate successive intervals. For example, during finger tapping, contact with the table surface might constitute such an event. In contrast, these patients show minimal or no impairment when producing repetitive movements in a smooth, continuous manner (Spencer et al., 2003). The dissociation is not dependent on utilization of haptic signals; when finger movements are made in the air, increased temporal variability is only observed when the patients are required to pause before each flexion phase. Moreover, the impairment in discrete movements is not related to the initiation and termination demands unique to these actions (Spencer et al., 2005).
We have proposed that the critical distinction between discrete and continuous movement timing is the way in which movements are controlled (Ivry et al., 2002; Spencer et al., 2003). For discrete movements, we hypothesize that an explicit process, mediated by the cerebellum, specifies the timing of successive events. This event timing hypothesis also provides a parsimonious account of the cerebellar contribution for temporal processing in perception and sensorimotor learning tasks. Continuous movements, by nature, lack an event structure; for these movements timing may be achieved through the control of a secondary variable, such as angular velocity, which does not involve the cerebellum.
In the present experiment we sought converging evidence for the role of the cerebellum in the control of discrete movements and, more specifically, sought to identify critical regions within the cerebellum for this form of control, using fMRI with neurologically healthy individuals. The movements required flexion–extension of the index finger without contacting a surface. Similar finger movements have been shown to evoke significant cerebellar activation during tasks requiring timed movements (e.g., Penhune et al., 1998; Ramnani and Passingham, 2001) and activation associated with these movements can be distinguished from that associated with movements produced by other effectors (Grodd et al., 2001). For discrete movements, participants were instructed to pause before each flexion cycle. For continuous movements, participants were instructed to move as smoothly as possible without pausing. We expected to observe cerebellar activation for both conditions. Of primary interest was the difference between the two movement types. In particular, we expected to observe areas in the cerebellum with greater activity during the discrete condition than the continuous condition. We hypothesized that no cerebellar regions would exhibit greater activity during continuous movements compared to discrete movements.
Materials and methods
Participants
Twenty-one right-handed adults (11 females; 18–38 years) from U. C. Berkeley were recruited for the experiment. Handedness was verified with the Edinburgh Handedness Inventory (Oldfield, 1971). Experimental procedures were approved by the Committee for Protection of Human Subjects at U. C. Berkeley. Participants gave informed consent after a description of the experiment was provided and were compensated financially for their participation.
Movement task
The task was to tap the index finger in one of four ways (2 movement rates × 2 movement types) by flexing and extending the finger at the metacarpo-phalangeal joint (see Spencer et al., 2003 Experiment 3). Prior to imaging, participants were trained to make repetitive movements with the index finger of the right hand using, in separate trials, a continuous or discrete mode of control. All movements were produced in the air without the finger contacting a surface. For the continuous task, participants were instructed to keep the finger moving at all times by making smooth transitions between flexion and extension. In the discrete task, participants were instructed to make abrupt flexion–extension movements with a brief pause prior to the next movement phase, resulting in staccato-like movements. We did not provide explicit instructions concerning the duration of the pause. Rather, participants were instructed to insert a consistent pause while maintaining the target cycle duration.
During training, prior to going into the scanner, the target cycle duration was specified by an auditory metronome, with an inter-tone onset time of 550 ms or 950 ms. Each training trial consisted of 10 tones and the participant was required to move in synchrony with the tones. Participants rested their forearm on the arm of the chair and held the hand flat, palm facing downward. For all conditions the participant was to initiate the flexion phase with the onset of each tone. Following this paced phase, the participants were instructed to continue moving at the established rate for 20 additional cycles. Finger movements were monitored with an Ascension miniBIRD 3-D tracking system (138-Hz sampling rate). Feedback from this system was used to calculate the mean movement rate for the 20 unpaced cycles. This information was provided to the participant at the end of each trial. In addition, the experimenter monitored the movements and provided verbal feedback to ensure that they were sufficiently discrete or continuous depending on the condition. Participants performed five trials of each of the four conditions (2 movement types × 2 rates).
In the subsequent imaging session, the participant lay supine in the MR scanner, arms along the side of the body. The right forearm was supported by cushions. The hand was extended beyond the cushions such that the index finger could comfortably move without contacting the gurney or the participant's body. Participants were instructed to hold the hand flat with the palm facing downward during scanning runs.
During each scanning run, 16 s movement epochs alternated with 16 s rest epochs. For the movement epochs, one of four visual prompts, projected on a screen in front of the participant, indicated the required condition. These prompts were the phrases “Slow Continuous”, “Fast Continuous”, “Slow Discrete”, and “Fast Discrete”. The participant was instructed to adopt the specified movement mode and rate for the 16 s epoch in which the prompt was visible. Between movement epochs, the word “Rest” was displayed on the monitor for 16 s; the participant was instructed to not move during this epoch. Participants were fitted with a 16 sensor, fiber-optic motion tracking glove (Fifth-Dimension Technology, Irvine, CA, sampling rate = 50 Hz). The output from the glove was monitored on-line by the experimenter to ensure that the participants maintained the required posture and moved at the appropriate times.
Subjects performed 4 scanning runs. Each run included 2 epochs of each of the four movement conditions, with the order randomized.
Verb generation task
The fMRI data were obtained with a high-field (4T) MRI scanner. Higher field strength is associated with reduced sensitivity in areas of high magnetic field inhomogeneity (see Norris, 2003 for a review). This is a particular risk for lateral and inferior aspects of the cerebellum. We therefore sought to independently establish that the MRI system was sensitive to activation in the lateral cerebellum. To this end, 14 of the participants were scanned while performing a verb-generation task (McCarthy et al., 1993; Peterson et al., 1988). This task was chosen because it has previously been shown to activate lateral regions of the cerebellar hemisphere (e.g., Seger et al., 2000). Concrete nouns or strings of X's were presented in the center of the display. Participants were instructed to silently think of a verb associated with the noun (e.g., for the word “Dog”, an appropriate response would be “Bark”). For the control condition, participants were instructed to passively view the string of X's. Each stimulus was presented for 2 s.
fMRI acquisition
Given our focus on the cerebellar signal associated with these two classes of movements, we chose MRI parameters (e.g., slice angle and thickness) that would maximize the signal strength from this region. Furthermore, in different groups of participants, we used two MR coils: a surface coil expected to provide maximum coverage and signal from the cerebellum and a standard whole-head coil with slice angles selected to cover the cerebellum and targeted cortical regions. A similar two-coil comparison has proven useful a previous study of cerebellar function (Le et al., 1998).
Functional images were acquired with a 4T Varian INOVA MR scanner (Palo Alto, CA) using a gradient echo-planar (EPI) pulse sequence (8 slices interleaved, thickness = 3.5 mm, gap = 0.5 mm, TR = 550 ms; 2-shot sequence; TE = 28 ms; matrix size = 64 × 64; FOV = 22.4 cm). The slice angle, approximately 56°, was selected to achieve optimal coverage of both the primary motor cortex and cerebellum. For each scanning run 323 functional volumes were acquired (approximately 5 min). At the end of the scanning session a high resolution structural 3-D MPFLASH scan was acquired (TR = 9 ms; TE = 4.8 ms; TI = 300 ms). This image was used for spatial normalization in order to do group random effects statistics (see below).
Acquisition parameters for the verb generation task were identical to the finger tapping task, with the exception that 478 functional images were collected across the 2 scanning runs (approximately 3 min per run).
The fMRI data were collected using two different receive coils. For 14 participants (7 females), a standard Varian whole-head transmit/receive coil was used to acquire the MR data (head coil group). For seven participants (4 females), we used a receive-only surface coil positioned over the cerebellum (surface coil group). We opted to include both coils given their relative strengths and weaknesses. The head coil allowed us to simultaneously measure activity in the cerebellum, M1, and SMA. However, due to magnetic field inhomogeneities caused by air-bone interfaces in sinus cavities and the ear canals, the signal from a whole-head coil may be poor at the margins of the brain, including lateral regions of the cerebellum. The surface coil was expected to increase the signal to noise ratio (SNR), providing higher spatial resolution relative to head coils (Fisher et al., 1985). Thus, the use of a surface coil in the second group of participants should provide the optimal cerebellar signal for our magnet; however, it would not be sufficient to measure cortical activity in M1 or SMA. The actual coverage provided by both coils is illustrated in Fig. 1.
Fig. 1.
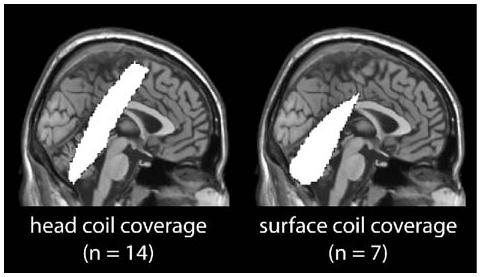
Extent of coverage (in white) for the head and surface coil groups.
Data processing and analysis: kinematic data
Kinematic analyses during scanning were based on the output of the sensor located at the base of the index finger on the motion tracking glove. This sensor provides a measure of angular deviation of the finger relative to when the hand is flat. This angular signal is an absolute difference; thus, the output signal does not differentiate between flexion and hyperextension about this joint. For the most part, the hyperextensions were relatively small and of short duration. Nonetheless, their presence required that we perform a threshold procedure to help identify kinematic events. For this purpose, a horizontal line was drawn through a position × time plot of the movement epoch. The vertical position of this line was adjusted such that unambiguous flexion phases fell above the line. Cycle duration was then automatically calculated as the time between successive points of peak flexion.
To quantify the relative discreteness of each trial, we also calculated the percentage of each cycle in which the velocity of the finger was more than 5% of the maximum velocity for that trial. Large values would correspond to trials in which the movements are continuous. This procedure was necessary given our inability to differentiate between small flexion and hyperflexion movements. It results in an underestimation of the movement phase of each trial. However, it proved sufficient to verify that the participants were following instructions in the scanner and allowed us to obtain rudimentary kinematic measures.
Due to technical problems with the motion glove, kinematic data were not obtained from two participants. We also excluded the kinematic data from a third participant because a significant portion of the trial appeared to involve hyperextension of the finger and we could not unambiguously distinguish these phases from flexion phases. Thus, the kinematic analyses are based on data from 18 participants.1 For these participants, we obtained kinematic measures on 86% of the trials. For the remaining 14%, the kinematic (flexion/hyperextension) output was ambiguous on some of the cycles. We conservatively opted to exclude these trials.
Data processing and analysis: fMRI data
All images were reconstructed from k-space using local software. Functional data were analyzed using SPM2 (Wellcome Institute of Cognitive Neurology, London, UK). The EPI images for each subject were corrected for differences in the slice acquisition time, and realigned to the first image in the series. Coverage from the functional volumes and the 4T environment made automated coregistration insufficient. Thus, the images were manually reoriented to be in the same coordinate frame as the anatomical T1 image. Each volume was normalized into the same coordinate frame as the MNI-template brain by using the individual T1-weighted anatomical image and warping it to the MNI-template using a 7 × 8 × 7 parameter nonlinear transform (Friston, 1995). The resulting transformation parameters were applied to all EPI images. After normalization, all images were smoothed using an 8 mm isotropic Gaussian kernel.
Estimations of task-related changes were performed using the general linear model (GLM) (Kiebel and Holmes, 2004) in SPM2. Separate parameter estimates were determined for each of the four movement conditions. Estimated head-movement parameters, obtained from the realignment process, and block effects were also included as covariates in the estimation of the GLM. For a given effect of interest, whole-brain contrast maps were determined for each participant. Assessment of statistical significance for a given contrast was determined using a random effects analysis by performing a one-sample t-test on the contrast maps. Because we have low degrees of freedom for this analysis, we used SnPM randomization (Nichols and Hayasaka, 2003) with 8-mm variance smoothing to detect areas of significant signal change that survive correction for multiple comparisons. Moreover, for display purposes we converted the standard SPM contrast maps to percent signal change for each condition by multiplying the parameter estimates for the conditions by the height of a regressor for a single block, dividing by the session means, and multiplying by 100.2
The analysis procedure was identical for the verb generation task except that we used the contrast images directly for display of activation rather than the contrast maps given that the primary interest was in the location of peak activation rather than the absolute signal change. Verb generation data was collected from 6 and 8 participants in the surface coil and head coil groups, respectively.
In addition to the whole-brain random effects analysis, we also performed a local region-of-interested (ROI) analysis. In Talairach space we identified two cerebellar and two cortical ROIs. The cerebellar ROIs were defined by the data from participants in the surface coil group and comparisons were examined in the data from participants in the head coil group. These ROIs consisted of a region in the superior vermis (2, −56, −22; x, y, z) and another in lobule V–VI (22, −56, −26) in the right hemisphere (Schmahmann et al., 2000). The vermal ROI was based on a contrast of discrete minus continuous conditions and the hemispheric ROI was based on a contrast in which the rest condition was subtracted from the mean of all of the movement conditions.
The cortical ROIs were identified using data from the head coil group based on an all-movements minus rest contrast, and taking the coordinates of the peak voxels for the largest two clusters. The first ROI was found in the primary motor cortex (M1; −44, −16, 52) and the second was found in the supplementary motor area (SMA; 4, 2, 60). The ROIs were defined as the location of the peak voxel in each region (uncorrected p=0.001). Each ROI was generated by taking a sphere of voxels surrounding each point (4 voxel radius, approximately 8 mm). Given the tight convolutions of the cerebellum, this ROI size was expected to be sufficient for identifying focal activity while excluding signal from adjacent areas.
Analyses of task-related effects were assessed using the Marsbar toolbox in SPM2. These analyses were based on data acquired from the head coil group because a) they provide an independent dataset from that used to generate the cerebellar ROIs and b) they have enough coverage to get signal in the upper cortical regions. Due to misaligned slice angles and individual differences in cortical structure, we did not have adequate coverage of the cortical ROIs in three participants. Therefore, the cortical ROI analyses were based on 11 participants.
Results
Movement kinematics
During training, participants had little difficulty matching their movement rates to the auditory metronome and maintained that rate during the unpaced phase of the trials. During scanning, we did not provide a metronome since we feared that the periodic pulses of the scanner might produce an interfering signal. Participants were very good at producing two different speeds, (F(1,71)=53.3, p<0.001) with the cycle duration for the fast movements averaged 57% of the cycle duration for the slow movements. However, the movements tended to be slower than those used during training and this effect was especially pronounced in the continuous conditions (Table 1). The amplitudes of the movements did not differ across tasks (F(1,71)=0.23, p=0.64) or rate (F(1,71)=0.3, p=0.58).
Table 1.
Summary of kinematic variables obtained during the scanning session when participants produced rhythmic movements in response to verbal cues without the assistance of a metronome
| Fast continuous | Slow continuous | Fast discrete | Slow discrete | |
|---|---|---|---|---|
| Cycle duration | 682.5(37.1) | 1232.2(107.1) | 634.7(20.2) | 967.5(37.9) |
| Pause duration | 128.8(12.9) | 227.9(19.2) | 220.4(19.6) | 378.6(38.2) |
| Movement duration | 553.7(27.3) | 1004.4(92.0) | 414.3(15.0) | 588.9(34.4) |
| Percent movement | 81.4(1.1) | 81.0(0.9) | 65.7(2.1) | 61.3(3.0) |
| Amplitude | 86.8(13.2) | 97.1(13.8) | 83.4(14.2) | 84.5(14.2) |
Values in parentheses are standard deviations.
To verify that the participants adopted different strategies when instructed to produce continuous or discrete movements, we calculated the percentage of each cycle in which the finger was moving with a velocity above 5% of the maximum velocity. In accordance with the instructions, the percent movement time was greater for continuous movements compared to discrete movements (F(1,71)=3.6, p<0.001) and this variable did not vary with movement rate (F(1,71)=1.08, p=0.30). Informal observation during the training phase and, to a limited extent, during scanning, verified that participants followed instructions to make smooth, gradual transitions on continuous trials and paused prior to each flexion phase on discrete trials.
For a given cycle duration, movement velocity must be greater in the discrete condition compared to the continuous condition. Likewise, the percentage of movement time must be greater in the continuous condition, assuming the participant pauses in the discrete condition. Given these inherent differences, the target rates were chosen with the goal of equating the movement portion of the fast continuous and the slow discrete conditions. As can be seen in Table 1, this objective was roughly accomplished. The mean duration of the movement phase was similar in the fast continuous and slow discrete conditions, (t(17)=0.76, p=0.46), as was the movement amplitude, (t(17)=0.53, p=0.60) From this, we infer that the velocity of the finger movements was similar in the two conditions.
Random effects analyses
All movements versus rest
Collapsed across both movement tasks and rates, cerebellar activation relative to rest was centered in vermal lobule VI (8, −66, −22) and extended laterally to right hemisphere lobule VI (22, −58, −28) for the head coil group (Fig. 2). The extent of this large region of activation included right hemisphere lobule V–VI, an area shown previously to have a prominent representation of the finger (e.g., Grodd et al., 2001). A very similar pattern of activation was observed in the surface coil group (10, −62, −18 to 28, −54, −28). Consistent with previous observations, this activation was also seen in similar regions in the left cerebellum but the contralateral activation failed to reach significance for either group.
Fig. 2.
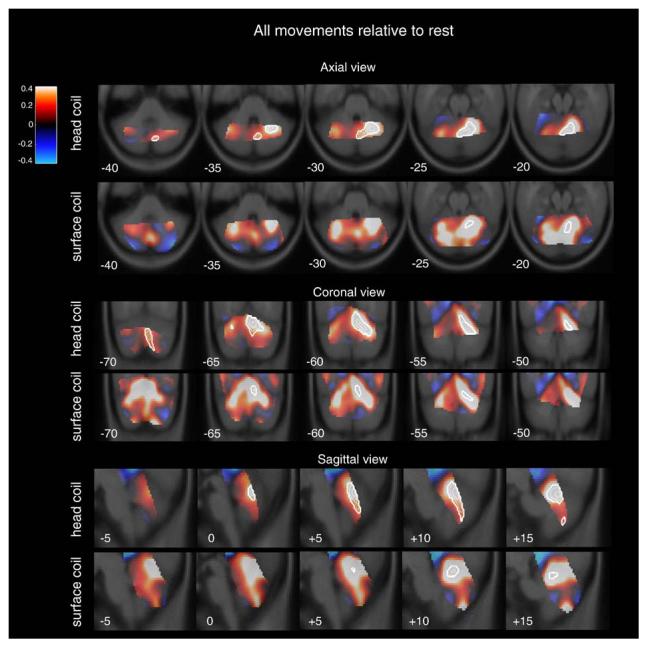
Percent signal change (color maps) within the cerebellum for all movement conditions, collapsed, relative to rest. Warm colors represent areas in which the signal was greater during movement; cool colors represent areas where signal was greater during rest. White contour overlays represent regions which reached statistical significance (p<0.05).
Activation in selected regions of the cortex was evaluated from the data of the head coil group. In a subtraction of all movement conditions minus rest, a prominent increase in the BOLD response was observed in left M1 (−42, −20, 52) and left SMA (2, 0, 60) (Fig. 3A).
Fig. 3.
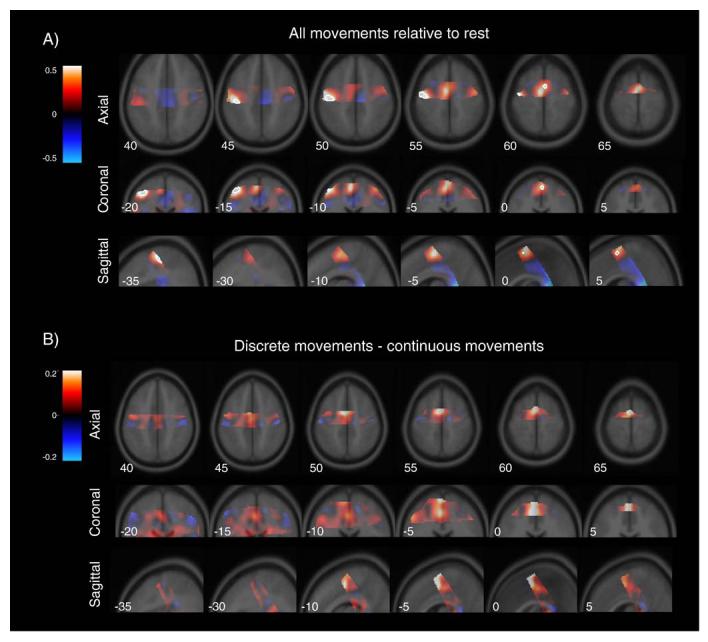
Percent signal change in cortical regions covered in the head coil group. A) All movements (warm colors) relative to rest (cool colors). B) Discrete movements (warm colors) relative to continuous movements (cool colors). Note the difference in scale for A and B.
Discrete versus continuous movements
We directly compared the discrete and continuous conditions, collapsing across the two rates. Given the overlap and consistency of the results within the cerebellum in the comparison of all movements minus rest, we chose to collapse the coil groups for this contrast to maximize power. As shown in Fig. 4, during discrete movements, significantly greater signal change was observed in the superior vermis, lobule V/VI (0, −62, −20). No other areas in the cerebellum, in either coil group, were more active in the discrete condition, nor were there any cerebellar regions in which the signal change was significantly greater for continuous movements.
Fig. 4.
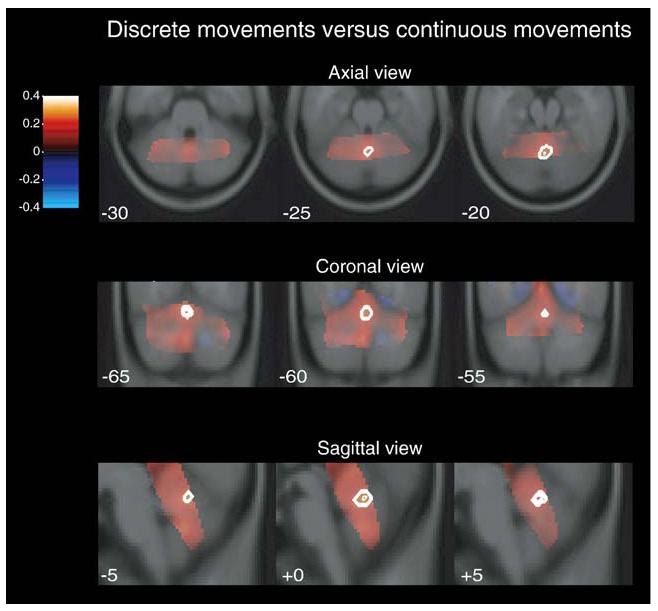
Percent signal change within the cerebellum collapsed across the head and surface coil groups for discrete movements (warm colors) relative to continuous movements (cool colors). White contours represent areas in which the signal was significantly greater (p<0.05) for discrete movements.
The increase in vermal activation during discrete movements compared to continuous movements was also present in each group when the analysis was performed separately (head coil: 8, −66, −22; surface coil: 3, −62, −18). To directly compare the results from the two scanning protocols, we used a bootstrapping method (Manly, 1997). The difference between the two coils for the discrete versus continuous comparison was not significant (p=0.11).
In the cortex, none of the voxels that were activated in the all-movement contrast showed a reliable difference between the discrete and continuous conditions (Fig. 3B). This null result includes those voxels within SMA and M1.
ROI analysis results
ROI analyses were performed for the superior vermis and right hemisphere lobule V of the cerebellum and for SMA and left M1 of the cortex (see Materials and methods). These results are presented in Fig. 5. A two-way ANOVA (task×rate) compared the percent signal change within each ROI. While our focus is on the contrast of discrete and continuous movements, it is also important to consider the effect of movement rate. For a given cycle duration, maximum velocity during the movement phase must be greater in the discrete conditions. Faster movements yield a greater change in the BOLD signal than slower movements in several areas including the cerebellum (Deiber et al., 1999; Jancke et al., 1999; Sadato et al., 1997). Thus, it is necessary to determine if the greater activation observed for discrete movements relative to continuous movements can be attributed to or is modulated by rate differences.
Fig. 5.
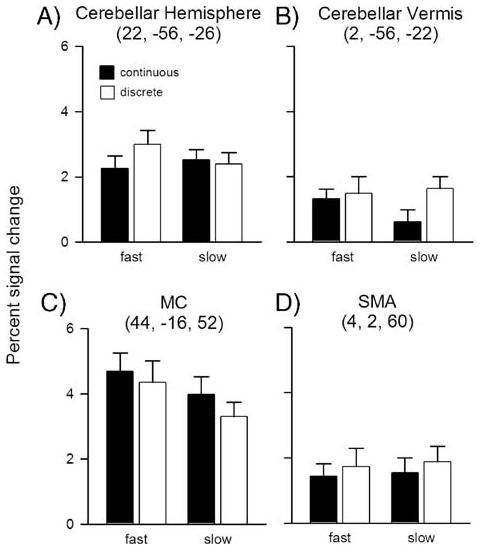
Percent signal change in each condition in A) right cerebellar hemisphere, B) cerebellar vermis, C) left motor cortex, and D) supplementary motor area. Error bars represent standard error. A and B are for the surface coil group only; C and D are for the head coil group only.
In accord with the random effects analysis, the signal change in the superior vermis ROI was greater for discrete movements than for continuous movements (main effect of task type: F(1,13)=9.9, p<0.01). The main effect of rate was not significant (F(1,13)=3.0, p=0.11), although the task×rate interaction approached significance (F(1,13)=4.0, p=0.07). We also performed a direct comparison of the slow discrete and fast continuous tasks given that kinematic variables such as movement velocity and amplitude were similar in these conditions. The signal change in the vermal ROI was greater for slow discrete movements relative to the fast continuous movements, although this effect, was only marginally significant (t(13)=1.6, p=0.06).
In the right cerebellar hemisphere, the main effects of task (F(1,13)=2.6, p=0.13) and rate (F(1,13)<1) were not reliable. The interaction term approached significance (F(1,13)=3.9, p=0.07), an effect driven by the increase in the activation of this region in the fast discrete condition (Fig. 5A). A rate effect has been observed in this region in other studies using traditional tapping tasks (Deiber et al., 1999; Jancke et al., 1999; Sadato et al., 1997).
Within the cerebral cortex, the main effect of task was not significant for either the M1 or SMA ROIs in the head coil group (M1: F(1,10)=3.8, p=0.08; SMA: F(1,10)=1.2, p=0.31). The main effect of rate was not significant in the SMA (F(1,10)<1), but was reliable in M1 (F(1,10)=9.1, p=0.01). The latter effect was due to an increase in activation for the fast movements compared to the slow movements. This observation is consistent with previous reports showing that M1 is sensitive to movement rate (Khushu et al., 2001; Rao et al., 1996). The interaction of task×rate was not significant in either cortical region (M1: F(1,10)<1; SMA: F(1,10)<1).
Assessment of cerebellar coverage with verb-generation task
As expected, a lateral region in right Crus I (36, −68, −34) was activated during covert verb generation compared to passive viewing of the string of X's (F(1,13)<1). This region is distinct from that identified in the contrast of the movement conditions to rest: the center is 18 voxels (36 mm) lateral to the lobule VI region activated during the movement conditions. Interestingly, while we expected the signal from lateral cerebellar cortex to be stronger with the surface coil, the pattern of activation in the verb-generation task was similar for the surface and head coil groups.
Discussion
Cerebellar vermis for discrete movement timing
Based on studies of individuals with cerebellar lesions (Spencer et al., 2003), we predicted that the cerebellum would be disproportionately activated during the production of rhythmic movements in which the successive cycles were marked by discrete events compared to when the cycle transitions were continuous. In accord with this prediction, activation of a focal region in lobule V/VI of the vermis was greater for the discrete conditions compared to the continuous conditions. Indeed, within our field of view, this was the only subcortical or cortical region responsive to this task manipulation.
A number of imaging studies have reported cerebellar activation associated with the timing of rhythmic movements (Elsinger et al., 2003; Lutz et al., 2000; Penhune et al., 1998; Ramnani and Passingham, 2001; Toma et al., 2003). The present study, however, suggests that the engagement of the cerebellum may depend on how such movements are produced, or more importantly, how they are represented. Despite the rather subtle difference between the discrete and continuous conditions, activation in the cerebellum was greater when the participants were required to insert a brief pause prior to the onset of each movement cycle. We have proposed that under such conditions, the task is represented as a concatenation of successively timed events and that the cerebellum provides the representation of this temporal event structure (Ivry et al., 2002). Continuous movements lack such a structure; for these movements, timing may be emergent.
Alternative accounts of the observed dissociation between the discrete and continuous conditions must be considered. First, the greater activation in the vermis may not be related to event timing per se, but rather to requirements associated with producing discrete movements. Our neuropsychological studies suggest that the impairment observed in patients with cerebellar lesions in timing of discrete movements is not due to monitoring and/or controlling the transitions between successive cycles (Spencer et al., 2005). In that study, participants pressed and released a response key in conditions with and without a temporal goal. Although all tasks required rapid movements to initiate and terminate each response, the patients were only impaired when required to insert a pause of a specified duration between the onset and offset phases. While this finding does not speak directly to the present imaging results, it does argue against the idea that the cerebellar contribution in the discrete task is due to the control requirements associated with the transitions.
Second, the greater activation for discrete relative to continuous movements may be associated with differences in movement frequency for these two tasks. Increases in cortical and subcortical activations corresponding to increases in movement frequency have been reported in a number of regions (Deiber et al., 1999; Jancke et al., 1999; Khushu et al., 2001; Rao et al., 1996; Sadato et al., 1997). The direct comparison of the slow discrete and fast continuous conditions provides the best match in terms of maximum velocity; it is also conservative in the sense that the percentage of movement time is much greater in the continuous condition. Nonetheless, in this restricted analysis, there remained a strong trend for activation to be greater for the discrete condition. Similarly, patients with cerebellar lesions show increased temporal variability when producing discrete movements even when the velocity profile of these movements is roughly matched to that produced during continuous movements (Spencer et al., 2003). Thus, we do not expect that the greater activation observed for discrete movements in the vermal region is directly related to kinematic differences between the discrete and continuous movements. Rate effects associated with discrete finger movements within the cerebellum have not been reported in the vermis; rather, these effects have been identified in hemispheric lobule V (Jancke et al., 1999).
Third, given that the vermis receives extensive input from the somatosensory system (Bushara et al., 2001; Robertson, 1984; Robertson et al., 1982), the increased activation in this area for the discrete conditions could result from more abrupt changes in proprioceptive signals generated with these movements. Alternatively, proprioceptive signals might be more important during discrete tapping because they help define the salient events that are required for producing rhythmic, discrete movements. Our experiment does not provide a means to evaluate these hypotheses, although we return to this issue below.
The midline region associated with the timing of discrete movements was unexpected. First, models of cerebellar–cortical interactions have linked lateral regions of the cerebellum with response planning (e.g., Allen and Tsukahara, 1974); event timing would seem to be an integral part of planning processes. Second, in patient studies, increases in timing variability have been associated with lesions of lateral aspects of the neocerebellum (Ivry et al., 1988; Spencer et al., 2003). Patients with focal midline lesions may also have difficulty producing rhythmic movements, but their deficit has been attributed to an impairment in response implementation, rather than central timing (Ivry et al., 1988).
The current work should motivate lesion analysis in future patient studies, as well as a re-examination of past studies. Lesion analysis in the Ivry et al. (1988) study was rather crude, limited to reproductions of selected CT/MRI slices. From these sections, it appears that for patients in the midline group, the lesions were generally more inferior than the activation site associated with the discrete movement conditions. Moreover, in a study involving a much larger number of patients (Harrington et al., 2004), increases in timing variability were restricted to patients with more superior lesions, many of which included superior vermis.
Consistent with the present report, a number of other imaging studies of timing have reported activation in superior vermis, in addition to other cerebellar foci (Ramnani and Passingham, 2001; Tesche and Karhu, 2000; Toma et al., 2003; Tregellas et al., 2006). In a recent study, Bengtsson et al. (2005) compared activation patterns during timed movements produced vocally or with finger movements. A region spanning vermal lobules V–VI was similarly activated in both conditions. This effector independence would be at odds with accounts relating the activation to motor or somatosensory task components rather than timing, although it is possible that subtle changes in the spatial distribution of the activation patterns could not be detected with fMRI. Activation in lobule V/VI of superior vermis has also been reported in perceptual tasks requiring event-based timing (Tesche and Karhu, 2000; Xu et al., 2006).
Role of the lateral hemisphere is discrete and continuous movements
We did observe task-related activation in the lobule V–VI portions of the ipsilateral cerebellar hemisphere and a trend of similar activation in the contralateral hemisphere. Notably, activation in this region was similar in the discrete and continuous conditions. This region includes the “hand area” of the cerebellum, and has previously been identified in imaging studies involving repetitive movement (e.g., Kawashima et al., 2000; Rao et al., 1997) or stimulation (Bushara et al., 2001; Takahashi et al., 2003) of the hand. We expect that the activation of lobule V–VI is related to more generic aspects of the sensorimotor requirements for finger movements rather than timing.
We were concerned that the signal from lateral neocerebellar regions might be insufficient to detect subtle differences between the discrete and continuous conditions. While we did have a significant drop-off in signal near the edges of the cerebellum, especially in the most posterior and inferior aspects, the verb generation task produced a strong activation in Crus 1, lateral and posterior to the hand region in lateral cerebellum. This result suggests that the absence of activation in lateral neocerebellum for both discrete and continuous movements is unlikely due to a sensitivity problem with the 4T magnet.
Cortical response in SMA and M1
SMA activity was observed for both the continuous and discrete conditions relative to rest. SMA activity has been reported in a number of previous studies involving timed movements (Elsinger et al., 2003; Lutz et al., 2000; Rao et al., 1997) as well as in some imaging studies of time perception (Ferrandez et al., 2003; Macar et al., 2002; Rao et al., 2001). Indeed, based on a review of the imaging literature, Lewis and Miall (2003b) hypothesized a critical role in motor timing for the SMA, perhaps in concert with the cerebellum. The current results do not implicate SMA in the conditions we associate with event timing, although the mean activation levels are greater in the two discrete conditions compared to the continuous conditions at similar rates (see Fig. 5D).
In M1, the BOLD response was greater for the fast movements, regardless of whether they were produced in a discrete fashion or continuously. Activation in M1 has previously been shown to be velocity dependent (Jancke et al., 1998; Rao et al., 1996), a result that we replicate here. It is noteworthy that the activation in primary motor cortex was distinct from that found in the cerebellar vermis (and the trend observed in SMA), suggesting that M1 and the cerebellum are associated with different movement parameters.
Conclusions
Discrete movements are marked by salient events—haptic contact, or in this study, the abrupt onset of each flexion cycle. We have proposed that the cerebellum plays a central role in the representation of the timing of these events. In accord with this prediction, a region within the cerebellum was more active when people produced rhythmic movements by imposing a short pause between each cycle compared to when the movements were produced without this discontinuity; this activation was centered in lobule V/VI of the vermis. The computational basis for this vermal activation remains to be elucidated. It may reflect an explicit temporal representation, indicating the timing of salient events, a computation that may be context-independent. Alternatively, given the extensive somatosensory projection to this vermal region, temporal control during rhythmic movements may involve monitoring intervals defined by sensory signals with respect to a temporal goal (see Aschersleben and Prinz, 1995).
Acknowledgments
This work was supported by NIH grants NS048012 and NS30256.
Footnotes
While the data from three individuals was excluded from the kinematic analyses, the data for these participants remains included in the imaging analysis given that the reasons for exclusion were merely due to technical issues in the kinematic recording. We expect their performance was similar to that of the other participants and, in an informal assessment of the imaging data, did not observe any abnormalities in their results.
References
- Allen GI, Tsukahara N. Cerebrocerebellar communications systems. Physio. Rev. 1974;54:957–1006. doi: 10.1152/physrev.1974.54.4.957. [DOI] [PubMed] [Google Scholar]
- Aschersleben G, Prinz W. Synchronizing actions with events: the role of sensory information. Percept. Psychophys. 1995;57:305–317. doi: 10.3758/bf03213056. [DOI] [PubMed] [Google Scholar]
- Bengtsson SL, Ehrsson HH, Forssberg H, Ullen F. Effector-independent voluntary timing: behavioural and neuroimaging evidence. Eur. J. Neurosci. 2005;22:3255–3265. doi: 10.1111/j.1460-9568.2005.04517.x. [DOI] [PubMed] [Google Scholar]
- Bushara KO, Wheat JM, Khan A, Mock BJ, Turski PA, Sorenson J, Brooks BR. Multiple tactile maps in the human cerebellum. NeuroReport. 2001;12(11):2483–2486. doi: 10.1097/00001756-200108080-00039. [DOI] [PubMed] [Google Scholar]
- Deiber MP, Honda M, Ibanez V, Sadato N, Hallett M. Mesial motor areas in self-initiated and externally triggered movements examined with fMRI: effect of movement type and rate. J. Neurophysiol. 1999;81:3065–3077. doi: 10.1152/jn.1999.81.6.3065. [DOI] [PubMed] [Google Scholar]
- Elsinger CL, Rao SM, Zimbelman JL, Reynolds NC, Blindauer KA, Hoffman RG. Neural basis for impaired time reproduction in Parkinson's disease: an fMRI study. J. Int. Neuropsychol. Soc. 2003;9(7):1088–1098. [Google Scholar]
- Ferrandez AM, Hugueville L, Lehericy S, Poline JB, Marsault C, Pouthas V. Basal ganglia and supplementary motor area subtend duration and perception: an fMRI study. NeuroImage. 2003;19:1532–1544. doi: 10.1016/s1053-8119(03)00159-9. [DOI] [PubMed] [Google Scholar]
- Fisher MR, Barker B, Amparo EG, Brandt G, Brant-Zawadzkil M, Hricak H, Higgins CB. MR Imaging using specialized coils. Radiology. 1985;157:443–447. doi: 10.1148/radiology.157.2.4048453. [DOI] [PubMed] [Google Scholar]
- Friston KJ. Statistical parametric mapping: ontology and current issues. J. Cereb. Blood Flow Metab. 1995;15:361–370. doi: 10.1038/jcbfm.1995.45. [DOI] [PubMed] [Google Scholar]
- Grodd W, Hulsmann E, Lotze M, Wildgruber D, Erb M. Sensorimotor mapping of the human cerebellum: fMRI evidence of somatotopic organization. Hum. Brain Mapp. 2001;13:55–73. doi: 10.1002/hbm.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrington DL, Lee RR, Boyd LA, Rapcsak SZ, Knight RT. Does the representation of time depend on the cerebellum? Effect of cerebellar stroke. Brain. 2004;127:1–14. doi: 10.1093/brain/awh065. [DOI] [PubMed] [Google Scholar]
- Ivry RB, Keele SW. Timing functions of the cerebellum. J. Cogn. Neurosci. 1989;1:136–152. doi: 10.1162/jocn.1989.1.2.136. [DOI] [PubMed] [Google Scholar]
- Ivry RB, Keele SW, Diener HC. Dissociation of the lateral and medial cerebellum in movement timing and movement execution. Exp. Brain Res. 1988;73:167–180. doi: 10.1007/BF00279670. [DOI] [PubMed] [Google Scholar]
- Ivry RB, Spencer RMC, Zelaznik HN, Diedrichsen J. The cerebellum and event timing. Ann. N. Y. Acad. Sci. 2002;978:302–317. doi: 10.1111/j.1749-6632.2002.tb07576.x. [DOI] [PubMed] [Google Scholar]
- Jancke L, Specht K, Mirzazade S, Loose R, Himmelbach M, Lutz K, Shah NJ. A parametric analysis of the “rate effect” in the sensorimotor cortex: a functional magnetic resonance imaging analysis in human subjects. Neurosci. Lett. 1998;252:37–40. doi: 10.1016/s0304-3940(98)00540-0. [DOI] [PubMed] [Google Scholar]
- Jancke L, Specht K, Mirzazade S, Peters M. The effect of finger-movement speed of the dominant and the subdominant hand on cerebellar activation: a functional magnetic resonance imaging study. NeuroImage. 1999;9(5):497–507. doi: 10.1006/nimg.1998.0426. [DOI] [PubMed] [Google Scholar]
- Jueptner M, Rijntjes M, Weiller C, Faiss JH, Timmann D, Mueller SP, Diener HC. Localization of a cerebellar timing process using PET. Neurology. 1995;45:1540–1545. doi: 10.1212/wnl.45.8.1540. [DOI] [PubMed] [Google Scholar]
- Kawashima R, Okuda J, Umetsu A, Sugiura M, Inoue K, Suzuki K, Tabuchi M, Tsukiura T, Narayan S, Nagasaka T, et al. Human cerebellum plays an important role in memory-timed finger movement: an fMRI study. J. Neurophysiol. 2000;83:1079–1087. doi: 10.1152/jn.2000.83.2.1079. [DOI] [PubMed] [Google Scholar]
- Khushu S, Kumaran SS, Tripathi RP, Gupta A, Jain PC, Jain V. Functional magnetic resonance imaging of the primary motor cortex in humans: response to increased functional demands. J. Biosci. 2001;26:205–215. doi: 10.1007/BF02703644. [DOI] [PubMed] [Google Scholar]
- Kiebel S, Holmes A. The general linear model. In: Frackowiak RSJ, Friston KJ, Frith CD, Dolan RJ, Price CJ, Zeki S, Ashburner J, Penny W, editors. Human Brain Function. 2nd ed. Elsevier Academic Press; San Diego: 2004. pp. 749–779. [Google Scholar]
- Le TH, Pardo JV, Hu X. 4 T-fMRI study of nonspatial shifting of selective attention: cerebellar and parietal contributions. J. Neurophysiol. 1998;79:1535–1548. doi: 10.1152/jn.1998.79.3.1535. [DOI] [PubMed] [Google Scholar]
- Lewis PA, Miall RC. Brain activation patterns during measurement of sub- and supra-second intervals. Neuropsychologia. 2003a;41:1583–1592. doi: 10.1016/s0028-3932(03)00118-0. [DOI] [PubMed] [Google Scholar]
- Lewis PA, Miall RC. Distinct systems for automatic and cognitively controlled time measurement: evidence from neuroimaging. Curr. Opin. Neurobiol. 2003b;13:250–255. doi: 10.1016/s0959-4388(03)00036-9. [DOI] [PubMed] [Google Scholar]
- Lutz K, Specht K, Shah NJ, Jancke L. Tapping movements according to regular and irregular visual timing signals investigated with fMRI. NeuroReport. 2000;11(6):1301–1306. doi: 10.1097/00001756-200004270-00031. [DOI] [PubMed] [Google Scholar]
- Macar F, Lejeune H, Bonnet M, Ferrara A, Pouthas V, Vidal F, Maquet P. Activation of the supplementary motor area and of attentional networks during temporal processing. Exp. Brain Res. 2002;142:475–485. doi: 10.1007/s00221-001-0953-0. [DOI] [PubMed] [Google Scholar]
- Mangels JA, Ivry R, Shimizu N. Dissociable contributions of prefrontal and neocerebellar cortex to time perception. Cogn. Brain Res. 1998;7:15–39. doi: 10.1016/s0926-6410(98)00005-6. [DOI] [PubMed] [Google Scholar]
- Manly B. Randomization, Bootstrap and Monte Carlo Methods in Biology. Chapman and Hall/CRC; New York: 1997. [Google Scholar]
- McCarthy G, Blamire AM, Rothman DL, Guetter R, Shulman RG. Echo-planar magnetic resonance imaging studies of frontal cortex activation during word generation in humans. Proc. Natl. Acad. Sci. 1993;90:4952–4956. doi: 10.1073/pnas.90.11.4952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichelli P, Alway D, Grafman J. Perceptual timing in cerebellar degeneration. Neuropsychologia. 1996;34:863–871. doi: 10.1016/0028-3932(96)00001-2. [DOI] [PubMed] [Google Scholar]
- Nichols T, Hayasaka S. Controlling the familywise error rate in functional neuroimaging: a comparative review. Stat. Methods Med. Res. 2003;12(5):419–446. doi: 10.1191/0962280203sm341ra. [DOI] [PubMed] [Google Scholar]
- Norris DG. High field human imaging. J. Magn. Reson. Imaging. 2003;18(5):519–529. doi: 10.1002/jmri.10390. [DOI] [PubMed] [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Penhune VB, Zatorre RJ, Evans AC. Cerebellar contributions to motor timing: a PET study of auditory and visual rhythm reproduction. J. Cogn. Neurosci. 1998;10(6):752–765. doi: 10.1162/089892998563149. [DOI] [PubMed] [Google Scholar]
- Peterson SE, Fox PT, Posner MI, Mintun M, Raichle ME. Positron emission tomographic studies of the cortical anatomy of single-word processing. Nature. 1988;331:585–589. doi: 10.1038/331585a0. [DOI] [PubMed] [Google Scholar]
- Ramnani N, Passingham RE. Changes in the human brain during rhythm learning. J. Cogn. Neurosci. 2001;13(7):952–966. doi: 10.1162/089892901753165863. [DOI] [PubMed] [Google Scholar]
- Rao SM, Bandettini PA, Binder JR, Bobholz JA, Hammeke TA, Stein EA, Hyde JS. Relationship between finger movement rate and functional magnetic resonance signal change in human primary motor cortex. J. Cereb. Blood Flow Metab. 1996;16:1250–1254. doi: 10.1097/00004647-199611000-00020. [DOI] [PubMed] [Google Scholar]
- Rao SM, Harrington DL, Haaland KY, Bobholz JA, Cox RW, Binder JR. Distributed neural systems underlying the timing of movements. J. Neurosci. 1997;17(14):5528–5535. doi: 10.1523/JNEUROSCI.17-14-05528.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao SM, Mayer AR, Harrington DL. The evolution of brain activation during temporal processing. Nat. Neurosci. 2001;4(3):317–323. doi: 10.1038/85191. [DOI] [PubMed] [Google Scholar]
- Robertson LT. Topographic features of climbing fiber input in the rostral vermal cortex of the cat cerebellum. Exp. Brain Res. 1984;55:445–454. doi: 10.1007/BF00235275. [DOI] [PubMed] [Google Scholar]
- Robertson LT, Laxer KD, Rushmer DS. Organization of climbing fiber input from mechanoreceptors to lobule V vermal cortex of the cat. Exp. Brain Res. 1982;46:281–291. doi: 10.1007/BF00237186. [DOI] [PubMed] [Google Scholar]
- Sadato N, Ibanez V, Campbell G, Deiber MP, LeBihan D, Hallett M. Frequency-dependent changes of regional cerebral blood flow during finger movements: functional MRI compared to PET. J. Cereb. Blood Flow Metab. 1997;17:670–679. doi: 10.1097/00004647-199706000-00008. [DOI] [PubMed] [Google Scholar]
- Schmahmann J, Doyon J, Toga AW, Petrides M, Evans AC. MRI Atlas of the Human Cerebellum. Academic Press; San Diego, CA: 2000. [DOI] [PubMed] [Google Scholar]
- Seger CA, Desmond JE, Glover GH, Gabrieli JDE. Functional magnetic resonance imaging evidence for right-hemisphere involvement in processing unusual semantic relationships. Neuropsychology. 2000;14:361–369. doi: 10.1037//0894-4105.14.3.361. [DOI] [PubMed] [Google Scholar]
- Smith A, Taylor E, Lidzba K, Rubia K. A right hemispheric frontocerebellar network for time discrimination of several hundreds of milliseconds. NeuroImage. 2003;20:344–350. doi: 10.1016/s1053-8119(03)00337-9. [DOI] [PubMed] [Google Scholar]
- Spencer RMC, Zelaznik HN, Diedrichsen J, Ivry RB. Disrupted timing of discontinuous movements by cerebellar lesions. Science. 2003;300:1437–1439. doi: 10.1126/science.1083661. [DOI] [PubMed] [Google Scholar]
- Spencer RMC, Ivry RB, Zelaznik HN. Role of the cerebellum in movements: control of timing or movement transitions? Exp. Brain Res. 2005;161(3):383–396. doi: 10.1007/s00221-004-2088-6. [DOI] [PubMed] [Google Scholar]
- Takahashi S, Abe K, Yanagihara T, Sakoda S, Tanaka H, Hirabuki N, Nakamura H, Fujita N. A functional MRI study of somatotopic representation of somatosensory stimulation in the cerebellum. Neuro-radiology. 2003;45:149–152. doi: 10.1007/s00234-002-0935-3. [DOI] [PubMed] [Google Scholar]
- Tesche CD, Karhu JJT. Anticipatory cerebellar responses during somatosensory omission in man. Hum. Brain Mapp. 2000;9:119–142. doi: 10.1002/(SICI)1097-0193(200003)9:3<119::AID-HBM2>3.0.CO;2-R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toma K, Ozawa M, Matsuo K, Nakai T, Fukuyama H, Sato S. The role of the human supplementary motor area in reactive motor operation. Neurosci. Lett. 2003;344:177–180. doi: 10.1016/s0304-3940(03)00417-8. [DOI] [PubMed] [Google Scholar]
- Tregellas JR, Davalos DB, Rojas DC. Effect of task difficulty on the functional anatomy of temporal processing. NeuroImage. 2006;32(1):307–315. doi: 10.1016/j.neuroimage.2006.02.036. [DOI] [PubMed] [Google Scholar]
- Xu D, Liu T, Ashe J, Bushara KO. Role of the olivo-cerebellar system in timing. J. Neurosci. 2006;26:5990–5995. doi: 10.1523/JNEUROSCI.0038-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]