

Abstract
Naïve T-cells divide and mature, both functionally and phenotypically, upon stimulation through the T-cell receptor. Although much is known about the overall changes that occur in naïve cells upon TCR stimulation, and the different memory/effector populations that arise following stimulation, the relationship between cell division and functional and phenotypical changes that occur after activation is poorly understood. Here, we examine the early stages of human naïve and antigen-experienced T-cell activation, and the relationship between cell division and acquisition of effector function during the transition from resting antigen-experienced or naïve T-cells into effector cells. Stimulated naïve T-cells proliferate prior to acquisition of effector function, as measured by cytokine production and expression of effector-associated cell surface molecules. Additionally, we show that interlukin-7 (IL-7) can drive proliferation of naïve T-cells without TCR:MHC peptide interactions. IL-7 alone does not, however, drive the proliferation of antigen-experienced T-cells. Memory T-cells will divide in response to exogenous IL-7 but only in the presence of naïve T-cells and IL-2. This study contributes to the current understanding of the mechanistic differences between naïve and memory T-cell responses by defining the functional and phenotypic changes that occur to T-cells after stimulation.
Keywords: naïve T-cells, antigen-experienced T-cells, cellular proliferation, T-cell stimulation, IFN-γ, CTLA-4, IL-7
Introduction
Upon initial activation through the T-cell receptor (TCR), naive T-cells begin to modulate expression of surface molecules associated with activation, costimulation, and adhesion. The expression pattern of these molecules has been used to further define effector and antigen-experienced T-cell subsets [1–4]. After a primary immunological response subsequent encounters with antigen lead to a rapid anamnestic response. This accelerated recall response is due to both an increased frequency of antigen-specific T-cells as well as qualitative changes in memory T-cells that allow them to elicit rapid effector function [5–8]. Memory T-cells also differ from naïve T-cells in migratory capabilities. After entering the antigen-experienced pool, memory T-cells circulate through peripheral lymphatics and accumulate in secondary lymphoid tissues [9] where they can be re-stimulated by their cognate antigen, whereas naïve T-cells continue to circulate through peripheral lymphatics awaiting initial antigenic stimulation. In addition to activation induced proliferation, T-cells are also capable of undergoing homeostatic proliferation [10,11]. This process is thought to involve weak interactions between TCR and self MHC:peptide as well as cytokines such as IL-7 [10,12,13].
To elucidate the relationship between cell division and the activation and homeostasis of human naïve and antigen-experienced T-cells, we monitored expression of CD11b, CD25, CD27, CD28, CD45RA, CD45RO, CD57, CD62L, CD69, CD95, CD134, CD152, IFN-γ, TNF-α, and IL-2 after mitogenic or cytokine stimulation of CFSE labelled naïve and antigen-experienced T-cells. These molecules were chosen as they have been used in the past to differentiate naïve, memory, and effector type T-cells [14]. We have defined functional and phenotypic changes that occur to CD4+ and CD8+ human naïve and antigen-experienced T-cells upon stimulation, as well as the temporal relationship of the expression patterns of these molecules with respect to cell division.
Materials and methods
Mononuclear cell preparations
Mononuclear cells were separated using Ficoll-Hypaque density gradient centrifugation. Naïve T-cells were obtained from human umbilical cord blood. The T-cell population was highly enriched (>97%) for naïve T-cells in cord blood as determined by staining for CD45RA or CD45RO, and CD27.
Antigen-experienced T-cells were obtained from venous blood of healthy adults. Antigen-experienced cells were purified by incubating cells with a FITC-conjugated antibody to CD45RA, then depleted by MACS sorting using anti-FITC magnetic microbeads (Miltenyi, Auburn, CA, USA). Following purification, T-cells were highly enriched (>99%) for CD45RO as determined by flow cytometry.
All blood samples were collected according to procedures approved by the internal review board and all subjects, or adult guardians of subjects, granted informed consent prior to sample acquisition.
At least two (usually three) different sources of umbilical cord blood or adult peripheral blood were used for each experiment to ensure described trends are not donor dependent.
T-cell stimulation
Cells were cultured at a concentration of 1 × 106 per ml in RPMI media (Gibco BRL, Rockville, MD, USA) supplemented with 10% heat inactivated fetal calf serum. Phytohemmagglutinin (PHA) (Sigma, St. Louis, MO, USA) was used at a concentration of 5 µg/ml, staphylococcal enterotoxin B (SEB) (Sigma) was used at 10 µg/ml, and IL-7 (R & D Systems, Minneapolis, MN, USA) was used at 5 ng/ml. Stimulator to responder cell ratios of 1 : 10 were used for the mixed lymphocyte reaction (MLR). Stimulator cells were irradiated with 3000 rads prior to addition into the MLR. Cells activated with PHA and SEB were incubated for 96 h, cells activated with MLR were cultured for 144 h, and cells driven to proliferate with IL-7 were incubated for 168 h. Experiments involving blockage of TCR:MHC interactions were accomplished using monoclonal antibodies against HLA A,B,C,DP,DQ,DR at 10 µg/ml (BD/Pharmingin, San Diego, CA, USA) as determined by titration experiments. Experiments involving depletion of IL-2 were accomplished using an IL-2 depleting antibody (BD/Pharmingin) at 10 µg/ml as determined by titration experiments.
CFSE labelling
Cells were suspended in phosphate buffered saline at a concentration of 1 × 106/ml. CFSE (Molecular Probes, Eugene, OR) was added to a final concentration of 0·25 µm. Cells were incubated in 0·25 µm CFSE in a 37°C water bath for 7 min. CFSE labelled cells were washed three times with PBS supplemented with 10% FCS prior to use.
Flow cytometry
Cells were stained for CD4 PerCP or CD8 PerCP (Becton Dickinson/Pharmingen), CD45RA APC (Caltag, Burlingame, CA, USA) or CD45RO APC (BD/Pharmingen), and one of either CD11b PE, CD25 PE (Caltag), CD27 PE, CD28 PE, CD152 PE, CD134 PE, CD62L PE, CD69 PE, CD95 PE (BD/Pharmingen), or CD57 PE (Beckman Coulter, Miami, FL, USA). Data acquisition was performed with a FACS Calibur (BD/Pharmingen) and data was analysed using Paint-A-Gate software (BD). Due to variability in CFSE loading compensation settings were adjusted for each experiment. As such, the mean fluorescent intensities (MFI) of cell surface molecules can only be compared within each experiment.
For detection of intracellular cytokines after the indicated culture periods cells were collected, washed, and re-stimulated phorbol myristial acetate (PMA) (Sigma) at 0·5 µg/ml, ionomycin (Sigma) at 10 µg/ml, and brefeldin A (Sigma) at 1 µg/ml for five hours as previously described [15]. Staining was then performed using CD4 or CD8 PerCP, CD45RA or CD45RO APC, and fix/permeablized followed by staining with one of IFN-γ PE, TNF-α PE, or IL-2 PE (all BD/Pharmingen) as described elsewhere [16].
Results
Naïve T-cells
In order to define the relationship between cell division and modulation of surface proteins or cytokine expression, we monitored the expression of a panel of surface markers and cytokines in both CD4+ and CD8+ T-cells activated by PHA, SEB, or MLR (Fig. 1). Four distinct patterns of differential expression with respect to cell division were noted: some makers were present before activation and maintained expression thereafter, others were present before activation and were down-modulated with cell division, yet others were not expressed on resting cells but were up-regulated after stimulation. Of these, some were more highly expressed with successive rounds of division while some became highly expressed before cell division and maintained thereafter. Moreover, some surface markers exhibited expression that was specific for the stimulation method employed. All markers stained are summarized in Table 1.
Fig. 1.

Cell division of stimulated naïve T-cells precedes effector function. Cord blood PBMC were stained with 0·25 µm CFSE and stimulated with PHA or SEB for 4 days, or were stimulated by MLR for 6 days. Panels demonstrate surface marker expression and cytokine production with respect to cell division for CD4+ or CD8+ gated naïve T-cells.
Table 1.
Surface markers and cytokines expressed by naïve and memory T-cells
| Naïve T-cells | Memory T-cells | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Resting | PHA Activated | SEB Activated | MLR Activated | Resting | PHA Activated | SEB Activated | MLR Activated | |||||||
| expression | expression | corr. w/ cell div. | expression | corr. w/ cell div. | expression | corr. w/ cell div. | expression | expression | corr. w/cell div. | expression | corr. w/ cell div. | expression | corr. w/ cell div. | |
| CD11b | – | – | – | – | – | + | ↑ | – | + | – | + | – | + | – |
| CD25 | – | + | ∼, ↑ | + | – | + | – | – | + | – | + | – | + | – |
| CD27 | + | + | ∼, ↓ | + | – | + | – | +, – | + | – | + | – | + | – |
| CD28 | + | + | ∼, ↓ | + | ∼, ↓ | + | – | +, – | + | – | + | – | + | – |
| CD45RA | + | + | ↓ | + | ↓ | + | ↓ | – | – | – | – | – | – | – |
| CD45RO | – | + | ∼, ↑ | + | – | + | – | + | + | – | + | – | + | – |
| CD57 | – | – | – | – | – | – | – | +, – | – | – | – | – | – | – |
| CD62L | + | + | ∼, ↓ | + | ∼, ↓ | + | – | +, − | + | – | + | – | + | – |
| CD69 | – | + | ↓ | + | ↓ | + | – | – | + | ↓ | + | ↓ | + | – |
| CD95 | – | + | – | + | – | + | – | – | + | – | + | – | + | – |
| CD134 | – | + | ↑ | + | ↑ | + | ↑ | – | + | – | + | – | + | – |
| CD152 | – | – | – | – | – | + | ↑ | – | + | – | + | – | + | – |
| IFN-γ | – | +, − | ↑ | + | ↑ | + | ↑ | – | + | – | + | – | + | – |
| IL-2 | – | + | ∼, ↑ | + | ∼, ↑ | + | ∼, ↑ | – | + | – | + | – | + | – |
| TNF-α | – | + | – | + | – | + | – | – | + | – | + | – | + | – |
+indicates presence, – indicates absence, ±indicates variation between CD4 and CD8+ T-cells, ∼ indicates moderate cell division dependence, +, – indicates expression by some CD4+ and CD8+ T-cells
indicates up-regulation with respect to cell division
indicates down-regulation with respect to cell division, corr. w/cell div. indicates correlation with cell division.
Cell surface molecules which had little dependence on cell division for their expression were CD25, CD28, CD45RO, and CD57. CD25 and CD45RO became expressed on activated cells prior to cell division and maintained expression thereafter, although the rapidity of CD25 and CD45RO expression was dependent on the method of stimulation. This phenomenon was especially evident in the CD8+ T-cells. SEB or MLR stimulation resulted in more rapid expression of CD25 and CD45RO than did PHA stimulation. CD57 did not become expressed when naïve T-cells were activated using any of these three stimuli.
Expression of CD45RA and CD69 decreased with respect to cell division. Expression of CD62L also decreased with cell division, but not as markedly as CD45RA and CD69, and was more dramatic in the CD8+ T-cell populations. CD45RA and CD69 were both down-modulated in a cell division dependent fashion, i.e. mitotic dilution, after stimulation. These data were confirmed by plotting mean fluorescent intensity (MFI) values with respect to cell division (data not shown).
Only one marker, CD134 (Ox-40), was up-regulated in a cell division dependent fashion for all three types of activation. Expression of CD134 on CD4+ T-cells reached maximum expression after two rounds of division and remained relatively constant thereafter (Fig. 1). The MFI of each division peak and the percentage of CD134 positive cells in each division peak showed that two rounds of division were required for CD134 to become expressed by more than 60% of the CD4+ T-cells, and that more than 60% of the CD4+ T-cells continued to express CD134 after two cell divisions (data not shown). CD134 did not become substantially expressed (<20%) on CD8+ T-cells. Expression of CD152 (CTLA-4), was only observed in MLR stimulation and was cell division dependent, in that three rounds of cell division were required for detection of cell surface CD152.
We also examined the relationship between cell division and cytokine secretion by activated naïve T-cells (Fig. 1). T-cells stimulated with SEB or MLR produced high levels of IFN-γ, while PHA stimulated T-cells produced little or no IFN-γ. Furthermore, regardless of stimulation, at least twice as many CD8+ T-cells made IFN-γ compared to CD4+ T-cells. We also found that undivided cells produced little IFN-γ, and that IFN-γ production by T-cells increased with cell division. IL-2 production was seen predominantly in CD4+ T-cells regardless of which stimulation method was employed. IL-2 production correlated with cell division, in that a higher percentage of dividing cells produced IL-2. TNF-α was produced by both subsets of T-cells independent of both cell division and stimulation method.
Antigen-experienced T-cells
Recently, a number of different nomenclatures have arisen to define different subpopulations of T-cells that have, at one time, been stimulated by antigen. Here, we use the term ‘antigen-experienced’ when we refer to PBMC taken from healthy adults that have been depleted of CD45RA+ cells. We use the term ‘memory’ when we refer to any T-cell that is not naïve. In order to compare resting, CD45RO+, antigen-experienced T-cell surface marker expression with that of naïve T-cells, purified antigen-experienced T-cells were activated with PHA, SEB, or by MLR and stained with the same panel of all surface markers and cytokines (Fig. 2). Compared to naïve T-cells, antigen-experienced T-cells exhibited substantially less dependence on cell division for surface marker modulation. Minor differences in cell division were observed between the antigen-experienced and naive T-cells after stimulation with PHA. This was consistent when using isolated antigen-experienced cells from five other healthy adult donors (data not shown). A smaller percentage (<5%) of the antigen-experienced T-cells also responded in the MLR compared to 20–30% of responding naïve T-cells. This was also reproducible using target cells and antigen-experienced effector cells from four different donors (data not shown).
Fig. 2.

.Stimulated antigen-experienced T-cells gain effector function without cell division. PBMC from healthy adult donors depleted of CD45RA+ T-cells, were stained with 0·25 µm CFSE and were then stimulated for 4 days with PHA or SEB, or for 6 days by MLR. Shown is a dot plot panel of surface markers and cytokine production for antigen-experienced CD4+ or CD8+ gated T-cells.
Cell surface molecules whose expression on antigen-experienced cells did not correlate with cell division were CD25, CD27, CD45RA, CD45RO, CD62L, and CD134. Antigen-experienced T-cells appeared to maintain expression of CD25, CD28, and CD45RO independent of cell division. Expression of CD25 or CD45RO by antigen-experienced T-cells did not significantly change after one round of division for CD4+ or CD8+ T-cells regardless of how the cells were stimulated. CD62L expression on antigen-experienced T-cells, whether initially CD62L positive or negative, did not appear to change after in vitro stimulation, summarized in Table 1.
Recent studies have shown that resting, memory CD8+ T-cells could re-express CD45RA after being re-stimulated [14]. We found that CD45RA expression did not increase in antigen-experienced CD4+ or CD8+ T-cells after in vitro stimulation. There were few CD45RA bright cells (< 5%) in the undivided subsets of PHA and MLR activated antigen-experienced cells, of these most expressed CD57 and did not divide in response to stimulation.
In contrast to naïve T-cells, CD134 expression in CD4+ antigen-experienced T-cells did not correlate with cell division. Naïve CD4+ T-cells expressed CD134 only after stimulation, and not until two rounds of replication were completed. However, resting CD4+ antigen-experienced T-cells expressed CD134 and maintained expression of CD134 after stimulation.
Expression of CD152 in antigen-experienced T-cells also differed from that in naïve T-cells, as CD152 expression was not limited to MLR stimulation. Figure 2 shows that CD152 was expressed when T-cells were activated with PHA, SEB, and by MLR and that this expression correlated with cell division. This was especially evident in SEB activated CD4+ T-cells, where expression levels of CD152 increased with respect to cell division.
Unlike naïve T-cells, the ability of antigen-experienced T-cells to produce cytokine after stimulation was not related to cell division. Figure 2 shows the cytokine production by antigen-experienced T-cells activated with PHA, SEB, or by MLR. Antigen-experienced T-cells produced IFN-γ, IL-2, and TNF-α in response to stimulation with SEB and PHA, prior to the first cell division. Cells that underwent division were able to produce these cytokines regardless of the stimulation method. As in naïve T-cells, IL-2 was preferentially produced by antigen-experienced CD4+ T-cells and had little, if any, cell division dependency. TNF-α was produced by both CD4+ and CD8+ antigen-experienced T-cells and was similarly not dependent on cell division.
IL-7 induced proliferation
In order to examine closely the effect of IL-7 proliferation and surface marker modulation by naïve T-cells, we cultured umbilical cord blood PBMC with IL-7 and monitored surface marker modulation with respect to cell division (Fig. 3). As can be seen, CD4+ and CD8+ T-cells that underwent one or two rounds of division maintained expression of CD45RA without significant expression of CD45RO or CD25. Interestingly, after two cell divisions, the T-cells were all CD45RO+.
Fig. 3.
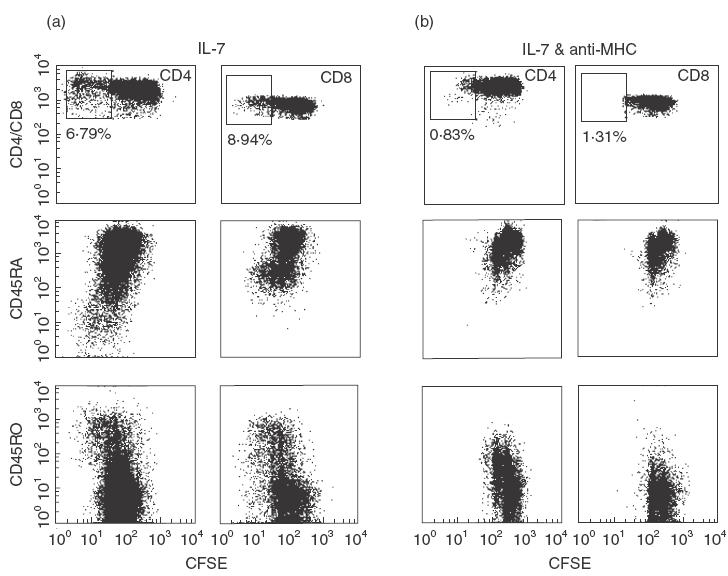
IL-7 drives proliferation of naïve T-cells in MHC independent fashion. Cord blood PBMC were stained with 0·25 µm CFSE and were then cultured for 9 days in 10 ng/ml IL-7 with isotypic control antibodies (a) or 10 µg/ml blocking antibodies recognizing HLA A, B, C, DR, DP, and DQ (b) Dot plots of CD4+ or CD8+ gated T-cells show expression patterns of CD45RA, and CD45RO with respect to cell division. Boxed areas of CD4 and CD8 plots represent the percentage of cultured cells that had proliferated more than three times for IL-7 alone (a) and IL-7+ anti-MHC antibodies (b).
In order to determine whether IL-7 is responsible for the differentiation of naïve T-cells into memory T-cells during the in vitro division assay, or whether it simply drives proliferation of pre-existing memory T-cells, we depleted the few antigen-experienced T-cells present in umbilical cord blood mononuclear cells and cultured the remaining cells with IL-7. No memory T-cells were seen after IL-7 induced proliferation, furthermore no T-cells had divided more than two times in the eight day incubation period (data not shown).
Interestingly, the majority of the T-cells observed after IL-7 induced proliferation were CD4+ T-cells. Normal CD4 : CD8 ratios in cord blood mononuclear cells are between 1 : 1 and 3 : 1. After culturing in IL-7 this ratio consistently increased. This preferential expansion of CD4+ T-cells in response to IL-7 may be attributed to higher expression levels of CD127 by CD4+ T-cells (data not shown).
In order to address the role of MHC:TCR interactions during IL-7 induced proliferation, we blocked these interactions using antibodies to MHC I and II (Fig. 3). CD4+ and CD8+ naïve T-cells were able to divide in the presence of either antibody, but memory T-cells were not able to divide when TCR interactions were blocked.
Expression of CD25 by memory T-cells that had divided more than two times when cultured with IL-7 suggested that IL-2 was critical to division of these cells. We therefore blocked IL-2 activity with an IL-2 depleting antibody (pretitred) in cultures supplemented with IL-7 (Fig. 4). When IL-2 was blocked in these cultures, no more than two cell divisions occurred, and no CD45RO/CD25+ T-cells were observed, regardless of whether the cord blood samples were CD45RO+ T-cell depleted. Naïve T-cells proliferating in IL-2 depleted cultures also failed to produce IFN-γ after PMA/ionomycin restimulation while T-cells cultured without IL-2 depleting antibody produced the cytokine after two rounds of cell division (Fig. 4).
Fig. 4.
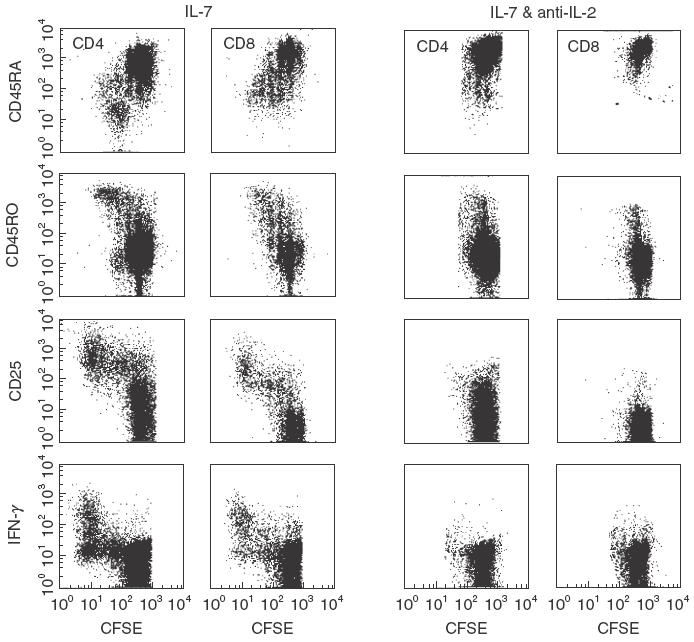
IL-2 is Required for IL-7 induced expansion of memory T-Cells present in umbilical cord blood. Cord blood PBMC were stained with 0·25 µm CFSE and were then cultured for 9 days in 10 ng/ml IL-7 with or without 10 µg/ml depleting antibodies to IL-2. After culturing, PBMC were stained with CD4 and CD45RO or CD45RA, and CD8 and CD45RA or CD45RO. Dot plots of CD4+ or CD8+ T-cells show expression patterns of CD45RA, CD45RO, CD25, and IFN-γ with respect to cell division.
Discussion
Much research has focused on correlations between T-cell effector function and phenotype [14,17–23]. It is difficult, however, to draw definitive conclusions from such data without a full understanding of the fine details of activation-induced surface marker expression and cytokine production. Therefore, our study sought to define and correlate these relationships with respect to cell division in naïve and antigen-experienced T-cells after stimulation with SEB, PHA, MLR, and in response to the cytokine IL-7.
Our data indicate that cell division is associated with both up- and down-regulation of certain surface molecules and cytokines following stimulation. This modulation can be different when comparing naïve and antigen-experienced T-cells. Here, we have demonstrated that mitotic dilution of certain cell surface markers, such as CD45RA and CD69 occurs during cell division. Additionally, other cell surface markers are up-regulated in a cell division-dependent fashion and this regulation differs between naïve and antigen-experienced T-cells. For example, in order to achieve a strong immune response to antigen, a surface molecule whose signals lead to decreased proliferation might be expected to have different regulation on naïve and antigen-experienced T-cells. Thus, we find that CD152, the ligation of which leads to proliferation inhibition, is expressed only after several divisions in the stimulated naïve T-cells, to allow for establishment of the precursor antigen-experienced pool. Conversely, CD152 expression in antigen-experienced T-cells occurs much faster, presumably to prevent large scale proliferation of the antigen-experienced T-cell population. Thus earlier expression would not hinder the immune response because of the higher antigen-experienced T-cell precursor frequency. Proteins involved in effector function may also be differentially regulated with respect to cell division in naïve and antigen-experienced T-cells. Initial expansion of activated naïve T-cells may be more important to the overall immune response than acquiring effector function. Only after initial expansion might it be important for activated naïve T-cells to acquire effector function, after which some differentiate into resting, antigen-experienced T-cells. Antigen-experienced T-cells, on the other hand, should be able to rapidly produce effector cytokine, prior to proliferation. Here, we measured cytokine production as a marker of effector function. Production of IFN-γ and IL-2 by activated naïve T-cells was dependent on cell division as shown in Fig. 1. Antigen-experienced T-cells were able to produce these cytokines rapidly, without cell division. This phenomenon is not restricted to naïve T-cells obtained from cord blood cells and naïve T-cells isolated from healthy adult donors show the same trend (data not shown). Interestingly TNF-α production occurred without naïve T-cell division, also displayed in Fig. 1, possibly due to the genetic location of the TNF-α gene within the MHC gene cluster.
In addition to naïve T-cell dependence on cell division for cytokine production other molecules associated with cytokine production should follow similar trends. CD134 is a TNF family member that has recently gained prominence as a costimulatory molecule [24]. Interactions between CD134 and its ligand are thought to occur after initial activation, and this prolongs expansion and enhances effector cytokine secretion [25]. We found that CD134 was expressed on naïve T-cells only after initial cell division, and its expression increased with subsequent rounds of cell division, directly correlating with the production of IFN-γ in CD4+ T-cells. Mechanisms responsible for the observed relationship between cell division and protein expression are not known but likely include the presence of specific transcriptional activators, absence of transcriptional repressors, and/or chromatin structure differences between naïve and memory T-cells.
The dependence upon cell division of cytokine production in naïve T-cells has also been observed in TCR transgenic mice [26,27]. However, other studies in mice have shown that small percentages of undivided naïve T-cells are capable of IFN-γ production, suggesting that cell division may not be required for cytokine production by naïve T-cells [28]. Our data suggests that cell division and activation are required for cytokine production by human naïve T-cells. These differences may be attributed to inherent differences between mouse and human T-cells, or may be due to some variations between experimental design.
The use of different stimulation methods allowed us to observe subtle differences in expression patterns of certain surface markers that may be attributed to different thresholds of activation [23], or to different mechanisms of stimulation. The MLR arises from a direct interaction between TCR, MHC, and peptide, whereas SEB stimulates both CD4+ and CD8+ T-cells by binding to specific TCR β-chains in a peptide independent fashion [29,30], while PHA bypasses the TCR entirely by binding to CD2 [31]. Interestingly, compared to naïve T-cells, a smaller percentage of antigen-experienced T-cells were responsive in the MLR reaction. This result would be expected, since any allo-specific T-cells in the subjects would not be expected to have undergone the naïve to memory transition. The antigen-experienced T-cells would be expected to be low, since they are outgrowths of specific populations specific for certain (non-MLR) antigen. Activation with PHA, SEB, and by MLR resulted in different cell surface molecule expression patterns which were more prevalent within the naïve T-cell pool. This suggests that naïve T-cell responses are neither uniform nor monolithic in nature, i.e. naïve T-cells can respond to different stimuli in different ways [32]. Antigen-experienced T-cells are already programmed to respond to antigen and, any secondary response, even to different stimuli, should mirror the initial antigen encounter.
Although antigenic stimulation results in the expansion of naïve T-cells and their differentiation into memory T-cells, there is evidence that homeostatic mechanisms operate to maintain naïve T-cell numbers in the periphery. These mechanisms appear to involve weak interactions via the TCR, are MHC dependent [12,33–35] and possibly cytokine driven [10]. We showed that IL-7 drove proliferation of naïve T-cells while maintaining their naïve phenotype, as previously shown, implying a role for IL-7 in homeostasis [13,36]. Additionally, we observed that CD4+ T-cells proliferating in the presence of IL-7 produce IL-2, even though it is unnecessary for their proliferation. Memory T-cells present in umbilical cord blood also proliferate in response to IL-7, but in contrast to naïve T-cells, their proliferation appeared to be dependent on IL-2. The proliferation rate for memory T-cells (responding to IL-7 in an IL-2 dependent manner) was more rapid than for naïve T-cells (responding to IL-7 alone).
Even though this proliferation is cytokine driven, it may be dependent on weak interactions between TCR:MHC-self peptide [12,33–35]. We found that IL-7 still drove the proliferation of naïve T-cells even when MHC molecules were blocked with monoclonal antibodies. The few memory T-cells present in umbilical cord blood, however, were hindered in their ability to divide in the presence of these antibodies. This suggests that in humans, as opposed to mice, homeostatic proliferation of naïve T-cells is independent of interactions between MHC and TCR. This idea has also been suggested based on data from humans containing mutations in TCR signalling molecules [37]. In addition, the inability of naïve T-cells proliferating in response to IL-7 to produce IFN-γ after proliferation implies that activation induced, and not homeostatic induced proliferation, enable T-cells to achieve effector function.
The fact that naïve T-cells proliferate in the presence of exogenous IL-7 may have special relevance to therapeutic use. Administration of thymopoetic cytokines such as IL-7 has been suggested as a therapeutic agent to increase immune reconstitution [38,39]. Out data suggests that not only can IL-7 be used to increase thymic output, but it could possibly be used to expand peripheral naïve T-cells [40,41]. As these T-cells proliferate with IL-7 administration, antigen-experienced T-cells may also proliferate, thus increasing the number of both naïve and memory T-cells.
Acknowledgments
This work was supported by grants AI47603 and AI35522 to RAK, AR45293 to LSD from the National Institutes of Health and Leukaemia Lymphoma Society of America translational research grant 6540–00, and amfAR grant #02680–28-RGV to D.C.D.
REFERENCES
- 1.Hintzen RQ, de Jong R, Lens SM, Brouwer M, Baars P, van Lier RA. Regulation of CD27 expression on subsets of mature T-lymphocytes. J Immunol. 1993;151:2426–35. [PubMed] [Google Scholar]
- 2.Michie CA, McLean A, Alcock C, Beverley PC. Lifespan of human lymphocyte subsets defined by CD45 isoforms. Nature. 1992;360:264–5. doi: 10.1038/360264a0. [DOI] [PubMed] [Google Scholar]
- 3.Mueller YM, De Rosa SC, Hutton JA, Witek J, Roederer M, Altman JD, Katsikis PD. Increased CD95/Fas-Induced Apoptosis of HIV-Specific CD8 (+) T Cells. Immunity. 2001;15:871–82. doi: 10.1016/s1074-7613(01)00246-1. [DOI] [PubMed] [Google Scholar]
- 4.Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–12. doi: 10.1038/44385. [DOI] [PubMed] [Google Scholar]
- 5.Dubois PM, Pihlgren M, Tomkowiak M, Van Mechelen M, Marvel J. Tolerant CD8 T cells induced by multiple injections of peptide antigen show impaired TCR signaling and altered proliferative responses in vitro and in vivo. J Immunol. 1998;161:5260–7. [PubMed] [Google Scholar]
- 6.McHeyzer-Williams MG, Altman JD, Davis MM. Enumeration and characterization of memory cells in the TH compartment. Immunol Rev. 1996;150:5–21. doi: 10.1111/j.1600-065x.1996.tb00693.x. [DOI] [PubMed] [Google Scholar]
- 7.McHeyzer-Williams MG, Davis MM. Antigen-specific development of primary and memory T cells in vivo. Science. 1995;268:106–11. doi: 10.1126/science.7535476. [DOI] [PubMed] [Google Scholar]
- 8.Murali-Krishna K, Lau LL, Sambhara S, Lemonnier F, Altman J, Ahmed R. Persistence of memory CD8 T cells in MHC class I-deficient mice. Science. 1999;286:1377–81. doi: 10.1126/science.286.5443.1377. [DOI] [PubMed] [Google Scholar]
- 9.Swain SL, Croft M, Dubey C, Haynes L, Rogers P, Zhang X, Bradley LM. From naive to memory T cells. Immunol Rev. 1996;150:143–67. doi: 10.1111/j.1600-065x.1996.tb00700.x. [DOI] [PubMed] [Google Scholar]
- 10.Ku CC, Murakami M, Sakamoto A, Kappler J, Marrack P. Control of homeostasis of CD8+ memory T cells by opposing cytokines. Science. 2000;288:675–8. doi: 10.1126/science.288.5466.675. [DOI] [PubMed] [Google Scholar]
- 11.Marrack P, Bender J, Hildeman D, et al. Homeostasis of alpha beta TCR+ T cells. Nat Immunol. 2000;1:107–11. doi: 10.1038/77778. [DOI] [PubMed] [Google Scholar]
- 12.Goldrath AW, Bevan MJ. Low-affinity ligands for the TCR drive proliferation of mature CD8+ T cells in lymphopenic hosts. Immunity. 1999;11:183–90. doi: 10.1016/s1074-7613(00)80093-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Soares MV, Borthwick NJ, Maini MK, Janossy G, Salmon M, Akbar AN. IL-7-dependent extrathymic expansion of CD45RA+ T cells enables preservation of a naive repertoire. J Immunol. 1998;161:5909–17. [PubMed] [Google Scholar]
- 14.Hamann D, Baars PA, Rep MH, Hooibrink B, Kerkhof-Garde SR, Klein MR, van Lier RA. Phenotypic and functional separation of memory and effector human CD8+ T cells. J Exp Med. 1997;186:1407–18. doi: 10.1084/jem.186.9.1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jason J, Larned J. Single-cell cytokine profiles in normal humans. comparison of flow cytometric reagents and stimulation protocols. J Immunol Meth. 1997;207:13–22. doi: 10.1016/s0022-1759(97)00079-3. [DOI] [PubMed] [Google Scholar]
- 16.Pitcher CJ, Quittner C, Peterson DM, Connors M, Koup RA, Maino VC, Picker LJ. HIV-1-specific CD4+ T cells are detectable in most individuals with active HIV-1 infection, but decline with prolonged viral suppression. Nat Med. 1999;5:518–25. doi: 10.1038/8400. [DOI] [PubMed] [Google Scholar]
- 17.Ahmed R, Gray D. Immunological memory and protective immunity: understanding their relation. Science. 1996;272:54–60. doi: 10.1126/science.272.5258.54. [DOI] [PubMed] [Google Scholar]
- 18.Byrne JA, Butler JL, Cooper MD. Differential activation requirements for virgin and memory T cells. J Immunol. 1988;141:3249–57. [PubMed] [Google Scholar]
- 19.Cho BK, Wang C, Sugawa S, Eisen HN, Chen J. Functional differences between memory and naive CD8 T cells. Proc Natl Acad Sci USA. 1999;96:2976–81. doi: 10.1073/pnas.96.6.2976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dutton RW, Bradley LM, Swain SL. T cell memory. Ann Rev Immunol. 1998;16:201–23. doi: 10.1146/annurev.immunol.16.1.201. [DOI] [PubMed] [Google Scholar]
- 21.Dutton RW, Swain SL, Bradley LM. The generation and maintenance of memory T and B cells. Immunol Today. 1999;20:291–3. doi: 10.1016/s0167-5699(98)01415-7. [DOI] [PubMed] [Google Scholar]
- 22.Gray D. Immunological memory. Annu Rev Immunol. 1993;11:49–77. doi: 10.1146/annurev.iy.11.040193.000405. [DOI] [PubMed] [Google Scholar]
- 23.Tanchot C, Lemonnier FA, Perarnau B, Freitas AA, Rocha B. Differential requirements for survival and proliferation of CD8 naive or memory T cells. Science. 1997;276:2057–62. doi: 10.1126/science.276.5321.2057. [DOI] [PubMed] [Google Scholar]
- 24.Godfrey WR, Fagnoni FF, Harara MA, Buck D, Engleman EG. Identification of a human OX-40 ligand, a costimulator of CD4+ T cells with homology to tumor necrosis factor. J Exp Med. 1994;180:757–62. doi: 10.1084/jem.180.2.757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gramaglia I, Weinberg AD, Lemon M, Croft M. Ox-40 ligand: a potent costimulatory molecule for sustaining primary CD4 T cell responses. J Immunol. 1998;161:6510–7. [PubMed] [Google Scholar]
- 26.Bird JJ, Brown DR, Mullen AC, et al. Helper T cell differentiation is controlled by the cell cycle. Immunity. 1998;9:229–37. doi: 10.1016/s1074-7613(00)80605-6. [DOI] [PubMed] [Google Scholar]
- 27.Gett AV, Hodgkin PD. Cell division regulates the T cell cytokine repertoire, revealing a mechanism underlying immune class regulation. Proc Natl Acad Sci USA. 1998;95:9488–93. doi: 10.1073/pnas.95.16.9488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ben-Sasson SZ, Gerstel R, Hu-Li J, Paul WE. Cell division is not a ‘Clock’ measuring acquisition of competence to produce IFN-gamma or IL-4. J Immunol. 2001;166:112–20. doi: 10.4049/jimmunol.166.1.112. [DOI] [PubMed] [Google Scholar]
- 29.Coppola MA, Blackman MA. Bacterial superantigens reactivate antigen-specific CD8+ memory T cells. Int Immunol. 1997;9:1393–403. doi: 10.1093/intimm/9.9.1393. [DOI] [PubMed] [Google Scholar]
- 30.Fuller CL, Braciale VL. Selective induction of CD8+ cytotoxic T lymphocyte effector function by staphylococcus enterotoxin B. J Immunol. 1998;161:5179–86. [PubMed] [Google Scholar]
- 31.Flynn KO, Krensky AM, Beverley PC, Burakoff SJ, Linch DC. Phytohaemagglutinin activation of T cells through the sheep red blood cell receptor. Nature. 1985;313:686–7. doi: 10.1038/313686a0. [DOI] [PubMed] [Google Scholar]
- 32.Picker LJ, Singh MK, Zdraveski Z, Treer JR, Waldrop SL, Bergstresser PR, Maino VC. Direct demonstration of cytokine synthesis heterogeneity among human memory/effector T cells by flow cytometry. Blood. 1995;86:1408–19. [PubMed] [Google Scholar]
- 33.Clarke SR, Rudensky AY. Survival and homeostatic proliferation of naive peripheral CD4+ T cells in the absence of self peptide: MHC complexes. J Immunol. 2000;165:2458–64. doi: 10.4049/jimmunol.165.5.2458. [DOI] [PubMed] [Google Scholar]
- 34.Ernst B, Lee DS, Chang JM, Sprent J, Surh CD. The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity. 1999;11:173–81. doi: 10.1016/s1074-7613(00)80092-8. [DOI] [PubMed] [Google Scholar]
- 35.Viret C, Wong FS, Janeway CA., Jr Designing and maintaining the mature TCR repertoire: the continuum of self-peptide: self-MHC complex recognition. Immunity. 1999;10:559–68. doi: 10.1016/s1074-7613(00)80055-2. [DOI] [PubMed] [Google Scholar]
- 36.Hassan J, Reen DJ. IL-7 promotes the survival and maturation but not differentiation of human post-thymic CD4+ T cells. Eur J Immunol. 1998;28:3057–65. doi: 10.1002/(SICI)1521-4141(199810)28:10<3057::AID-IMMU3057>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 37.Elder ME, Skoda-Smith S, Kadlecek TA, Wang F, Wu J, Weiss A. Distinct T cell developmental consequences in humans and mice expressing identical mutations in the DLAARN motif of ZAP-70. J Immunol. 2001;166:656–61. doi: 10.4049/jimmunol.166.1.656. [DOI] [PubMed] [Google Scholar]
- 38.Abdul-Hai A, Or R, Slavin S, Friedman G, Weiss L, Matsa D, Ben-Yehuda A. Stimulation of immune reconstitution by interleukin-7 after syngeneic bone marrow transplantation in mice. Exp Hematol. 1996;24:1416–22. [PubMed] [Google Scholar]
- 39.Bolotin E, Smogorzewska M, Smith S, Widmer M, Weinberg K. Enhancement of thymopoiesis after bone marrow transplant by in vivo interleukin-7. Blood. 1996;88:1887–94. [PubMed] [Google Scholar]
- 40.Fry TJ, Connick E, Falloon J, et al. A potential role for interleukin-7 in T-cell homeostasis. Blood. 2001;97:2983–90. doi: 10.1182/blood.v97.10.2983. [DOI] [PubMed] [Google Scholar]
- 41.Fry TJ, Mackall CL. Interleukin-7. master regulator of peripheral T-cell homeostasis? Trends Immunol. 2001;22:564–71. doi: 10.1016/s1471-4906(01)02028-2. [DOI] [PubMed] [Google Scholar]