

Abstract
Rationale:
Although reinstatement of extinguished cocaine self-administration is widely used as an animal model of relapse, it is unclear which behavioral effects of the drug stimulus (i.e., unconditioned, discriminative or reinforcing) mediate the increases in responding after extinction.
Objective:
To examine the influence of experience with cocaine as a reinforcer on the ability of response-independent cocaine injections to increase extinguished responding.
Methods:
Effects of noncontingent injections of cocaine (0.01-1.0 mg/kg, IV) were assessed in two groups of cynomolgus monkeys: monkeys with extensive histories of cocaine self-administration when responding was maintained under a concurrent fixed ratio (FR) 50 schedule of saline and food presentation (n=8), and cocaine-naïve monkeys (n=5) responding under an FR 50 schedule of food presentation. In the latter group, the effects of noncontingent cocaine and food (1 or 5 pellets) were examined before and after a brief history of cocaine (0.03 mg/kg/inj) self-administration under an FR 50 schedule.
Results:
In the cocaine-experienced subjects responding under a concurrent schedule of saline and food availability, non-contingent cocaine dose-dependently increased injection-lever responding. In the initially cocaine-naïve subjects, no dose of cocaine increased extinguished food-maintained responding before or after a brief exposure to cocaine self-administration. In contrast, non-contingent delivery of five food pellets significantly increased extinguished food-maintained responding after cocaine self-administration.
Conclusions:
These results support the view that, under self-administration conditions, the discriminative stimulus effects of cocaine play a prominent role in the ability of cocaine to increase extinguished responding.
Keywords: cocaine, extinction, reinstatement, relapse, self-administration, nonhuman primates
One of the most difficult obstacles in treating drug addiction is preventing relapse to drug use (O'Brien 1997, 2005). The most prominent preclinical animal model of relapse involves the extinction of drug-maintained responding followed by exposure to stimuli (noncontingent drug injections, drug-conditioned contextual cues or stressful stimuli) that result in an increase in responding (see Shaham et al. 2003). This “reinstatement” model has been used extensively to examine the neurobiological and behavioral mechanisms underlying relapse (Stretch et al. 1971; de Wit and Stewart 1981; Shalev et al. 2002). Although the model possesses face validity to human relapse, the construct validity of the model (i.e., the similarity of mechanisms that underlie the model and the clinical situation), remains to be firmly established (Epstein and Preston 2003; Katz and Higgins 2003; Epstein et al. 2006). For example, the ability of a noncontingent cocaine injection to reinstate extinguished responding previously maintained by cocaine is commonly interpreted as “drug seeking” or “craving,” suggesting that increases in responding result from exposure to the reinforcing stimulus (SR) effects of cocaine. However, the prominence of SR effects in reinstatement is not firmly established. Several behavioral effects of drugs may play a role in increasing extinguished responding, including discriminative stimulus (SD, a stimulus that provides information about the availability of reinforcement under prevailing conditions) and “direct” behavioral effects (i.e., stimulant effects not explicitly related to contingencies between behavioral responses and reinforcing stimuli).
Several investigators have studied the response-reinstating effects of cocaine in the absence of its SR effects by examining extinguished responding that was previously maintained by food rather than cocaine itself. These studies have supported the involvement of multiple drug effects in increasing extinguished responding. For example, data reported by Odum and Shahan (2004) highlighted the importance of SD effects in reinstatement. In those studies, rodents responded under a variable-interval schedule of food presentation; amphetamine was administered either before or after the behavioral session in separate groups of subjects. Thus, in the group that received amphetamine prior to the session, the interoceptive (internal stimulus) effects of amphetamine served as an SD for the availability of food. During extinction, amphetamine significantly increased lever pressing in the subjects in whom amphetamine was administered before, but not after, food-reinforced training sessions. Recent studies have extended this observation to cocaine (Cador et al. 2006). Results of the Odum and Shahan (2004) study additionally supported a role for direct effects of amphetamine on responding through the observation that amphetamine slightly but significantly increased responding in rats who had received only saline during training. Previous studies have similarly demonstrated that noncontingent drug administration can increase extinguished food-reinforced responding (Skinner and Heron 1937; but see de Wit and Stewart 1981). Taken together these studies suggest that drug effects other than SR effects can play a role in reinstating extinguished behavior.
As noted above, previous studies isolated the influence of SD versus SR effects of drugs by scheduling food as the reinforcer rather than drug (Odum and Shahan 2004; Cador et al. 2006). In an earlier study, Franks and Lattal (1976) found that the ability of non-contingent food presentation to reinstate extinguished food-reinforced responding was influenced by the schedule of reinforcement prior to extinction. That is, food presentation during priming sessions was serving as an SD that influenced the frequency of responding during reinstatement. The present study was explicitly designed to examine the importance of the SR effects of cocaine in reinstatement by studying the ability of noncontingent cocaine injections to reinstate extinguished food-maintained responding under conditions in which monkeys had experience with the SR effects of cocaine, but when the interoceptive effects of cocaine did not serve as an SD for food availability. First, to demonstrate that cocaine could reinstate responding in cynomolgus monkeys, we examined reinstatement in monkeys with extensive experience self-administering cocaine in the context of a food, drug choice procedure (see Czoty et al. 2005). When choice was between saline and food such that monkeys allocated nearly 100% of responses to the food-associated lever (Paronis et al. 2002; Negus 2003; Czoty et al. 2005), response-independent injections of cocaine were administered. Because these subjects had extensive histories and exposure to the SD and SR effects of cocaine under this schedule, we anticipated that non-contingent injections of cocaine at the start of the session would result in a dose-dependent shift in response allocation to the injection lever, as has been reported previously in monkeys (Gasior et al. 2004). To more directly investigate the specific role of reinforcing effects in reinstatement, the ability of cocaine to increase extinguished food-reinforced responding was examined in a separate group of initially cocaine-naïve monkeys before and after a brief experience self-administering cocaine. Observation of reinstatement of extinguished food-reinforced responding prior to experience with cocaine as a reinforcer would clearly indicate a minimal role for SD and SR effects in reinstatement, suggesting instead the involvement of direct effects of cocaine on responding. Observing reinstatement only after self-administration experience would suggest that SR effects, acquired by cocaine during self-administration, played a prominent role in reinstatement. Finally, observation that reinstatement did not occur before or after monkeys acquired a cocaine self-administration history would suggest an importance of SD rather than SR effects. The ability of response-independent delivery of food to reinstate extinguished food-maintained responding was also examined before and after cocaine self-administration.
METHODS
Subjects
Thirteen adult male cynomolgus monkeys (Macaca fascicularis) served as subjects. Five of the monkeys were drug and experimentally-naïve at the outset of these studies and lived individually in stainless steel cages. The remaining 8 monkeys had extensive histories of cocaine self-administration and exposure to D1 agonists (Czoty et al. 2004, 2005), and were socially housed. Although monkeys occupied both dominant and subordinate ranks in the social hierarchy, no influence of social rank was observed in this experiment; thus, data are collapsed across ranks. All monkeys were fitted with aluminum collars (Primate Products, Redwood City, Calif., USA) and trained to approach the front of the cage to be guided into a restraint chair (Primate Products) using a specially designed stainless steel pole (Primate Products). Monkeys' were weighed weekly and fed enough food daily (Purina Primate Chow and fresh fruit) to maintain at least 95% of free-feeding weight; water was continuously available in the home cage. All procedures were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Animal Care and Use Committee of Wake Forest University. Environmental enrichment was provided as outlined in the Animal Care and Use Committee of Wake Forest University Nonhuman Primate Environmental Enrichment Plan.
Apparatus
The apparatus consisted of a ventilated, sound-attenuating experimental chamber (1.5 × 0.74 × 0.76 m; Med Associates, East Fairfield, VT). Two retractable response levers (5 cm wide) were located on one side of the chamber with a horizontal row of three stimulus lights 14 cm above each lever and a food receptacle located between the levers. The receptacle was connected with a Tygon tube to a pellet dispenser (Gerbrands Corp., Arlington, MA) located on the top of the chamber for delivery of 1-g banana-flavored food pellets (Bio-Serv, Frenchtown, NJ). An infusion pump (Cole-Parmer, Inc., Chicago, IL) was located on the top of the chamber. Experimental sessions were conducted 5-7 days/week while monkeys were seated in a primate restraint chair within the operant conditioning chamber.
Catheter Implantation
All subjects were surgically prepared with a chronic indwelling venous catheter into a major vein (femoral, internal or external jugular or brachial) and subcutaneous vascular access port (Access Technologies, Skokie, Ill., USA) under sterile conditions as previously described (Czoty et al. 2004). To prolong patency, each port and catheter was flushed with a solution of heparinized saline (100 U/ml) each day.
Procedure
Experiment 1: Effects of noncontingent cocaine on response allocation during concurrent availability of saline and food
Subjects (n=8) in this experiment had extensive experience self-administering cocaine under a concurrent FR 50 schedule of cocaine and food presentation described previously (Czoty et al. 2005). Briefly, 50 consecutive responses on one lever produced an injection of cocaine; completion of an FR 50 on the other lever produced a food pellet. Responses emitted on the alternate lever before completion of an FR 50 reset the response requirement. Sessions ended after 30 total reinforcers had been earned or 60 min had elapsed, whichever came first. Under this schedule, response-contingent cocaine (0.003-0.1 mg/kg) presentation resulted in dose-dependent increases in the percentage of responses emitted on the injection lever (Czoty et al. 2005). In the present studies, saline was substituted for a dose of cocaine that resulted in >90% injection-lever responding (0.03 or 0.1 mg/kg per injection). When response allocation stabilized, such that < 5% of the total responses were allocated to the injection lever for three consecutive days, a noncontingent injection of cocaine (0.01-0.56 mg/kg, i.v.) was administered immediately before the session. Each dose was administered in mixed order across monkeys and the entire cocaine dose-response curve was determined under the saline, food choice conditions before re-establishing cocaine self-administration.
Experiment 2: Effects of cocaine self-administration on reinstatement of extinguished food-reinforced responding
Cocaine-naïve subjects (n=5) were trained to respond under an FR 50 schedule of food presentation on the right lever. Sessions began with illumination of the white light above the lever; completion of 50 responses resulted in the white light being extinguished, illumination of the red light for 10 sec and delivery of a food pellet, followed by a 10-sec timeout period in which the chamber was dark and responding had no scheduled consequences. The left lever was not extended into the chamber. Sessions lasted until 30 reinforcers had been obtained or 60 min had elapsed. Once response rates and number of reinforcers were deemed stable, the food pellet dispenser was unplugged and food-maintained responding was extinguished. Under these conditions, 50 responses still produced changes in stimulus lights but no food pellet was delivered. Responding was deemed extinguished when the number of reinforcers earned was less than 20% of those earned during baseline sessions for three consecutive sessions. Next, food pellets (1 or 5) or cocaine injections (0.03-1.0 mg/kg, IV) were administered non-contingently immediately prior to the session. It is important to note that, because catheters were flushed with heparinized saline immediately before each extinction session, data from non-cocaine priming sessions represent the effects of a priming injection of saline. Food pellets were delivered into the food receptacle via manual operation of the pellet dispenser. After a food or cocaine prime was examined during an extinction session, at least one baseline extinction session was conducted before the next prime was administered. The order of presentation of these stimuli was randomized across subjects.
When the effects of noncontingent cocaine and food had been determined, monkeys were exposed to cocaine self-administration on the opposite (left) lever in the presence of the white light; the right lever remained retracted during cocaine self-administration sessions. Cocaine- and food-reinforced responding were maintained on opposite levers so that the contingencies for the different reinforcers would be associated with topographically different responses. With the white light illuminated, responding was maintained under an FR 50 schedule of 0.03 mg/kg per injection cocaine for 11 ± 0.6 sessions (range: 10-13 sessions). The dose was chosen based on previous results within our laboratory that this is the lowest dose that maintains responding above saline (control) levels (Czoty et al., 2005) and the duration of self-administration was chosen to conservatively assure that acquisition of cocaine self-administration had occurred, which we previously established to be within 7 sessions (e.g., Nader et al., 2002). At the end of that period, food-reinforced responding was re-established on the original (right) lever while the left lever remained retracted. When food-maintained responding was stable, responding was extinguished and the effects of noncontingent food and cocaine were re-determined as described above.
Drugs
For noncontingent injections, a stock solution of cocaine HCl (National Institute on Drug Abuse, Bethesda, MD, USA) was dissolved in sterile saline to a concentration of 10-100 mg/ml. Doses (0.01-1.0 mg/kg, IV) were administered in a volume of 1.0 ml/10 kg. For cocaine self-administration, cocaine was prepared in 250 ml sterile saline and delivered at a rate of approximately 1.5 ml/10 sec.
Data Analysis
The primary dependent variables examined were % injection-lever responding when saline was the alternative to food and total stimulus presentations (food presentations and saline injections) in Experiment 1 and number of fixed-ratio requirements completed during extinction in Experiment 2. Data are represented as mean ± SEM. To determine whether cocaine or food significantly increased responding above extinction levels, separate one-way repeated-measures analyses of variance (ANOVA) were conducted for noncontingent cocaine and food before and after exposure to cocaine self-administration, followed by Dunnett's post-hoc tests when significant main effects were observed. To test whether cocaine self-administration experience significantly altered the ability of cocaine or drug administration to increase extinguished responding, data were analyzed using a two-way repeated-measures ANOVA with a post-hoc Fishers LSD test. Differences were considered significant at the 95% level of confidence (p<0.05).
RESULTS
Experiment 1: Effects of noncontingent cocaine on response allocation during concurrent availability of saline and food
When choice was between cocaine (0.1 mg/kg per injection) and food, ≥80% of responses were emitted on the injection-associated lever (see Czoty et al., 2005). When saline was substituted for cocaine, response allocation shifted to the food-associated lever within 10 sessions in all monkeys (Fig. 1 above “S”). Under these conditions, monkeys typically received the maximum number of food reinforcers (30). Noncontingent administration of cocaine before the behavioral session dose-dependently and significantly increased responding on the injection lever (F5,42=14.4, p<0.001; Fig. 1); post-hoc analysis indicated that the response distribution after administration of 0.03, 0.1, 0.3 and 0.56 mg/kg cocaine was significantly different from that observed in the absence of noncontingent cocaine. On average, monkeys received at least 29 total stimulus presentations after all primes except 0.56 mg/kg. After that dose, monkeys received 18.3 ± 10.9 total presentations.
Figure 1.
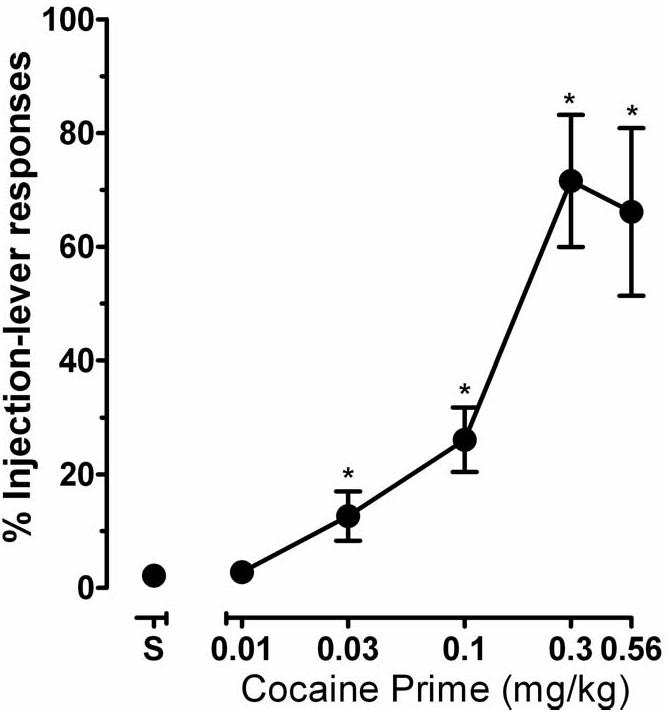
Effects of noncontingent injections of cocaine (0.01-0.56 mg/kg) on response distribution under a concurrent schedule of saline and food presentation in monkeys with extensive cocaine self-administration histories. Ordinate, percent of total responses emitted on the injection lever, Abscissa, dose of noncontingent cocaine injection. Point above “S” represent response distribution when saline was the alternative to food. Data are expressed as mean ± SEM. *, significant difference from response distribution in the absence of the response-independent cocaine (p<0.05).
Experiment 2: Effects of cocaine self-administration on reinstatement of extinguished food-reinforced responding
Food pellets reliably maintained responding in each monkey (Table 1), and subjects always earned the maximum available reinforcers. When the food pellet dispenser was unplugged, response rates decreased substantially (Table 1); monkeys completed 1.5 ± 0.8 FRs per session. Approximately 10 sessions were required to reach the extinction criteria. When administered noncontingently prior to extinction sessions, neither food nor cocaine significantly increased the number of FRs completed (Fig. 2).
Table 1.
Rates of responding§ under each experimental condition in Experiment 2.
| Subject | Food- maintained (Before SA) |
Extinction (Before SA) |
Cocaine- maintained (0.03mg/kg/inj) |
Food- maintained (After SA) |
Extinction (After SA) |
|---|---|---|---|---|---|
| C-7078 | 1.69 ± 0.38 | 0.08 ± 0.09 | 0.33 ± 0.15 | 1.89 ± 0.17 | 0.03 ± 0.02 |
| C-7079 | 4.74 ± 1.41 | 0.10 ± 0.15 | 0.51 ± 0.24 | 4.50 ± 1.05 | 0.06 ± 0.05 |
| C-7083 | 3.26 ± 1.09 | 0.04 ± 0.01 | 0.58 ± 0.18 | 4.59 ± 0.66 | 0.04 ± 0.02 |
| C-7423 | 2.12 ± 1.04 | 0.01 ± 0.01 | 0.27 ± 0.11 | 2.93 ± 0.29 | 0.10 ± 0.13 |
| C-7424 | 2.52 ± 0.07 | 0.04 ± 0.04 | 0.28 ± 0.03 | 2.57 ± 0.43 | 0.05 ± 0.02 |
| Group | 2.87 ± 0.54 | 0.05 ± 0.02 | 0.39 ± 0.14 | 3.30 ± 0.54 | 0.05 ± 0.01 |
Individual data are expressed as mean ± SD of the last three sessions under each condition. Group data are expressed as mean ± SEM.
Figure 2.
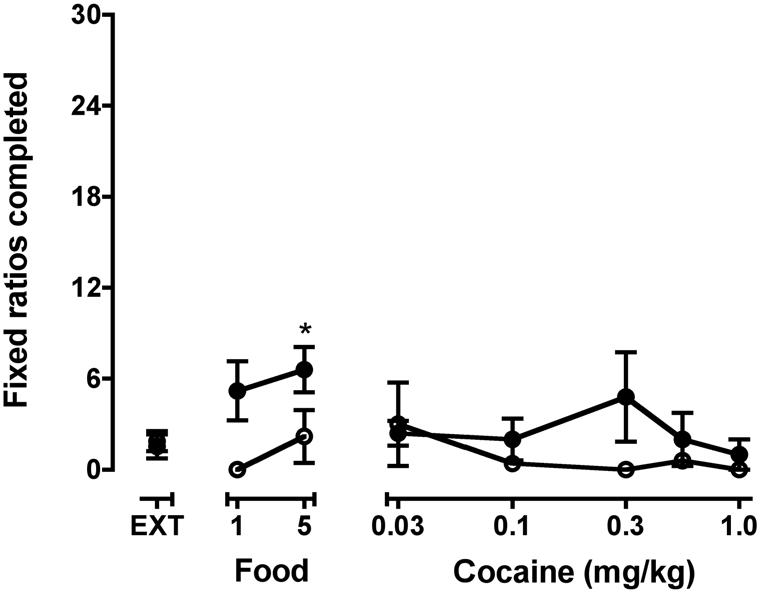
Effects of noncontingent cocaine or food on the number of fixed ratios completed during extinction before (open symbols) and after (closed symbols) a brief history of cocaine self-administration. Points above EXT are the mean (± SEM) data from last three extinction sessions. *, significantly different from EXT conditions (p<0.05).
Next, the conditions were changed to study cocaine self-administration on the other (left) lever. All subjects readily self-administered 0.03 mg/kg cocaine under an FR 50 schedule for 11.0 ± 0.6 sessions. On average, monkeys earned 12.3 ± 2.2 injections per session, with total cocaine intake over this period averaging 4.03 ± 0.7 mg/kg. Mean response rates when cocaine was available were higher (t4=5.77, p<0.01) than response rates during food extinction (Table 1). Saline substitutions were not conducted because we did not want to confound the time period after the brief self-administration history by substituting saline.
After monkeys were exposed briefly to cocaine self-administration, responding was again maintained by food presentation. Average response rates during this period and the effects of extinction on response rates and on the average number of FRs completed were nearly identical to effects observed during the earlier phase of the study (Table 1, Fig. 2). Also, resumption of food-maintained responding and number of sessions required to reach extinction were similar to those before the brief history of cocaine self-administration. When response-independent food pellets and cocaine injections were again administered before extinction sessions as was done prior to the brief period of cocaine self-administration, the number of completed FRs was significantly increased above extinction levels by food (F2,8=4.9, p<0.05) but not by cocaine. At the 0.3 mg/kg dose, there was an increase in extinguished responding, albeit not significant, with a high variability as two subjects completed 12 FRs while the other three completed 0 FRs. Using total responses instead of completed FRs as the primary dependent variable did not change the statistical outcomes (data not shown).
DISCUSSION
The ability of response-independent cocaine injections to reinstate extinguished responding in subjects who have previously self-administered cocaine is well-documented (Spealman et al. 1999; Shaham et al. 2003). In such subjects, cocaine has been experienced as a reinforcer and the interoceptive effects of cocaine have come to function as an SD signaling cocaine availability. Either of these stimulus functions could result in increases in responding when a noncontingent cocaine injection is administered during extinction. The main purpose of the present study was to examine whether experience with cocaine as a reinforcer influenced the ability of noncontingent cocaine injections to reinstate extinguished food-maintained responding under conditions in which the interoceptive effects of cocaine did not serve as an SD signaling availability of food reinforcement.
In cynomolgus monkeys with extensive experience self-administering cocaine in the context of a food, drug choice procedure, noncontingent cocaine injections increased responding on the injection-associated lever when saline was the alternative to food, as has been observed previously in rhesus monkeys (Gasior et al. 2004). The assessment of reinstatement of cocaine choice differs from traditional reinstatement procedures in which responding is maintained on a single manipulandum. Most notably, under a food/drug choice procedure, substitution of saline for cocaine results in a shift in allocation of behavior from the injection- to the food-associated lever as opposed to the decrease in responding produced by extinction in typical reinstatement procedures. This feature of choice studies may better reflect the clinical situation in that abusers may allocate their behavior to a number of potential reinforcers. Moreover, the reinforcing effects of cocaine are almost never extinguished in drug abusers. In addition, this choice procedure permits the assessment of drug effects on a baseline of ongoing behavior, rather than extinguished responding that occurs at very low rates. The process of extinction itself represents a form of learning that results in an array of neuroadaptive changes which impact the behavioral effects of cocaine (Myers and Davis 2002; Bouton et al. 2004; Self et al. 2004). Although some differences in the effects of direct dopamine receptor agonists have been observed in choice versus traditional reinstatement procedures (e.g., Self et al. 1996; Khroyan et al. 2000; Gasior et al. 2004), cocaine reliably reinstates extinguished responding previously maintained by cocaine under simple schedules (e.g., Khroyan et al. 2000) as well as under choice conditions (Gasior et al. 2004; present results). In a separate group of monkeys, noncontingent cocaine injections did not affect extinguished food-maintained responding when monkeys were cocaine-naïve or after monkeys were exposed to a brief history of cocaine self-administration. Although it is possible that response-reinstating effects of cocaine injections may have been observed after a longer period of self-administration in these monkeys, imparting SR effects to cocaine by explicitly scheduling it as a reinforcer did not enable cocaine to reinstate extinguished food-maintained responding implies that SR effects per se are not sufficient to cause a stimulus to reinstate behavior.
The present results are consistent with those of Odum and Shahan (2004) and Cador et al. (2006) which support a primary role for SD effects of amphetamine and cocaine, respectively, in reinstating extinguished responding previously maintained by food. In those studies, amphetamine and cocaine were only able to reinstate extinguished responding in animals in which amphetamine served as a SD for food availability during training. That none of the subjects in those studies had experience with amphetamine or cocaine as a SR suggests strongly that SD effects play a more prominent role than SR effects in reinstatement. In the present study, during the brief exposure to cocaine self-administration, the SD effects of cocaine predicted availability of cocaine rather than food. Thus it is not surprising that subsequent exposure to the SD effects of cocaine failed to increase extinguished responding that had been maintained by another reinforcer (in this case, food). Although increased responding could have resulted because the cocaine SD signaled cocaine availability to the subjects, this possibility was minimized by arranging conditions such that cocaine and food were self-administered on separate manipulanda. Had food- and cocaine-maintained responding occurred on the same response lever, it is likely that reinstatement would have been observed.
Data from the present study suggest that direct response rate-increasing effects of cocaine do not play a major role in reinstatement. This conclusion differs from that of Odum and Shahan (2004) who observed a significant increase in extinguished responding following amphetamine administration in drug-naïve rats. However, this effect, while statistically significant, was extremely small (from approximately 1% to approximately 3% to of pre-extinction levels). In an earlier study by Skinner and Heron (1937), the authors concluded that caffeine- and benzedrine-induced increases in extinguished food-maintained responding were due to direct effects of the drug on behavior (i.e., unconditioned effects). The discrepancy between that study and the present results is more difficult to resolve, but may reflect the use of “relatively large” doses in the earlier study. Nonetheless, it is somewhat surprising that noncontingent cocaine did not increase extinguished response rates in view of the well-established observation that psychostimulants generally increase low baseline rates of responding (Kelleher and Morse 1968). Earlier research on rate-dependent effects of amphetamine identified responding that has no programmed consequences as an exception to rate dependency (Kelleher and Morse 1968; Dews and Wenger 1977). Although responding in extinction fits this description, these authors were addressing responding that never had programmed consequences; it is unclear whether extinguished responding was considered an exception to the rate dependency of effects of stimulant drugs. Nonetheless, the present results suggest that under the present conditions, the direct rate-increasing effects of cocaine do not increase extinguished responding.
Unlike response-independent cocaine, noncontingent administration of food significantly increased extinguished responding previously maintained by food. These results extend to nonhuman primates an effect that has been observed previously in rodents (Reid 1958; Franks and Lattal, 1976). The results are consistent with the view that, during training, food acquired both SR and SD effects indicating food availability that increased responding under extinction conditions. It is unclear, however, why this effect was observed only during the second determination. It is possible that a higher number of food pellets would have reinstated responding under both conditions.
In summary, the results of the present studies reinforce the importance of SD effects in the ability of cocaine to reinstate extinguished responding, and extend the conditions under which a drug can reinstate extinguished food-maintained responding. Although these data highlight SD effects and de-emphasize SR effects of drugs, it is clear that, as is true for SR effects, the overlap between SD effects and reinstatement is not complete. Not all drugs that can reproduce the SD effects of cocaine in drug discrimination experiments are able to reinstate extinguished cocaine-maintained responding (Spealman et al. 1999; Shaham et al. 2003). A more detailed understanding of the neuropharmacological phenomena that underlie the SD, SR and response-reinstating effects of cocaine will help to unravel the distinct neurobiological mechanisms by which these behavioral effects are produced, and may provide new directions for pharmacotherapies designed to promote abstinence and prevent relapse. For example, recent findings suggest that total cocaine intake, rather than simply a history of cocaine self-administration, produce neurobiological changes (Czoty et al., in press). Such findings suggest that there are interactions between stimulus effects of cocaine (unconditioned, SD and SR) and neuropharmacology that may determine the behavioral effects of drugs.
ACKNOWLEDGEMENTS
This research was supported by National Institute on Drug Abuse grants DA-10584, F31 DA-020281, DA-12460 and DA-16455. The authors acknowledge the excellent technical assistance of Robert Gould, Michelle Icenhower, Ciara McCabe, Susan Nader and Matthew Dickens.
REFERENCES
- Bouton ME. Context and behavioral processes in extinction. Learn Mem. 2004;11:485–494. doi: 10.1101/lm.78804. [DOI] [PubMed] [Google Scholar]
- Cador M, Isingrini E, Keiflin R. Systemic cocaine can reinstate an instrumental response not directed towards cocaine but previously performed under cocaine. Soc Neurosci Abstract. 2006;189:12. [Google Scholar]
- Czoty PW, Gage HD, Nader SH, Reboussin BA, Bounds M, Nader MA. Acquisition of cocaine self-administration does not alter dopamine D2 receptor or transporter availability in rhesus monkeys. J Addiction Med. 2007 doi: 10.1097/ADM.0b013e318045c038. in press. [DOI] [PubMed] [Google Scholar]
- Czoty PW, Morgan D, Shannon EE, Gage HD, Nader MA. Characterization of dopamine D1 and D2 receptor function in socially housed cynomolgus monkeys self-administering cocaine. Psychopharmacology. 2004;174:381–388. doi: 10.1007/s00213-003-1752-z. [DOI] [PubMed] [Google Scholar]
- Czoty PW, McCabe C, Nader MA. Assessment of the relative reinforcing strength of cocaine in socially housed cynomolgus monkeys using a choice procedure. J Pharmacol Exp Ther. 2005;312:96–102. doi: 10.1124/jpet.104.073411. [DOI] [PubMed] [Google Scholar]
- de Wit H, Stewart J. Reinstatement of cocaine-reinforced responding in the rat. Psychopharmacology. 1981;74:134–143. doi: 10.1007/BF00432175. [DOI] [PubMed] [Google Scholar]
- Dews PB, Wenger GR. Rate-dependency of the behavioral effects of amphetamine. In: Thompson T, Dews PB, editors. Advances in Behavioral Pharmacology. Vol. 1. Academic Press; New York: 1977. pp. 167–227. [Google Scholar]
- Epstein DH, Preston KL. The reinstatement model and relapse prevention: a clinical perspective. Psychopharmacology. 2003;168:31–41. doi: 10.1007/s00213-003-1470-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epstein DH, Preston KL, Stewart J, Shaham Y. Toward a model of drug relapse: an assessment of the validity of the reinstatement procedure. Psychopharmacology. 2006;189:1–16. doi: 10.1007/s00213-006-0529-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franks GJ, Lattal KA. Antecedent reinforcement schedule training and operant response reinstatement in rats. Anim Learn Behav. 1976;4:374–378. [Google Scholar]
- Gasior M, Paronis CA, Bergman J. Modification by dopaminergic drugs of choice behavior under concurrent schedules of intravenous saline and food delivery in monkeys. J Pharmacol Exp Ther. 2004;308:249–259. doi: 10.1124/jpet.103.052795. [DOI] [PubMed] [Google Scholar]
- Katz JL, Higgins ST. The validity of the reinstatement model of craving and relapse to drug use. Psychopharmacology. 2003;168:21–30. doi: 10.1007/s00213-003-1441-y. [DOI] [PubMed] [Google Scholar]
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD. Dopamine D1- and D2-like receptor mechanisms in relapse to cocaine-seeking behavior: effects of selective antagonists and agonists. J Pharmacol Exp Ther. 2000;294:680–687. [PubMed] [Google Scholar]
- Kelleher RT, Morse WH. Determinants of the specificity of behavioral effects of drugs. Ergeb der Physiol Biolog Chem Exp Pharmakol. 1968;60:1–56. doi: 10.1007/BFb0107250. [DOI] [PubMed] [Google Scholar]
- Myers KM, Davis M. Behavioral and neural analysis of extinction. Neuron. 2002;36:567–584. doi: 10.1016/s0896-6273(02)01064-4. [DOI] [PubMed] [Google Scholar]
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR, Friedman DP, Porrino LJ. Effects of cocaine self-administration on striatal dopamine systems in rhesus monkeys: initial and chronic exposure. Neuropsychopharmacology. 2002;27:35–46. doi: 10.1016/S0893-133X(01)00427-4. [DOI] [PubMed] [Google Scholar]
- O'Brien CP. A range of research-based pharmacotherapies for addiction. Science. 1997;278:66–70. doi: 10.1126/science.278.5335.66. [DOI] [PubMed] [Google Scholar]
- O'Brien CP. Anticraving medications for relapse prevention: a possible new class of psychoactive medications. Am. J. Psychiatry. 2005;162:1423–1431. doi: 10.1176/appi.ajp.162.8.1423. [DOI] [PubMed] [Google Scholar]
- Odum AL, Shahan TA. D-amphetamine reinstates behavior previously maintained by food: importance of context. Behavioral Pharmacology. 2004;15:513–516. doi: 10.1097/00008877-200411000-00007. [DOI] [PubMed] [Google Scholar]
- Reid RL. The role of the reinforcer as a stimulus. Br J Psychol. 1958;49:202–209. doi: 10.1111/j.2044-8295.1958.tb00658.x. [DOI] [PubMed] [Google Scholar]
- Self DW, Barnhart WJ, Lehman DA, Nestler EJ. Opposite modulation of cocaine-seeking behavior by D1- and D2-like dopamine receptor agonists. Science. 1996;271:1586–1589. doi: 10.1126/science.271.5255.1586. [DOI] [PubMed] [Google Scholar]
- Self DW, Choi KH, Simmons D, Walker JR, Smagula CS. Extinction training regulates neuroadaptive responses to withdrawal from chronic cocaine self-administration. Learn Mem. 2004;11:648–657. doi: 10.1101/lm.81404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaham Y, Shalev U, Lu L, De Wit H, Stewart J. The reinstatement model of drug relapse: history, methodology and major findings. Psychopharmacology. 2003;168:3–20. doi: 10.1007/s00213-002-1224-x. [DOI] [PubMed] [Google Scholar]
- Shalev U, Grimm JW, Shaham Y. Neurobiology of relapse to heroin and cocaine: a review. Pharmacol Rev. 2002;54:1–42. doi: 10.1124/pr.54.1.1. [DOI] [PubMed] [Google Scholar]
- Skinner BF, Heron WT. Effects of caffeine and Benzedrine upon conditioning and extinction. Psychological Record. 1937;1:340–346. [Google Scholar]
- Spealman RD, Barrett-Larimore RL, Rowlett JK, Platt DM, Knroyan TV. Pharmacological and environmental determinants of relapse to cocaine-seeking behavior. Pharmacol Biochem Behav. 1999;64:327–336. doi: 10.1016/s0091-3057(99)00049-0. [DOI] [PubMed] [Google Scholar]
- Stretch R, Gerber RJ, Wood SM. Factors affecting behavior maintained by response-contingent intravenous infusions of amphetamine in squirrel monkeys. Can J Physiol Pharmacol. 1971;49:593–598. doi: 10.1139/y71-075. [DOI] [PubMed] [Google Scholar]