

Abstract
In lepidopterous larvae the maxillary palps contain a large portion of the sensory equipment of the insect. Yet, knowledge about the sensitivity of these cells is limited. In this paper a morphological, behavioral, and electrophysiological investigation of the maxillary palps of Yponomeuta cagnagellus (Lepidoptera: Yponomeutidae) is presented. In addition to thermoreceptors, CO2 receptors, and gustatory receptors, evidence is reported for the existence of two groups of receptor cells sensitive to plant volatiles. Cells that are mainly sensitive to (E)-2-hexenal and hexanal or to (Z)-3-hexen-1-ol and 1-hexanol were found. Interestingly, a high sensitivity for benzaldehyde was also found. This compound is not known to be present in Euonymus europaeus, the host plant of the monophagous Yponomeuta cagnagellus, but it is a prominent compound in Rosaceae, the presumed hosts of the ancestors of Y. cagnagellus. To elucidate the evolutionary history of this sensitivity, and its possible role in host shifts, feeding responses of three Yponomeuta species to benzaldehyde were investigated. The results confirm the hypothesis that the sensitivity to benzaldehyde evolved during the ancestral shift from Celastraceae to Rosaceae and can be considered an evolutionary relict, retained in the recently backshifted Celastraceae-specialist Y. cagnagellus.
Keywords: Sensory physiology, Plant odours, Yponomeuta cagnagellus, Adaptive speciation, Lepidoptera
Introduction
Recently, progress has been made in our understanding of speciation in herbivorous insects, and mathematical models demonstrate that adaptive speciation driven by selection on ecological traits is theoretically plausible (Kondrashov and Kondrashov 1999; Dickman and Doebeli 1999; Doebeli 2005). However, as yet not much work has been done on the actual nature of these traits. The theoretical results make clear that host shifts and concomitant changes in host acceptance behaviour are important factors in adaptive speciation, but the underlying changes in the neural system are poorly studied and understood. Only recently work addressing this type of question is starting to appear (cf. Olsson et al. 2005a, b) .
Although host acceptance in phytophagous insects is usually viewed as solely governed by the adult (“mother knows best”), it should be noted that in many species larval host acceptance is equally important. Lepidopterous larvae in particular often display striking food preferences, which are based on a small set of chemoreceptors (Schoonhoven and van Loon 2002), thus making caterpillars promising models to study the neural basis of host plant recognition, host shifts, and the evolution of insect–plant relations. However, such an approach requires detailed knowledge about the responses of the involved receptors, information that is almost completely lacking.
External chemoreceptors in caterpillars are located on the antennae, on the maxillae, and in the epipharynx. The gustatory sensilla styloconica on the galea are important in host acceptance behaviour, and a large body of data is available about their sensitivity (see Schoonhoven and van Loon 2002; Schoonhoven et al. 2005 for review). The other sets of receptors, the epipharyngeal organs on the inside of the labrum, and the sensilla on the antennae and the maxillary palps, are more difficult to study and as a consequence less well known. The maxillary palp receptors in particular have received very little attention. Except for the pioneering studies of Schoonhoven and Dethier (1966), recordings from the maxillary nerve of the silkworm by Hirao et al. (1976, and a recent paper on gustatory receptors in the palp of Choristoneura fumiferana (Albert 2003), no electrophysiological data on the maxillary palps of Lepidoptera appears to be available. However, behavioural work indicates that the influence of the palps on feeding can be as significant as that of the taste sensilla on the galea (Hanson and Dethier 1973; Glendinning et al. 1998; de Boer 1993, 2006).
The small ermine moth genus Yponomeuta Latreille (Lepidoptera: Yponomeutidae) consists of some 75 species (Gershenson and Ulenberg 1998). Most are specialised on Celastraceae, and phylogenetic analysis (Menken 1996; Turner H, Lieshout N, van Ginkel W, Menken SBJ, submitted; Ulenberg, personal communication) strongly suggest that this association is the ancestral state in the genus. Moreover, mapping host use upon the phylogeny indicates an intriguing evolutionary scenario: the genus most likely originated in the Far East and dispersed to the western Palearctic concomitant with a (probably single) shift to Rosaceae and further to Salicaceae (Menken 1996). Interestingly, one lineage within this clade, Y. cagnagellus (Hübner), shifted back to the ancestral host, so this species is secondarily associated with Celastraceae. Yponomeuta plumbellus on the other hand is present in Western Europe, but this species is more closely related to the clades in the Far East, and retained the ancestral relation with Celastraceae.
The feeding of the Western European Yponomeuta species, and in particular Y. cagnagellus, is relatively well studied. Both the behaviour (Gerrits-Heybroek et al. 1978; van Drongelen 1980; Kooi and van de Water 1988; Peterson et al. 1990) and the underlying sensory physiology (van Drongelen 1979; Roessingh et al. 1999) have been addressed. The results demonstrate a clade of predominantly monophagous species with well-adapted sensory systems displaying high sensitivity for sugar alcohols characteristic for the host–plant families, as well as reduced sensitivities for host-specific deterrents. However, this whole body of work is focused on gustatory responses of styloconic sensilla on the galea that may provide only part of the relevant sensory inputs (de Boer 2006).
Here we present a morphological, behavioural, and electrophysiological survey of chemoreceptors on the maxillary palps in the larvae of Yponomeuta cagnagellus. In addition, we performed a comparative study into the effects of benzaldehyde on feeding behaviour of three related Yponomeuta species. Benzaldehyde strongly stimulated sensory cells in the maxillary palp of Y. cagnagellus, but its role in feeding behaviour is unknown. This compound is of particular interest because it is prominent in Rosaceae such as Prunus spinosa (Humpf and Schreier 1992), the host of Y. padellus (L), but absent from Euonymus europaeus, the host of Y. cagnagellus and Y. plumbellus (Denis and Schiffermüller) (Fig. 1). In this comparison we take advantage of the fact that the phylogenetic tree indicates that Y. plumbellus, although present in Western Europe, belongs to a clade that still displays the original ancestral association with Celastraceae. Yponomeuta padellus is in the middle of the clade that shifted to the benzaldehyde containing Rosaceae. Yponomeuta cagnagellus on the other hand is now feeding on the Celastraceous E. europaeus, that does not contain benzaldehyde, but this species most likely originated from an ancestor that was feeding on Rosaceae. An analysis of the phylogenetic pattern of the response to benzaldehyde in these three species will therefore contribute to our understanding of the flexibility of sensory systems and the role of sensory stimuli in host shifts and adaptive speciation.
Fig. 1.
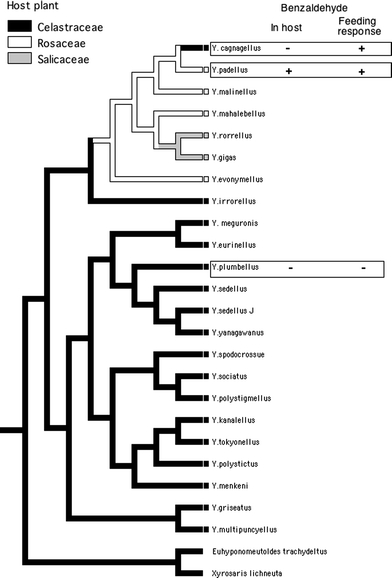
Phylogeny of Yponomeuta, based on the internal transcribed spacer region (ITS-1) and on 16S rDNA (16S) and cytochrome oxidase (COII) mitochondrial genes, using maximum parsimony (Turner H, Lieshout N, van Ginkel W, Menken SBJ, submitted). Host plant use is plotted on the tree and the presence of benzaldehyde in the host and feeding responses to this compound are indicated for the three tested species
Materials and methods
Insects
Egg batches of Y. cagnagellus and Y. padellus were collected in the field in The Netherlands from their host plants: Y. cagnagellus from E. europaeus and Y. padellus from P. spinosa and Crataegus spp. Yponomeuta plumbellus larvae were collected as fourth instars. Egg batches were kept at 4°C and 80% relative humidity until needed. Larvae were reared in 10 cm wide Petri dishes (at 25°C, ambient humidity, L18:D6 h photoperiod) on host foliage from potted plants grown in the greenhouse (E. europaeus for Y. cagnagellus and Y. plumbellus; P. spinosa for Y. padellus). During the rearing period larvae were fed ad libitum. For the electrophysiological experiments. 1- to 4-day-old fifth instars were used that were starved for 2–3 h. In feeding experiments the caterpillars were used during the whole duration of the fifth stadium. Between feeding experiments, caterpillars were kept at 4°C.
Morphology
The morphology of the palps was studied using scanning EM. Fixation and drying often induce shrinking and cause collapse of soft tissue parts. We avoided these problems by using Cryo-SEM. Larvae were used directly after moulting into the fifth stadium. The preparation was stabilized by quick immersion in nitrogen slush (60 K), coated with a 200 Å thick layer of gold and viewed in the frozen state using a Philips 535 SEM equipped with a Hexland CT1000/CP2000 and a Jeol 35C SEM equipped with an EMSCOPE SP2000A cryo preparation assembly.
Host plant odour composition
One of the problems in determining the function of unknown sensory organs is the choice of the stimuli to be tested. To identify a set of relevant substances, the chemical composition of the headspace of E. europaeus, the host of Y. cagnagellus, was characterized. Branches (30 cm long) of E. europaeus growing in the field in Wageningen, the Netherlands, were cut with sharp garden scissors and transferred to the lab. About 2 kg of branches was placed in a 20-l stainless steel vessel within 30 min after cutting. Care was taken not to damage the leaves. With a membrane pump air was passed through KOH pellets, molecular sieves 5A and 3×, active charcoal, and a tenax pre-trap. This cleaned air flowed for 7–15 h at a rate of 70 ml/min through the stainless steel vessel and subsequently through the tenax-TA sample trap. The collected volatiles were released from the tenax by the TCT technique (Thermo-desorption cold trap; Dicke et al. 1990). The TCT unit (Chrompack, Middelburg, The Netherlands) was heated (250°C) for 3 min and desorbed compounds were collected on a Sil5CB coated, fused silica capillary at −100°C. Flash heating of the cold trap provided sharp injection of the compounds into the GC/MS unit (Chrompack Sil19CB column, 25 m × 0.25 mm, df 0.25 μm). Electron impact ionisation was carried out at 70 eV on a VG MM 7070F mass spectrometer.
Electrophysiological recording and odour stimuli
A diagram of the recording set-up is presented in Fig. 2. An isolated caterpillar head was mounted on a silver wire loop and connected via a 10-mm connector to the input terminal of a laboratory-built amplifier (Ri > 109 Ω, Ci < 1 pF, Ib < 1 pA). The sensory cells of the sensilla on the maxillary palp are located below the base of the terminal segment (Schoonhoven and Dethier 1966; Albert 1980). To gain access to these cells, the connecting membrane between the two most distal segments was pierced with a sharpened tungsten electrode. The actual single cell recordings were made using a glass capillary electrode filled with 3 M KCl (impedance about 40 MΩ) inserted through the hole and connected to ground (so actually serving as the indifferent electrode). The position of the glass electrode was carefully adjusted until action potentials could be recorded. The inside of the palp is probed with the tip until activity is picked up, analogous to the way in which recordings from the CNS are made. It should be noted that although this procedure has the advantage of allowing the recoding from single cells with good S/N ratios, and easy identification of separate cells, the drawback is that the position of the tip (and therefore the identity of the recorded cell) is not known, and not reproducible between recordings.
Fig. 2.
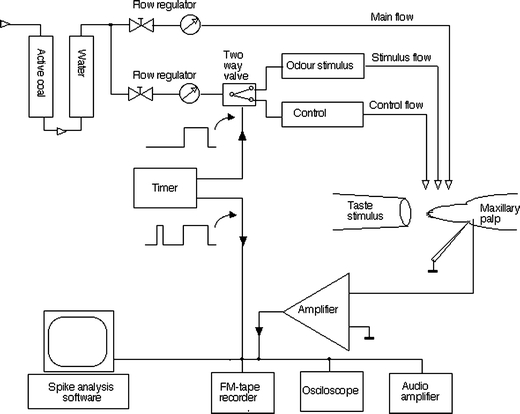
Schematic diagram of the electrophysiological set-up used to measure responses to both gustatory and olfactory stimuli. Open arrows indicate airflow, black arrows electrical signals. Note that in contrast to standard tip recording the stimulus pipette is electrically isolated
The preparation was placed in a continuous stream of clean, moistened air (80 ml/s, 25 cm/s). A second (control) air stream (2 ml/s) was injected in the main stream through an empty pasteur pipette. Odour stimuli were delivered by switching this second stream for 1 s through a pasteur pipette containing the stimulus. For stimulation with CO2, a single injected stream of commercial available bottled CO2 (Hoekloos, the Netherlands) was switched on. This injected stream could be adjusted to produce CO2 concentrations in the main stream between 0.1 and 10%. Olfactory stimuli were diluted in paraffin oil (10−3, 10−2, and 10−1 v/v). Twenty-five microlitres of these solutions was applied to filter paper strips (6.0 × 0.5 cm) and was placed in pasteur pipettes. The following compounds (99% pure) were used: 1-hexanol and benzaldehyde (Fluka, Switzerland); (Z)-3-hexenyl acetate and (E)-2-hexenal (Z)-3-hexen-1-ol (Roth, West Germany); hexanal (98% pure) (Merck, USA); limonene, citral (97% pure), and geraniol (98% pure) (Aldrich, Germany). All stimuli were applied in random order with at least 30 s between them. Longer intervals were used after strong stimuli. Preparations were discarded after 60 min and were changed after a successful recording. Odour stimuli were stored at 4°C between recording sessions, and replaced every few days.
The AC amplified action potentials from the sensory cells were recorded on tape (Racal FM taperecorder). To facilitate computer-assisted analysis, the valve signal and a pre-pulse 1 s earlier were recorded on an additional channel.
Analysis of recordings
The recordings contained activity from one to four sensory cells but usually not more than two. Recordings with more than three cells were discarded. Multi-cellular responses were analysed with the help of an interactive computer program based on the methods described in Roessingh et al. (1992). Spikes were characterized primarily by amplitude. Amplitude boundaries for different cells were taken from an amplitude versus time plot. The separation of the cells is an interactive process in which spike amplitude and spike interval distributions are used as criteria to judge the separation. After the separation, responses for each cell were calculated as the difference in action potential frequency in the last second before the onset of stimulation and the first reaction second. A cell was assumed to respond to a stimulus if an increase of at least ten spikes could be evoked. Cells with weaker reactions were assumed to be sensitive to untested stimuli and were not further analysed. To test the significance of observed differences in spike frequency, analysis of variance was used on log transformed response data.
No-choice feeding assay
A new no-choice feeding assay was designed to investigate the effect of benzaldehyde odour on larval feeding. The experimental setup is illustrated in Fig. 3. A clean air stream was split into two branches, with airflow meters to control the flow. Each branch was connected to a cluster of 18 Petri dishes, each dish containing a feeding caterpillar. In the control group, clean air was passed over the feeding larvae. The air for the experimental group was passed through a small bottle of benzaldehyde diluted in paraffin oil. A bypass with a needle valve allowed mixing in clear air and fine-tuning of the concentration of benzaldehyde (i.e. 50% flow through the bypass reduces the final concentration with a factor 2. Six different dilutions (v/v) of benzaldehyde in paraffin oil were used in the experiments, viz. 0.5 × 10−4, 0.5 × 10−3, 0.2 × 10−2, 0.5 × 10−2, 1 × 10−2, and 0.5 × 10−1. The total airflow was approximately 50 ml/s, resulting in a flow of approximately 1.4 ml/s through each Petri dish.
Fig. 3.
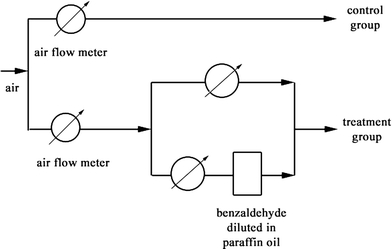
Feeding assay air flow diagram. Both the control group and treatment group consisted of 18 Petri dishes
A leaf disc with a diameter of 11 mm made from fresh host leaves was placed in each Petri dish on top of a piece of filter paper slightly wetted with distilled water. To reduce baseline palatability and detect feeding stimulation and/or inhibition, the leaf discs were extracted in alcohol and then washed in distilled water for 10 min. The duration of extraction needed to induce a weak feeding response depending upon both the host plants and leaf quality. For Y. cagnagellus, E. europaeus leaf discs were soaked in 96% EtOH for 24 h. Prunus spinosa leaf discs for Y. padellus were soaked for 2 h in 70% EtOH. Yponomeuta plumbellus appears later in the season than the other two species and feeds on slightly older E. europaeus foliage. For these older leaves extraction of leaf discs for 1 h in EtOH was sufficient to reduce palatability. The feeding experiments lasted 4–5 h. Usually 1 or 2 h after the start of the experiment, caterpillars started eating. The experiment was terminated after at least two third of the insects had been eating for more than 3 h.
To quantify the amount of feeding, the remainders of the leaf discs were scanned with an EPSON perfection 1250 scanner. The surface area was measured on a Macintosh computer using the open source program NIH Image version 1.62 (developed at the U.S. National Institutes of Health and available on the Internet at http://www.rsb.info.nih.gov/nih-image/). The differences in consumption between treatment and control groups were evaluated using a Mann–Whitney test in SPSS (SPSS Inc. 2001). For graphical display a feeding index (C − T)/(C + T) × 100% (ranging from −100 to 100%), was calculated, in which C and T denote, respectively, the mean of the remaining leaf surface in the control group and in the treatment group.
Results
Morphology
The palps of Y. cagnagellus are more elongated than in most other species of Lepidoptera, however, the sensory equipment does conform to the general pattern of ditrysian species as outlined by Grimes and Neunzig (1986). These authors divide the eight sensilla on the apex in a medial group (M1, M2), an apical group (A1, A2, A3), and a lateral group (L1, L2, L3). These groups can also be recognised in Y. cagnagellus, although the position of the sensilla is more linear than in the generalised pattern (4c).
Fig. 4.
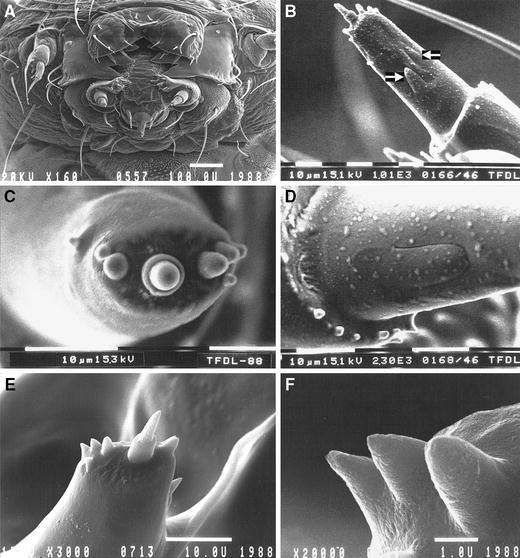
Cryo-SEM micrographs of Yponomeuta cagnagellus.a Ventral view of the head. b Maxillary palp showing the digitiform sensillum (leftmost arrow) and a large campaniform sensillum (rightmost arrow). c Overview of the tip of the most distal segment. Note the linear arrangement of the sensilla. d Detail of the digitiform sensillum. e On the apex of the palp, seven sensilla basiconica and one sensillum styloconicum are present. From left to right, L1, L2, L3, A1, a styloconic A2, A3, a partly hidden M1, and M2 can be seen (terminology after Grimes and Neunzig 1986). f High magnification of the lateral side of the apex. The two leftmost sensilla belong to the lateral group. The slightly more blunt sensillum on the right is A1. Note the absence of surface structures
On the apex seven sensilla basiconica and one sensillum styloconicum can be distinguished. On the side of the segment, a digitiform sensillum and at least one campaniform sensillum are visible (4b). With the exception of the styloconic sensillum, which has a prominent base, the sensilla on the apex are simple blunt cones. The lateral and medial sensilla, for which a gustatory function has been suggested, seem to be slightly more tapered (Fig. 4e, f) but the difference is only slight. Structural details like grooved surfaces or apical pores were not observed.
Host plant odour composition
The compounds used for olfactory stimulation were chosen on the basis of the headspace analysis of E. europaeus. Whole undamaged plants release only limited amounts of volatiles (Buttery et al. 1985; van Poecke and Dicke 2004), and indeed small quantities were found in the present study. The only compounds that could be detected consistently were C6-based “green odours” (Visser and Avé 1978). Other compounds (limonene, beta-ocimene, alpha-copaene, beta-bourbonene, benzaldehyde) could not be detected reliably. Based on this result (Z)-3-hexenyl acetate (E)-2-hexenal, 1-hexanal (Z)-3-hexen-1-ol, hexanol, limonene, citral, geraniol, and benzaldehyde were chosen as olfactory stimuli. Benzaldehyde is not known from E. europaeus, but was nevertheless included in the analysis because it is a major compound in the Rosaceous hosts of some other Yponomeuta species that are closely related to Y. cagnagellus.
Electrophysiology
In experiments on 69 preparations, a total of 85 single cells were analysed. Twenty-four cells did not respond to any of the applied stimuli. Of the remaining receptors, 22 cells showed no response to odours, but responded to a 100 mM KCl solution. However, these gustatory responses were only weak (21 ± 4 spikes maximum) so the adequate stimulus was most likely not KCl. Gustatory responses will therefore not be considered here any further. Twenty-two other cells responded strongly to C6 compounds and to benzaldehyde (Fig. 5). For 19 of those cells more or less complete response spectra could be obtained (Fig. 6). At least two groups could be distinguished. Cells one to five responded to the aldehydes (E)-2-hexenal and hexanal. The other cells respond to the leaf alcohol (Z)-3-hexen-1-ol, to 1-hexanol, and to benzaldehyde. No cells were found that responded strongly to (Z)-3-hexen-1-yl acetate, or to the terpenoids limonene (6 cells tested), citral (3 cells tested) and geraniol (5 cells tested).
Fig. 5.
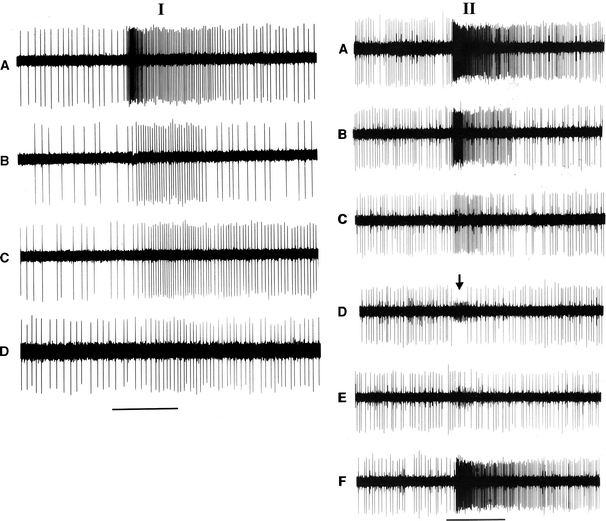
I Responses of a (E)-2 hexenal cell in the maxillary palp of Yponomeuta cagnagellus to plant odour compounds. A Hexanal 10−1, B Hexanal 10−2. C (Z)-3-hexen-1-ol 10−1. D (Z)-3-hexen-1-ol 10−2. The stimulus period (1 s) is indicated by the horizontal bar. II Responses of a (Z)-3-hexen-1-ol best cell in the maxillary palp of Yponomeuta cagnagellus to plant odour compounds. A (Z)-3-hexen-1-ol 10−1. B (Z)-3-hexen-1-ol 10−2, C (Z)-3-hexen-1-ol 10−3. D (E)-2-hexenal 10−2, E hexanal 10−2, F benzaldehyde 10−2. Note in D and E the presence of an aldehyde-sensitive cell in the noise (arrow). The stimulus duration (1 s) is indicated by the horizontal bar
Fig. 6.
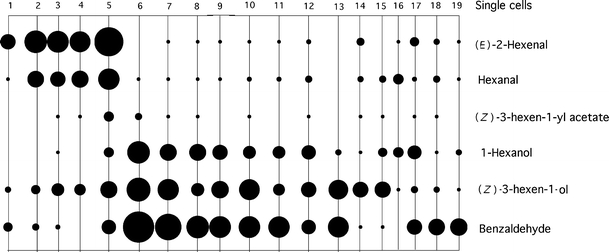
Response spectra for 19 cells sensitive to plant odour compounds in the maxillary palps of Yponomeuta cagnagellus at a concentration of 10−2. Response intensity is indicated by the diameter of the circles. Empty spots are missing observations
Dose response relations for green odours in the two main cell groups are given in Fig. 7. In Fig. 7a, the responses of the (E)-2-hexenal sensitive cells to the other green odours are plotted. The same is done in Fig. 7b for the (Z)-3-hexen-1-ol cells. In the alcohol-sensitive group, several cells responded to both (Z)-3-hexen-1-ol and 1-hexanol (cells 6–12) but cells that reacted mainly to one of the compounds were also found (cells 13–19).
Fig. 7.
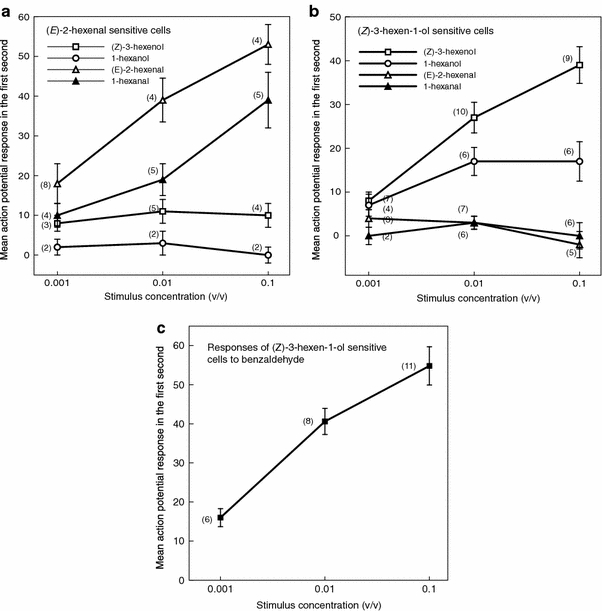
Dose–response relations of olfactory receptor cells in the cells of Yponomeuta cagnagellus. a (E)-2-hexenal sensitive cells (data from cell 1 to 5 in Fig. 6, plus three additional cells). b (Z)-3-hexen-1-ol sensitive cells (data from cell 6 to 19 in Fig. 6). c. benzaldehyde sensitive cells (data from cell 6 to 16 in Fig. 6). The number of sensory cells actually recorded for each datapoint is given between brackets, vertical bars indicate the SE
The (Z)-3-hexen-1-ol and 1-hexanol cells were often also highly sensitive to benzaldehyde (Fig. 7c). Evidence indicating that this sensitivity could be attributed to another cell (e.g. superpositions or irregular spike interval histograms) was not observed. The “aldehyde” cells in our sample did not show this sensitivity and only two cells responded weakly to benzaldehyde.
Although the sensitivity to benzaldehyde was often associated with that for the leaf alcohols, examples of both benzaldehyde insensitive cells (cell 14–15) and apparent benzaldehyde specialists (cells 18–19) were found.
Fourteen cells responded selectively to CO2 (Fig. 8). The dose–response curve (Fig. 9) is almost linear from 0.2 to 2.5% CO2. No significant response to other stimuli was found for these cells. All tested C6 compounds sometimes inhibited spike activity in CO2 cells, but only at high concentrations (10−1 v/v). The terpenoids limonene, citral, and geraniol never inhibited the CO2 cells (n = 14).
Fig. 8.
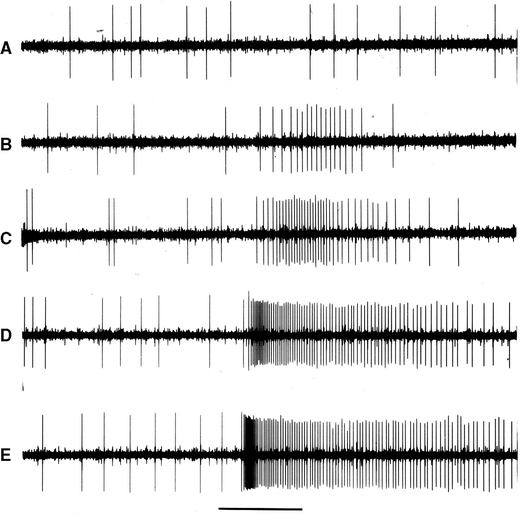
Response of a CO2 sensitive cell in the maxillary palp of Yponomeuta cagnagellus. a to e are responses to, respectively, 0.26, 1.0, 1.5, 2.5, and 10% CO2. The time marker indicates 1 s
Fig. 9.
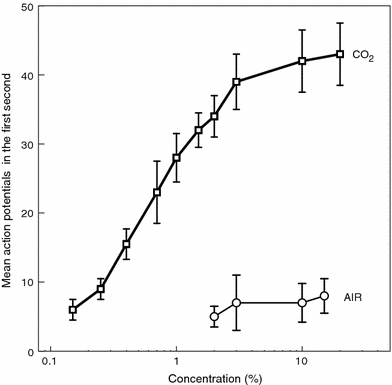
Dose–response relationship of 12 CO2 sensitive cells in the palp of Yponomeuta cagnagellus stimulated with CO2 or clean air. Vertical bars indicate 95% confidence interval
Three cells did not react to chemical stimuli but responded to changes in temperature induced by a hot or cold metal rod in the vicinity of the palp. A drop in temperature evoked a strong response. An increase in temperature inhibited the cells (Fig. 10), i.e. these cells functioned as cold receptors.
Fig. 10.
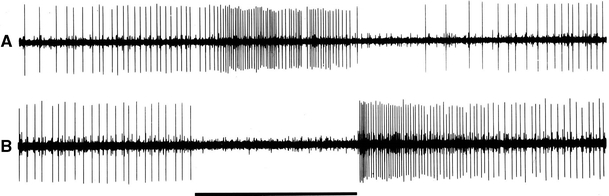
Response of a cold receptor in the maxillary palp of Yponomeuta cagnagellus. a Stimulation with a cold metal rod at an upwind distance of about 10 mm. b Stimulation with a warm metal rod at an upwind distance of about 10 mm. Stimulus bar 2.5 s
Feeding behaviour modulated by benzaldehyde
Feeding assays were conducted for Y. cagnagellus at 5 different concentrations of benzaldehyde on 72 insects per treatment (Fig. 11). Benzaldehyde had a positive feeding index at concentrations of 0.5 × 10−3, 0.2 × 10−2, and 0.5 × 10−2. However, the stimulating effect of benzaldehyde was significant only at the concentration of 0.2 × 10−2 M, with a feeding index of about 12% (percentage leaf disk left on benzaldehyde and control was, respectively, 60.9 ± 5.3 and 76.8 ± 4.4%, Mann–Whitney U = 427, P = 0.013). At higher concentrations, benzaldehyde tended to inhibit the feeding of Y. cagnagellus larvae. For example, the feeding index was −3% at the concentration of 0.5 × 10−1 but feeding inhibition was not significant compared to the control group.
Fig. 11.
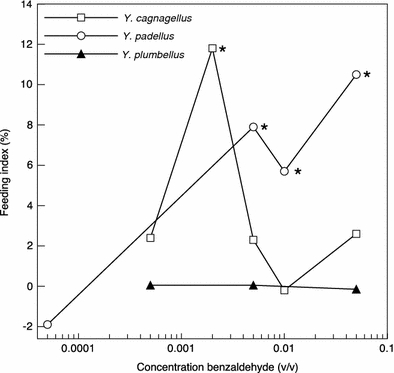
Feeding index of three Yponomeuta species. FI = (C − T)/(C + T) (ranging from −100 to 100%). C and T denote, respectively, the mean of the remaining leaf surface in the control group and in the treatment (benzaldehyde) group. Feeding indexes with P < 0.05 are indicated with an asterisk
The feeding response of Y. padellus was tested at four concentrations, i.e. 0.5 × 10−4, 0.5 × 10−2, 1 × 10−2, and 0.5 × 10−1 (Fig. 11). Benzaldehyde significantly stimulated the feeding of caterpillars at all concentrations except at the lowest concentration (percentage leaf disk left on benzaldehyde and control at 0.5 × 10−2 was, respectively, 76.5 ± 3.7 and 84.5 ± 3.4%, Mann–Whitney U 375, P = 0.003, At 1 × 10−2 76.6 ± 3.8 and 90.4 ± 1.7%, Mann–Whitney U = 321, P = 0.001 and at 0.5 × 10−1 70 ± 3.8% and 82.8 ± 3.3% Mann–Whitney U = 408, P < 0.007). In Y. plumbellus, no effect of benzaldehyde on food intake was found, and the insects appeared to be fully indifferent to the compound (Fig. 11).
Discussion
Morphology
A description of the external structure of the maxillary palps of ditrysian larvae is given by Grimes and Neunzig (1986). From their results it is clear that location, size, and shape of palpal sensilla are constant in lepidopterous larvae. Eight sensilla are always found on the apex. In addition, a digitiform sensillum and up to four campaniform sensilla or pore plates are present in the wall of the most distal segment. Grimes and Neunzig (1986) make a distinction between the more primitive, endophagous feeders, and larger, exophagous feeders. In endophagous species, the walls of the A sensilla are smooth, and A2 is often two-tiered (styloconic). In the more advanced, exophagous groups A2 is basiconic and the A sensilla are wrinkled or pockmarked. The Yponomeutidae are regarded as relatively primitive Ditrysia (Kristensen and Skalski 1999), and indeed a styloconic A2 and smooth A sensilla are found (Fig. 4; see also van Drongelen 1979). On morphological grounds a gustatory function for L and M sensilla has been suggested (Grimes and Neunzig 1986), and for Choristoneura fumiferana this was confirmed by recordings (Albert 2003). In Helicoverpa armigera, however, some sensilla show ultrastructural features characteristic for both olfaction and taste (Keil 1996) and it is therefore difficult to assign a function based on mophology alone. Although we recorded responses to gustatory as well as olfacory stimuli, we stimulated all eight sensilla simultaneously and this method precludes assignment of the observed responses to particular sensilla. The wall of the most distal segment of the palp bears at least one campaniform sensillum. This sensillum is probably equivalent to the large plate sensillum described in C. fumiferana (Albert 2003), Euxoa messoria (Devitt and Smith 1982), and H. amigera (Keil 1996). The plate sensilla are multiporous, multiple innervated, and possess branching dendrites. If this is also true of Y. cagnagellus, these sensilla are presumably olfactory organs where some of the neural responses reported in this paper could originate.
In the wall of the most distal segment, a digitiform sensillum is present which is normally innervated by one neuron (Schoonhoven and Dethier 1966; Albert 1980; Devitt and Smith 1982; Keil 1996). In E. messoria and Heliotis amigera this sensillum appears to possess a laminated outer dendritic segment. Such laminated dendrites have been associated with cold receptors (see Altner and Loftus 1985), as well as with CO2 receptors (Chu-wang et al. 1975; McIver and Siemicki 1984; Lee et al. 1985; Bogner et al. 1986). In Y. cagnagellus both cold receptors and CO2 receptors were found. The digitiform sensillum of Y. cagnagellus might therefore house a CO2 receptor as suggested by Keil (1996) but without further electrophysiological evidence cold reception cannot be excluded.
Carbon dioxide reception
The ecological function of the CO2 receptor in Y. cagnagellus is not clear. CO2 reception is well known in soil dwelling organisms that must locate plant roots (Johnson and Gregory 2006). CO2 sensitivity is also common in adult lepidoptera (Bogner 1990; Stange and Stowe 1999). Helicoverpa armigera for instance is in principle capable of detecting modulation of CO2 concentrations normally occurring in the environment (Stange 1992). The CO2 receptor of Y. cagnagellus and other lepidopterous larvae is clearly less sensitive than that of adult Lepidoptera, however, CO2 receptors with a similar low sensitivity range have been described in the tick Amblyomma varigatum (Steullet and Guerin 1992). During the day, CO2 levels near the surface of green leaves will be 0.038% or lower, while this level may rise to a maximum of 1.2% during nocturnal respiration (see Bogner 1990 for references). Although the latter concentration is just within the range of the larval receptor, it seems unlikely that the CO2 receptor is used to assess subtle changes in the respiratory status of the leaf (as a messure of leaf quality). Alternative hypotheses are that the receptor is used to monitor CO2 levels in the communal nest during periods of tight clustering, or that it facilitates aggregation behaviour. However, as yet experimental data to support these ideas is lacking.
Temperature perception
Only qualitative data was collected from the three observed cold receptors. The actual drop in temperature induced by the hot and cold metal rods used, as stimulus is not known. The cold receptors might be used for monitoring the microclimate (Schoonhoven 1967) and in principle could be used to determine smaller temperature variations associated with leaf quality (Dethier and Schoonhoven 1968).
Plant odour responses
Stimulation with plant odours yielded cells of at least two spectral types: aldehyde-sensitive cells and alcohol-sensitive cells. The two groups are clearly separated from each other. However, within each group responses to several chemically very different stimuli were found. A single receptor cell that reacts to different types of compounds used to be interpreted as a reflection of the presence of several receptor “sites” on the membrane (Kaissling 1976; Selzer 1984; Hansen 1978; Wiezoreck and Wolff 1989). With the identification of the actual receptor genes and advanced labeling techniques it was suggested that normally only a single receptor protein is expressed in a particular sensory cell, the “one receptor–one neuron” rule (see Gaillard et al. 2004 for review). Recently however, it was shown that in Drosophila olfactory organs this rule does not always hold, and coexpression of different receptor proteins was observed (Goldman et al. 2005). In C. elegans coexpression must also occur, simply because there are too many receptor genes to express for the limited number of sensory neurons. We observed single cells that responded to structurally different compounds such as (Z)-3-hexen-1-ol and benzaldehyde, but also cells that responded to only one of these two compounds. These observations can in each case still be explained by the expression of a single receptor protein per cell, with a different binding specificity (i.e. a cell that responds to several compounds expresses a receptor with a broad specificity, while a cell that responds to only a single compound expresses a narrowly tuned receptor protein). Alternatively, cells might be broadly tuned because they express an assembly of several narrowly tuned receptor proteins with different sensitivity spectra. In either case, following this line of reasoning a simple quantitative shift in the numbers of receptor proteins could in principle result in a qualitative change in host-acceptance behaviour and host shifts, processes that were postulated as major factors in the speciation process in Yponomeuta species (Menken et al. 1992; Menken and Roessingh 1998).
From Fig. 6 is clear that although at least two groups of cells with comparable spectral responses can be distinguished (cells 1–5, cells 6–19), however, there are no sharp boundaries between these classes (cf. cell 5), and considerable variation does exist. Since care was taken to record only from cells that were reliable and reproducibly responding, it is likely that this variation does reflect natural variability in the receptor cell population. The separation in aldehyde- and alcohol sensitive cells should therefore at present merely be considered as a convenient way to summarise the data. Until more recordings are made, it cannot be excluded that a continuum from more specialised to, more generalised cells exists.
In the present study, no cells with a primary response to (Z)-3-hexenyl acetate were found in the maxillary palps of Y. cagnagellus, but receptors sensitive to this compound have been found on the antennae of this species (Roessingh, unpublished results) as well as on the antennae of Pieris brassicae larvae (Visser and de Jong 1988). In this respect it is interesting to note that in Manduca sexta fibers from the maxillary nerve travel into a core of neuropile in the suboesophageal ganglion that also receives antennal axons (Kent and Hildebrand 1987).
The maxillary palps of caterpillars contain 15–30 chemoreceptors (Schoonhoven and Dethier 1966; Albert 1980; Devitt and Smith 1982). If the 85 cells in the present study represent a random sample from the population in the palps, then the proportion of neurons sensitive to each of the tested stimuli could be estimated. However, it must be stressed that the position of the electrode was not reproducable between recordings. As a consequence, the recordings by no means represent a random sample. In spite of this limitation it can be concluded that receptors for the tested terpenoids and for (Z)-3-hexen-1-yl acetate are relatively scarce, since no cells primarily sensitive to these stimuli were encountered.
The role of benzaldehyde in feeding behaviour
Because benzaldehyde is absent from E. europaeus, Euonymus feeders such as Y. plumbellus and Y. cagnagellus are not expected to be sensitive to this compound, or use it as a repellent. The electrophysiological results clearly show that benzaldehyde is stimulating the sensory cells in the maxillary palp of Y. cagnagellus; the straightforward interpretation would be that in Y. cagnagellus benzaldehyde signals a non-host compound, and therefore low acceptability. On the other hand, as benzaldehyde is prominently present in Rosaceae, it can be expected to be a feeding stimulant for Rosaceae feeders like Y. padellus, a species that is closely related to Y. cagnagellus.
From a phylogenetic perspective a different hypothesis can thus be formulated. Yponomeuta cagnagellus is located within the European clade that feeds mostly on Rosaceae, which do contain benzaldehyde. Considering that the most recent common ancestor of this clade was presumably a Rosaceae feeder, the sensitivity of Y. cagnagellus to benzaldehyde could be viewed as a maintained sensitivity to a feeding stimulant from the ancestral host.
Under this hypothesis both Y. cagnagellus and Y. padellus are expected to be stimulated by benzaldehyde. Yponomeuta plumbellus, a Celastraceae feeder outside the European clade, is expected to be unresponsive because it has never been associated with Rosaceae nor have its ancestors. The results from the feeding experiment obtained from the three species are consistent with the latter scenario. Benzaldehyde stimulated feeding in Y. cagnagellus and Y. padellus and increased food intake by 6–12%. Yponomeuta plumbellus did not respond to benzaldehyde at all. The sensory response to benzaldehyde in Y. cagnagellus is therefore probably an evolutionary relict of the association with Rosaceae of the ancestral lineage of this species. In a previous study (van Drongelen 1979), dulcitol sensitivity was found in Y. evonymellus and Y. padellus, species feeding on Rosaceae that do contain very little or nodulcitol. In this case it was similarly concluded that the dulcitol sensitivity might be an evolutionary relict from the ancestral host association with Celastraceae.
Although feeding in both Y. cagnagellus and Y. padellus was stimulated by benzaldehyde, Y. padellus (associated with a benzaldehyde-containing hosts) responded over a much larger concentration range than Y. cagnagellus (a species that is using a benzaldehyde-free host). It should be noted that care must be taken when interpreting these results. It is not clear what the natural concentrations of host plant odours that are experienced by the larvae, since a steep gradient in the boundary layer exists, and concentrations near the leaf surface can be much higher than those obtained from headspace samples (Schoonhoven et al. 2005). However, since the sensory responses show good dose–response relations, and the behaviour yields a clear optimum, we feel that the used concentrations are covering the natural range. It will be interesting to investigate the actual genetics underlying the observed sensitivity differences between the species.
Acknowledgments
We wish to thank Felix Thiel (TFDL, Wageningen) for invaluable help with the Cryo SEM, Teris van Beek and J. Cozijnsen for assistance and advice concerning the headspace analysis, Maarten Posthumus for the GC/MS work, Louis Schoonhoven for critical reading of the manuscript, and Birgit Nübel for rearing Yponomeuta larvae. Special thanks to Erich Städler and the Forschungsanstalt Wädenswil for providing time and facilities to complete an early version of this manuscript.
References
- Albert PJ (1980) Morphology and innervation of mouthpart sensilla in larvae of the spruce budworm, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). Can J Zool 58:842–851
- Albert PJ (2003) Electrophysiological responses to sucrose from a gustatory sensillum on the larval maxillary palp of the spruce budworm, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). J Insect Physiol 49:733–738 [DOI] [PubMed]
- Altner H, Loftus R (1985) Ultrastructure and function of insect thermo- andhygroreceptors. Ann Rev Entomol 30:273–295 [DOI]
- Bogner F (1990) Sensory physiological investigation of carbon dioxide receptors in lepidoptera. J Insect Physiol 36:951–957 [DOI]
- Bogner F, Boppré M, Ernst KD, Boeckh J (1986) CO2 sensitive receptors on labial palps of Rhodogastria moths (Lepidoptera: Arctiidae): physiology, fine structure and central projection. J Comp Physiol A 158:741–749 [DOI] [PubMed]
- Buttery RG, Xu C, Ling LC (1985) Volatile components of wheat leaves (and stems): possible insect attractants. J Agric Food Chem 33:115–117 [DOI]
- Chu-Wang, I-Wu, Axtell RC, Kline DL (1975) Antennal and palpal sensilla of the sand fly Culicoides furens (Poey). Int J Insect Morphol Embryol 4:131–149 [DOI]
- de Boer G (1993) Plasticity in food preference and diet-induced differential weighting of chemosensory information in larval Manduca sexta. J Insect Physiol 39:387–398 [DOI]
- de Boer G (2006) The role of the antennae and maxillary palps in mediating food preference by larvae of the tobacco hornworm, Manduca sexta. Entomol Exp Appl 119:29–38 [DOI]
- Dethier VG, Schoonhoven LM (1968) Evaluation of evaporation by cold and humidity receptors in caterpillars. J Insect Physiol 14:1049–1054 [DOI] [PubMed]
- Devitt BD, Smith JJB (1982) Morphology and fine structure of mouthpart sensilla in the dark-sided cutworm Euxoa messoria (Harris) (Lepidoptera: Noctuidae). Int J Insect Morphol Embryol 11:255–270 [DOI]
- Dicke M, van Beek TA, Posthumus MA, Ben Dom N, van Bokhoven H, de Groot AE (1990) Isolation and identification of violatile kairomones that affect acarine predator–prey interactions. J Chem Ecol 16:381–396 [DOI] [PubMed]
- Dieckmann U, Doebeli M (1999) On the origin of species by sympatric speciation. Nature 400:354–357 [DOI] [PubMed]
- Doebeli M (2005) Adaptive speciation when assortative mating is based on female preference for male marker traits. J Evol Biol 18:1587–1600 [DOI] [PubMed]
- Gaillard I, Rouquier S, Giorgi D (2004) Olfactory receptors. Cell Mol Life Sci 61:456–469 [DOI] [PMC free article] [PubMed]
- Gerrits-Heybroek EM, Herrebout WM, Ulenberg SA,Wiebes JT (1978) Host plant preference of five species of small ermine moths (Lepidoptera: Yponomeutidae). Entomol Exp Appl 24:160–168 [DOI]
- Gershenson ZS, Ulenberg SA (1998) The Yponomeutinae (Lepidoptera) of the World exclusive of the Americas. Verh K Ned Akad Wet, afd Natuurkunde, 2e reeks
- Glendinning JI, Valcic S, Timmermann BN (1998) Maxillary palps can mediate taste rejection of plant allelochemicals by caterpillars. J Comp Physiol A 183:35–43 [DOI] [PubMed]
- Goldman Al van Naters WV, Lessing D, Warr CG, Carlson JR (2005) Coexpression of two functional odor receptors in one neuron. Neuron 45:661–666 [DOI] [PubMed]
- Grimes LR, Neunzig HH (1986) Morphological survey of the maxillae in last stage larvae of the suborder Ditrysia (Lepidoptera): palpi. Ann Entomol Soc Am 79:491–509
- Hansen K (1978) Insect chemoreception. In: Hazelbauer GL (ed) Taxis and behavior. Chapman & Hall, London, pp 233–292
- Hanson FE, Dethier VG (1973) Role of gustation and olfaction in food plant discrimination in the tobacco hornworm, Manduca sexta. J Insect Physiol 19:1019–1034 [DOI] [PubMed]
- Hirao T, Yamaoka K, Arai N (1976) Studies in mechanism of feeding in the silkworm, Bombyx mori L. II. Control of mandibular biting by olfactory information through maxillary sensilla basiconica. Bull Sericult Exp Stat 26:385–410
- Humpf H-U, Schreier P (1992) 3-Hydroxy-5,6-epoxy-β-ionol β-d-glucopyranoside and 3-hydroxy-7,8-dihydro-β-ionol β-d-glucopyranoside: new C13 norisoprenoid glucoconjugates from sloe tree (Prunus spinosa L.) leaves. J Agric Food Chem 40:1898–1901 [DOI]
- Johnson SN, Gregory PJ (2006) Chemically mediated host–plant location and selection by root-feeding insects. Physiol Entomol 31:1–13 [DOI]
- Kaissling KE (1976) The problem of specificity in olfactory cells In: Benz G (ed) Structure–activity relationships in chemoreception. IRL, London, pp 137–148
- Kent KS, Hildebrand JG (1987) Cephalic sensory pathways in the central nervous system of larval Manduca sexta (Lepidoptera: Sphingidae). Phil Trans R Soc Lond B 315:1–36 [DOI] [PubMed]
- Keil TA (1996) Sensilla on the maxillary palps of Helicoverpa armigera caterpillars in search of the CO2-receptor. Tissue Cell 28:703–717 [DOI] [PubMed]
- Kooi RE, van de Water TPM (1988) Yponomeuta cagnagellus an Euonymus-feeder. Proc Kon Ned Acad Wet C 91:223–232
- Kondrashov AS, Kondrashov FA (1999) Interactions among quantitative traits in the course of sympatric speciation. Nature 400:351–354 [DOI] [PubMed]
- Kristensen, NP, Skalski AW (1999) Phylogeny and Paleontology. In: Kristensen NP (ed) Lepidoptera: moths and butterflies, vol 1, pp 7–25. Evolution, systematics and biogeography, Handbook of Zoology 4 (35). de Gruyter, Berlin
- Lee JK, Selzer R, Altner H (1985) Lamellated outer dendritic segments of a chemoreceptor within wall-pore sensilla in the labial palp-pit organ of the butterfly Pieris rapae L. Cell Tissue Res 240:333–342 [DOI]
- McIver SB, Siemicki R (1984) Fine structure of pegs on the maxillary palps of adult Toxorhynchites brevipalpis Theobald (Diptera: Culicidae). Int J Insect Morphol Embryol 13:11–20 [DOI]
- Menken SBJ (1996) Pattern and process in the evolution of insect–plant associations: Yponomeuta as an example. Entomol Exp Appl 80:297–305 [DOI]
- Menken SBJ, Roessingh P (1998) Evolution of insect–plant associations: sensory perception and receptor modifications direct food specialization and host shifts in phytophagous insects. In: Howard DJ, Berlocher SH (eds) Endless forms: species and speciation. Oxford University Press, New York, pp 145–156
- Menken SBJ, Herrebout WM, Wiebes JT (1992) Small ermine moths (Yponomeuta): their host relations and evolution. Ann Rev Entomol 37:41–66 [DOI]
- Olsson SB, Linn CE Jr, Roelofs WL (2005a) The chemosensory basis for behavioral divergence involved in sympatric host shifts: I: characterizing olfactory receptor neuron classes responding to key host volatiles. J Comp Physiol A 192:279–288 [DOI] [PubMed]
- Olsson SB, Linn CE Jr, Roelofs WL (2005b) The chemosensory basis for behavioral divergence involved in sympatric host shifts II: olfactory receptor neuron sensitivity and temporal firing pattern to individual key host volatiles. J Comp Physiol A 192:289–300 [DOI] [PubMed]
- Peterson SC, Herrebout WM, Kooi RE (1990) Chemosensory basis of host-colonization by small ermine moth larvae. Proc K Ned Akad Wet Ser C Biol Med Sci 93:287–294
- Roessingh P (1992) Computer-aided analysis of chemosensory data. In: Menken SBJ, Visser JH, Harrewijn P (eds) Proceedings of 8th symposium on insect–plant relationships. Kluwer, Dordrecht
- Roessingh P, Hora KH, van Loon JJA, Menken SBJ (1999) Evolution of gustatory sensitivity in Yponomeuta caterpillars: sensitivity to the stereo-isomers dulcitol and sorbitol is localised in a single sensory cell. J Comp Physiol A 184:119–126 [DOI]
- Schoonhoven LM (1967) Some cold receptors in larvae of three Lepidoptera species. J Insect Physiol 13:821–826 [DOI]
- Schoonhoven LM, Dethier VG (1966) Sensory aspects of host–plant discrimination by lepidopterous larvae. Arch Neerl Zool 16:497–530
- Schoonhoven LM, van Loon (2002) An inventory of taste in caterpillars: each species its own key. Acta Zool Acad Sci Hung 48(Suppl 1):215–263
- Schoonhoven LM, van Loon JJA, Dicke M (2005) Insect–plant biology. Oxford University Press, Oxford
- Selzer R (1984) On the specificities of antennal olfactory receptor cells of Periplaneta americana. Chem Senses 8:375–395 [DOI]
- SPSS Inc. (2001) SPSS for Macintosh, Rel. 10.0. SPSS, Chicago
- Stange G (1992) High resolution measurement of atmospheric carbon dioxide concentrations by the labial palp organ of the moth Heliothis armigera (Lepidoptera: Noctuidae). J Comp Physiol A 172:317–324
- Stange G, Stowe S (1999) Carbon-dioxide sensing structures in terrestrial arthropods. Microsc Res Tech 47:416–427 [DOI] [PubMed]
- Steullet P, Guerin PM (1992) Perception of breath components by the tropical bont tick, Amblystomma variegatum Fabricius (Ixodidae) J Comp Physiol A 170:665–676 [DOI] [PubMed]
- Turner H, Lieshout N, van Ginkel W, Menken SBJ Molecular phylogeny of the small ermine moth genus Yponomeuta (Lepidoptera: Yponomeutidae). (submitted) [DOI] [PMC free article] [PubMed]
- van Drongelen W (1979) Contact chemoreception of host plant specific chemicals in larvae of various Yponomeuta species (Lepidoptera). J Comp Physiol 134:265–279 [DOI]
- van Drongelen W (1980) Behavioural responses of two small ermine moth species (Lepidoptera: Yponomeutidae) to plant constituents. Entomol Exp Appl 28:54–58 [DOI]
- van Poecke RMP, Dicke M (2004) Indirect defence of plants against herbivores: using Arabidopsis thaliana as a model plant. Plant Biol 6:387–401 [DOI] [PubMed]
- Visser JH, Avé DA (1978) General green leaf volatiles in the olfactory orientation of the colorado beetle, Leptinotarsa decemlineata. Entomol Exp Appl 24:738–749 [DOI]
- Visser JH, de Jong R (1988) Olfactory coding in the perception of semiochemicals. J Chem Ecol 14:2005–2018 [DOI] [PubMed]
- Wieczorek H, Wolff G (1989) The labellar sugar receptor of Drosophila. J Comp Physiol A 164:825–834 [DOI]