

Abstract
In two experiments, direct and indirect semantic priming were measured using event-related potentials. In Experiment 1, participants rated the relatedness between prime and target on a seven-point scale. In Experiment 2, participants simply read the primes and targets as they monitored for a semantic category in probe filler items. Significant direct and indirect N400 priming effects were observed in both experiments. In Experiment 1, the indirect N400 priming effect remained significant when indirectly related and unrelated word pairs were matched for participants’ explicit relatedness judgments. In both experiments, the indirect N400 priming effects were preserved when indirectly related and unrelated word pairs were matched on more global and objective measures of semantic similarity. These findings are discussed in the context of current theoretical models of semantic memory and semantic priming.
Keywords: ERP, N400, semantic, memory, priming
It has long been established that reaction times are decreased to words preceded by semantically related words (“tiger”-“stripes”) in comparison with words preceded by semantically unrelated words (“truck”-“stripes”). This is known as the semantic priming effect. One of the mechanisms often invoked to explain this effect is an automatic spread of activation through semantic memory. In this model, semantic memory is conceptualized as a network of words or concepts (Anderson, 1983; Collins & Loftus, 1975). If an individual sees or hears a word (e.g., “tiger”), its node in semantic memory will be activated, and this activation will automatically spread from this node to those of related concepts (e.g., “stripes”). If the target word corresponding to this partially preactivated or primed node is then presented, the individual’s response to that target will be facilitated. Despite the theoretical importance of this model, clear evidence for the spread of activation through semantic memory is still lacking, and several other candidate mechanisms—some controlled and some automatic—have been proposed to account for the semantic priming effect.
The two controlled or strategic processes most often discussed in the context of semantic priming tasks are expectancy generation and semantic matching. In expectancy generation, the participant internally generates predictions about which word will appear next, and priming results from the target being a member of this expectancy set. In semantic matching, the participant consciously utilizes the relationship between the word pairs to bias decision making during a particular experimental task. Both of these mechanisms are thought to occur when the time interval between the presentation of the prime and target—the stimulus onset asynchrony (SOA)—is relatively long (usually more than 400 ms) and when the total proportion of related words (the relatedness proportion: RP) within the stimulus set is more than about 33% (Neely, 1977). Semantic matching strategies are particularly likely to contribute to priming in traditional lexical decision tasks. Here, a high proportion of the targets are pronounceable nonwords derived from words that are unrelated to the prime. Participants are asked to make a decision about whether the target string of letters is a real word or a nonword (Meyer & Schvaneveldt, 1971). If the prime and target are related, the correct response must be a word. If, on the other hand, the prime and target are unrelated, there is a “nonword bias”: The correct response is likely to be a nonword. This nonword bias is thought to slow participants’ lexical decisions to targets preceded by unrelated relative to related primes (Neely & Keefe, 1989).
To reduce the effects of controlled and strategic processing on priming, several experimenters have reduced the SOA, the RP, and/or proportion of nonwords in LD tasks or have devised tasks that discourage any decision making (e.g., naming) that reduce any obvious pairing between prime and target (e.g., a “double lexical decision” in which participants make decisions to both prime and target simultaneously or a “single presentation lexical decision” in which participants make a decision to each word as it appears; Chwilla & Kolk, 2002; for a review, see Hutchinson, 2003) or that attempt to preclude conscious processing of the prime word through masking (Kiefer, 2002; Kiefer & Brendel, 2006; Misra & Holcomb, 2003). The semantic priming effect survives despite these manipulations, providing evidence for automatic processing. However, this still does not provide conclusive evidence for spreading activation, as other automatic mechanisms of priming have been proposed. In compound cue theory (Ratcliff & McKoon, 1988), rather than activating stored concepts in long-term memory, the prime and target are thought to join together in short-term memory. If the compound formed from a semantically related prime and target is more familiar than a compound formed from a semantically unrelated prime and target, priming will occur. Semantic feature overlap, most often invoked in the context of distributed memory models (Masson, 1991, 1995), asserts that when a prime is presented, it activates its semantic features in long-term memory. If a target is then presented that shares a sufficient number of semantic features with the prime (e.g., “dog” and “cat” both are small animals with four legs), priming will occur as a result of the preactivation of these overlapping features.
One of the strongest pieces of evidence supporting the spread of activation through semantic memory is the demonstration of an indirect or mediated semantic priming effect. In traditional semantic priming tasks, the prime and the target are directly associatively or categorically related. In indirect priming, the prime is not directly related to the target but, rather, is associated with another word that is not explicitly presented, which is, in turn, associated with the target. For example, the words “lion” and “stripes” are not directly semantically related, but are only connected through the third, unseen mediator “tiger.” Indirect priming is thought to be best explained by spreading activation theory (McNamara 1992; for a review, see Neely, 1991): The mediating word is thought to be activated by the prime; this spread of activation, in turn, activates the target. In contrast, other candidate mechanisms for automatic priming do not easily explain an indirect semantic priming effect. In compound cue theory, indirectly related compounds would not be familiar; targets would only be primed by indirectly related words if these word pairs happened to be weakly but directly related (McKoon & Ratcliff, 1992). In addition, semantic feature overlap is an unlikely mechanism, as indirectly related word pairs tend not to share common semantic features.
Indirect semantic priming effects, however, have been difficult to demonstrate in behavioral studies using traditional lexical decision tasks. One explanation for this is that controlled processes such as semantic matching are as unlikely to find an association between prime and target to indirectly related word pairs as for unrelated word pairs. The nonword bias on indirectly related word pairs therefore cancels out any indirect priming due to automatic spreading activation (see discussion by Neely, 1991). Some evidence for this explanation comes from a study by Balota and Lorch (1986) that reported a significant indirect priming effect during a word pronunciation task (where there was no binary decision) but not for lexical decision. In addition, McNamara and Altaribba (1988) reported an indirect priming effect in double- and single-presentation lexical decision tasks that both discourage controlled semantic matching. This indirect priming effect, however, was only seen when lists did not contain directly related word pairs. They suggested that, in the presence of the directly related word pairs, indirectly related pairs were treated as unrelated because participants used the most obvious semantic relationships to speed their lexical decisions. Chwilla and Kolk (2002) replicated this list composition effect. Using a double lexical decision task, they found indirect priming only when lists excluded directly related word pairs. Moreover, in a second experiment, when lists also included three-step pairs (e.g., “mane”-“stripes” via “lion” and “tiger”), they demonstrated priming for indirectly related pairs (e.g., “lion”-“stripes”). Priming for three-step pairs, however, was only seen in lists that excluded both directly related and indirectly related pairs. Once again, this list effect suggested the use of strategy in seeking the clearest relationships between word pairs and using such relationships to speed decisions.
Individual differences in prefrontal functioning have also recently been shown to influence the amount of semantic priming during lexical decision. Kiefer, Ahlegian, and Spitzer (2005) reported increased indirect semantic priming for individuals with lower working memory capacity. It is possible that individuals with lower working memory capacity were less able to efficiently use semantic matching strategies, and therefore their reaction times were sensitive to automatic spreading activation.
To summarize, reaction time studies have revealed some evidence for indirect semantic priming when tasks are used that encourage automatic processing and reduce strategic processing, but these effects have been small and inconsistent. Event-related potentials (ERPs) are a direct, online measure of brain activity with a temporal resolution in the order of milliseconds, which might provide a more sensitive index of automatic processes such as spreading activation. The N400 component, a negative-going waveform peaking around 400 ms after target onset, has been shown to be highly sensitive to semantic relationships between words. Words preceded by semantically related words elicit a smaller, less negative N400 than words preceded by unrelated words (Bentin, McCarthy, & Wood, 1985; Rugg, 1985). This modulation is known as the N400 effect (Kutas & Hillyard, 1980) and is thought to reflect the difficulty with which a word can be semantically integrated into its preceding context (Holcomb, 1993).
When examining indirect semantic priming through traditional lexical decision tasks, some ERP researchers have reported significant indirect N400 priming effects (Kiefer, Weisbrod, Kern, Maier, & Spitzer, 1998; Weisbrod et al., 1999), but others have not demonstrated this effect (Hill, Strube, Roesch-Ely, & Weisbrod, 2002). Other studies have compared standard lexical decision tasks with single and double lexical decision tasks and have also manipulated other experimental parameters in an attempt to reduce strategic processing. Silva-Pereyra et al. (1999) reported a significant N400 effect to indirectly related targets in a standard lexical decision task, but only when RP was low. Chwilla, Kolk, and Mulder (2000) found no indirect N400 priming effect when lists also contained directly related items, and only a small effect in the second half of the experiment when lists contained only indirectly related and unrelated items. When a double lexical decision task was used (primes and targets presented simultaneously for 400 ms), a significant indirect N400 priming effect was seen in both pure and mixed lists.
Taken together, these ERP experiments generally support the behavioral findings in suggesting that indirect priming can be seen when experimental conditions such as a low RP, a short SOA, and the reduction of obvious pairing between prime and target discourage the use of strategies. The assumption is that these experimental manipulations reduce the influence of strategic processes such as semantic matching and allow the effects of spreading activation to emerge. However, all ERP studies and most behavioral studies of indirect priming to date have used some form of the lexical decision task where it is difficult to entirely eliminate the effects of strategic processes because a behavioral response is required for each trial. The current study aimed to explore mechanisms of indirect priming by contrasting the effects of two different tasks on N400 modulation to targets preceded by directly related, indirectly related, and unrelated primes. An SOA of 350 ms was used to encourage automatic processing. In Experiment 1, participants were explicitly instructed to rate the semantic relationships between word pairs on each trial on a seven-point scale. This allowed us to determine whether participants’ subjective ratings of relationships between the indirectly related word pairs could be dissociated from their ERP responses. In addition to examining ERPs in relation to participants’ explicit ratings, we also examined ERPs in relation to more objective measures of semantic association (using the Latent Semantic Analysis, LSA). In Experiment 2, participants were not required to make any decision to targets that were preceded by the different types of prime, but simply monitored for a semantic category in probe filler items. Again, we determined if N400 modulation to the indirectly related targets could be dissociated from LSA measures of semantic association. The presence of an indirect N400 priming effect to indirectly related word pairs that were subjectively perceived as being unrelated, and were objectively nonassociated, would provide strong evidence for an automatic spread of activation.
EXPERIMENT 1
In this experiment, participants were asked to rate the relatedness of word pairs on a seven-point scale as we measured ERPs to targets in three types of word pairs: directly related pairs (e.g., “tiger”-“stripes”), indirectly related pairs (e.g., “lion”-“stripes”), and unrelated pairs (e.g., “truck”-“stripes”). We focused on three questions. First, we were interested in whether indirectly related word pairs would receive ratings that were in between those of the directly related and unrelated pairs. Ratings in between unrelated and directly related word pairs would suggest the operation of some strategy whereby participants’ search through semantic memory led them to successfully detect and classify the indirect relationships as being less semantically related than the directly related pairs and more semantically related than the unrelated pairs. On the other hand, ratings of the indirectly related word pairs that were closer to the unrelated word pairs than to the directly related word pairs would suggest that participants were not engaging in such a strategy.
Second, we were interested in the precise relationship between these ratings and the modulation of the N400 component. If indirectly related targets evoked N400s in between those of directly related and unrelated targets, even when participants’ ratings did not show a similar linear pattern, this would suggest that explicit semantic matching might not entirely explain the indirect N400 priming effect, leaving open the possibility that this effect was at least in part mediated by automatic, nonstrategic mechanisms. Some evidence that explicit semantic matching may not be effective in increasing indirect priming comes from an ERP experiment by Chwilla et al. (2000). Here, participants were informed of the presence of indirectly related items and were asked to try to use that information to speed their lexical decisions. Although the semantic matching instruction had the effect of enlarging overall N400 amplitudes and broadening their time windows, no N400 effect was observed to indirectly related targets. In other words, even when participants were instructed to engage in semantic matching they did not find it an effective strategy to increase the degree of indirect N400 priming.
Our third question was whether more global and objective measures of semantic similarity could explain the modulation of the N400 to indirectly related word pairs. It has been argued that indirectly related word pairs may, in fact, share semantic similarities and that any attenuation of the N400 to such indirectly related targets might result from such similarities. This potential explanation for indirect priming is important theoretically because it would provide a mechanism by which indirect priming could occur through automatic mechanisms other than the spread of activation, that is, through forming a compound cue or through sharing common semantic features. To examine the relationship of the indirectly related word pairs in more detail, we submitted them to a LSA. The LSA not only examines co-occurrence frequency, but also takes into account the similarity of contexts in which words occur, yielding global semantic similarity values (SSVs; Landauer & Dumais, 1997; Landauer, Foltz, & Laham, 1998). Chwilla and Kolk (2002) found that the mean SSVs derived using a LSA of word pairs that had both two-step indirect relationships (e.g., “lion”-“stripes” via “tiger”) and three-step indirect relationships (e.g., “mane”-“stripes” via “lion” and “tiger”) were greater than those of unrelated word pairs. They suggested that priming of these word pairs might not necessarily require the activation of a mediating concept, but could occur through the formation of a compound cue or through semantic feature overlap. To determine whether any indirect priming in the current stimulus set was driven by greater SSVs in the indirectly related word pairs relative to unrelated word pairs, we repeated these ERP analyses after matching for SSVs.
Methods
Participants
Eighteen (9 male and 9 female; mean age: 20) Tufts undergraduate students were paid for participation. All participants were right-handed (as assessed by a modified version of the Edinburgh Handedness Inventory; Oldfield, 1971) had normal or corrected-to-normal vision, were native speakers of English, had no history of traumatic head injury, and were not taking any medication acting on the central nervous system. Written consent was obtained from all participants according to the established guidelines of the Tufts University Institutional Review Board.
Design and Stimulus Materials
Two hundred and ten word triplets were developed such that target words (e.g., “stripes”) were paired with directly related primes (e.g., “tiger”) and indirectly related primes (e.g., “lion”). Directly related word pairs constituted a mixture of categorically related word pairs in which prime and target were both exemplars of a particular category (e.g., “cat”-“mouse”), associatively related word pairs in which prime and target tend to co-occur in speech but do not necessarily share semantic features (e.g., “ring”-“finger”), and functionally related word pairs in which prime and target are strongly associated by their co-occurrence in performing a particular action (e.g., “hammer”-“nail”). Most directly related word pairs were concrete nouns but some were abstract (e.g., “question”-“answer”), and sometimes nouns were paired with familiar adjectives (e.g., “sky”-“blue”), or verbs (e.g., “sweep”-“broom”).
Indirectly related primes were categorically or associatively related to the prime of the directly related word pairs (e.g., “lion” is associatively related to “tiger”) but they were judged to show no categorical, associative, or functional relationship to the target of the directly related word pairs (“lion” shows no associative, functional, or categorical relationship to “stripes”). The indirectly related pairs of 45 of these triplets were gathered from several published studies of indirect priming (e.g., Balota & Lorch, 1986; McNamara, 1992; Weisbrod et al., 1999). An additional 165 triplets were developed for the purposes of this study. To verify that, in the indirectly related pairs, the mediator could be identified objectively, 18 Tufts undergraduates who did not participate in the ERP study were given the final list of 210 indirectly related pairs and asked to produce a word or concept that would link the two words. For all indirectly related pairs, either the precise mediating word (66%) or the mediating concept (33%) was correctly identified.
Finally, unrelated word pairs were constructed by pairing a target with a prime of a related word pair from another list (see counterbalancing below). These unrelated word pairs were judged by the experimenter as showing no categorical, associative, or functional relationship to their target. An example stimulus set is given in Table 1.
Table 1.
Example of Stimuli, Word Length and Frequency as a Function of Priming Condition
| Example | Mean (SD) frequency | Mean (SD) word length | |
|---|---|---|---|
| Directly related primes | Tiger | 96.39 (149.58) | 5.11 (1.51) |
| Indirectly related primes | Lion | 69.05 (155.87) | 5.60 (1.66) |
| Unrelated primes | Truck | 84 (123.49) | 5.30 (1.50) |
| Target words | Stripes | 93.14 (469.6) | 5.09 (1.46) |
Target words were counterbalanced across three lists such that, in each list, there were 70 directly related pairs, 70 indirectly related pairs, and 70 unrelated pairs. Thus, across all participants, the same targets were seen in all three conditions. There were no significant differences in the frequency (Kučera & Francis, 1967) of prime words across the three conditions (direct vs. indirect, t[418] = 1.834, p>.05; indirect vs. unrelated, t[418] = -1.090, p>.1; direct vs. unrelated, t[418] = 0.925, p>.1). Mean frequencies and word lengths are shown in Table 1.
ERP Procedure
Participants were asked to rate the relatedness of word pairs on a scale of 1 to 7 (1 = not related, 7 = highly related). They were not given any information about the nature of the relationships between words, and were simply asked to go with their first instincts and not to dwell on any single word pair. Participants entered their ratings on a number pad immediately after reading the second word in the pair.
Each participant was given 30 practice trials at the start of the experiment and was randomly assigned to one of the three lists used for counterbalancing. Participants sat in a comfortable chair in a dimly lit room, separate from the experimenter and computers. Each trial (one word pair) began with presentation of a fixation point at the center of the screen for 500 ms, followed by a 500 ms blank screen, followed by the first word. Each word appeared on the screen for 250 ms with an interstimulus interval (ISI) of 100 ms separating words. After the second word in the pair, the screen was blank for 1250 ms (to avoid contamination of the data with vertical eye movement as participants prepared to enter their responses) and then a nonlinguistic cue remained on the screen until a response was entered.
Electrophysiological Recording
Twenty-nine active tin electrodes were held in place on the scalp by an elastic cap (Electro-Cap International, Inc., Eaton, OH; see Figure 1). Several of these electrodes were placed in standard International 10-20 system locations. These included five sites along the midline (FPz, Fz, Cz, Pz, and Oz) and eight lateral (four over each hemisphere) sites (F3/F4, C3/C4, T3/T4, and P3/P4). Eight extended 10-20 system sites were also used (FC1/FC2, FC5/FC6, CP1/CP2, and CP5/CP6) and eight additional 10-20 sites were altered to form a circle around the perimeter of the scalp. These altered sites included FP1′/FP2′ (33% of the distance along the circle between T3/T4), F7′/F8′ (67% of the distance between FPz and T3/T4), T5′/T6′ (33% of the distance between T3/T4 and Oz), and O1′/O2′ (67% of the distance between T3/T4 and Oz). Electrodes were placed below the left eye and at the outer canthus of the right eye to monitor vertical and horizontal eye movements. Electrodes were also placed over the left mastoid (reference) and over the right mastoid (recorded actively to monitor for differential mastoid activity). All EEG electrode impedances were maintained below 5 k (impedance for eye electrodes was less than 10 kΩ).
Figure 1.
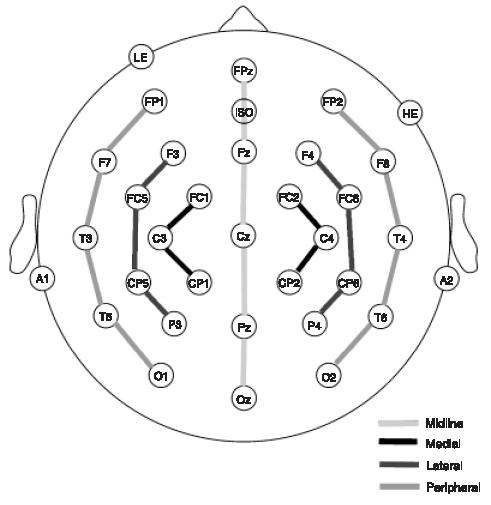
The electrode montage used in Experiments 1 and 2.
The EEG signal was amplified by an Isolated Bioelectric Amplifier System Model HandW-32/BA (SA Instrumentation Co., San Diego, CA) with a bandpass of 0.01 to 40 Hz and was continuously sampled at 200 Hz by an analogue-to-digital converter. The stimuli presented to participants and their behavioral responses were simultaneously monitored by the digitizing computer.
ERP Data Analysis
Averaged ERPs were formed off-line from trials free of ocular and muscular artifact. The averaged ERPs elicited by target words were quantified by calculating the mean amplitude values (relative to a 100 ms prestimulus baseline) for the voltage points 300-500 ms and 600-800 ms after stimulus onset.
The resulting data for each time window were analyzed with analyses of variance (ANOVAs) for repeated measures. ERPs were analyzed separately for midline and lateral electrode sites. The ANOVA for midline electrodes included an electrode site factor (from anterior to posterior: FPz, Fz, Cz, Pz, Oz). To examine distributional differences across the scalp in both eft-right and anterior-posterior dimensions, we carried out ANOVAs comparing right- and left-sided parasagittal columns: medial, lateral, and peripheral. The medial analysis had three levels of electrode site (FC1/FC2, C3/C4, CP1/CP2) and two levels of hemisphere. The lateral analysis had four levels of electrode site (F3/F4, FC5/FC6, CP5/CP6, P3/P4) and two levels of hemisphere. The peripheral analysis had five levels of electrode site (FP1′/FP2′, F7F8′, T3/T4, T5′/T6′, O1′/O2′) and two levels of hemisphere.
Three levels of priming condition were within-subject variables in initial ANOVAs (in addition to the within-subject variables described above). Significant main effects and interactions in these ANOVAs were followed up by planned simple effects ANOVAs that allowed comparisons between each condition.
To protect against Type 1 error resulting from violations of sphericity, the Geisser-Greenhouse correction (Greenhouse & Geisser, 1959) was applied to all repeated measures with more than one degree of freedom in the numerator. In these cases, we report the original degrees of freedom and the corrected probability level. In reporting whether or not a particular effect was significant, we used a standard significance level of alpha = .05 as, in all cases, we were testing specific a priori hypotheses. In addition, we report significance at p<.01 and p<.001 for each ANOVA in the results tables.
Results
Fewer than 9% of total trials were rejected for artifact. Grand-average ERPs elicited by target words in the three priming conditions are shown at all electrode sites in Figure 2. Beginning 300 ms after target onset, a clear separation of the three priming conditions can be seen, continuing through approximately 800 ms after target onset. Between 300 and 500 ms, a negative-going component with a peak amplitude at around 400 ms—the N400—was observed. The N400 was followed by a late positive component (LPC) between 600 and 800 ms. Below we report statistical analyses comparing ERPs elicited by target words within these time windows.
Figure 2.
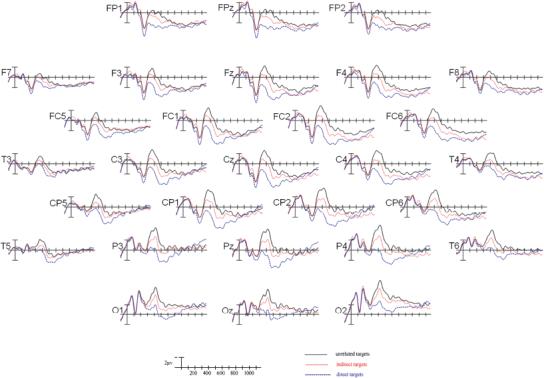
Grand average ERPs time-locked to target onset in all three priming conditions in the relatedness ratings task (Experiment 1).
ERPs: Effects of Priming Condition
300-500 ms: the N400
During the 300-500 ms time window, an overall ANOVA revealed a significant main effect of Priming Condition at all electrode columns (midline: F[2,34] = 44.813, p<.001; medial: F[2,34] = 43.077, p<001; lateral: F[2,34] = 36.380, p<.001; peripheral: F[2,34] = 27.041, p<.001) and significant Priming Condition × Electrode Site interactions at all columns (midline: F[8,136] = 3.217, p<.05; medial: F[4.68] = 4.006, p<.05; lateral: F[6,102] = 4.926, p<.01; peripheral: F[8,136] = 4.378, p<.01). Planned simple effects ANOVAs were carried out to determine how the three priming conditions differed from each other.
Comparisons between directly related targets and unrelated targets revealed a significant main effect of Priming Condition at all columns (Table 2Ai, left), and significant Priming Condition × Electrode Site interactions at all columns (Table 2Ai, right), reflecting the posterior distribution of this effect (see Figure 3A,B). These differences were larger over the right hemisphere, as evidenced by significant Priming Condition × Hemisphere interactions at lateral, F(1,17) = 13.159, p<.01, and peripheral, F(1,17) = 6.752, p<.05, columns.
Table 2.
Simple Effects ANOVAs at N400 and LPC Time Windows, Experiment 1
| Main effect of priming |
Interactions between priming and electrode site |
|||
|---|---|---|---|---|
| Degrees of freedom | F value | Degrees of freedom | F value | |
| A. Directly related vs. unrelated | ||||
| i. N400: 300-500 ms. | ||||
| Midline | 1,17 | 54.157*** | 4,68 | 6.780** |
| Medial | 1,17 | 53.381*** | 2,34 | 8.115** |
| Lateral | 1,17 | 44.705*** | 3,51 | 9.516*** |
| Peripheral | 1,17 | 33.746*** | 4,68 | 6.960** |
| ii. LPC: 600-800 ms. | ||||
| Midline | 1,17 | 11.668** | 4,68 | 2.232 |
| Medial | 1,17 | 12.566** | 2,34 | 6.954** |
| Lateral | 1,17 | 10.730** | 3,51 | 4.879* |
| Peripheral | 1,17 | 9.343** | 4,68 | 8.817** |
| B. Indirectly related vs. unrelated | ||||
| i. N400: 300-500 ms. | ||||
| Midline | 1,17 | 16.115** | 4,68 | 1.303 |
| Medial | 1,17 | 16.461** | 2,34 | 1.777 |
| Lateral | 1,17 | 14.555** | 3,51 | 3.508* |
| Peripheral | 1,17 | 11.843** | 4,68 | 1.239 |
| ii. LPC: 600-800 ms. | ||||
| Midline | 1,17 | 2.523 | 4,68 | 0.621 |
| Medial | 1,17 | 3.755 | 2,34 | 0.095 |
| Lateral | 1,17 | 2.248 | 3,51 | 0.177 |
| Peripheral | 1,17 | 1.258 | 4,68 | 0.565 |
| C. Directly related vs. indirectly related | ||||
| i. N400: 300-500 ms. | ||||
| Midline | 1,17 | 81.379*** | 4,68 | 1.840 |
| Medial | 1,17 | 71.108*** | 2,34 | 2.446 |
| Lateral | 1,17 | 56.804*** | 3,51 | 2.052 |
| Peripheral | 1,17 | 38.262*** | 4,68 | 3.961* |
| ii. LPC: 600-800 ms. | ||||
| Midline | 1,17 | 6.308* | 4,68 | 2.813 |
| Medial | 1,17 | 5.226* | 2,34 | 5.104* |
| Lateral | 1,17 | 5.986* | 3,51 | 2.458 |
| Peripheral | 1,17 | 7.967* | 4,68 | 5.614* |
p<.05
p<.01
p<.001.
Figure 3.
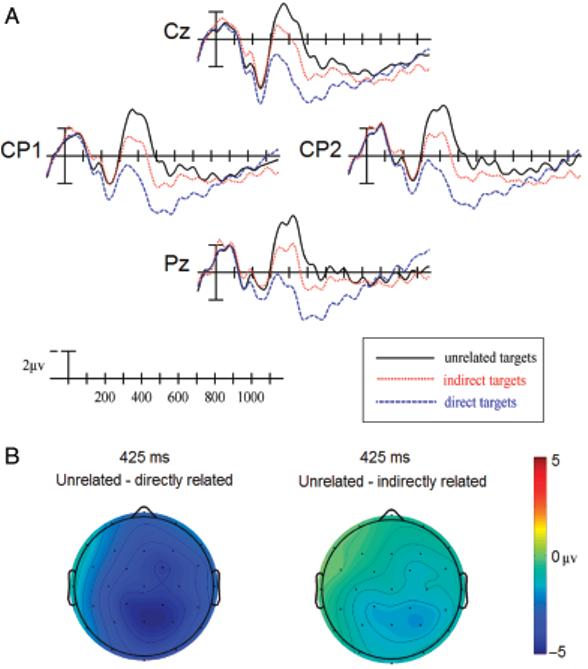
A: ERPs to target words at four centro-parietal sites in Experiment 1. B: Scalp distribution of comparisons between unrelated-directly related and unrelated-indirectly related in Experiment 1.
In comparing indirectly related targets with unrelated targets, significant main effects of Priming Condition were seen at all electrode columns (Table 2Bi, left), and significant Priming Condition × Electrode Site interactions were found at the lateral column (Table 2Bi, right), indicating a posteriorly distributed effect (see Figure 3A,B).
The amplitude of the N400 to the indirectly related targets appeared to be in between that of the unrelated and directly related targets. This was confirmed by a comparison between the indirectly and directly related targets that revealed a significant main effect of Priming Condition at all electrode columns (Table 2Ci, left), and a significant Priming Condition × Electrode Site interaction at the peripheral column (Table 2Ci, right).1
600-800 ms: the LPC
An overall ANOVA in this time window revealed significant main effects of Priming Condition at all columns (midline: F[2,34] = 7.634, p<.01; medial: F[2,34] = 7.905, p<.01; lateral: F[2,34] = 7.026, p<.01; peripheral: F[2,34] = 6.531, p<.01), and Priming Condition × Electrode Site interactions at medial, F(2,34) = 3.765, p<.05, peripheral, F(8,136) = 5.329, p<.01, columns.
Directly related targets appeared to elicit significantly more positive LPCs than unrelated targets, particularly at posterior sites. This was confirmed by a significant main effect of Priming Condition at all columns (Table 2Aii, left) and significant priming condition by electrode site interactions at all but the midline column (Table 2Aii, right). This effect also appeared more distinct over the right hemisphere. This impression was confirmed by significant Priming Condition × Hemisphere interactions at the lateral electrode column, F(1,17) = 5.987, p<.05.
In comparing indirectly related to unrelated targets, there were no significant main effects of Priming Condition (Table 2Bii, left) or significant Priming Condition × Electrode Site interactions (Table 2Bii, right). However, significant Priming Condition × Hemisphere interactions at the medial, F(1,17) = 7.188, p<.05, and lateral, F(1,17) = 6.158, p<.05, columns and a significant Priming Condition × Electrode Site × Hemisphere interaction at the peripheral column, F(4,68) = 2.900, p<.05, reflected a more positive LPC to indirectly related than to unrelated targets over posterior and right-lateralized sites.
Finally, the comparison between the directly related and indirectly related targets revealed significant main effects of Priming Condition at all columns (Table 2Cii, left) and interactions between Priming Condition and Electrode Site at medial and lateral columns (Table 2Cii, right), indicating that directly related targets produced more positive LPCs than did indirectly related targets, particularly at posterior sites.
Behavioral Ratings and Their Contribution to the Indirect N400 Effect
Participants used all seven points on the ratings scale when entering their decisions about relatedness. The mean ratings across all participants for directly related words, indirectly related words, and unrelated words respectively were 5.7 (SD: 1.2), 4.0 (SD: 0.5), and 1.6 (SD: 0.3). An ANOVA conducted with mean relatedness ratings revealed a significant effect of Priming Condition, F(2,34) = 4.191, p<.05. Follow-up pairwise comparisons indicated that this difference could be accounted for primarily by significantly higher relatedness ratings in the direct priming condition than in the unrelated condition, F(1,17) = 6.758, p<.05. There was also a trend toward higher ratings to the directly related than to the indirectly related word pairs, F(1,17) = 4.349, p<.1. However, no significant difference was found between ratings to the indirectly related and unrelated word pairs, F(1,17) = 1, p>.3.
To determine whether differences in ratings alone could explain the N400 effect to indirectly related targets, we matched the relatedness ratings of indirectly related and unrelated targets by including only those word pairs that had been rated a 4 or lower (56% of the total indirectly related items; mean rating indirect = 1.58; mean rating unrelated = 1.47) so that there was no significant difference between them, t(17) = 0.822, p = .42. When only these targets were included in an ANOVA comparing the mean amplitude in the N400 time window between the unrelated and indirectly related conditions, significant main effects of priming condition were seen at all electrode columns (midline: F[1,18] = 10.326, p<.01; medial: F[1,18] = 10.275, p<.01; lateral: F[1,18] = 7.958, p<.05; peripheral: F[1,18] = 5.984, p<.05; see Figure 4A). Thus, the indirect N400 priming effect was preserved in these analyses.
Figure 4.
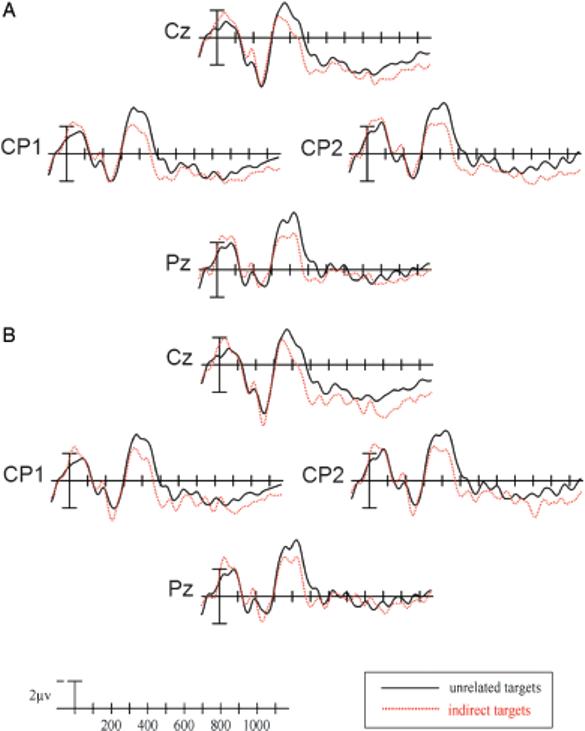
A: Indirectly related and unrelated targets matched by relatedness ratings in Experiment 1. B: Indirectly related and unrelated targets matched by LSA in Experiment 1.
Global Semantic Similarity Values and Their Contribution to the Indirect N400 Effect
We examined all word pairs on a term-to-term basis with an LSA using tasaALL space (Landauer & Dumais, 1997; Landauer et al., 1998). This yielded SSVs for all word pairs except for three word pairs that generated missing values and were left out of any subsequent analysis. Mean SSVs for directly related, indirectly related, and unrelated pairs were as follows: 0.40 (SD: 0.22), 0.23 (SD: 0.17), and 0.08 (SD: 0.07). In keeping with the findings of Chwilla and Kolk (2002), SSVs for indirectly related word pairs were significantly larger than those of unrelated pairs, t(205) = 12.1278, p<.001. Directly related word pairs had significantly larger SSVs than unrelated pairs, t(205) = 20.099, p<.001, or indirectly related pairs, t(205) = 8.434, p<.001).
To determine whether these differences alone could explain the N400 effect to indirectly related targets, we removed more than half of the word pairs such that the mean SSV of the indirectly related word pairs matched that of the unrelated pairs (indirect: 0.08 [SD: 0.05]; unrelated: 0.08 [SD: 0.07]). Simple effects ANOVAs comparing mean N400 amplitudes of these indirectly related word pairs and their unrelated control pairs revealed significant main effects of priming condition at all electrode columns (midline: F[1,18] = 10.649, p<.01; medial: F[1,18] = 10.949, p<.01; lateral: F[1,18] = 10.388, p<.01; peripheral: F[1,18] = 7.542, p<.05; see Figure 4B), indicating that there was still an indirect semantic N400 priming effect.
Discussion
Indirectly related targets evoked an N400 in between that of directly related and unrelated targets. This N400 was followed by an LPC effect—not a surprising result, because participants were making explicit judgments about the stimuli, and post-N400 positivities, presumably associated with late decision-based processes, are frequently seen in word priming studies (Donchin & Coles, 1988; Holcomb, 1988). The pattern and distribution of differences between the three priming conditions found in this later time window seem to point to carryover effects from the N400 time window. Targets that elicited less negative N400s also elicited more positive LPCs and vice versa.
These N400 effects indicate that unrelated target words were most difficult to semantically integrate, directly related targets were the easiest, and indirectly related targets were in between the two. Given the explicit nature of the task, a likely explanation for this pattern of findings would be that the participants were using a semantic matching strategy whereby they perceived indirectly related word pairs to be half as related as directly related items and used this information to facilitate semantic integration. However, the behavioral data indicated that participants did not perceive indirectly related pairs as being significantly more related than unrelated pairs. The wide variability in ratings responses to indirectly related word pairs suggests these indirect relationships are more difficult to classify, even when participants are asked to explicitly attend to such relationships. Most interestingly, even when we matched ratings between the indirectly related and unrelated word pairs, we still observed a significant indirect N400 priming effect. It therefore seems unlikely that an explicit strategy of looking for relationships between prime and target is the only explanation for the pattern of N400 modulation observed in this experiment.
Having ruled out explicit strategic processes as entirely accounting for our indirect N400 priming effect, we next considered which automatic mechanism might best explain our results. Compound cue and semantic feature overlap both predict that semantic priming will occur when word pairs share common contexts or semantic features. However, we observed an indirect N400 effect even when such global semantic similarity, extracted through a LSA, was matched between the unrelated and indirectly related word pairs, suggesting automatic spreading activation as a potential mechanism. This explanation is also consistent with the graded modulation of the N400 across the three conditions. A spreading activation theory predicts decreased facilitation with increased associative distance and therefore explains why the N400 amplitude was larger to indirectly primed words than to directly primed words.
In sum, even when indirectly related word pairs were perceived as unrelated and were matched on global similarity values, targets still evoked a significant N400 effect that was smaller than the N400 effect evoked by directly related targets. This provides supportive evidence for automatic spreading activation theories. However, the explicit task performed in this experiment makes it difficult to entirely rule out the contribution of strategic processes. To reduce such strategic processes, we carried out Experiment 2, in which participants performed an implicit task that did not require them to attend to the relationship between prime and target. The presence of an indirect priming N400 effect under such conditions would provide further evidence for the automatic spread of activation.
EXPERIMENT 2
As noted above, all ERP experiments examining indirect priming thus far have utilized various forms of the lexical decision task. Although the use of single presentation lexical decision (e.g., Silva-Pereyra et al., 1999) and double lexical decision (e.g., Chwilla et al., 2000) may serve to attenuate strategic processing, it is still possible that the requirement to make a decision for all word pairs could obscure automatic priming processes.
In Experiment 2, participants viewed the same set of stimuli as for Experiment 1 but performed an implicit semantic categorization task (Misra & Holcomb, 2003) in which they were instructed to monitor all words (primes and targets) for occasional exemplars from a designated category of probe words (food words), and to press a button when such items were detected. No response was required to other (nonfood) words. The use of this task ensured that participants attempted to process both the primes and targets at a deep semantic level and that they focused equal attentional resources on all words being presented. A similar stepwise reduction in N400 amplitude in such an implicit task across unrelated, indirectly related, and directly related word pairs would suggest the operation of automatic processes.
To determine which theoretical account best explained any automatic indirect semantic priming N400 effect, we once again carried out a LSA with our stimuli. If an indirect N400 priming effect held up after indirectly related and unrelated word pairs were matched on their semantic similarity values, this would provide more compelling evidence for automatic spreading activation theory.
Methods
Participants
Eighteen Tufts undergraduate students who did not take part in Experiment 1 (6 male and 12 female; mean age: 20) were paid for participation and fulfilled the same criteria as those in Experiment 1.
Stimuli
Word pairs were the same as those in Experiment 1, except that nine word pairs that contained food-related words were removed. A total of 201 directly related, indirectly related, and unrelated word pairs were counterbalanced across three lists so that each list contained 67 of each. A probe condition for the semantic categorization task was added; probe items were always food words and comprised 20% of the total stimulus set. Half of the probe items appeared in the prime position, and half appeared in the target position. Filler words were added to complete food word pairs, and they were always unrelated to the food words.
ERP Procedure, Recording, and Data Analysis
Participants were told that several words would appear in a row, and that their task was to simply press a button whenever they saw a food word. Because, unlike in Experiment 1, participants were not required to respond to each word pair, trials were presented in a slightly different fashion. As in Experiment 1, each trial began with a fixation point centered on the screen for 500 ms, followed by a 500-ms blank screen. Prime and target words were then each presented for 250 ms, and the ISI between the prime and target was 100 ms. This trial was immediately followed by a second word pair trial with a 900-ms ISI between the first target and the onset of the second prime to avoid component overlap. In between every two trials, a 900-ms blank screen was followed by a blink cue that remained on the screen for 3000 ms. Participants were given 18 practice trials (36 word pairs) and were given feedback on their performance prior to beginning the experiment.
EEG recording and data analysis were identical to that described in Experiment 1.
Results
Less than 6% of trials were rejected for artifact. Participants correctly identified 96.94% (SD = 5.18%) and 97.78% (SD = 2.56%) of the probe items in the prime and target positions, respectively, indicating that they were attending to the task.
300-500ms: the N400
As in Experiment 1, directly related targets appeared to elicit the smallest N400, unrelated targets the largest, and indirectly related targets in between the two (see Figure 5). These effects appeared to be more robust at posterior than anterior sites.
Figure 5.
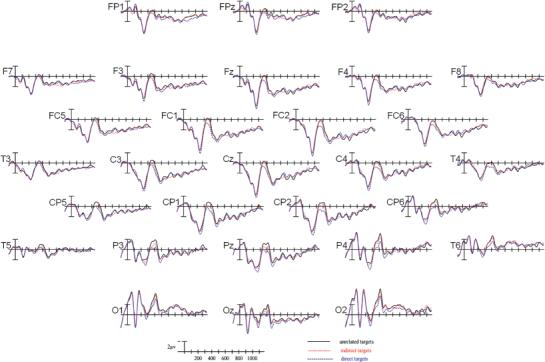
Grand average ERPs time-locked to target onset in all three priming conditions in the semantic categorization task (Experiment 2).
An overall ANOVA revealed significant main effects of priming condition at all electrode columns (midline: F[2,34] = 4.765, p<.05; medial: F[2,34] = 4.478, p<.05; lateral: F[2,34] = 4.688, p<.05; peripheral: F[2,34] = 5.363, p<.05). The contributions of the three priming conditions to these effects were further explored through planned simple effects ANOVAs.
In comparing directly related targets to unrelated targets, a significant main effect of Priming Condition was seen at all columns (Table 3A, left). This effect was posteriorly distributed, as evidenced by significant Priming Condition × Electrode Site interactions at lateral and peripheral columns (Table 3A, right, see Figure 6A,B). No interactions between Priming Condition and Hemisphere were seen, all Fs<1.200, all Ps>.3.
Table 3.
Simple Effects ANOVAs at N400 Time Window, Experiment 2
| Main effect of priming |
Interactions between priming and electrode site |
|||
|---|---|---|---|---|
| Degrees of freedom | F value | Degrees of freedom | F value | |
| A. Directly related vs. unrelated | ||||
| Midline | 1,17 | 6.342* | 4,68 | 1.237 |
| Medial | 1,17 | 6.569* | 2,34 | 1.595 |
| Lateral | 1,17 | 6.284* | 3,51 | 3.932* |
| Peripheral | 1,17 | 6.550* | 4,68 | 3.579* |
| B. Indirectly related vs. unrelated | ||||
| Midline | 1,17 | 0.891 | 4,68 | 0.409 |
| Medial | 1,17 | 1.030 | 2,34 | 1.397 |
| Lateral | 1,17 | 1.090 | 3,51 | 1.463 |
| Peripheral | 1,17 | 0.490 | 4,68 | 1.331 |
| C. Directly related vs. indirectly related | ||||
| Midline | 1,17 | 6.451* | 4,68 | 0.439 |
| Medial | 1,17 | 6.081* | 2,34 | 0.620 |
| Lateral | 1,17 | 6.309* | 3,51 | 0.650 |
| Peripheral | 1,17 | 10.123** | 4,68 | 0.603 |
p<.05
p<.01.
Figure 6.
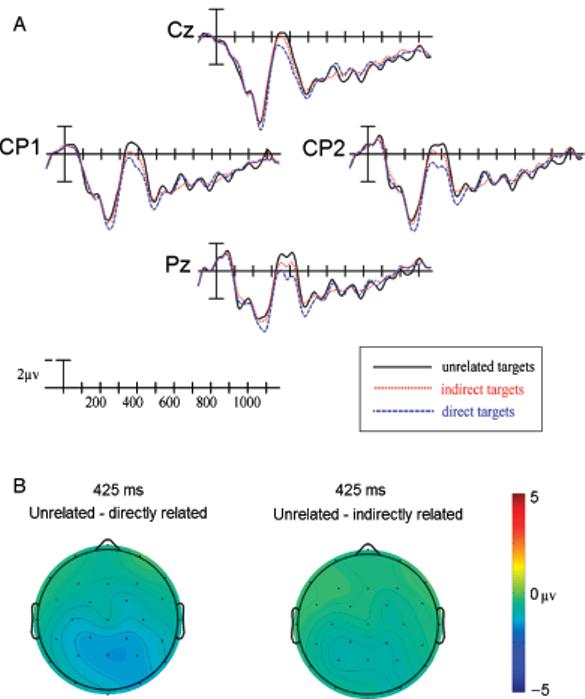
A: ERPs to target words at four centro-parietal sites in Experiment 2. B: Scalp distribution of comparisons between unrelated-directly related and unrelated-indirectly related in Experiment 2.
In comparing indirectly related targets to unrelated targets, the separation of waveforms appeared maximal at left-lateralized sites, particularly posteriorly (see Figure 6A,B). There were no significant main effects of Priming Condition (Table 3B, left) or significant Priming Condition × Electrode Site interactions (Table 3B, right), but there were significant Priming Condition × Hemisphere interactions at all electrode columns (medial: F[1,17] = 5.228, p<.05; lateral: F[1,17] = 6.788, p<.05; peripheral: F[1,17] = 5.902, p<.05). In addition, there were significant Priming Condition × Electrode Site × Hemisphere interactions at the medial, F(2,34) = 6.220, p<.01, and lateral columns, F(3,51) = 3.637, p<.05. To determine the source of these interactions, follow-up ANOVAs were carried out at paired electrode sites for the medial, lateral, and peripheral columns, and these revealed that indirectly related targets elicited the smallest N400s over posterior and left-lateralized sites (see Table 4). Finally, paired t tests were conducted at the sites where the indirect N400 priming effect appeared maximal and yielded further confirmation that indirect targets elicited significantly smaller N400s than unrelated targets (CP5: t = 2.463, p<.05; P3: t = 2.336, p<.05; T5: t = 2.386, p<.05).
Table 4.
Paired Electrode Site ANOVAs:Indirectly Related vs. Unrelated
| Priming × Hemisphere |
||
|---|---|---|
| Degrees of freedom | F value | |
| Medial column | ||
| FC1-FC2 | 1,17 | 0.013 |
| C3-C4 | 1,17 | 9.906** |
| CP1-CP2 | 1,17 | 8.067* |
| Lateral column | ||
| F3-F4 | 1,17 | 0.418 |
| FC5-FC6 | 1,17 | 2.901 |
| CP5-CP6 | 1,17 | 12.363** |
| P3-P4 | 1,17 | 10.587** |
| Peripheral column | ||
| FP1-FP2 | 1,17 | 1.063 |
| F7-F8 | 1,17 | 5.671* |
| T3-T4 | 1,17 | 3.341† |
| T5-T6 | 1,17 | 7.774* |
| O1-O2 | 1,17 | 1.707 |
p<.1
p<.05
p<.01.
Directly related targets evoked less negative N400s than indirectly related targets, reflected by significant main effects of Priming Condition at all columns (Table 3C, left). These effects were posteriorly distributed and right lateralized as reflected by significant Priming Condition × Hemisphere interactions at all columns (medial: F[1,17] = 6.346, p<.05; lateral: F[1,17] = 9.689, p<.01; peripheral: F[1,17] = 10.630, p<.01), as well as by significant Priming Condition × Electrode Site × Hemisphere interactions at all columns (Table 3C, right).2
Thus, once again a significant N400 effect was obtained for both directly and indirectly primed targets as compared to unrelated targets, and directly related targets elicited a significantly less negative N400 than indirectly related targets.
600-800ms
No significant main effect of Priming Condition was seen and no significant Priming Condition × Electrode Site interactions were observed, all Fs<2.8, all Ps>.07.
Global Semantic Similarity Values and Their Contribution to the Indirect N400 Effect
We once again carried out LSA “pairwise comparisons” using tasaALL space, generating SSVs for each word pair. The elimination of nine word pairs did not significantly influence mean SSVs, and again directly related items had significantly greater SSVs than indirectly related, t(200) = 11.784, p<.001, or unrelated word pairs, t(200) = 19.727, p<.001, and indirectly related word pairs had significantly larger SSVs than unrelated word pairs, t(200) = 12.113, p<.001.
We then repeated analyses after matching the mean SSVs of the indirectly related and unrelated word pairs (indirect: 0.08 [SD: 0.05]; unrelated: 0.08 [SD: 0.07]). These analyses only included 75 of the original 201 indirectly related pairs, thus reducing our power to detect effects. Nonetheless, simple effects ANOVAs comparing mean N400 amplitudes of these indirectly related word pairs and the unrelated word pairs revealed a qualitatively similar pattern of results (see Figure 7). Significant Priming Condition × Hemisphere interactions were observed at the medial, F(1,17) = 8.594, p<.01, and lateral, F(1,17) = 5.722, p<.05, columns and significant Priming Condition × Electrode Site × Hemisphere interactions were seen at the medial, F(2,34) = 7.588, p<.01, and lateral, F(3,51) = 3.133, p<.05, columns. It is possible that this subset of word pairs differed critically from the entire stimulus set on characteristics that might influence N400 amplitude such as frequency or concreteness, and that these differences led to the N400 effects observed in these analyses. If this were the case, one would predict an interaction between N400 amplitude and stimulus set (total vs. SSV-matched). To test this prediction, we carried out two-way ANOVAs that included both mean N400 amplitude and stimulus set as factors. We did not observe any interactions between Priming and Stimulus Set, all Fs<1, nor was there a main effect of Stimulus Set in either the indirectly related, all Fs<1, or unrelated, all Fs<1, conditions. It is therefore very unlikely that the results of the SSV-matched analyses reported here reflect any factor other than priming condition.
Figure 7.
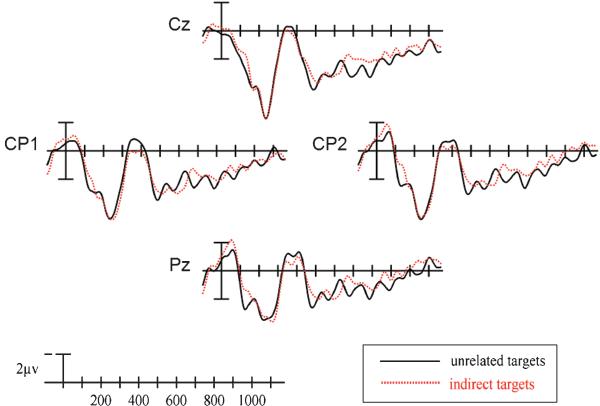
Indirectly related and unrelated targets matched by LSA in Experiment 2.
Discussion
Significant priming, as indexed by mean amplitude of the N400 component, was observed in a semantic categorization task for both directly related and indirectly related targets. The overall magnitude of this effect was smaller than that of Experiment 1, but a similar pattern of results was seen. Indirectly related targets elicited N400s in between those of directly related and unrelated targets. These findings are consistent with the hypothesis that indirect priming is, at least in part, a product of automatic processes and occurs regardless of the level of task explicitness.
After equalizing the SSVs between indirectly related and unrelated pairs, we still observed a small significant N400 priming effect for indirectly related targets. It therefore seems unlikely that any global semantic similarity contributing to automatic indirect priming through compound cue formation or through semantic feature overlap can entirely explain the modulation of the N400 to indirectly related targets. Rather, these findings suggest that automatic spreading activation contributed to indirect semantic priming.
However, because the associative strength between prime and target predicts the amount of facilitation under spreading activation theories, an alternative explanation might be that a subset of our indirectly related items had relatively high associative strength, thus generating intermediate N400 amplitudes in between directly related and unrelated items. Although it is doubtful that such items would have been preserved in our SSV-matched analyses, it is possible that they influenced the analyses that included the entire stimulus set. By examining the frequency with which our indirectly related primes were generated in response to their targets and vice versa, we would be able to more convincingly rule out this possibility.
EXPERIMENT 3
To investigate the associative strength of our materials, we conducted a free-association task where we asked participants to generate five associates to either primes from the directly related word pairs, primes from the indirectly related word pairs, or target words. In this way, we were able to assess both the forward and backward association strengths of the directly related and indirectly related word pairs, as well as the association strength between the primes of the indirectly related word pairs and their theoretical mediators. Assuming that our indirectly related word pairs had greater associative distance than our directly related word pairs and were linked together only by a mediating concept, we made three predictions: (1) that our directly related items would have significantly higher associative strength in both the forward and backward direction than our indirectly related items, (2) that few of the indirectly related words would be produced as associates in either direction, and (3) that the associative strength from indirect prime to our theoretical mediating word would be much higher than from indirect prime to target.
Methods
Participants
Thirty Tufts undergraduate students who did not participate in Experiments 1 or 2 were either paid for participation or received course credit.
Procedure
Participants were asked to read words listed on a Microsoft Excel sheet and type the first five words that came to mind for each of the words they read. They were instructed to try to avoid chaining their responses together and to try to only type in words that came to mind based on the initial word, not their own responses. Three lists were counterbalanced such that participants either viewed a list of primes from the directly related word pairs, primes from the indirectly related word pairs, or target words.
Results
As expected, the mean association strength for the directly related items was significantly higher than that of the indirectly related items in both the forward, t(9) = 8.943, p<.001, and backward, t(9) = 12.312, p<.001, direction, and the indirectly related items were almost never generated as associates in either the forward or backward direction (mean association strength direct forward: 6%; mean direct backward: 7%; mean indirect forward: 0.05%; mean indirect backward: 0.04%: see Table 5 for a more complete breakdown of association frequencies by first through fifth positions). Finally, the associative strength of indirect primes to their theoretical mediating words was much greater than the associative strength from indirect prime to target in either the forward or backward direction (forward: t[9] = 15.174, p<.001; backward: t[9] = 15.019, p<.001; mean associative strength mediator: 6%).
Table 5.
Association Frequencies by Condition
| Association frequencies |
||
|---|---|---|
| Forward | Backward | |
| Direct | ||
| First | 236/2100 | 343/2100 |
| Second | 115/2100 | 147/2100 |
| Third | 87/2100 | 88/2100 |
| Fourth | 67/2100 | 86/2100 |
| Fifth | 70/2100 | 55/2100 |
| Indirect | ||
| First | 9/2100 | 3/2100 |
| Second | 8/2100 | 13/2100 |
| Third | 13/2100 | 10/2100 |
| Fourth | 11/2100 | 3/2100 |
| Fifth | 12/2100 | 12/2100 |
| Mediator | ||
| First | 316/2100 | — |
| Second | 130/2100 | — |
| Third | 86/2100 | — |
| Fourth | 72/2100 | — |
| Fifth | 66/2100 | — |
Nonetheless, because a few participants did produce the indirectly related associates to a small proportion of items in either the forward or backward direction, it is possible that this small subgroup of items was responsible for our observed N400 effects. To rule out this possibility, we repeated our main ERP analyses for both experiments after removing these indirectly related word pairs and their matched unrelated control pairs. For both experiments, significant indirect N400 priming effects were still present. In Experiment 1, main effects at all electrode columns remained (midline: F[1,17] = 13.001, p<.01; medial: F[1,17] 13.517, p<.01; lateral: F[1,17] = 12.369, p<.01; peripheral: F[1,17] = 10.401, p<.01). For Experiment 2, Condition × Electrode Site × Hemisphere interactions were preserved at the medial, F(2,34) = 9.284, p<.01, and lateral columns, F(3,51) = 6.497, p<.01, and a significant Condition × Hemisphere interaction was preserved at the peripheral column, F(1,17) = 6.202, p<.05, whereas a trend remained at the lateral column, F(1,17) = 3.549, p = .08.
Discussion
Taken together, these results strongly suggest that our indirectly related word pairs had genuine mediated relationships, as opposed to being weakly but directly related. In particular, the extremely low frequencies with which the indirectly related items were generated in either the forward or backward direction suggests that these word pairs did not share direct relationships. Additionally, participants were much more likely to generate the mediating word in response to the indirect prime, indicating that this mediating word was truly the “first step” in our two-step pairs. Even so, we observed that some indirect items were generated as associates by some participants, and therefore we repeated our initial ERP analyses after removing these items and found that the indirect N400 priming effects were largely unchanged.
General Discussion
We observed a reliable N400 effect to indirectly primed words using both an explicit semantic judgment task (Experiment 1) and using an implicit semantic categorization task (Experiment 2). The explicit relatedness judgment task in Experiment 1 was associated with N400s that were larger in amplitude, spanned a broader time window, and were more widely distributed than the implicit task in Experiment 2. Chwilla et al. (2000) reported a similar finding: An explicit semantic matching instruction had the effect of enlarging N400 amplitudes and expanding the N400 time window.
Of most theoretical significance are two observations that we consider in more detail below. First, an indirect N400 priming effect was observed even when we matched the indirectly related and unrelated word pairs for participants’ explicit ratings (Experiment 1) and for global semantic similarity (Experiments 1 and 2). Second, in both experiments, the indirectly related targets evoked N400s with amplitudes that were in between those of directly related and unrelated targets.
In Experiment 1, an indirect N400 effect was observed even when participants perceived the indirectly related word pairs as being unrelated. In both experiments, an indirect N400 effect was observed even when indirectly related word pairs had few features in common and did not co-occur (as measured by SSVs generated by Latent Semantic Analysis). These observations are, to some degree, internally consistent. If indirectly related word pairs always tended to share more semantic features or contextual similarities than unrelated pairs (as indexed by the global semantic similarity values), it would be hard to imagine why participants could not then access such semantic similarity information and use this information when asked to explicitly rate the semantic relationships between the word pairs.
Although our indirect N400 priming effects had remained significant after matching our stimuli for both participant ratings and SSVs, we wanted to satisfy any concerns that these effects may have been driven by a small number of indirectly related word pairs with higher associative strength. In Experiment 3, we collected free-association data that demonstrated that our indirectly related items were very infrequently generated as associates, in sharp contrast with both our directly related items and the theoretical mediating items for indirectly related word pairs. These findings suggest that our indirectly related word pairs did not simply have relatively weak direct relationships. Nevertheless, because some indirect items were generated as associates by a few participants, to more stringently test the strength of our effects we repeated our initial ERP analyses after removing these items. The indirect N400 priming effects remained significant.
Taken together, these findings suggest that the indirect priming effect cannot be accounted for entirely by strategic semantic matching mechanisms, by compound cuing, or by semantic feature overlap between the indirectly related word pairs. If the indirect N400 priming effect arose primarily from semantic matching, one would expect participants to rate indirectly related word pairs as significantly more related than unrelated word pairs, and this effect should disappear after matching participants’ explicit relatedness ratings to indirectly related and unrelated word pairs. However, our participants did not perceive indirectly related items as significantly more related than unrelated items, and we demonstrate a significant indirect N400 priming effect even after matching for relatedness ratings between conditions. Alternatively, if this effect was primarily automatic, but the mechanism underlying this effect was compound cue or semantic feature overlap, one would predict that the indirect N400 priming effect could be explained by the indirectly related word pairs having higher semantic similarity values than unrelated word pairs. However, after matching indirectly related and unrelated word pairs on a measure of global semantic similarity, we still observed a small significant N400 effect for indirectly primed words. This leaves open just one explanation for the indirect priming effect to these matched word pairs: a spread of activation from prime to mediator to target.
The second important observation across the two experiments is the gradation of modulation of the N400: Indirectly primed words elicited N400s in between those of directly related and unrelated targets. This was a widespread effect in Experiment 1 when participants performed the explicit task, and was more marked at posterior and left-lateralized sites in Experiment 2 when participants performed the implicit task. This pattern of modulation supports a model of spreading activation that predicts a decrease in facilitation with increasing semantic distance. In contrast, the operation of strategic processes such as semantic matching does not predict this pattern, but rather that priming should occur in an allor-none fashion, depending on whether participants recognize the indirectly related word pairs as being related or unrelated. Similarly, unless the indirectly related and directly related word pairs shared weak but direct semantic relationships or half as many semantic features, other models of automatic priming also fail to predict this stepwise reduction in priming. We sought to rule out any differences that could be explained by these other mechanisms in two ways: First, we matched indirectly related and unrelated word pairs on their semantic similarity values; second, we gathered free association norms and excluded any indirectly related items that were produced as associates in either the forward or backward direction. After repeating our ERP analyses for both of these reduced data sets, we still observed a stepwise reduction in N400 amplitude from directly related to indirectly related to unrelated target words.
Conclusion
In conclusion, our findings provide evidence that at least some of the indirect N400 priming effect can be accounted for by the spread of activation within semantic memory from prime to mediator to target. Although we do not submit that spreading activation is the only potential means of achieving indirect semantic priming, we suggest that, of the current theoretical frameworks in the literature, spreading activation is the one that best explains these findings. Future studies using masked primes that preclude the use of any type of strategic processing will further examine the automaticity of this mechanism.
Footnotes
In addition to contrasting the N400 evoked by the directly and indirectly related targets, we also conducted comparisons between difference scores indexing the direct and indirect N400 effects: The direct N400 priming effect (calculated as the difference in mean N400 amplitudes) was significantly larger than the indirect N400 priming effect (midline: F[1,17] = 81.379, p<.001; medial: F[1,17] = 71.108, p<.001; lateral: F[1,17] = 56.804, p<.001; peripheral: F[1,17] = 38.262, p<.001).
The direct N400 priming effect was significantly larger than the indirect N400 priming effect (midline: F[1,17] = 6.451, p<.05; medial: F[1,17] = 6.081, p<.05; lateral: F[1,17] = 6.309, p<.05; peripheral: F[1,17] = 10.123, p<0.01).
This research was supported by NIH (R01 MH071635, K23 MH02034, R01 HD25889, and R01 HD043251) and the Institute for Mental Illness and Neuroscience Discovery (MIND). Gina R. Kuperberg was also supported by the NARSAD (with the Sidney Baer Trust) and a Claflin Distinguished Scholars Award from Massachusetts General Hospital.
REFERENCES
- Anderson JR. The architecture of cognition. Harvard University Press; Cambridge, MA: 1983. [Google Scholar]
- Balota DA, Lorch RF. Depth of automatic spreading activation: Mediated priming effects in pronunciation but not in lexical decision. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1986;12:336–345. [Google Scholar]
- Bentin S, McCarthy G, Wood CC. Event-related potentials, lexical decision and semantic priming. Clinical Neurophysiology. 1985;60:343–355. doi: 10.1016/0013-4694(85)90008-2. [DOI] [PubMed] [Google Scholar]
- Chwilla DJ, Kolk HHJ. Three-step priming in lexical decision. Memory and Cognition. 2002;30:217–225. doi: 10.3758/bf03195282. [DOI] [PubMed] [Google Scholar]
- Chwilla DJ, Kolk HHJ, Mulder G. Mediated priming in the lexical decision task: Evidence from event-related potentials and reaction time. Journal of Memory and Language. 2000;42:314–341. [Google Scholar]
- Collins AM, Loftus EF. A spreading-activation theory of semantic processing. Psychological Review. 1975;82:407–428. [Google Scholar]
- Donchin E, Coles MG. Is the P300 component a manifestation of context updating? Behavioral and Brain Sciences. 1988;11:357–427. [Google Scholar]
- Greenhouse SW, Geisser S. On methods in the analysis of profile data. Psychometrika. 1959;24:95–112. [Google Scholar]
- Hill H, Strube M, Roesch-Ely D, Weisbrod M. Automatic vs. controlled processing in semantic priming—Differentiation by event-related potentials. International Journal of Psychophysiology. 2002;44:197–218. doi: 10.1016/s0167-8760(01)00202-1. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Semantic priming and stimulus degradation: Implications for the role of the N400 in language processing. Psychophysiology. 1993;30:47–61. doi: 10.1111/j.1469-8986.1993.tb03204.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Automatic and attentional processing: An event-related brain potential analysis of semantic priming. Brain and Language. 1998;35:66–85. doi: 10.1016/0093-934x(88)90101-0. [DOI] [PubMed] [Google Scholar]
- Hutchinson KA. Is semantic priming due to association strength or feature overlap? A microanalytic review. Psychonomic Bulletin & Review. 2003;10:785–813. doi: 10.3758/bf03196544. [DOI] [PubMed] [Google Scholar]
- Kiefer M. The N400 is modulated by unconsciously perceived masked words: Further evidence for an automatic spreading activation account of N400 priming effects. Cognitive Brain Research. 2002;13:27–39. doi: 10.1016/s0926-6410(01)00085-4. [DOI] [PubMed] [Google Scholar]
- Kiefer M, Ahlegian M, Spitzer M. Working memory capacity, indirect semantic priming, and Stroop interference: Patterns of interindividual prefrontal performance differences in healthy volunteers. Neuropsychology. 2005;19:332–344. doi: 10.1037/0894-4105.19.3.332. [DOI] [PubMed] [Google Scholar]
- Kiefer M, Brendel D. Attentional modulation of unconscious ‘automatic’ processes: Evidence from event-related potentials in a masked priming paradigm. Journal of Cognitive Neuroscience. 2006;18:184–198. doi: 10.1162/089892906775783688. [DOI] [PubMed] [Google Scholar]
- Kiefer M, Weisbrod M, Kern I, Maier S, Spitzer M. Right hemisphere activation during indirect semantic priming: Evidence from event-related potentials. Brain and Language. 1998;64:377–408. doi: 10.1006/brln.1998.1979. [DOI] [PubMed] [Google Scholar]
- Kučera H, Francis WN. Computational analysis of presentday American English. Brown University Press; Providence, RI: 1967. [Google Scholar]
- Kutas M, Hillyard SA. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science. 1980;207:203–205. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Landauer TK, Dumais ST. A solution to Plato’s problem: The latent semantic analysis theory of acquisition, induction, and representation of knowledge. Psychological Review. 1997;104:211–240. [Google Scholar]
- Landauer TK, Foltz PW, Dumais ST. Introduction to latent semantic analysis. Discourse Processes. 1998;25:259–284. available at: http://lsa.colorado.edu.
- Masson MEJ. A distributed memory model of context effects in word identification. In: Besner D, Humphreys GW, editors. Basic processes in reading: Visual word recognition. Lawrence Earlbaum Associates; Hillsdale, NJ: 1991. pp. 233–263. [Google Scholar]
- Masson MEJ. A distributed memory model of semantic priming. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1995;21:3–23. [Google Scholar]
- McKoon G, Ratcliff R. Spreading activation versus compound cue accounts of priming: Mediated priming revisited. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1992;18:1155–1172. doi: 10.1037//0278-7393.18.6.1155. [DOI] [PubMed] [Google Scholar]
- McNamara TP. Theories of priming: I. Associative distance and lag. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1992;18:1173–1190. [Google Scholar]
- McNamara TP, Altarriba J. Depth of spreading activation revisited: Semantic mediated priming occurs in lexical decisions. Journal of Memory and Language. 1988;27:545–559. [Google Scholar]
- Meyer DE, Schvaneveldt RW. Facilitation in recognizing pairs of words: Evidence of a dependence between retrieval operations. Journal of Experimental Psychology. 1971;90:227–234. doi: 10.1037/h0031564. [DOI] [PubMed] [Google Scholar]
- Misra M, Holcomb PJ. Event-related potential indices of masked repetition priming. Psychophysiology. 2003;40:115–130. doi: 10.1111/1469-8986.00012. [DOI] [PubMed] [Google Scholar]
- Neely JH. Semantic priming and retrieval from lexical memory: Roles of inhibitionless spreading activation and limited capacity attention. Journal of Experimental Psychology. 1977;106:226–254. [Google Scholar]
- Neely JH. Semantic priming effects in visual word recognition: A selective review of current findings and theories. In: Besner D, Humphreys GW, editors. Basic processes in reading: Visual word recognition. Lawrence Earlbaum Associates; Hillsdale, NJ: 1991. pp. 264–336. [Google Scholar]
- Neely JH, Keefe DE. Semantic context effects on visual word processing: A hybrid prospective/retrospective processing theory. In: Bower GH, editor. The psychology of learning and motivation: Advances in research and theory. Vol. 24. Academic Press; New York: 1989. pp. 207–248. [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Ratcliff R, McKoon G. A retrieval theory of priming in memory. Psychological Review. 1988;95:385–408. doi: 10.1037/0033-295x.95.3.385. [DOI] [PubMed] [Google Scholar]
- Rugg MD. The effects of semantic priming and word repetition on event-related potentials. Psychophysiology. 1985;22:642–647. doi: 10.1111/j.1469-8986.1985.tb01661.x. [DOI] [PubMed] [Google Scholar]
- Silva-Pereyra J, Harmony T, Villanueva G, Fernandez T, Rodriguez M, Galan L, et al. N400 and lexical decisions: Automatic or controlled processing? Clinical Neuropsychology. 1999;110:813–824. doi: 10.1016/s1388-2457(99)00009-7. [DOI] [PubMed] [Google Scholar]
- Weisbrod M, Kiefer M, Winkler S, Maier S, Hill H, Roesch-Ely A, et al. Electrophysiological correlates of direct versus indirect semantic priming in normal volunteers. Cognitive Brain Research. 1999;8:289–298. doi: 10.1016/s0926-6410(99)00032-4. [DOI] [PubMed] [Google Scholar]