

Abstract
The current study evaluated the effects of drugs that increase GABA levels by activation of GABAB receptors (baclofen and CGP44532) or by inhibition of GABA reuptake (tiagabine) on the reinstatement of extinguished lever responding produced by priming doses of cocaine in baboons (i.e., cocaine-seeking). Cocaine self-injection was established and maintained under a fixed ratio (FR10) schedule of reinforcement during daily 2-h sessions. Lever responding was extinguished by substituting vehicle (saline) for cocaine until the number of self-injections decreased to 10 or less per session for 2 consecutive sessions (defined as extinction). Once extinction occurred, priming doses of cocaine (0.1-3.2 mg/kg, IV) were administered during extinction conditions. Administration of priming doses of cocaine significantly increased cocaine-seeking in a dose dependent manner. Cocaine-seeking produced by priming doses of cocaine were attenuated by pretreatment with baclofen (N=5) or CGP44532 (N=5) but not tiagabine (N=3). The doses of baclofen (0.32 mg/kg), and CGP445532 (0.32 mg/kg) that reduced cocaine-seeking produced by cocaine priming doses did not reinstate cocaine-seeking and did not produce overt effects when administered alone. These data indicate that GABAB agonists may reduce relapse to cocaine taking.
Keywords: reinstatement, reinforcement, GABAB receptor agonist, relapse, self-administration, primates
1. Introduction
Important new targets for the development of potential medications for cocaine addiction are the multiple neurotransmitter systems that influence the dopamine (DA) brain reward systems. The inhibitory neurotransmitter gamma-aminobutyric acid (GABA) has extensive neuronal connections in mesolimbic areas of the brain, which appear to inhibit dopaminergic neurons (Bardo, 1998). Thus, drugs that enhance GABAergic neurotransmission and thereby inhibit dopamine are likely candidates as potential treatment medications for cocaine dependence (Sofuoglu and Kosten, 2006). GABAergic neurotransmission can be increased by positive modulators at GABAA or GABAB receptors, as well as drugs that increase endogenous GABA by increasing GABA synthesis, inhibiting GABA-transaminase, or blocking reuptake of GABA.
The effects of GABAergic drugs on cocaine craving and relapse are still under investigation in both human and animal studies. GABAB agonists, in particular, show promise as potential treatment medications for cocaine abuse (Brebner et al., 2002; Sofuoglu and Kosten, 2005). In rodents, pretreatment with GABAB agonists dose-dependently reduced the cocaine-induced DA release in mesolimbic regions (Fadda et al., 2003), reduced cocaine-induced facilitation of brain stimulation reward (Dobrovitsky et al., 2002), and decreased cocaine-induced seizures (Gasior et al., 2004). Baclofen, the prototypical GABAB agonist, has been reported to decrease cocaine self-administration in rats (Roberts et al., 1996; Roberts & Andrews, 1997). In an initial open labeled clinical trial, baclofen (20 mg t.i.d.), combined with group counseling three times per week, reduced self-reports of craving and cocaine use in 10 cocaine abusers (Ling et al., 1998). The decreased self-reports of cocaine use by baclofen-treated patients was confirmed by a decrease in cocaine positive urines. In a subsequent 16-week double-blind study in 35 cocaine-dependent subjects, baclofen (20 mg t.i.d.) reduced cocaine use and increased the number of cocaine-free urines (Shoptaw et al., 2003) but did not significantly alter cocaine craving. Recent brain imaging studies have demonstrated that cocaine-related cues increase limbic activation (as measured by PET) and cocaine craving in humans (Childress et al., 1999). Pretreatment with baclofen (10-20 mg b.i.d. for 10 days) blocked both cocaine-cue induced craving and limbic activation in cocaine abusers (n=3) (Childress et al., 2000; Franklin et al., 2000). Recently characterized phosphinic acid analogues of GABA, such as CGP44532 (Froestl et al., 1993, Froestl et al., 1995), have also been effective at reducing cocaine self-administration. Brebner et al. (1999) reported decreased cocaine self-administration in rats under a discrete-trials procedure and decreased break points in a cocaine-maintained progressive ratio procedure. Compared to baclofen, administration of CGP44532 to rhesus monkeys produced more robust muscle relaxant effects without sedation or decreased vigilance (Froestl et al., 1995).
In addition to the GABAB agonist baclofen, clinical trials have been initiated for other compounds that increase GABAergic tone (Sofuoglu and Kosten, 2006). For example, tiagabine which is used for the treatment of epilepsy has been shown to reduce subjective ratings of “stimulated” and “crave cocaine” in participants administered cocaine (Sofuoglu et al., 2005) and appeared to reduce cocaine use in early treatment trials (Gonzalez et al., 2003; Winhusen et al., 2005). Tiagabine increases endogenous GABA by blocking reuptake of synaptically released GABA in both the presynaptic neurons and glial cells (Suzdak and Jansen, 1995). Tiagabine does not alter GABA release and lacks affinity for other neurotransmitter binding sites and/or reuptake sites (e.g., DA, serotonin, norepinephrine) (Suzdak and Jansen, 1995). Based on its mechanism of action and these preliminary findings, further investigation of the effects of tiagabine on cocaine-mediated behaviors is warranted (Sofuoglu and Kosten, 2006).
In animals, models of craving and relapse include responding under second-order schedules and the reinstatement of extinguished drug-taking behavior (Carroll and Comer, 1996; deWit and Stewart, 1981; Markou et al., 1993). In second-order schedules, subjects respond according to one schedule for a brief presentation of a stimulus such as a light (i.e., the unit schedule) and then responses emitted under the unit schedule are reinforced by drug injection according to another schedule of reinforcement (Schindler et al., 2002). It is well established that cocaine maintains responding under second-order schedules (Arroyo et al., 1998; Goldberg et al., 1976; Johanson, 1982; Katz, 1979). The mechanisms involved in reinstatement of drug-seeking behaviors (relapse) may differ from those involved in the maintenance of drug-taking behaviors (Norman et al., 1999; Self and Nestler, 1998; Spealman et al., 1999). Reinstatement procedures usually involve a combination of self-administration and extinction procedures. For example, subjects are first trained to press a lever to self-inject a drug. Once stable drug-maintained lever responding is established, a period of extinction is imposed in which lever responses no longer result in drug delivery. When lever responding decreases, animals are then exposed to different stimuli and lever responding in the absence of drug reinforcement (defined as “drug-seeking” behavior) can be measured. Noncontingent cocaine ‘priming’ injections reliably reinstate cocaine-seeking in both rats and monkeys (deWit and Stewart, 1981; Gerber and Stretch, 1975; Slikker et al., 1984). Presentation of stimuli that have been previously associated with cocaine injections will also reinstate lever responding after a period of extinction (Davis and Smith, 1976; deWit and Stewart, 1981).
Although studies have evaluated the effects of GABAB agonists on ongoing cocaine-taking (self-administration) behaviors (for review see Brebner et al., 2002; Cousins et al., 2002), relatively few studies have evaluated the effects of GABAB agonists on cocaine seeking behaviors. Baclofen reduced both cocaine self-administration and lever responding maintained by cocaine-associated cues under a second order schedule in rats (Di Ciano and Everitt, 2003). Campbell et al. (1999) reported that baclofen dose-dependently reduced the reinstatement of extinguished cocaine-taking behavior produced by priming injections of cocaine in rats.
As described above, noncontingent cocaine ‘priming’ injections will reliably reinstate extinguished lever responding previously maintained by cocaine (defined as cocaine-seeking). The current study investigated the effects of GABAB agonists baclofen and CGP44532 and the GABA reuptake inhibitor tiagabine on cocaine-seeking behavior in baboons. Baclofen, CGP44532, tiagabine, or vehicle pretreatments were administered before priming injections of cocaine or its vehicle and lever responding in the absence of drug reinforcement was determined. Previously, this laboratory evaluated the effects of a range of doses of baclofen, CGP44532 and tiagabine on food-maintained operant responding in baboons (Weerts et al., 2005). Since decreases in food-maintained responding may indicate a non-specific general suppression of behavior, only the highest dose of baclofen, CGP44532 and tiagabine that did not decrease food-maintained responding in baboons (0.32 mg/kg in each case) was used in the current study.
2. Methods
2.1. Subjects
Subjects were ten adult male baboons (Papio anubis, Primate Imports, NY, NY) weighing 23 to 36 kg at the beginning of the experiment. Each baboon had an intravenous catheter that was surgically implanted using procedures described previously (Lukas et al., 1982). The chronic indwelling silastic catheter exited in the midscapular region of the back and was protected by a tether/harness/vest system that permitted the baboon free movement inside the cage (Lukas et al., 1982). One baboon (HK) was experimentally naïve. All other baboons had histories with self-administration of cocaine and food under the procedures used in the current study (Weerts et al., 2005; Weerts and Griffiths, 2003). In the current study, baboons DM, JN, JP, SI, and ZA completed the baclofen conditions, baboons AB, BA, HU, KR and ZA completed CGP44532 conditions, and baboons HK, KR, and ZA completed the tiagabine conditions. Baboons that received more than one pretreatment drug (KR, ZA) completed all of the conditions (as described below) for each pretreatment drug.
Baboons were not food restricted during the studies. All subjects received monkey biscuits, two pieces of fresh produce and a multivitamin at approximately 11:00 h each day. Water was available from a drinking spout 24 h/day and volumes consumed each day were recorded. Baboons were anesthetized with ketamine hydrochloride (preceded by atropine sulfate) every 2 weeks for physical examinations and to perform medical checks/care of catheter exit sites. The facilities were maintained in accordance with USDA and AALAC standards. The Johns Hopkins Medical Institution Animal Care and Use Committee approved the experimental protocol.
2.2. Apparatus
The baboons were housed in standard stainless steel primate cages, which also served as experimental chambers. Each cage was equipped with a bench running the length of one wall and an “intelligence panel” mounted on the rear wall. The intelligence panel contained a Lindsleystyle pull and release operandum (Med-Associates, Georgia, VT) for drug self-injection which was mounted in the lower left quadrant of the panel within easy reach of the baboon when seated on the bench. A colored (e.g., yellow or green) “jewel light” (1 cm diameter) was mounted above the lever. Although not used in the current study, a recessed food hopper, which contained a white translucent panel (5 × 5 cm), was located in the center of the panel. A second 5 × 5 cm “baylight,” that could be lit by a green bulb, was located in the upper left quadrant of the intelligence panel. A speaker for delivery of white noise and tones was mounted on the back of the intelligence panel. During the sessions, masonite panels were placed on the sides of the cage to provide a visual barrier from the baboons on either side of the subject, but subjects could see other baboons housed across from them. The room ceiling lights were brightly illuminated for 13 h/day (6:00-19:00h) and were dimly illuminated for the remaining 11 h/day. Experimental control and data collection were accomplished using MED-PC/Med State (Med Associates, East Fairfield, VT) software and hardware interfaced with personal computers.
The catheter, which was protected by the tether/harness/vest system, was attached to an 18-gauge single-channel liquid swivel (Instec-Soloman, Plymouth Meeting, PA) mounted on the top of the cage. The swivel was connected to a valve system that allowed delivery of the sterile drug and vehicle solutions from two separate peristaltic pumps (Model 66 Harvard Apparatus, Natick, MA). Injections of drug or vehicle (2 ml/injection) were delivered at a rate of 0.25 ml per s. Drug solutions were injected into the valve system by means of the first pump and then flushed into the vein with 2 ml of vehicle via a second peristaltic pump. A third pump (Watson-Marlow model 403U/R1, Atlantic Technology Group, Richmond, VA) continuously administered heparinized (10 units/ml) saline at a rate of 10 ml/h to maintain catheter patency. The infusion system was a closed system that was maintained using aseptic technique. The pumps, infusion systems, IV solutions, and drinking water bottles were located on a grating above the cage.
2.3. Drugs
Cocaine hydrochloride (National Institute on Drug Abuse Drug Supply Program, Rockville, MD) was dissolved in sterile 0.09% saline and sterilized by filtration (25 mm Swinnex, Minipore Corp., Bedford, MA). Cocaine priming doses were administered in 2-ml IV infusions 5-min before the self-injection session. Baclofen (Sigma-Aldrich, St. Louis, MO), CGP44532 (3-Amino-2-(S)-hydroxy-propyl)-methyl-phosphinic acid) (Novartis AG, Basel Switzerland), and tiagabine (Cephalon, Inc., West Chester, PA) were dissolved in 2-mls of sterile water and injected IM 30-min before the self-injection session began.
2.4. Procedures
2.4.1. Self-injection
The availability of each injection was signaled by a 5-s tone, and continuous illumination of the jewel light over the lever. Each injection was dependent on completion of 10 responses (i.e., a fixed ratio reinforcement schedule). When the jewel light over the lever was illuminated, a pull and release of the Lindsley lever produced a 0.1 s feedback tone. Upon completion of the response requirement, the jewel light over the lever was extinguished, a 4-min timeout began, and the injection was delivered. To signal drug delivery and the onset of drug effects, the green baylight was illuminated for the first 3 min of the timeout period. After the timeout, the availability of the next injection was again signaled by a tone and illumination of the jewel light. A maximum of 30 injections was available during each daily 2-h session. The cocaine dose used (0.032 mg/kg/injection) was one that maintained 25-30 injections under the 2 h procedure as determined in previous studies (Weerts et al., 2005; Weerts and Griffiths, 1999; Weerts and Griffiths, 2003)
2.4.2. Reinstatement of cocaine-seeking
The reinstatement procedure was similar to one used previously (Weerts and Griffiths, 2003). Reinforcement, extinction and reinstatement tests were conducted over separate daily sessions. First, baseline rates of ≥ 25 self-injections for cocaine were established (defined as criterion performance). After at least 3 consecutive sessions in which criterion performance for cocaine self-injection was maintained, lever responding was extinguished by substituting the vehicle (saline) for cocaine. The criterion for extinguished behavior was < 10 injections per session for 2 consecutive sessions and no increasing trends (i.e., the number of injections for the last session was the same as or lower than the number of injections for the preceding session). All reinstatement tests were conducted only if the extinguished behavior criterion was met.
During reinstatement sessions, a priming dose of a cocaine (0.1 – 1.8 mg/kg) or its vehicle (saline) was administered IV 5-min before a session in which no drug was available (i.e., only saline injections delivered). Doses higher than 1.8 mg/kg cocaine were not used since a previous study showed that 3.2 mg/kg cocaine produced low rates of lever responding and full body tremors (Weerts and Griffiths, 2003). For interaction tests, the pretreatment drug or its vehicle (sterile water) were administered IM 30-min before the session and 25 min before the IV priming dose of cocaine. The doses of baclofen (0.32 mg/kg), CGP44532 (0.32 mg/kg), and tiagabine (0.32 mg/kg) selected for interaction tests in the current study were doses that did not decrease lever responding maintained by food in the same 2-h procedure in a previous study (Weerts et al., 2005). The session following a test session was always the baseline condition (i.e., cocaine was substituted for saline). The cocaine self-injection baseline condition was then maintained until the number of reinforcers delivered was ≥ 25 for 3 consecutive days. Once criterion performance was obtained, saline was again substituted for cocaine until the extinction criterion was met again and the next cocaine priming dose or interaction test session was conducted. This procedure of first establishing criterion performance for the self-administration of cocaine and then for extinction was repeated before each priming dose or interaction test session. Thus, reinstatement tests occurred about once a week. For each drug group, every baboon completed both treatment conditions (i.e. cocaine priming doses alone and cocaine priming doses plus pretreatment).
2.5. Data summary
Data collection included the actual times of onset of each drug/vehicle injection trial (i.e., when an injection was available as indicated by illumination of the jewel light over the “drug” lever), the number of drug lever responses emitted when injections were available (i.e., during illumination of the jewel light for the “drug” lever), the number of injections delivered, the number of lever responses emitted during timeout, actual times for completion of the response requirement for each injection, and the number of injections delivered.
A single subject design was used in which each baboon served as his own control (Sidman, 1960). Baseline rates of cocaine self-administration and saline extinction were the last 3 days of each condition that preceded each reinstatement test. First, statistical analysis of variance (ANOVA) was conducted to compare the number of saline injections taken after priming doses of cocaine between groups (baclofen group vs. CGP44532 group vs. tiagabine group). Second, self-injection data for each drug group were then analyzed separately using a 2-way repeated measures ANOVA for treatment condition (i.e., cocaine priming doses alone vs. cocaine priming dose plus pretreatment with baclofen, CGP44532, or tiagabine). Two additional analysis were conducted. First, latency (in s) from the start of the session until administration of the first saline injection was analyzed separately for each pretreatment group using a 2-way repeated measures ANOVA. Second, an identical analysis was conducted for local response rate (“run rate”). Response rate was defined as the number of responses per s; run rate was calculated using only the time (in s) from the first to the last response in each ratio (that is, excluding any pausing that occurred prior to initiation of responding). A p<0.05 was accepted as significant. When a significant effect for the interaction for treatment condition and cocaine dose was found, then post-hoc Bonferroni t-tests comparisons were conducted.
3. Results
Cocaine (0.032 mg/kg) maintained self-administration in all baboons during the daily 2-h baseline sessions. All baboons obtained near maximal numbers of injections (25 to 30) when cocaine was available. The mean run rate (responses/s) during the cocaine baseline sessions was calculated separately for each pretreatment group. Cocaine maintained high run rates in each pretreatment group [responses/s (±1 SEM): baclofen, 2.45 (0.51); CGP44532, 3.37 (0.53); tiagabine, 3.4 (0.93)]. When saline was substituted for cocaine, the number of self-injections decreased over consecutive days. The number of saline injections on the day before reinstatement tests ranged from 1-8.
The three pretreatment groups (baclofen vs. CGP44532 vs. tiagabine) were not significantly different from each other for baseline levels of cocaine self-injection or for saline self-injection under extinction conditions. Across pretreatment groups, there was a significant main effect for cocaine priming dose (F (4,40)=52.43, P < 0.0001). Although only saline was available for injection, administration of priming doses of cocaine (0.1-1.8 mg/kg, i.v.) following extinction periods increased lever responding (i.e., cocaine-seeking) in a dose-dependent manner; the number of ratios completed (which resulted in saline injections) increased as a function of the priming dose administered (Fig. 1).
Figure 1.
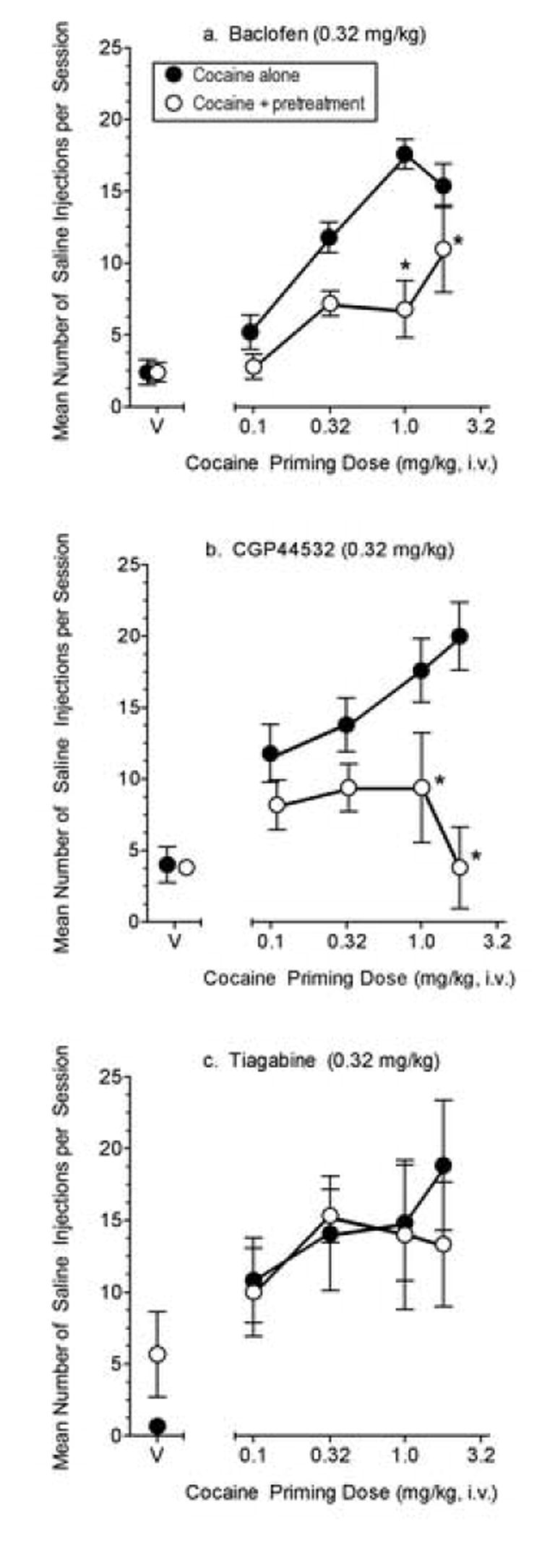
Effects a. baclofen (N=5), b. CGP44532 (N=5), and c. tiagabine (N=3) on the reinstatement of extinguished lever responding by cocaine priming dose injections. Data shown are the group mean number of ratios completed on the cocaine-associated lever after priming injections of cocaine alone (closed circles) and in combination with the pretreatment (open circles) when only saline was available for self-injection and lever responding had been extinguished. Pretreatments (0.32 mg/kg baclofen, 0.32 mg/kg CGP44532, or 0.32 mg/kg tiagabine) or vehicle (V) were administered i.m. 30 min before the saline self-injection session and then cocaine (0.1-1.8 mg/kg, i.v.) or its vehicle were administered 5 min before the saline self-injection session. Data points above “V” are the group mean number of ratios completed on the cocaine-associated lever following administration of the pretreatment alone or its vehicle alone during the saline extinction period. * represent significant (p<0.05) difference for pretreatment when compared to the same dose of cocaine alone
As shown in Fig. 1a, pretreatment with baclofen attenuated the reinstatement of cocaine-seeking produced by priming injections of cocaine. ANOVA of the baclofen group data confirmed a significant main effect for 0.32 mg/kg baclofen pretreatment (F(1,20)=21.32, P < 0.001) and cocaine priming dose (F(4,20)=22.63, P < 0.0001). There was also a significant interaction for baclofen pretreatment and cocaine dose (F(4,20)=3.48, P < 0.03). Bonferroni t-tests indicated a significant difference between means for treatment conditions (baclofen vs. vehicle) following 1.0 (P < 0.001) and 1.8 (P < 0.05) mg/kg priming injections of cocaine.
Like baclofen, pretreatment with 0.32 mg/kg CGP44532 attenuated the reinstatement of cocaine-seeking produced by priming injections of cocaine (Fig. 1b). ANOVA of the CGP44532 group data confirmed that there were significant main effects for CGP44532 pretreatment (F(1,20)=46.74, P < 0.0001) and cocaine priming dose (F(4,20)=3.72, P < 0.02). There was also a significant interaction for CGP44532 pretreatment and cocaine dose (F(4,20)=8.22, P < 0.0004). Bonferroni t-tests indicated a significant difference between means for treatment conditions (CGP44532 vs. vehicle) following 1.0 (p< 0.01) and 1.8 (p< 0.001) mg/kg priming injections of cocaine.
In contrast, pretreatment with 0.32 mg/kg tiagabine did not systematically affect the number of saline self-injections following cocaine priming doses (Fig. 1c). Results of ANOVA of the tiagabine group data confirmed a main effect of cocaine priming dose (F(4,16)=8.76, P<.001). Tiagabine pretreatment (vehicle vs. 0.32 mg/kg tiagabine) and the interaction for tiagabine pretreatment and cocaine dose treatment were not significant.
Interestingly, the latency to the first injection of saline following administration of the IV cocaine priming doses progressively increased in a dose dependent manner (F(3,30)=10.77, p<.0001). Table 1 shows the mean (SEM) latency (s) to the first saline injection following IV administration of cocaine priming doses alone and with 0.32 mg/kg baclofen, 0.32 mg/kg CGP44532, or 0.32 mg/kg tiagabine pretreatments. Pretreatment with baclofen, CGP44532, or tiagabine did not systematically affect the latency to the first injection. In addition, once the first response in the ratio occurred, the remaining response requirement was completed at a high rate, regardless of the cocaine priming dose or the pretreatment condition. Table 2 shows the mean run rate (and SEM) for injections of saline following administration of the IV cocaine priming doses alone (no pretreat) and following pretreatment prior to administration of the IV cocaine priming (pretreat) for each treatment group. Pretreatment with baclofen, CGP44532 or tiagabine prior to the cocaine priming injection did not significantly or systematically change the local response rate (run rate). Administration of 0.32 mg/kg baclofen, 0.32 mg/kg CGP44532 or 0.32 mg/kg tiagabine alone also did not significantly alter run rates.
Table 1.
Mean (SEM) latency (in sec) to the first saline injection following IV administration of cocaine priming doses alone and with 0.32 mg/kg baclofen, 0.32 mg/kg CGP44532, or 0.32 mg/kg tiagabine pretreatments
| Baclofen Group | CGP44532 Group | Tiagabine Group | ||||
|---|---|---|---|---|---|---|
| Cocaine
Priming Dose (mg/kg) |
No
Pretreat |
Pretreat | No
Pretreat |
Pretreat | No
Pretreat |
Pretreat |
| Vehicle | 90.4
(35.5) |
51.0
(32.4) |
134.8
(31.3) |
39.0
(13.6) |
101.0
(93.0) |
7.5
(0.5) |
| 0.1 mg/kg | 107.4
(69.3) |
230.5
(186.5) |
196.2
(128.9) |
193.0
(71.4) |
23.7
(17.2) |
117.0
(44.5) |
| 0.32 mg/kg | 415.0
(99.0) |
484.4
(230.4) |
341.8
(131.5) |
503.8
(176.0) |
350.3
(179.9) |
310.0
(165.7) |
| 1.0 mg/kg | 1157.6
(192.0) |
1022.5
(344.0) |
1340.7
(521.0) |
1661.3
(602.1) |
1383.8
(742.8) |
1634.0
(999.1) |
| 1.8 mg/kg | 2217.6
(523.6) |
2028.8
(590.4) |
1303.4
(218.8) |
2112.9
(1182.2) |
876.3
(539.7) |
155.3
(90.7) |
Table 2.
Mean (SEM) run rate (r/sec) to the first saline injection following IV administration of cocaine priming doses alone and with 0.32 mg/kg baclofen, 0.32 mg/kg CGP44532, or 0.32 mg/kg tiagabine pretreatments
| Baclofen Group | CGP44532 Group | Tiagabine Group | ||||
|---|---|---|---|---|---|---|
| Cocaine
Priming Dose (mg/kg) |
No
Pretreat |
Pretreat | No
Pretreat |
Pretreat | No
Pretreat |
Pretreat |
| Vehicle | 1.89
(0.46) |
1.03
(0.53) |
3.80
(1.15) |
2.13
(0.84) |
4.12
(2.09) |
3.02
(1.65) |
| 0.1 mg/kg | 2.50
(0.89) |
0.74
(0.47) |
3.17
(1.15) |
2.33
(0.77) |
4.07
(2.02) |
2.33
(1.02) |
| 0.32 mg/kg | 1.48
(0.51) |
2.49
(0.97) |
2.48
(0.82) |
2.58
(0.81) |
1.78
(0.84) |
1.50
(0.80) |
| 1.0 mg/kg | 1.39
(0.73) |
2.75
(1.05) |
1.83
(0.65) |
2.04
(0.93) |
2.77
(1.35) |
1.15
(1.00) |
| 1.8 mg/kg | 0.79
(0.26) |
0.95
(0.34) |
2.55
(1.00) |
2.58
(0.75) |
2.78
(1.33) |
2.50
(1.00) |
Although pretreatments did not alter the latency or rate of responding, the GABAB agonists baclofen and CGP445432 did alter the pattern of responding over time during the 2-hr sessions. Fig. 2 shows within session patterning of injections over time in a representative baboon for each pretreatment (baclofen, CGP44532 and tiagabine) across the different testing conditions. As shown in Fig 2a, when cocaine was available for self-injection, the latency to complete the first response requirement and receive the first injection of cocaine was very short (i.e., within the first few minutes after the session began) and a regular pattern of evenly spaced injections were received throughout the 2 h session. In comparison, during saline extinction (Fig 2b), low numbers of saline injections were taken and the within session pattern of injections was irregular; injections occurred in groups near the beginning of the session. The rate and pattern of responding after vehicle substitution were characterized by greater pausing between bursts of responding; but when responding did occur, the local run rate was generally >1 response/s. Administration of IV priming injections of cocaine during saline extinction reinstated lever responding and the within session pattern of injections was related to the dose of cocaine administered. The increased latency to complete the first response requirement and receive the first self-injection of saline following priming injections of 0.32 and 1.0 mg/kg cocaine (as shown for the group in Table 1) is evident in the patterning of injections shown in Figs 2c and 2d. After the first injection, the pattern of saline injections produced by a priming dose of 0.32 mg/kg cocaine were similar to those maintained by cocaine for the first 60-min but were more erratic over time when compared to the cocaine baseline sessions (Fig 2c). Injections continued to be received throughout the session following the 1.0 mg/kg cocaine priming injection (Fig 2d). When either baclofen (Fig 2c and 2d, left panels) or CGP44532 (Fig 2c and 2d, center panels) were administered before the cocaine priming injection, a similar pattern of saline injections was obtained at the beginning of the session, but responding (and therefore injections received) terminated sooner and few injections were received later in the session. In contrast, when the tiagabine pretreatment was administered prior to the 0.32 mg/kg (Fig 2c right panel), or the 1.0 mg/kg cocaine priming injection (Fig 2d, right panel), the pattern of saline injections was comparable to that following the cocaine priming injections alone (no pretreat).
Figure 2.
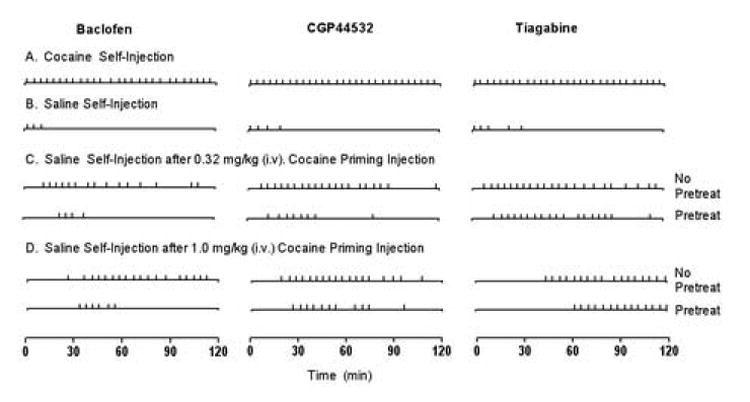
Within session pattern of injections over time and across experimental conditions. Data for a representative baboon are shown for each pretreatment group (Baboon JA for baclofen, Baboon BA for CGP44532, and Baboon KR for tiagabine). Each tick mark along an axis represents an injection of cocaine (A only) or saline (B, C, and D). The cocaine baseline self-injection (A) session and saline extinction sessions (B) were randomly chosen, but were determined to be representative for each baboon. Saline self-injections are shown following priming injections of 0.32 mg/kg cocaine (C) with and without administration of a pretreatment drug and following priming injections of 1.0 mg/kg cocaine (D) with and without administration of a pretreatment drug.
4. Discussion
In the current study, 0.032 mg/kg/injection cocaine maintained high numbers of self-injection (25-30 per session) and a single intravenous priming injection of cocaine increased extinguished lever responding on the cocaine-associated lever when no drug was available (i.e., reinstated cocaine-seeking behavior) in all baboons. As in previous studies in our laboratory (Weerts and Griffiths, 2003), cocaine-seeking increased as a function of the priming dose administered, with the greatest increases in number of saline injections obtained at 1.0-1.8 mg/kg, i.v. cocaine. In the current study, the pattern of responding and injections received over the time course of the session revealed that as the priming dose of cocaine increased, the latency to the first injection also increased. In rats trained to self-inject cocaine under a second order schedule, priming doses of IV cocaine also have been shown to increase the latency to initiate responding when the doses of cocaine administered were similar to or higher than the training/maintenance dose of cocaine (Markou et al., 1999). Thus, the magnitude of the cocaine priming dose administered in relation to the training/maintenance dose of cocaine influences both the pattern drug-seeking behavior, and the latency to initiate responding.
In the current study, pretreatment with baclofen or CGP44532 decreased reinstatement of cocaine-seeking produced by cocaine priming doses, where as tiagabine did not. A number of studies in rodents have shown that GABAB agonists, including baclofen and CGP44532, can reduce cocaine self-administration under various schedules of reinforcement (Brebner et al., 1999; Campbell et al., 1999; Roberts and Andrews, 1997; Roberts et al., 1996; Shoaib et al., 1998). Baclofen pretreatment also has been shown to reduce both the rate of acquisition and the percentage of rats that acquired criterion levels of cocaine self-administration (Campbell et al., 2002). When effects on food-maintained responding were also determined in studies in rats, the effects of baclofen and CGP44532 on food-maintained responding were less than its effects on cocaine self-administration (Brebner et al., 1999; Roberts et al., 1996; Shoaib et al., 1998). In primates, however, the effects of GABAB agonists on the maintenance of cocaine self-administration have been less robust. Recently, we reported that baclofen and CGP44532 suppressed cocaine self-administration and food-maintained responding in a similar manner when ongoing rates of responding were the same for both reinforcers (Weerts et al., 2005). Baclofen also failed to suppress cocaine self-administration in rhesus monkeys even though a high dose (10 mg/kg) of baclofen substantially reduced food-maintained responding and produced motor incoordination (Stafford and Glowa, 2000).
Similar to previous studies in rats (Campbell et al., 1999; Di Ciano and Everitt, 2003), the reinstatement of cocaine seeking produced by priming doses of cocaine was attenuated by low doses of both baclofen and CGP44532 in baboons in the current study. Inspection of the pattern of responding and injections received over the time course of the 2-hour session suggests that neither baclofen nor CGP44532 decreased the likelihood of initiating cocaine-seeking but, once initiated, facilitated the termination (extinction) of responding. The effects of baclofen and CGP44532 in the current study also do not appear to be related to the demonstrated sedative and muscle-relaxant effects of the drugs. The doses of baclofen (0.32 mg/kg) and CGP44532 (0.32 mg/kg) that attenuated cocaine seeking did not produce overt effects and did not significantly reduce local response rates (i.e., run rates) in the current study. As reported previously, lever responding maintained by food using the same 2-h procedure as in the current study were not altered by 0.32 mg/kg baclofen and 0.32 mg/kg CGP44532 (Weerts et al., 2005). In the current study, then, effects were seen at doses that did not produce impairment and therefore suggest that the effects of baclofen and CGP44532 on cocaine-induced reinstatement are not related to motor impairment or other non-specific effects, but are likely related to a decrease in the reinforcing effects of the priming dose of cocaine.
Both baclofen and CGP44532 were more effective at blocking the reinstatement of cocaine-seeking in baboons in the current study when compared to their effects on ongoing cocaine self-administration (Weerts et al., 2005). The current data are consistent with baclofen’s effects on cocaine seeking-behavior in rats. Specifically, baclofen dose-dependently reduced the reinstatement of extinguished drug-taking behavior produced by priming injections of cocaine (Campbell et al., 1999), and lever responding maintained by cocaine-associated cues under a second order schedule (Di Ciano and Everitt, 2003). The efficacy of both GABAB agonists in reducing cocaine seeking may be related to an interaction between extinction and the unit dose of cocaine. Campbell and colleagues (1999) reported that baclofen was more effective in suppressing self-administration behavior maintained by lower doses of cocaine (Campbell et al., 1999). Similarly, a recent laboratory study (Haney et al., 2006) found that baclofen decreased self-administration of a low dose of smoked cocaine (12 mg) without altering self-administration of a higher dose (50 mg) in non-opioid dependent human subjects. Baclofen did decrease subject estimated dollar value of the high dose of smoked cocaine (50 mg), but failed to alter subjective effects (e.g. ratings of ‘High,’ ‘good Drug Effect’ or ‘Stimulated’) produced by smoked cocaine. Thus, when the findings of all of these studies are considered, GABAB agonists may be most effective in reducing relapse to drug taking in the event of re-exposure to low doses of cocaine or cocaine-associated cues.
In a previous study in baboons the effects of GABAergic modulators including tiagabine on behavior maintained by either food or cocaine were investigated using the same 2-hr procedure (Weerts et al., 2005). Similar numbers of reinforcers per session were maintained by both food and cocaine. In that study, tiagabine (1.0 mg/kg) reduced both cocaine self-administration and food-maintained behavior, although there was a trend toward greater suppression of cocaine-maintained behavior. Still, the significant decrease in food-maintained behavior indicates concurrent motor impairment or other nonspecific effects. The dose selected for reinstatement tests in the current procedure was one that did not produce non-specific effects (i.e., did not decrease food-maintained behavior) when administered alone (Weerts et al., 2005). In the current study, tiagabine did not reduce cocaine-seeking and also did not systematically affect the response latency, response rates or patterns of injections produced by the priming injections of cocaine. Thus, tiagabine failed to reduce cocaine seeking at doses that did not also produce non-specific effects. While higher doses of tiagabine may have decreased the number of saline injections during extinction in the present procedure, it would be unclear whether this was a result of a non-specific suppression of responding. The results in human studies have also been mixed. One human laboratory study found that tiagabine did not alter cocaine-induced increases in heart rate and blood pressure but did reduce some subjective effects of intravenously administered cocaine (i.e., decreased subject ratings of “stimulated” and “craving” but not ratings of “high” or “feel effects of last dose”) (Sofuoglu et al., 2005). In contrast, another laboratory study found that tiagabine failed to alter the subjective or reinforcing effects of cocaine and cocaine-induced increases in heart rate and blood pressure (Lile et al., 2004). Preliminary placebo-controlled clinical trials have reported tiagabine reduced cocaine use, although effects were mostly observed after 5-8 weeks of tiagabine treatment (Gonzalez et al., 2003; Winhusen et al., 2005). Thus, it can not be ruled out that long-term administration, dose stabilization and the possible development of tolerance to motor effects of tiagabine may be required for treatment efficacy in cocaine dependent subjects.
In conclusion, the current findings for the GABAB agonists CGP44532 and baclofen are consistent with data from preliminary clinical trials for baclofen in cocaine dependent subjects. Baclofen administration combined with group therapy has been reported to reduce self-reported cocaine use and increase cocaine negative urines in cocaine-dependent subjects in an open labeled clinical trial (Ling et al., 1998) and in a 16-week double-blind study (Shoptaw et al., 2003). The current data in baboons suggest that GABAB agonists such as baclofen and the less sedative CGP44532, may be useful in decreasing relapse to cocaine taking in abstinent patients by facilitating extinction in the case of a “slip.”
Acknowledgments
The authors thank Samuel Womack for excellent technical assistance. This research was supported by NIH/NIDA grant R01 DA13621 (EMW). The National Institute on Drug Abuse Drug Supply Program provided cocaine. CGP44532 was provided as a gift from Novartis Pharma AG and tiagabine was provided as a gift from Cephalon Inc.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Arroyo M, Markou A, Robbins TW, Everitt BJ. Acquisition, maintenance and reinstatement of intravenous cocaine self-administration under a second-order schedule of reinforcement in rats: effects of conditioned cues and continuous access to cocaine. Psychopharmacology (Berl) 1998;140:331–344. doi: 10.1007/s002130050774. [DOI] [PubMed] [Google Scholar]
- Bardo MT. Neuropharmacological mechanisms of drug reward: beyond dopamine in the nucleus accumbens. Crit Rev Neurobiol. 1998;12:37–67. doi: 10.1615/critrevneurobiol.v12.i1-2.30. [DOI] [PubMed] [Google Scholar]
- Brebner K, Childress AR, Roberts DC. A potential role for GABA(B) agonists in the treatment of psychostimulant addiction. Alcohol. 2002;37:478–484. doi: 10.1093/alcalc/37.5.478. [DOI] [PubMed] [Google Scholar]
- Brebner K, Froestl W, Andrews M, Phelan R, Roberts DC. The GABA(B) agonist CGP 44532 decreases cocaine self-administration in rats: demonstration using a progressive ratio and a discrete trials procedure. Neuropharmacology. 1999;38:1797–1804. doi: 10.1016/s0028-3908(99)00094-5. [DOI] [PubMed] [Google Scholar]
- Campbell UC, Lac ST, Carroll ME. Effects of baclofen on maintenance and reinstatement of intravenous cocaine self-administration in rats. Psychopharmacology (Berl) 1999;143:209–214. doi: 10.1007/s002130050937. [DOI] [PubMed] [Google Scholar]
- Campbell UC, Morgan AD, Carroll ME. Sex differences in the effects of baclofen on the acquisition of intravenous cocaine self-administration in rats. Drug Alcohol Depend. 2002;66:61–69. doi: 10.1016/s0376-8716(01)00185-5. [DOI] [PubMed] [Google Scholar]
- Carroll ME, Comer SD. Animal models of relapse. Exp Clin Psychopharmacol. 1996;4:11–18. [Google Scholar]
- Childress AR, Franklin T, McElgin W, Acton P, O’Brien CP. College on the Problems of Drug Dependence. San Juan: Puerto Rico; 2000. GABAergics may blunt limbic activation during cue-induced cocaine craving. [Google Scholar]
- Childress AR, Mozley PD, McElgin W, Fitzgerald J, Reivich M, O’Brien CP. Limbic activation during cue-induced cocaine craving. Am J Psychiatry. 1999;156:11–18. doi: 10.1176/ajp.156.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cousins MS, Roberts DC, de Wit H. GABA(B) receptor agonists for the treatment of drug addiction: a review of recent findings. Drug Alcohol Depend. 2002;65:209–220. doi: 10.1016/s0376-8716(01)00163-6. [DOI] [PubMed] [Google Scholar]
- Davis WM, Smith SG. Role of conditioned reinforcers in the initiation, maintenance, and extinction of drug-seeking behavior. Pavlov J Biol Sci. 1976;11:222–236. doi: 10.1007/BF03000316. [DOI] [PubMed] [Google Scholar]
- deWit H, Stewart J. Reinstatement of cocaine-reinforced responding in the rat. Psychopharmacology. 1981;75:134–143. doi: 10.1007/BF00432175. [DOI] [PubMed] [Google Scholar]
- Di Ciano P, Everitt BJ. The GABA(B) receptor agonist baclofen attenuates cocaine- and heroin-seeking behavior by rats. Neuropsychopharmacology. 2003;28:510–518. doi: 10.1038/sj.npp.1300088. [DOI] [PubMed] [Google Scholar]
- Dobrovitsky V, Pimentel P, Duarte A, Froestl W, Stellar JR, Trzcinska M. CGP 44532, a GABAB receptor agonist, is hedonically neutral and reduces cocaine-induced enhancement of reward. Neuropharmacology. 2002;42:626–632. doi: 10.1016/s0028-3908(02)00007-2. [DOI] [PubMed] [Google Scholar]
- Fadda P, Scherma M, Fresu A, Collu M, Fratta W. Baclofen antagonizes nicotine-, cocaine-, and morphine-induced dopamine release in the nucleus accumbens of rat. Synapse. 2003;50:1–6. doi: 10.1002/syn.10238. [DOI] [PubMed] [Google Scholar]
- Franklin T, Kampman K, Ehrman R, O’Brien C, Childress AR. College on the Problems of Drug Dependence. San Juan: Puerto Rico; 2000. Impact of GABAergics in a novel “cue + cocaine challenge” paradigm. [Google Scholar]
- Froestl W, Mickel SJ, Bittiger H. Potent GABAB agonists and antagonists. Curr Opin Ther Patents. 1993;3:561–567. [Google Scholar]
- Froestl W, Mickel SJ, Hall RG, et al. Phosphinic acid analogues of GABA. New potent and selective GABAB agonists. J Med Chem. 1995;38:3297–3312. doi: 10.1021/jm00017a015. [DOI] [PubMed] [Google Scholar]
- Gasior M, Kaminski R, Witkin JM. Pharmacological modulation of GABA(B) receptors affects cocaine-induced seizures in mice. Psychopharmacology (Berl) 2004;174:211–219. doi: 10.1007/s00213-003-1743-0. [DOI] [PubMed] [Google Scholar]
- Gerber GJ, Stretch R. Drug-induced reinstatement of extinguished self-administration behavior in monkeys. Pharmacol Biochem Behav. 1975;3:1055–1061. doi: 10.1016/0091-3057(75)90016-7. [DOI] [PubMed] [Google Scholar]
- Goldberg SR, Morse WH, Goldberg DM. Behavior maintained under a second-order schedule by intramuscular injection of morphine or cocaine in rhesus monkeys. J Pharmacol Exp Ther. 1976;199:278–286. [PubMed] [Google Scholar]
- Gonzalez G, Sevarino K, Sofuoglu M, Poling J, Oliveto A, Gonsai K, George TP, Kosten TR. Tiagabine increases cocaine-free urines in cocaine-dependent methadonetreated patients: results of a randomized pilot study. Addiction. 2003;98:1625–1632. doi: 10.1046/j.1360-0443.2003.00544.x. [DOI] [PubMed] [Google Scholar]
- Haney M, Hart CL, Foltin RW. Effects of baclofen on cocaine self-administration: opioid- nd non-opioid dependent volunteers. Neuropsychopharm. 2006;31:1814–1821. doi: 10.1038/sj.npp.1300999. [DOI] [PubMed] [Google Scholar]
- Johanson CE. Behavior maintained under fixed-interval and second-order schedules of cocaine or pentobarbital in rhesus monkeys. J Pharmacol Exp Ther. 1982;221:384–393. [PubMed] [Google Scholar]
- Katz JL. A comparison of responding maintained under second-order schedules of intramuscular cocaine injection or food presentation in squirrel monkeys. J Exp Anal Behav. 1979;32:419–431. doi: 10.1901/jeab.1979.32-419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ling W, Shoptaw S, Majewska D. Baclofen as a cocaine anti-craving medication: a preliminary clinical study. Neuropsychopharmacology. 1998;18:403–404. doi: 10.1016/S0893-133X(97)00128-0. letter. [DOI] [PubMed] [Google Scholar]
- Lile JA, Stoops WW, Glaser PE, Hays LR, Rush CR. Acute administration of the GABA reuptake inhibitor tiagabine does not alter the effects of oral cocaine in humans. Drug Alcohol Depend. 2004;76:81–91. doi: 10.1016/j.drugalcdep.2004.04.010. [DOI] [PubMed] [Google Scholar]
- Lukas SE, Griffiths RR, Bradford LD, Brady JV, Daley L. A tethering system for intravenous and intragastric drug administration in the baboon. Pharmacol Biochem Behav. 1982;17:823–829. doi: 10.1016/0091-3057(82)90366-5. [DOI] [PubMed] [Google Scholar]
- Markou A, Arroyo M, Everitt BJ. Effects of contingent and non-contingent cocaine on drug-seeking behavior measured using a second-order schedule of cocaine reinforcement in rats. Neuropsychopharm. 1999;20:542–555. doi: 10.1016/S0893-133X(98)00080-3. [DOI] [PubMed] [Google Scholar]
- Markou A, Weiss F, Gold LH, Caine SB, Schulteis G, Koob GF. Animal models of drug craving. Psychopharmacology. 1993;112:163–182. doi: 10.1007/BF02244907. [DOI] [PubMed] [Google Scholar]
- Norman AB, Norman MK, Hall JF, Tsibulsky VL. Priming threshold: a novel quantitative measure of the reinstatement of cocaine self-administration. Brain Res. 1999;831:165–174. doi: 10.1016/s0006-8993(99)01423-7. [DOI] [PubMed] [Google Scholar]
- Roberts DC, Andrews MM. Baclofen suppression of cocaine self-administration: demonstration using a discrete trials procedure. Psychopharmacology (Berl) 1997;131:271–277. doi: 10.1007/s002130050293. [DOI] [PubMed] [Google Scholar]
- Roberts DC, Andrews MM, Vickers GJ. Baclofen attenuates the reinforcing effects of cocaine in rats. Neuropsychopharmacology. 1996;15:417–423. doi: 10.1016/0893-133X(96)00002-4. [DOI] [PubMed] [Google Scholar]
- Schindler CW, Panlilio LV, Goldberg SR. Second-order schedules of drug self-administration in animals. Psychopharmacology (Berl) 2002;163:327–344. doi: 10.1007/s00213-002-1157-4. [DOI] [PubMed] [Google Scholar]
- Self DW, Nestler EJ. Relapse to drug-seeking: neural and molecular mechanisms. Drug Alcohol Depend. 1998;51:49–60. doi: 10.1016/s0376-8716(98)00065-9. [DOI] [PubMed] [Google Scholar]
- Shoaib M, Swanner LS, Beyer CE, Goldberg SR, Schindler CW. The GABAB agonist baclofen modifies cocaine self-administration in rats. Behav Pharmacol. 1998;9:195–206. [PubMed] [Google Scholar]
- Shoptaw S, Yang X, Rotheram-Fuller EJ, Hsieh YC, Kintaudi PC, Charuvastra VC, Ling W. Randomized placebo-controlled trial of baclofen for cocaine dependence: preliminary effects for individuals with chronic patterns of cocaine use. J Clin Psychiatry. 2003;64:1440–1448. doi: 10.4088/jcp.v64n1207. [DOI] [PubMed] [Google Scholar]
- Sidman M. Tactics of Scientific Research. Basic Books; New York: 1960. [Google Scholar]
- Slikker W, Jr, Brocco MJ, Killam KF., Jr Reinstatement of responding maintained by cocaine or thiamylal. J Pharmacol Exp Ther. 1984;228:43–52. [PubMed] [Google Scholar]
- Sofuoglu M, Kosten TR. Novel approaches to the treatment of cocaine addiction. CNS Drugs. 2005;19:13–25. doi: 10.2165/00023210-200519010-00002. [DOI] [PubMed] [Google Scholar]
- Sofuoglu M, Kosten TR. Emerging pharmacological strategies in the fight against cocaine addiction. Expert Opin Emerg Drugs. 2006;11:91–98. doi: 10.1517/14728214.11.1.91. [DOI] [PubMed] [Google Scholar]
- Sofuoglu M, Poling J, Mitchell E, Kosten TR. Tiagabine affects the subjective responses to cocaine in humans. Pharmacol Biochem Behav. 2005;82:569–573. doi: 10.1016/j.pbb.2005.10.013. [DOI] [PubMed] [Google Scholar]
- Spealman RD, Barrett-Larimore RL, Rowlett JK, Platt DM, Khroyan TV. Pharmacological and environmental determinants of relapse to cocaine-seeking behavior. Pharmacol Biochem Behav. 1999;64:327–336. doi: 10.1016/s0091-3057(99)00049-0. [DOI] [PubMed] [Google Scholar]
- Stafford D, Glowa JR. College on the Problems of Drug Dependence. San Juan: Puerto Rico; 2000. Effects of intravenous baclofen pretreatment on food- and cocaine-maintained responding in rhesus monkeys. [Google Scholar]
- Suzdak PD, Jansen JA. A review of the preclinical pharmacology of tiagabine: a potent and selective anticonvulsant GABA uptake inhibitor. Epilepsia. 1995;36:612–626. doi: 10.1111/j.1528-1157.1995.tb02576.x. [DOI] [PubMed] [Google Scholar]
- Weerts EM, Froestl W, Griffiths RR. Effects of GABAergic modulators on food and cocaine self-administration in baboons. Drug Alcohol Depend. 2005;80:369–376. doi: 10.1016/j.drugalcdep.2005.05.006. [DOI] [PubMed] [Google Scholar]
- Weerts EM, Griffiths RR. Cocaine, caffeine and CGS15943 reinstate extinguished cocaine-taking behavior in baboons. Society for Neuroscience Abstracts. 1999;25:305. [Google Scholar]
- Weerts EM, Griffiths RR. The adenosine receptor antagonist CGS15943 reinstates cocaine-seeking behavior and maintains self-administration in baboons. Psychopharmacology (Berl) 2003;168:155–163. doi: 10.1007/s00213-003-1410-5. [DOI] [PubMed] [Google Scholar]
- Winhusen TM, Somoza EC, Harrer JM, Mezinskis JP, Montgomery MA, Goldsmith RJ, Coleman FS, Bloch DA, Leiderman DB, Singal BM, Berger P, Elkashef A. A placebo-controlled screening trial of tiagabine, sertraline and donepezil as cocaine dependence treatments. Addiction. 2005;100(Suppl 1 ):68–77. doi: 10.1111/j.1360-0443.2005.00992.x. [DOI] [PubMed] [Google Scholar]