

Short abstract
Recent microarray studies of mouse and human osteoblast differentiation in vitro have identified novel transcription factors that may be important in the establishment and maintenance of differentiation.
Abstract
Recent microarray studies of mouse and human osteoblast differentiation in vitro have identified novel transcription factors that may be important in the establishment and maintenance of differentiation. These findings help unravel the pattern of gene-expression changes that underly the complex process of bone formation.
Osteoblasts, the bone-forming cells, have the unique function of producing and then mineralizing the bone matrix. Although substantial progress has been made in understanding the molecular basis of osteoblast differentiation and function, many aspects remain unknown. Osteoblasts differentiate from their mesenchymal precursors in a complex process that is orchestrated by the timely activation of specific transcription factors that regulate the expression of certain genes and thus define the osteoblast phenotype. The genes encoding two of these transcription factors, Runx2/Cbfa1 and Osterix (Osx), have been identified as master controllers of the osteoblastic lineage, and the absence of either one results in a complete lack of a mineralized skeleton [1-3]. Many other transcription factors have been shown to regulate osteoblast function, including homeobox proteins: MSX1, MSX2, DLX3, and DLX5; members of the AP1 family; C/EBP β, CBFB, Twist and, more recently, effectors of the β-catenin/Wnt signaling pathway (reviewed in [4-6]). As new transcription factors that regulate Osteoblasts are discovered and the complexity of the osteoblast-differentiation program becomes more apparent, we can see that our current picture of this process is partial, and a unified view of the interplay and timing of the different transcriptional regulators is still elusive.
Genome-scale analysis of osteoblast differentiation
Microarray analysis has recently been applied by several investigators in an attempt to further understand the molecular programs that define osteoblast differentiation. Several cellular models have been used, including committed osteogenic precursors of murine and human origin [7-10], immortalized human cells at various stages of differentiation [11], and uncommitted mesodermal progenitor cells [12-16]. The variety of cell sources and models underscore a potential difficulty with comparing studies that use this approach. A bone-forming cell goes through many phases in its lifetime, from early commitment to organic-matrix production, mineralization and apoptosis or terminal differentiation into an osteocyte, and its gene-expression profile varies widely depending upon the differentiation stage. Changes in gene expression must therefore be interpreted in a way that takes into consideration the cell context and differentiation phase.
A second limitation of the current microarray methods stems from the need to use a stimulator to induce osteoblast differentiation in vitro. In the classic murine calvaria (cranial vault) cell model, ascorbic acid and β-glycerolphosphate are used to induce matrix production and mineralization. Other models, in particular human osteoblasts, require 'stronger' stimulators, such as bone morphogenetic protein-2 or dexamethasone, which have specific and often diverse modulatory effects on osteoblast gene expression, thus complicating the distinction between effects of the stimulator and changes due to osteoblast differentiation. As a paradigm for this type of study, we will focus on the recent work of Qi et al. [12], who used a unique model of human osteoblasts.
Novel transcription factors involved in osteoblast differentiation
The cell model used by Qi et al. [12] is based on mesenchymal progenitor cells isolated from the marrow of human donors. These mesenchymal progenitor cells can be induced towards the osteoblast lineage by incubation with dexamethasone in the presence of ascorbic acid and β-glycerolphosphate. Using a commercially available microarray, the authors compared the profile of genes expressed in undifferentiated mesenchymal progenitor cells (baseline) to those induced in the presence of the stimulators over a seven-day time course. As a critically important validation step, genes previously proven to be hallmarks of differentiated osteoblasts, including osteocalcin, type I collagen, RUNX2, MSX2 and alkaline phosphatase, were found to be up-regulated with time in culture after osteogenic stimulation. The extent of up-regulation of many of these genes was only marginal, however, and some did not even meet the criteria of significant change, which was set at greater than two-fold increase relative to the baseline. This result highlights another major limitation of the method: given that the expression of proven master genes may not change dramatically, as in this case, what is the correct threshold for deciding whether a change in mRNA abundance is significant? More to the point, is a change in mRNA levels a real index of the importance of a gene in cell function?
Despite these limitations, microarray data do provide important novel information. Among the genes found to be differentially expressed (greater or less than two-fold) in the study by Qi et al. [12], 41 encoded transcription factors, many of which were not previously known to be involved in osteoblast differentiation, yet they were found to co-cluster in expression profile with osteoblast-specific transcription factors. Table 1 lists the transcription factors most abundantly regulated during osteoblast differentiation in the study by Qi et al. [12]. A number of little-characterized zinc-finger proteins were upregulated throughout the process of osteoblast differentiation, including many Znf family members (Znf9 is associated with myotonic dystrophy [17], and Znf74 with DiGeorge syndrome; DiGeorge syndrome is characterized by the absence or hypoplasia of the thymus and parathyroid glands, resulting in hypocalcemia, heart defects, short stature and craniofacial deformities [18]). ZNF133, ZNF143 and ZNF177 exhibited the most robust up-regulation throughout the seven-day time course, peaking at levels 3.07-, 3.08- and 5-77-fold that of baseline, respectively. In contrast, ZNF6 and EGR1 were markedly down-regulated. Interestingly, the roles of these zinc-finger proteins have not been studied in bone, although their potential importance for bone-forming cells was heralded by the identification of OSX, another zinc-finger protein that is critical to osteoblast function, which acts downstream of RUNX2/CBFA1 [1]. Other structurally related zinc-finger family members, SP1 and SP3, are also emerging as strong regulators of osteoblast genes [19,20], thus highlighting the critical role of this class of transcriptionally active factors in osteoblast commitment and differentiation. The finding that members of the zinc-finger protein family are crucial to the process of osteoblast differentiation represents an important contribution originating from gene array studies.
Table 1.
Transcription factors most abundantly regulated during osteoblast differentiation
| Gene | Full name | Fold change |
| Increased | ||
| DGSI | DiGeorge syndrome critical region gene | 7.10 |
| ZNF177 | Zinc-finger protein 177 | 5.77 |
| NCOR2 | Nuclear receptor co-repressor 2 | 4.92 |
| CRABP1 | Cellular retinoic acid binding protein 1 | 4.29 |
| IRX2A | Iroquois-class homeobox protein | 3.84 |
| NR1H2 | Nuclear receptor subfamily 1, group H, member 2 | 3.65 |
| GATA1 | Globin transcription factor 1 | 3.40 |
| POU2F1 | POU domain, class 2, transcription factor 1 | 3.16 |
| MAD4 | Mad4 homolog | 3.08 |
| ZNF143 | Zinc-finger protein 143 (clone pHZ-1) | 3.08 |
| ZNF133 | Zinc-finger protein 133 (clone pHZ-13) | 3.07 |
| SHOX2 | Short stature homeobox 2 | 3.01 |
| Decreased | ||
| MYC | v-Myc homolog | 5.00 |
| ZNF6 | Zinc-finger protein 6 | 3.57 |
| CA150 | Transcription factor CA150 | 3.33 |
| TCEB1 | Transcription elongation factor B (Elongin C) | 3.23 |
The data are from the microarray study of Qi et al. [12] and fold change describes the relative expression of the given factor in osteoblasts relative to expression in the mesenchymal progenitor cells.
Several of the mammalian Sox genes, encoding homeobox-containing transcription factors related to the SRY sex-determining gene, are involved in regulating chondrocyte (cartilage-forming cell) differentiation and function during endochondral ossification. Qi et al. [12] show that SOX-4 and SOX-22 are also upregulated during osteoblast differentiation from mesenchymal progenitor cells. SOX-4 is of particular interest since it is expressed in hypertrophic chondrocytes (cartilage-forming cells) at the zone of mineralizing cartilage and in osteoblasts. Further, SOX-4 expression in the skeletal tissue is modulated by parathyroid hormone, a critical regulator of calcium and bone metabolism [21]. Up-regulation of SOX-4 has also been reported by two other microarray studies of osteoblastogenesis [13,16].
Homeobox transcription factors - in particular MSX1, MSX2, DLXs, DLXs, ALX4, SHOX and HOXD13 - coordinate skeletal patterning and modulate the mature function of osteoblasts. Additional homeobox factors identified by Qi et al. [12] as being up-regulated during differentiation of osteoblasts from mesenchymal progenitor cells include IRX2A, POU2F1, SHOX2 and HOXB6. The Iroquois class of homeobox proteins, IRX1 and IRX2, are highly expressed during digit formation in mouse embryos [22]; IRX2 is also expressed in the vertebrae and developing skull, and although it is not specific for the skeleton, it is up-regulated (more than 3.8-fold) during osteoblastogenesis [12]. The idea of a role for the Iroquois homeobox genes in bone formation is strengthened by the observation that IRX3 expression increases during osteoblast differentiation [8]. Another intriguing factor is SHOX2 (short stature homeobox 2), which is up-regulated three-fold and has a similar expression profile to that of its homolog, SHOX, which is associated with skeletal abnormalities in the sex-chromosome disorder Turner syndrome [23].
Interestingly, the work of Qi et al. [12] also reveals that a number of hematopoiesis-associated transcription factors are markedly upregulated during osteoblastogenesis. In addition to HOXB6, the expression of GATA1, GATA3 and the Kruppel-like factor KLF1 increases during osteogenic differentiation, thus offering new clues as to the interaction between the hematopoietic and mesenchymal lineages. As further proof of this link, four transcription factors associated with DiGeorge syndrome were found to be up-regulated during osteoblastogenesis. Apart from ZNF74, noted above, DGSI (seven-fold increase), PNUTL1 and PNUT2 (2.49 and 2.29-fold increase, respectively) all are upregulated in the human mesenchymal progenitor cell differentiation model. Given the association of DiGeorge syndrome with hypoparathyroidism and the attendant skeletal abnormalities, it is likely that these four transcription factors may be important modulators of parathyroid hormone signaling in bone.
In summary, the use of genome-wide approaches to identify genes involved in the differentiation of osteoblasts provides a snapshot of this cell's inventory of transcription factors. But, extracting a biological function from this inventory offers an even greater challenge (Figure 1). In fact, if one compares the results of the different microarray analyses published so far, one will see that rarely do two studies produce similar results; the use of different arrays, different methods of data analysis, different cell models and stimulators and, last but not least, different criteria for defining a change as significant, all contribute to this large variability. While a wealth of new information can be gathered by genome-wide approaches, a systematic molecular analysis of the genes identified by these methods is mandatory if we are to fully understand the real biological role of the newly identified 'osteoblast' genes.
Figure 1.
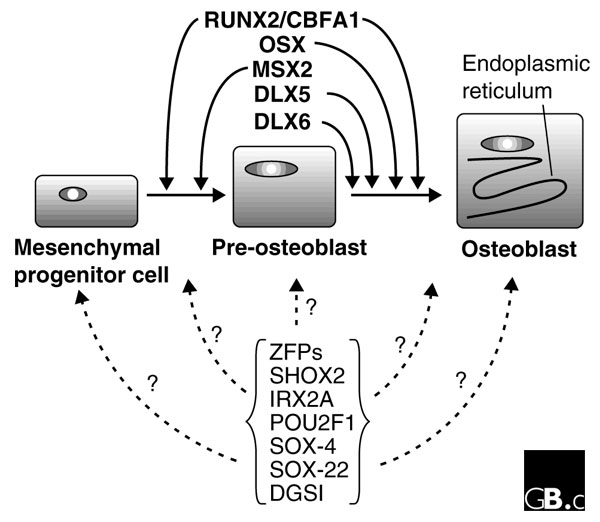
Transcriptional control of osteoblastogenesis. The transcription factors RUNX2, OSX, MSX2, DLX5 and DLX6 play critical roles in coordinating the differentiation from a mesenchymal progenitor cell into a fully differentiated osteoblast. On the basis of the changes in relative abundance of mRNAs and the occurrence of mutants with phenotypes that affect bone formation, a family of zinc-finger proteins (ZFPs) and the factors SHOX2, IRX2A, POU2F1, SOX-4, SOX-22 and DGSI are new likely candidates to be modulators of the progression through the osteoblast lineage from mesenchymal progenitor cells, although the precise step(s) at which they act is not currently known. See text for further details.
Acknowledgments
Acknowledgements
This work has been supported by National Institutes of Health grants R01 AR42155, AR43470 (to RC) and T32 AR07033 (to JPS), and by a grant from the National Aeronautics and Space Administration (NRA 99-HESD-02-110).
References
- Nakashima K, Zhou Z, Kunkel G, Zhang Z, Deng JM, Behringer RR, De Crombrugghe B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell. 2002;108:17–29. doi: 10.1016/s0092-8674(01)00622-5. [DOI] [PubMed] [Google Scholar]
- Otto F, Thornell AP, Crompton T, Denzel A, Gilmour KC, Rosewell IR, Stamp GW, Beddington RS, Mundlos S, Olsen BR, et al. Cbfa1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell. 1997;89:765–771. doi: 10.1016/s0092-8674(00)80259-7. [DOI] [PubMed] [Google Scholar]
- Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell. 1997;89:755–764. doi: 10.1016/s0092-8674(00)80258-5. [DOI] [PubMed] [Google Scholar]
- Yang X, Karsenty G. Transcription factors in bone: developmental and pathological aspects. Trends Mol Med. 2002;8:340–345. doi: 10.1016/S1471-4914(02)02340-7. [DOI] [PubMed] [Google Scholar]
- Sandell LJ, Towler DA. Transcription of bone and cartilage genes. Curr Opin Orthop. 2002;13:375–381. doi: 10.1097/00001433-200210000-00008. [DOI] [Google Scholar]
- De Crombrugghe B, Lefebvre V, Nakashima K. Regulatory mechanisms in the pathways of cartilage and bone formation. Curr Opin Cell Biol. 2001;13:721–727. doi: 10.1016/S0955-0674(00)00276-3. [DOI] [PubMed] [Google Scholar]
- Raouf A, Seth A. Discovery of osteoblast-associated genes using cDNA microarrays. Bone. 2002;30:463–471. doi: 10.1016/S8756-3282(01)00699-8. [DOI] [PubMed] [Google Scholar]
- Seth A, Lee BK, Qi S, Vary CPH. Coordinate expression of novel genes during osteoblast differentiation. J Bone Miner Res. 2000;15:1683–1696. doi: 10.1359/jbmr.2000.15.9.1683. [DOI] [PubMed] [Google Scholar]
- Beck GR, Zerler B, Moran E. Gene array analysis of osteoblast differentiation. Cell Growth Differ. 2001;12:61–83. [PubMed] [Google Scholar]
- Doi M, Nagano A, Nakamura Y. Genome-wide screening by cDNA microarray of genes associated with matrix mineralization by human mesenchymal stem cells in vitro. Biochem Biophys Res Commun. 2002;290:381–390. doi: 10.1006/bbrc.2001.6196. [DOI] [PubMed] [Google Scholar]
- Billiard J, Moran RA, Whitley MZ, Chatterjee-Kishore M, Gillis K, Brown EL, Komm BS, Bodine PVN. Transcriptional profiling of human osteoblast differentiation. J Cell Biochem. 2003;89:389–400. doi: 10.1002/jcb.10514. [DOI] [PubMed] [Google Scholar]
- Qi H, Aguiar DJ, Williams SM, La Pean A, Pan W, Verfaillie CM. Identification of genes responsible for osteoblast differentiation from human mesodermal progenitor cells. Proc Natl Acad Sci USA. 2003;100:3305–3310. doi: 10.1073/pnas.0532693100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Locklin RM, Riggs BL, Hicok KC, Horton HF, Byrne MC, Khosla S. Assessment of gene regulation by bone morphogenetic protein 2 in human marrow stromal cells using gene array technology. J Bone Miner Res. 2001;16:2192–2204. doi: 10.1359/jbmr.2001.16.12.2192. [DOI] [PubMed] [Google Scholar]
- Vaes BLT, Dechering KJ, Feijen A, Hendriks JMA, Lefevre C, Mummery CL, Olijve W, Van Zoelen EJJ, Steegenga WT. Comprehensive microarray analysis of bone morphogenetic protein 2-induced osteoblast differentiation resulting in the identification of novel markers of bone development. J Bone Miner Res. 2002;17:2106–2118. doi: 10.1359/jbmr.2002.17.12.2106. [DOI] [PubMed] [Google Scholar]
- De Jong DS, Van Zoelen EJJ, Bauerschmidt S, Olijve W, Steegenga WT. Microarray analysis of bone morphogenetic protein, transforming growth factor b, and activin early response genes during osteoblast differentiation. J Bone Miner Res. 2002;17:2119–2129. doi: 10.1359/jbmr.2002.17.12.2119. [DOI] [PubMed] [Google Scholar]
- Balint E, Lapointe D, Drissi H, van der Meijden C, Young DW, Van Wijnen AJ, Stein JL, Stein GS, Lian JB. Phenotype discovery by gene expression profiling: mapping of biological processes linked to BMP-2-mediated osteoblast differentiation. J Cell Biochem. 2003;89:401–426. doi: 10.1002/jcb.10515. [DOI] [PubMed] [Google Scholar]
- Liquori CL, Ricker K, Moseley ML, Jacobsen JF, Kress W, Naylor SL, Day JW, Ranum LPW. Myotonic dystrophy type 2 caused by a CCTG expansion in intron 1 of ZNF9. Science. 2001;293:864–867. doi: 10.1126/science.1062125. [DOI] [PubMed] [Google Scholar]
- Aubry M, Demczuk S, Desmaze C, Aikem M, Julien JP, Rouleau GA. Isolation of a zinc finger gene consistently deleted in DiGeorge syndrome. Hum Mol Gen. 1993;2:1583–1587. doi: 10.1093/hmg/2.10.1583. [DOI] [PubMed] [Google Scholar]
- Stains JP, Lecanda F, Screen J, Towler DA, Civitelli R. Gap junctional communication modulates gene transcription by altering the recruitment of Sp1 and Sp3 to connexin-response elements in osteoblast promoters. J Biol Chem. 2003;278:24377–24387. doi: 10.1074/jbc.M212554200. [DOI] [PubMed] [Google Scholar]
- Yeung F, Law WK, Yeh CH, Westendorf JJ, Zhang Y, Wang R, Kao C, Chung LWK. Regulation of human osteocalcin promoter in hormone-independent human prostate cancer cells. J Biol Chem. 2002;277:2468–2476. doi: 10.1074/jbc.M105947200. [DOI] [PubMed] [Google Scholar]
- Reppe S, Rian E, Jemtland R, Olstad OK, Gautvik VT, Gautvik KM. Sox-4 messenger RNA is expressed in the embryonic growth plate and regulated via the parathroid hormone/parathyroid homrone-related protein receptor in osteoblast-like cells. J Bone Miner Res. 2000;15:2402–2412. doi: 10.1359/jbmr.2000.15.12.2402. [DOI] [PubMed] [Google Scholar]
- Zulch A, Becker MB, Gruss P. Expression pattern of Irx1 and Irx2 during mouse digit development. Mech Dev. 2001;106:159–162. doi: 10.1016/S0925-4773(01)00411-7. [DOI] [PubMed] [Google Scholar]
- Clement-Jones M, Schiller S, Rao E, Blaschke RJ, Zuniga A, Zeller R, Robson SC, Binder G, Glass I, Strachan T, et al. The short stature homeobox gene SHOX is involved in skeletal abnormalities in Turner syndrome. Hum Mol Gen. 2000;9:695–702. doi: 10.1093/hmg/9.5.695. [DOI] [PubMed] [Google Scholar]