

Abstract
Interleukin 15 (IL-15) mRNA is expressed in a wide variety of tissue types. However, with the exception of some T cell lines, IL-15 transcript expression has not been described in T cells. Herein we demonstrate that IL-15 mRNA can be detected in freshly isolated normal T cells and T cell lines. Furthermore, its expression is 3- to 4-fold higher in human T cell lymphotropic virus type I (HTLV-I)-infected T cells. By using reporter constructs bearing the 5′ regulatory region of the IL-15 gene, we observed a positive correlation between HTLV-I Tax protein expression and IL-15 promoter activity in HTLV-I-infected T cells. Additionally, by using a Jurkat T cell transfectant that expresses Tax under an inducible promoter, we demonstrated that the expression of IL-15 mRNA increased 3-fold as Tax was expressed, suggesting that the Tax protein activates IL-15 transcription. An NF-κB consensus sequence is located at the −75 and −65 region of the IL-15 5′ regulatory region. Mutations in the NF-κB motif or deletion of this sequence abrogated the promoter activity in both HTLV-I-positive and Jurkat Tax-transfectant cells. These data represent evidence for trans-activation of the IL-15 gene by the HTLV-I Tax protein through an NF-κB motif and suggest a potential role for IL-15 in HTLV-I-associated diseases such as adult T cell leukemia and HTLV-I-associated myopathy/tropical spastic paraparesis.
Interleukin 15 (IL-15) is a cytokine whose functional characteristics are similar to those of interleukin 2 (IL-2), including its ability to stimulate T and B cell proliferation, induce lymphokine-activated killer (LAK) cells and enhance B cell differentiation (1–3). There are also reports supporting a role for IL-15 in natural killer cell development and function (4–6). In contrast to IL-2, which is predominantly produced by activated T cells, IL-15 mRNA appears to be more abundantly expressed in a wide variety of other cells and tissues including skeletal muscle (7), epithelial cells (8), astrocytes and macroglia (9), macrophages (10), and monocytes (3, 11). Herein, in contrast to previous reports (3), we provide evidence that T cells also express IL-15 mRNA.
Infection of T cells by human T cell lymphotropic virus type I (HTLV-I) immortalizes these cells as a prelude to adult T cell leukemia (ATL) (12). It had been suggested that transformation of T cells in the early phase of ATL is mediated by the HTLV-I Tax protein (13). The Tax protein is encoded by the pX region of HTLV-I and is essential for viral gene expression by driving its transcription through the cAMP-responsive element (CRE) in the viral long terminal repeat (14). Furthermore, Tax is known to trans-activate many cellular growth-promoting genes, including the IL-2, IL-2 receptor α (IL-2Rα) (15), and the c-fos protooncogene (16). The activation of cellular and viral genes is mediated through a number of cis-acting DNA elements including CRE (14), serum-responsive element, and NF-κB (16, 17).
NF-κB defines a family of dimeric transcription factors composed of combinations of members of the Rel/NF-κB family such as p50, p52, p65 (RelA), c-Rel, and RelB (18, 19). The predominant species is the p50–p65 complex, which is retained in the cytoplasm by its inhibitor IκB-α. Particular cell stimuli such as mitogens, cytokines, and HTLV-I Tax protein cause proteolytic degradation of the IκB-α subunits in the cytoplasm and subsequent translocation of NF-κB proteins into the nucleus, which results in trans-activation of certain genes (17, 20). Many of the signals known to activate NF-κB result in phosphorylation and subsequent degradation of IκB. The kinases responsible for phosphorylation of IκB have recently been described (21, 22).
Although HTLV-I has been known to be etiologically related to ATL, the molecular mechanism of HTLV-I action in transforming the T cells is not completely understood. The overexpression of IL-2 and IL-2Rα, which may be mediated by Tax, seems to be a critical event in polyclonal proliferation of T cells in the early stages of the disease. However, IL-2 expression becomes undetectable during the late stage of ATL, which argues against an IL-2 autocrine loop as the cause of clonal T cell proliferation at this stage (23, 24). It is still not clear which factor(s) is required for the HTLV-I-associated T cell activation in the late stage of ATL when there is no IL-2 production.
In this study, we demonstrate that IL-15 mRNA expression is elevated in the ex vivo leukemic cells of ATL patients and in some HTLV-I-infected T cell lines. By analyzing the IL-15 promoter region, we show that the IL-15 gene is responsive to the HTLV-I Tax protein. We also provide data supporting the involvement of NF-κB in transcription of the IL-15 gene, which makes this gene susceptible to Tax regulation mediated by NF-κB. Thus, our data point to IL-15 as a potential candidate in the development or maintenance of HTLV-I associated diseases.
MATERIALS AND METHODS
Cell Culture.
All of the cells were cultured in RPMI 1640 medium containing 10% fetal calf serum and incubated at 37°C and 5% CO2/95% air. Jurkat cells were activated for 4 h by using phytohemagglutinin (PHA; 1 μg/ml) and phorbol 12-myristate 13-acetate (PMA; 20 ng/ml).
Isolation of Peripheral T Cells.
T cells from 25 ml of the peripheral blood were enriched by sheep red blood cell rosetting. Subsequently, about 108 of these cells were used to separate T cells from other cells by negative selection using antibodies conjugated to magnetic beads as described in the MACS T cell isolation (Miltenyl Biotech, Auburn, CA) protocol. The result was a population of T cells that was 99.7% pure as determined by flow cytometry analysis.
RNase Protection Assay (RPA).
Total RNA was isolated by using the PUREScript RNA isolation kit (Gentra Systems). Ten micrograms of the total RNA was used in each assay (PharMingen) using IL-15, IL-2, and GAPDH probes as described in the manufacture’s instructions.
Plasmids.
Tax cDNA was obtained by using reverse transcription-coupled PCR (Invitrogen) from MT-2 cells. After TA cloning (Invitrogen), the fragment was subcloned into the expression vector pEF.Neo (a gift from Mayumi Naramura, National Institutes of Health) at HindIII and NotI sites.
IL-15 Promoter Cloning.
We used the GenomeWalker library (CLONTECH) to clone the IL-15 5′ regulatory region. Two antisense primers, 5′-CTCCATCGAGTCCTGGCTGCAGG-3′ and 5′-CGACAGCCACCGCAGCCGGGA-3′, were used in two consecutive PCRs to amplify the upstream region of the IL-15 exon 1. The 1,170-bp amplified fragment was TA cloned into PCR2.1 plasmid (Invitrogen) and later into the alkaline phosphatase reporter plasmid (CLONTECH) at the KpnI and XhoI sites. Deletion constructs and NF-κB mutant constructs were prepared by using the Exo mung bean deletion kit and the QuickChange site-directed mutagenesis kit (Stratagene). The mutated sites were introduced into the NF-κB motif by using two complementary primers with sense primer as GCGGGGCTGG(G → T)(G → A)CTCCTCGATGTCC.
S1 Nuclease Protection Assay.
The S1 assay kit (Ambion, Austin, TX) was used to detect the transcription initiation site in the IL-15 mRNA obtained from activated monocytes.
Reporter Assay.
Ten micrograms of the reporter construct was transfected into the 10 × 106 cells by using electroporation at 220 V and 950 μF for HTLV-I-infected T cells and 280 V and 975 μF for Jurkat cells. After 48 h, the supernatant of the transfected cells was collected. The reporter activity was measured according to the manufacturer’s protocol for the Great EscAPe SEAP reporter system (CLONTECH). Five micrograms of the β-galactoside reporter construct (CLONTECH) were cotransfected in each reporter assay to normalize the transfection efficiency. The promoter activity was expressed as the fold induction of the observed reporter activity of the construct over that of the negative control, which is the reporter construct with no insert.
Electrophoretic Mobility Gel Shift Assay.
Five micrograms of cell extract were incubated with 100,000 cpm of the probe in a buffer containing 100 mM KCl, 20 mM Hepes, 0.5 mM DTT, 0.2 mM EDTA, and 20% glycerol for 30 min at 25°C. The supershift assays were done with 1 μl of the antibody against p50 and p65 (17), which was added to the binding reaction prior to addition of the probe and incubated on ice for 30 min.
RESULTS
Detection of the IL-15 mRNA in HTLV-I-Positive and -Negative T Cells.
To determine whether IL-15 mRNA is present in T cells and whether HTLV-I infection has any impact on its expression, we compared the levels of the IL-15 mRNA in normal and HTLV-I-infected T cells. We performed RPA with total RNA obtained from HTLV-I-negative T cell lines [such as Jurkat, CEM, Kit225-K6 (IL-2 dependent), Kit225-IG3 (IL-2 independent), 8402T, A301 (CD4+), and HuT78] and HTLV-I-infected T cell lines (such as HuT102, MT-1, MT-2, and MJ). We also examined peripheral blood (PB) T cells obtained from four normal individuals and seven patients with ATL for their expression of IL-15 transcripts.
As shown in Fig. 1A, IL-15 mRNA expression was higher in all the T cells from patients with ATL when compared with that of T cells from normal individuals (one normal individual is shown). For T cell lines (Fig. 1B), IL-15 mRNA levels were comparable among seven HTLV-I-negative T cell lines with the exception of HuT78 (see Discussion). The IL-15 transcript expression was elevated in HuT102, MT-2, and MJ when compared with the HTLV-I-negative T cell lines. HuT102 is a special case because the R fragment of the HTLV-I long terminal repeat region is fused to the 5′ region of the IL-15 mRNA forming a chimeric transcript (11, 25) that is associated with the overexpression of the IL-15 mRNA. Nevertheless, MT-2 and MJ cells express 3-fold more IL-15 mRNA than does the Jurkat cell line. However, the IL-15 mRNA level in MT-1 was comparable to that of Jurkat. Because MT-1 cells do not express detectable levels of Tax protein, we, therefore, considered the possibility that Tax may be involved in IL-15 expression.
Figure 1.

IL-15 mRNA expression in various T cell populations, as examined by RPA. (A) IL-15 RNA from T cells from PB of a normal individual before (lane 1) and after (lane 2) activation (PHA at 1 μg/ml and PMA at 20 ng/ml for 4 h). Lanes 3–9 contain RNA from the T cells of seven HTLV-I-infected patients with ATL. (B) Comparison of the IL-15 mRNA level between HTLV-I-positive (lanes 9–12) and -negative T cell lines (lanes 1–8). Lanes 1 and 2 contain the Jurkat T cell line at a resting state (lane 1) and an activated state (lane 2) (PHA at 1 μg/ml and PMA at 20 ng/ml for 4 h). Lanes 3–8 include the HTLV-I-non-expressing T cell lines CEM, A301, 8402T, Kit225-K6, Kit225-IG3, and Hut78, respectively. Lanes 9–12 contain HTLV-I-expressing T cell lines in the order HuT102, MT-1, MT-2, and MJ.
To explore this possibility, we used two inducible Jurkat Tax transfectants, JPX-9, which generates wild-type Tax, and JPX-m, which produces a nonfunctional Tax mutant after addition of ZnCl2 (26). As shown in Fig. 2, the IL-15 mRNA expression was increased about 4-fold upon induction of Tax in JPX-9 but not in the mutant JPX-m. To confirm that IL-15 mRNA is induced by Tax using a different approach, we transiently expressed the Tax protein in the Jurkat T antigen (27) cell line (Fig. 2). Although the endogenous level of the IL-15 transcript in the Jurkat T antigen cell line is higher than that of parental Jurkat, transfection of the Tax construct was associated with a 3-fold increase in IL-15 mRNA expression. These data suggest that the Tax protein is involved in the elevation of the IL-15 transcript levels observed in HTLV-I-infected T cells.
Figure 2.
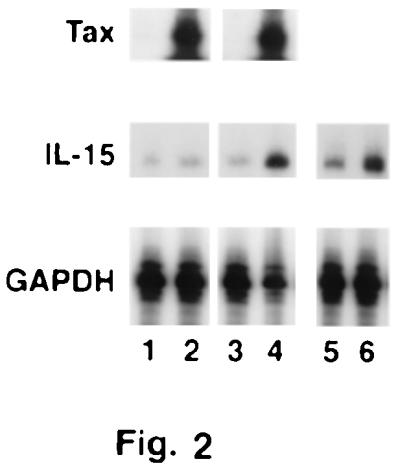
Effect of Tax expression on IL-15 mRNA expression. Lanes 1 and 2 are JPX-m (nonfunctional mutant) before and 24 h after induction of the Tax protein by ZnCl2, respectively. Lanes 3 and 4 are JPX-9 before and 24 h after induction of the Tax protein by ZnCl2, respectively. Lanes 5 and 6 are the Jurkat T antigen T cell line before and 48 h after transient transfection with the Tax/pEF.Neo construct, respectively.
Cloning of the Human IL-15 Gene Promoter.
To study the mechanisms underlying the constitutive expression of IL-15 mRNA in T cells and the effect of the HTLV-I on it, we cloned a 1,170-bp fragment of the 5′ flanking region of the IL-15 gene. The sequence of this fragment is shown in Fig. 3A. Schematic representations of the conserved regions of the mouse and human IL-15 promoters are shown in Fig. 3B. The transcription initiation site was determined by S1 nuclease mapping and RNA obtained from lipopolysaccharide (LPS)- and interferon-γ-activated monocytes (11) (Fig. 3C). As judged by the sequence, there is no conventional TATA box upstream of the transcription initiation site.
Figure 3.

(A) Sequence of the human IL-15 5′ upstream region. The transcription initiation site and the NF-κB motif are in boldface type and underlined. Part of IL-15 exon 1, which is cloned in h15/sAP, is also underlined. (B) Schematic demonstration of some of the conserved motifs between mouse and human IL-15 promoters. (C) S1 nuclease protection assay to determine the transcription initiation site (+1 position). Total RNA from activated monocytes (by LPS and interferon γ) was used for the S1 nuclease assay, which was electrophoresed alongside the sequence ladder of the cloned IL-15 5′ upstream region to determine the +1 position.
To evaluate the promoter activity of the cloned 5′ regulatory region of the IL-15 gene, we subcloned this fragment into the secreted alkaline phosphatase reporter plasmid, designated h15/sAP (27). This reporter construct was then transiently transfected into T cell lines to determine its promoter activity. The promoter activity was represented as the fold induction compared with that of the negative control. The reporter activity of the h15/sAP construct was 6-fold higher in HuT102, 4-fold higher in MT-1, and about 10-fold higher in MT-2 and MJ when compared with the negative control (Fig. 4A). In contrast, when the same construct was transfected into the control resting and activated Jurkat cells [activated by PMA and PHA (28)], less than a 2-fold increase in reporter activity was observed (Fig. 4A). We next measured the reporter activity of the h15/sAP construct in Jurkat Tax transfectants, JPX-9 and JPX-m. The h15/sAP construct was transfected into JPX-9 and JPX-m cells and Tax protein was induced by addition of ZnCl2. As shown in Fig. 4B, the reporter activity of the h15/sAP construct increased as much as 4-fold when wild-type Tax protein was induced in JPX-9. These data suggest that the HTLV-I Tax protein trans-activates the IL-15 gene promoter.
Figure 4.

Reporter assays of the IL-15 promoter construct h15/sAP in HTLV-I-infected T cell lines and Jurkat (resting and activated) cells, as well as two Jurkat Tax transfectants, JPX-m and JPX-9. Ten micrograms of the h15/sAP construct were cotransfected with 5 μg of the β-galactoside plasmid used to normalize the transformation efficiency. The average of the results of three reporter assays was used to generate each graph.
NF-κB Is Involved in Transcription of the IL-15 Gene.
To determine which element of the IL-15 promoter is responsible for its constitutive transcription and for its responses to the HTLV-I Tax protein, we made serial deletions at the 5′ end of the h15/sAP construct. These constructs were transfected into the Jurkat cells (Fig. 5A), MJ as a representative of the HTLV-I-positive cell lines (Fig. 5B), JPX-9, and JPX-m cell lines (Fig. 5C). The D2(−587), D3(−417), D4(−320), and D5(−201) deletion constructs manifested reduced promoter activity when compared with wild type, with the final value of less than 2-fold induction over the negative control. However, deletion of the 5′ region of the cloned IL-15 promoter in the D7(−141) construct increased the reporter activity about 3-fold more than the full-length promoter clone in Jurkat, MJ, and JPX-9 treated with ZnCl2 (Fig. 5 A–C). This observation suggests that there is a factor(s) that may bind to a motif in the 5′ region of nucleotide −141 of the cloned IL-15 promoter that down-regulates its transcription. Deletion of this region relieves the inhibitory effect on the IL-15 gene transcription and results in increased reporter activity. Removal of the region between nucleotides −141 and −15 in D7(−141) and D8(−15) constructs reduced the promoter activity to less than the h15/sAP level (Fig. 5 A–C), indicating that this region of the IL-15 promoter is required for its basal transcriptional activity. Because we identified an NF-κB motif at the position of the −75 and −65 nucleotides, we decided to evaluate the contribution of the NF-κB family of transcription factors to IL-15 transcription. We introduced mutations in the NF-κB site in the h15/sAP construct and transfected it into the cell lines examined previously. As shown in Fig. 5 A–C, mutation in NF-κB site abrogates IL-15 promoter activity, which suggests the involvement of NF-κB proteins in the transcription of the IL-15 gene.
Figure 5.

Reporter assay of the IL-15 promoter deletion constructs in different cell lines. (A) Jurkat resting and activated (PHA/PMA). (B) MJ an HTLV-I-infected T cell line. (C) JPX-9 and JPX-m cells 24 h after addition of ZnCl2. (D) Schematic representation of each of the deletion constructs generated in original cloned IL-15 5′ upstream region (h15/sAP). The reporter assays were performed as described in Fig. 4.
NF-κB Binds Specifically to Its Motif in the IL-15 Promoter.
To determine whether the NF-κB proteins and its motif in the IL-15 promoter are functionally related, we performed electrophoretic mobility shift assays (29) comparing the IL-15 NF-κB site and consensus NF-κB (cNF-κB) of the Ig κ light chain gene. Extracts from COS cells transfected with p50 and p65 NF-κB subunits were used to determine whether the IL-15 NF-κB motif can bind NF-κB proteins. As shown in Fig. 6A, the IL-15 NF-κB site binds to a protein complex that migrates at approximately the same level as the p50/p50 complex of the cNF-κB. This protein complex was supershifted on addition of an anti-p50 antibody. There may also be a p65-containing complex because addition of an anti-p65 antibody supershifted a protein, although it did not migrate to the same position as the cNF-κB p65 supershift. The nature of this shifted band is undefined.
Figure 6.

Electrophoretic mobility shift assay using the IL-15 NF-κB motif GGGCTGGGGCTCCTCGATGTC and Ig κB NF-κB as consensus NF-κB (cNF-κB) AGTTGAGGGGACTTTCCCAGGC (where the underlined sequences are NF-kB binding site). (A) Extracts obtained from COS cells transfected with p65 and p50 expression constructs were used with cNF-κB (lanes 1–3) or IL-15 NF-κB (lanes 4–6). The typical p50–p65 heterodimer and p50–p50 homodimer can be readily seen with cNF-κB (lane 1). The IL-15 NF-κB forms a complex that comigrates with the p50–p50 homodimer of the cNF-κB as shown by an arrow (lane 4). Supershifts of this complex gave rise to two bands almost at the same height, as indicated by an arrow (lanes 5 and 6). (B) Five micrograms of the cellular extract from resting Jurkat (lanes 1 and 8), PHA/PMA-activated Jurkat (lanes 2–4 and 9–13), or ZnCl2-treated (Tax expressing) JPX-9 (lanes 5–7 and 14–18) were used in each reaction. The formation of the p50–p65 heterodimer and p50–p50 homodimer can be seen with cNF-κB and activated or Tax-expressing Jurkat cells. The p50 and p65 supershifts are marked for the cNF-κB probe. By using the IL-15 NF-κB probe, two species can be detected, as shown by the arrows. The specificity of the binding of these complexes was examined by a competition assay using a 10 and 100 times excess of the unlabeled IL-15 NF-κB probe added to the activated or Tax-expressing Jurkat extract (lanes 10, 11, 15, and 16). The p50 and p65 supershifts of these complexes resulted in the two bands that migrated almost at the same level as indicated by an arrow (lanes 12–13 and 17–18).
To examine the NF-κB complexes formed in PHA/PMA-activated or Tax-expressing Jurkat T cells, we performed the electrophoretic mobility shift assay with extracts obtained from these cells (Fig. 6B). Two bands representing a p50–p65 heterodimer and a p50–p50 homodimer can be recognized in Jurkat extracts with the cNF-κB probe. However, the DNA–protein complexes formed with Jurkat extracts and the IL-15 NF-κB probe migrated faster than p50–p65 heterodimer in cNF-κB. One complex migrated at the same level as the p50–p50 homodimer of cNF-κB and the other complex migrated faster than that as marked in Fig. 6B. Both represent specific binding because the addition of the unlabeled probe competed for the binding sites. To characterize the nature of this DNA–protein complex, we performed supershift assays using antibodies against p50 and p65 proteins. As shown in Fig. 6B, when IL-15 NF-κB and cNF-κB probes were used, a super-shift was detected with anti-p50 or anti-p65 antibody in both PHA/PMA-treated Jurkat and Tax-induced JPX-9. However, the anti-p65 supershift of the IL-15 NF-κB site migrates faster than that of cNF-κB. Thus, the data suggest that the IL-15 NF-κB motif can bind NF-κB proteins including the p50 protein, whereas its binding to p65 containing complexes is less clear.
DISCUSSION
Although it has been reported that when assessed by Northern blot analysis, T cells do not express IL-15 mRNA (3), by using an RPA, we demonstrated that T cells obtained ex vivo from normal and ATL patient PB and all the T cell lines examined in this study express IL-15 mRNA. Because the basal level of T cell IL-15 transcript is low, the Northern blot analysis of the total RNA used in previous reports was not sensitive enough to detect IL-15 mRNA expressed by T cells. However, with RPA, we were able to detect the low levels of IL-15 mRNA expressed in these cells and to distinguish 3- to 4-fold increases in the levels of IL-15 mRNA expression by HTLV-I-positive T cell lines as compared with that of HTLV-I-negative T cell lines. In the rare situations where no IL-15 mRNA was detected by RPA, we were able to detect such transcripts by performing reverse transcription-coupled PCR. Thus, on the basis of this and previous studies, it appears that IL-15 transcripts have a virtually universal distribution.
The levels of IL-15 mRNA in all the HTLV-I-negative T cell lines, with the exception of HuT78, were comparable to those of normal PB T cells. The exception to this observation, HuT78, is a T lymphoma cell line that has a rearranged NF-κB2 gene that results in abnormal NF-κB2 proteins (p100/p52) with no transcriptional repressor functions (30), which may contribute to the overexpression of the IL-15 transcript observed with this cell line.
When the IL-15 mRNA level expressed by T cells examined in this study are compared, it is apparent that the ex vivo leukemic cells of randomly selected ATL patients express higher levels of the IL-15 mRNA than normal PB T cells and HTLV-I-negative T cell lines. Because we have presented evidence that Tax can induce IL-15 transcription in Jurkat cells, it is reasonable to suggest that the Tax protein expressed by MT-2 and MJ is responsible for the higher IL-15 mRNA expression. However, it is intriguing to note that IL-15 mRNA expression is higher in the PB T cells of ATL patients when compared with that of MT-2 and MJ, considering the fact that T cells from PB obtained from ATL patients do not express levels of Tax mRNA that can be detected by RPA. Nonetheless, we could detect the Tax transcript at a minimal level with reverse transcription-coupled PCR (unpublished data). A more probable explanation for the augmented IL-15 mRNA expression of ATL cells is suggested by the studies by Kitajima et al. (31) in which they demonstrated that in mice at the late stage of ATL, Tax antisense oligonucleotides had no apparent effect on the cell growth of the HTLV-I-positive cells, whereas NF-κB antisense oligonucleotides inhibited the cell growth. These observation suggest that in the late stages of ATL, NF-κB activation is necessary for the maintenance of the ATL phenotype. It appears that at the late stage of ATL when there is only minimal Tax expression, the NF-κB pathway remains active, which may account for the persistent expression of certain genes such as those encoding IL-15 and IL-2Rα.
Our data suggest that the NF-κB pathway may be involved in the constitutive production of IL-15 mRNA in HTLV-I-infected T cells and the Tax-expressing Jurkat cell line. As we showed by our reporter assays, the introduction of a mutation in the NF-κB motif site or deletion of this region in the IL-15 promoter constructs reduced the reporter activity dramatically in Jurkat and HTLV-I-infected T cells. We also demonstrated that the IL-15 NF-κB motif can bind NF-κB proteins. Although binding a complex containing the p50 protein is more evident, further studies are required to investigate whether any other NF-κB protein can bind as well.
Although the NF-κB pathway is activated by variety of stimuli (19), it often results in distinct transcriptional activity of certain genes. For instance, Tax and PMA/PHA treatment of T cells activation the NF-κB pathway but only Tax causes the induction of IL-15 mRNA. This may be due to the involvement of Tax at different levels of IL-15 transcription. Activation of monocytes by LPS and interferon γ also results in IL-15 mRNA induction (11). Although LPS is known to use NF-κB as one of its transcriptional mediators, induction of the IL-15 transcripts in monocytes may be due to the cooperation of this factor with other transcriptional elements, such as interferon regulatory factor 1, which is known to be activated by interferons (32) and whose motif was found to be present in the IL-15 promoter region. These considerations suggest that activation of the NF-κB pathway alone is not sufficient for the induction of the IL-15 gene.
The fact that IL-15 mRNA expression is elevated in ATL leukemic cells raises a potential role for IL-15 in the pathogenesis and/or maintenance of this disease. We have demonstrated that IL-15 mRNA expression by ATL cell lines is associated with the synthesis and secretion of this cytokine (N.A., unpublished observations). This constitutive expression of IL-15 could account for some of the reported observations with ATL. For example, the activation of HTLV-I-infected T cells is sensitive to rapamycin, a drug that inhibits the IL-2Rγc pathway shared with IL-15 (33). Moreover, the Janus family tyrosine kinases (JAK) 1 and 3, which are associated, respectively, with IL-2Rβ and -γc subunits used by IL-2 and IL-15 (34–36), were constitutively phosphorylated in HTLV-I-infected T cell lines and leukemic cells of selected ATL patients (37–39). Downstream signal transducers and transcription activators STAT-3 and 5 were also constitutively activated (39). These observations suggest the engagement of the IL-2/15R pathway in HTLV-I-infected T cells, even in the absence of IL-2. In this regard, as noted, IL-15 uses the IL-2Rβ and -γc chains of the IL-2R (40, 41) and the same JAK and STAT pathways used by IL-2 (36, 42). Thus, it is possible that an IL-15 autocrine loop may account for this JAK/STAT activation and may even contribute to HTLV-I-induced T cell proliferation in ATL.
The present study brings up a potential role for IL-15 in another HTLV-I-associated disease, HTLV-I-associated myelopathy/tropical spastic paraparesis (HAM/TSP). The HTLV-I-infected T cells of patients with HAM/TSP express Tax and manifest abnormal spontaneous proliferation when studied ex vivo (43). This proliferation can be partially inhibited by the addition of antibodies to IL-2 or to the private IL-2R IL-2Rα, suggesting a role for IL-2 in this proliferation (44). However, the addition of Mikβ1, an antibody to the IL-2R/IL-15Rβ chain, also reduces this proliferation (44). This antibody does not inhibit the action of IL-2 on the high-affinity IL-2R but inhibits IL-15 function, an observation that suggests a role for IL-15 in the abnormal T cell proliferation observed in HAM/TSP. Understanding the role of IL-15 in HTLV-I-associated diseases may assist in the development of rational therapeutic approaches for these disorders.
ABBREVIATIONS
- IL
interleukin
- HTLV-I
human T cell lymphotropic virus type I
- ATL
adult T cell leukemia
- HAM/TSP
HTLV-I-associated myopathy/tropical spastic paraparesis
- PB
peripheral blood
- RPA
RNase protection assay
- IL-2R
IL-2 receptor
- PHA
phytohemagglutinin
- PMA
phorbol 12-myristate 13-acetate
- LPS
lipopolysaccharide
Footnotes
References
- 1.Burton J D, Bamford R N, Peters C, Grant A J, Kurys G, Goldman C K, Brennan J, Roessler E, Waldmann T A. Proc Natl Acad Sci USA. 1994;91:4935–4939. doi: 10.1073/pnas.91.11.4935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Armitage R J, Macduff B M, Eisenman J, Paxton R, Grabstein K H. J Immunol. 1995;154:483–490. [PubMed] [Google Scholar]
- 3.Grabstein K H, Eisenman J, Shanebeck K, Rauch C, Srinivasan S, Fung V, Beers C, Richardson J, Schoenborn M A, Giri J. Science. 1994;264:965–968. doi: 10.1126/science.8178155. [DOI] [PubMed] [Google Scholar]
- 4.Leclercq G, Debacker V, De Smedt M, Plum J. J Exp Med. 1996;184:325–336. doi: 10.1084/jem.184.2.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ogasawara, K., Hida, S., Azimi, N., Tagaya, Y., Sato, T., Yokochi-Fukuda, T., Waldmann, T. A., Taniguchi, T. A. & Taki, S. (1998) Nature (London), in press. [DOI] [PubMed]
- 6.Mrozek E, Anderson P, Caligiuri M A. Blood. 1996;87:2632–2640. [PubMed] [Google Scholar]
- 7.Quinn L S, Haugk K L, Grabstein K H. Endocrinology. 1995;136:3669–3672. doi: 10.1210/endo.136.8.7628408. [DOI] [PubMed] [Google Scholar]
- 8.Reinecker H, MacDermott R P, Mirau S, Dignass A, Podolsky D K. Gastroenterology. 1996;111:1706–1713. doi: 10.1016/s0016-5085(96)70036-7. [DOI] [PubMed] [Google Scholar]
- 9.Lee Y B, Satoh J, Walker D G, Kim S U. Neuroreport. 1996;7:1062–1066. doi: 10.1097/00001756-199604100-00022. [DOI] [PubMed] [Google Scholar]
- 10.Doherty T M, Seder R A, Sher A. J Immunol. 1996;156:735–741. [PubMed] [Google Scholar]
- 11.Bamford R N, Battiata A, Burton J D, Sharma H, Waldmann T A. Proc Natl Acad Sci USA. 1996;93:2897–2902. doi: 10.1073/pnas.93.7.2897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Poiesz B J, Ruscetti F W, Gazdar A F, Bunn P A, Minna J D, Gallo R C. Proc Natl Acad Sci USA. 1980;77:7415–7419. doi: 10.1073/pnas.77.12.7415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grassmann R, Dengler C, Muller-Fleckenstein I, Fleckenstein B, McGuire K, Dokhelar M C, Sodroski J G, Haseltine W A. Proc Natl Acad Sci USA. 1989;86:3351–3355. doi: 10.1073/pnas.86.9.3351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jeang K-T, Boros I, Brady J, Randonovich M, Khory G. J Virol. 1988;62:4499–4509. doi: 10.1128/jvi.62.12.4499-4509.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Inoue J, Seiki M, Taniguchi T, Tsuru S, Yoshida M. EMBO J. 1986;5:2883–2888. doi: 10.1002/j.1460-2075.1986.tb04583.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fujii M, Sassone-Corsi P, Verma I M. Proc Natl Acad Sci USA. 1988;85:8526–8530. doi: 10.1073/pnas.85.22.8526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kanno T, Brown K, Franzoso G, Siebenlist U. Mol Cell Biol. 1994;14:6443–6451. doi: 10.1128/mcb.14.10.6443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Grimm S, Bauerele P A. Biochem J. 1993;290:297–308. doi: 10.1042/bj2900297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Siebenlist U, Franzoso G, Brown K. Annu Rev Cell Biol. 1994;10:405–455. doi: 10.1146/annurev.cb.10.110194.002201. [DOI] [PubMed] [Google Scholar]
- 20.Grilli M, Chiu J, Lenardo M J. Int Rev Cytol. 1993;143:1–62. doi: 10.1016/s0074-7696(08)61873-2. [DOI] [PubMed] [Google Scholar]
- 21.Stancovski I, Baltimore D. Cell. 1997;91:299–302. doi: 10.1016/s0092-8674(00)80413-4. [DOI] [PubMed] [Google Scholar]
- 22.Mercurio F, Zhu H, Murray B W, Shevchenko A, Bennett B, Li J W, Young D, Barbosa M, Mann M, Manning A, Rao A. Science. 1997;281:860–866. doi: 10.1126/science.278.5339.860. [DOI] [PubMed] [Google Scholar]
- 23.Maeda M, Arima N, Daitoku Y, Kashihara M, Okamoto H, Uchiyama T, Shrono K, Matsuoka M, Hattori T, Yodi J. Blood. 1987;70:1407–1411. [PubMed] [Google Scholar]
- 24.Tendler C L, Greenberg S J, Burton J D, Danielpour D, Kim S J, Blattner W A, Manns A, Waldmann T A. J Cell Biochem. 1991;46:302–311. doi: 10.1002/jcb.240460405. [DOI] [PubMed] [Google Scholar]
- 25.Tagaya Y, Bamford R N, Defillipis A P, Waldmann T A. Immunity. 1996;4:329–336. doi: 10.1016/s1074-7613(00)80246-0. [DOI] [PubMed] [Google Scholar]
- 26.Ohtani K, Nakamura M, Saito S, Nagata K, Sugamura K, Hinuma Y. Nucleic Acids Res. 1989;17:1589–1604. doi: 10.1093/nar/17.4.1589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Northrop J P, Ullman K S, Crabtree G R. J Biol Chem. 1993;268:2917–2923. [PubMed] [Google Scholar]
- 28.Baldwin A S, Sharp P A. Mol Cell Biol. 1987;7:305–313. doi: 10.1128/mcb.7.1.305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fried M, Crothers D M. Nucleic Acids Res. 1981;9:6505–6525. doi: 10.1093/nar/9.23.6505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhang J, Chang C C, Lombardi L, Dalla-Favera R. Oncogene. 1994;9:1931–1937. [PubMed] [Google Scholar]
- 31.Kitajima I, Shinohara T, Bilakovics J, Brown D A, Xu X, Nerenberg M. Science. 1992;258:1792–1795. doi: 10.1126/science.1299224. [DOI] [PubMed] [Google Scholar]
- 32.Tanaka N, Taniguchi T. Adv Immunol. 1992;52:263–280. doi: 10.1016/s0065-2776(08)60877-9. [DOI] [PubMed] [Google Scholar]
- 33.Hollsberg P, Wucherpfenning K, Ausubel L, Calvo V, Bierer B, Hafler D. J Immunol. 1992;148:3256–3263. [PubMed] [Google Scholar]
- 34.Whitthuhn B A, Silvennoinen O, Mirua O, Lai K S, Cwick C, Liu E T, Ihle J N. Nature (London) 1994;370:153–157. doi: 10.1038/370153a0. [DOI] [PubMed] [Google Scholar]
- 35.Johnston J A, Kawamura M, Kirken R A, Chen Y-Q, Blake R A, Shibuya K, Ortaldo J R, McVicar D W, O’Shea J J. Nature (London) 1994;370:151–153. doi: 10.1038/370151a0. [DOI] [PubMed] [Google Scholar]
- 36.Johnston J A, Bacon C M, Finbloom D S, Rees R C, Kaplan D, Shibuya K, Ortaldo J R, Gupta S, Chen Y Q, Giri J D, O’Shea J J. Proc Natl Acad Sci USA. 1995;92:8705–8709. doi: 10.1073/pnas.92.19.8705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Migone T, Lin J, Cereseto A, Mulloy J C, O’Shea J, Franchini G, Leonard W J. Science. 1995;269:79–81. doi: 10.1126/science.7604283. [DOI] [PubMed] [Google Scholar]
- 38.Xu X, Kang S-H, Heidenreich O, Okerholm M, O’Shea J J, Nerenberg M I. J Clin Invest. 1995;3:1548–1555. doi: 10.1172/JCI118193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Takemoto S, Mulloy J C, Cereseto A, Migone T, Patel B K R, Matsuoka M, Yamaguchi K, Takatsuki K, Kamihira S, White J D, et al. Proc Natl Acad Sci USA. 1997;94:13897–13902. doi: 10.1073/pnas.94.25.13897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Giri J G, Ahdieh M, Eisenman J, Shanebeck K, Grabstein K, Kumaki S, Namen A, Park L S, Cosman D, Anderson D. EMBO J. 1994;13:2822–2830. doi: 10.1002/j.1460-2075.1994.tb06576.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bamford R N, Grant A J, Burton J D, Peters C, Kurys G, Goldman C K, Brennan J, Roessler E, Waldmann T A. Proc Natl Acad Sci USA. 1994;91:4940–4944. doi: 10.1073/pnas.91.11.4940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lin J X, Migone T S, Tsang M, Friedmann M, Weatherbee J A, Zhau L, Yamauchi A, Bloom E T, Mietz J, Leonard W J. Immunity. 1995;2:331–339. doi: 10.1016/1074-7613(95)90141-8. [DOI] [PubMed] [Google Scholar]
- 43.Jacobson, S., Zaminovic, V., Mora, C., Rodgers-Johnson, P., Sheremata, W. A., Gibbs, C. J., Gajdusek, C. & McFarlin, D. E. (1988) Ann. Neurol. 23, Suppl., S196–S200. [DOI] [PubMed]
- 44.Tendler C L, Greenberg S J, Blattner W A, Manns A, Murphy E, Fleisher T, Hanchard B, Morgan O, Burton J D, Waldmann T A. Proc Natl Acad Sci USA. 1990;87:5218–5222. doi: 10.1073/pnas.87.13.5218. [DOI] [PMC free article] [PubMed] [Google Scholar]