

Abstract
Anakinra, the recombinant form of IL-1 receptor antagonist (IL-1Ra), has been approved for clinical use in the treatment of rheumatoid arthritis as the drug Kineret™, but it must be administered daily by subcutaneous injection. Gene transfer may offer a more effective means of delivery. In this study, using prostaglandin E2 production as a measure of stimulation, we quantitatively compared the ability of anakinra, as well as that of IL-1Ra delivered by gene transfer, to inhibit the biologic actions of IL-1β. Human synovial fibroblast cultures were incubated with a range of doses of anakinra or HIG-82 cells genetically modified to constitutively express IL-1Ra. The cultures were then challenged with recombinant human IL-1β either simultaneously with addition of the source of IL-1Ra or 24 hours later. In a similar manner, the potencies of the two sources of IL-1Ra were compared when human synovial fibroblasts were challenged with IL-1β produced constitutively by genetically modified cells. No significant difference in inhibitory activity was observed between recombinant protein and IL-1Ra provided by the genetically modified cells, under static culture conditions, even following incubation for 4 days. However, under culture conditions that provided progressive dilution of the culture media, striking differences between these methods of protein delivery became readily apparent. Constitutive synthesis of IL-1Ra by the genetically modified cells provided sustained or increased protection from IL-1 stimulation over time, whereas the recombinant protein became progressively less effective. This was particularly evident under conditions of continuous IL-1β synthesis.
Keywords: arthritis, gene therapy, IL-1, IL-1 receptor antagonist, synoviocytes
Introduction
IL-1 has been implicated as a pathogenic mediator in numerous inflammatory and degenerative conditions, including rheumatoid arthritis (RA) and osteoarthritis (OA) [1]. The IL-1 receptor antagonist (IL-1Ra), a naturally occurring inhibitor of the biologic actions of IL-1, has obvious therapeutic potential in such diseases [2]; indeed recombinant human IL-1Ra (anakinra) has recently been approved for use in patients with RA as the drug Kineret™ (Amgen, Inc., Thousand Oaks, CA, USA).
Limitations of IL-1Ra as a pharmaceutical include its lack of oral availability and its short biologic half-life. This is why in clinical application Kineret™ must be administered by daily subcutaneous injection. Even then, it remains unlikely that a therapeutic concentration of IL-1Ra will be maintained between injections [3]; IL-1Ra is rapidly eliminated in the kidney, resulting in a serum half-life of 4–6 hours following intravenous injection into healthy, human volunteers. This problem is exacerbated by the pronounced spare receptor effect of IL-1. According to the literature [4-6] it is necessary to maintain an IL-1Ra : IL-1 molar ratio of 10–100 or more to achieve a strong inhibitory effect.
We have proposed IL-1Ra gene transfer as a means of overcoming these problems [7]. The advantages of IL-1Ra gene delivery include its ability to engender the continuous production of therapeutic concentrations of IL-1Ra at defined anatomic locations for extended periods of time – potentially for life. Moreover, it is theoretically possible to regulate levels of IL-1Ra gene expression in a manner commensurate with disease activity [8]. IL-1Ra gene therapy has been evaluated in a number of different animal models of RA and OA, with extremely promising results [9-18]. Indeed, a phase I human study of IL-1Ra gene therapy in RA [19] was recently successfully completed.
During the preclinical development of IL-1Ra gene therapy, we often noticed that transfer of the IL-1Ra gene provided a far greater biologic effect than administration of the recombinant protein. An example is provided by the treatment of antigen-induced arthritis in rabbits. Lewthwaite and coworkers [20] reported that repeated injection of recombinant human IL-1Ra had no effect in this model of RA beyond inhibition of the synovial fibrosis occurring in the chronic stage of the disease. Otani and colleagues [16], in contrast, observed a dramatic beneficial effect on cartilage matrix metabolism, and a moderate anti-inflammatory effect when administering IL-1Ra locally to joints via ex vivo gene transfer.
There exist several possible explanations for the improved effectiveness of IL-1Ra when delivered as a gene rather than as a recombinant protein. The most likely of these are as follows. First, gene transfer results in continuous, rather than intermittent, protein delivery, thus maintaining a constant supply of IL-1Ra at a concentration sufficient to inhibit the biologic actions of IL-1. Second, gene delivery produces a molecule that has been subjected to authentic post-translational processing. Because the recombinant molecule lacks glycosylation and has an extra amino-terminal methionine, the native molecule may have greater biologic potency than the recombinant one.
The present study was designed to compare quantitatively the relative effectiveness of these two avenues of protein delivery under controlled conditions in vitro. Cultures of primary human synovial fibroblasts (HSFs) were treated with human IL-1Ra, either administered as the recombinant protein or by co-culture with fibroblasts genetically engineered to express and secrete human IL-1Ra in a constitutive manner. Stimulation from human IL-1β was then provided by addition of recombinant IL-1β protein or by co-culture with fibroblasts genetically engineered to constitutively secrete high levels of human IL-1β [21]. Using prostaglandin E2 (PGE2) levels in conditioned media as a readout of IL-1 stimulation in the respective cultures, protection from IL-1 stimulation by each method was evaluated under static and dynamic culture conditions, the latter of which were designed to resemble more closely the circumstance of an arthritic joint in which IL-1 is chronically produced.
The data suggest that the recombinant and transgenic molecules are similarly potent. Although the gene delivery procedure may benefit marginally from increased concentration at the cellular level, the advantage of gene transfer as a means of drug delivery arises from the sustained availability of IL-1Ra that this method permits.
Materials and method
Materials
Ham's F12 medium, Dulbecco's modified Eagle medium (DMEM), fetal bovine serum (FBS), penicillin-streptomycin, type II collagenase, dispase, and Geneticin™ were supplied by Gibco-BRL (Rockville, MD, USA). Zeocin™ was obtained from Invitrogen (Carlsbad, Ca, USA). Recombinant human IL-1β and IL-1Ra were purchased from R&D Systems (Minneapolis, MN, USA). ELISA kits for PGE2 and IL-1Ra were purchased from Dynatech (Ann Arbor, MI, USA) and R&D Systems, respectively. ELISA kits for human IL-1β were purchased from Endogen (Woburn, MA, USA).
Reporter cell cultures
Human synovial tissues were recovered from joints of OA patients undergoing total joint replacement surgery. HSFs were isolated by sequential digestion of synovial fragments with 1.5% dispase for 2 hours at 37°C and 0.2% collagenase for 2 hours. After washing in phosphate-buffered saline the cells were cultured in 25 cm2 dishes in DMEM with 10% FBS and 1% penicillin-streptomycin. After the third passage, the type B synovial cells were trypsinized, counted, and cultured at a density of 5 × 105 cells per well in 24-well plates with 1 ml DMEM supplemented with 10% FBS and 1% penicillin-streptomycin.
Engineered cell lines
To generate a cell line that provided a source of constitutive production and secretion of transgenic IL-1Ra, the rabbit synovial cell line HIG-82 [22] was cultured in 25 cm2 flasks containing 4 ml Ham's F12 medium with 10% FBS and 1% penicillin-streptomycin. Cells were grown to approximately 75% confluence and incubated in the presence of 8 μg/ml polybrene with 2 ml supernatant containing amphotropic retrovirus DFG-IRAP-zeor containing the human IL-1Ra and Streptoalloteichus hindustanus bleomycin-resistance (Sh ble; zeor) genes. The latter allowed positive selection of the transduced cells in medium containing Zeocin at 0.5 mg/ml. These cells, HIG-82-IL-1Ra+, were found to secrete approximately 3 μg IL-1Ra/ml per 106 cells over 24 hours. The HIG-82 cells were chosen for this purpose because they do not produce PGE2 in response to IL-1β. Non-transduced HIG-82 cells used as negative controls were cultured in Ham's F12 medium with 10% FBS and 1% penicillin-streptomycin.
To generate cells that constitutively expressed human IL-1β, skin was first harvested from a euthanized Wistar rat (Charles River Laboratories, Wilmington, MA, USA), minced with a scalpel, and digested for 2 hours at 37°C under gentle agitation with 0.2% clostridial collagenase. Dermal cells were recovered by centrifugation of the digestion mixture at 5000 rpm in a table top centrifuge, and then cultured in 25 cm2 flasks in DMEM with 10% FBS and 1% penicillin-streptomycin. Adherent cells were transduced with an amphotropic retrovirus, DFG-hIL-1β-neo, which encodes the mature form of human IL-1β fused to the leader sequence of human parathyroid hormone to enable efficient secretion, and neomycin phosphotransferase [21]. Retroviral transductants were positively selected in complete DMEM containing Geneticin at 0.5 mg/ml. These cells were found to secrete approximately 250 ng human IL-1β/ml per 106 cells over 24 hours.
IL-1 receptor antagonist and IL-1β treatment
Human IL-1Ra or IL-1β was delivered as a recombinant protein or by expression of its cDNA from genetically modified cells. For the addition of cells, cultures of HIG-82-IL-1Ra+, or dermal fibroblasts secreting IL-1β, were trypsinized, washed in PBS, and resuspended in complete DMEM. The cells were then counted using a hemocytometer and the appropriate number suspended in 50 μl DMEM for subsequent addition to the multiwell HSF cultures.
Biological assays
PGE2, IL-1β, and IL-1Ra concentrations in conditioned media were measured using ELISA according to the manufacturers' instructions. These assays do not show any cross-reactivity with other prostanoids, or rabbit and rat forms of IL-1 or IL-1Ra. Under certain culture conditions, such as those described in Figs 2,3,4,5, approximately 1/20, or 50 μl of the media volume was removed periodically and replaced, in order to allow analysis at strategic time points without significantly altering the evolving culture conditions.
Figure 2.
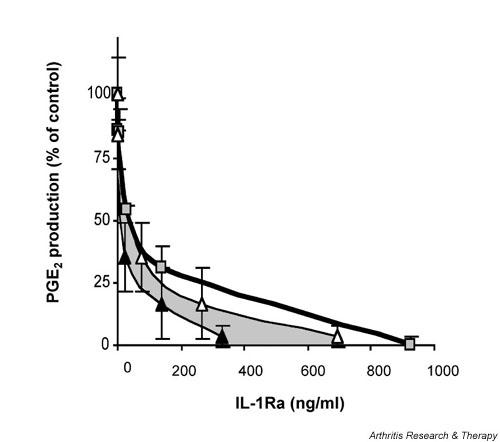
Comparison of the relative inhibitory activity of HIG-82-IL-1Ra+ cells to recombinant IL-1Ra when added to HSF cultures 24 hours before stimulation with IL-1β. As in Fig. 1, HSF were plated in multiwell plates and incubated with a range of doses of rIL-1Ra or HIG-82-IL-1Ra+ cells. Twenty-four hours following the addition of the protein or cells, 5 ng recombinant IL-1β was added, and the cultures were incubated an additional 48 hours. PGE2 production at 48 hours after IL-1stimulation for the IL-1Ra treated groups was normalized to control, namely IL-1 stimulated HSF cultures that did not receive IL-1Ra. Shown in the graph is the relationship between PGE2 production at48 hours after IL-1 stimulation, and IL-1Ra concentration immediately before and 48 hours after IL-1 stimulation. PGE2/IL-1Ra levels for cultures receiving the IL-1Ra producing cells are represented by triangles; black indicates before IL-1 stimulation and white indicates 48 hours after. The shaded boxes represent the PGE2/IL-1Ra levels for cultures receiving recombinant IL-1Ra. A single set of data points is shown for the wells receiving the recombinant protein because the IL-1Ra concentration did not change over time. The shaded region between the curves is shown to emphasize the change in IL-1Ra levels over time in the wells receiving the IL-1Ra producing cells. Experiments were performed in triplicate, and each data point repesents the mean value ± SD. HSF, human synovial fibroblast; IL-1Ra, IL-1 receptor antagonist; PGE2, prostaglandin E2.
Figure 3.
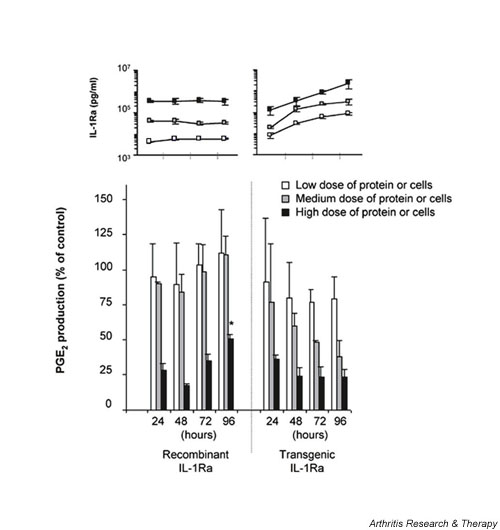
Comparison of the relative inhibitory activity of HIG-82-IL-1Ra+ cells to recombinant IL-1Ra following extended incubation. HSF cultures were incubated with one of three doses of rIL-1Ra (3, 30, or 300 ng/ml) or IL-1Ra producing cells (3 × 103, 2 × 104, or 1 × 105) that secreted corresponding levels of transgenic IL-1Ra protein within 24 hours. From the results of Fig. 2 these doses provided either low (approximately 10–15%), medium (approximately 25–50%), or high level (approximately 70–80%) inhibition of IL-1β. Twenty-four hours following the addition of the source of IL-1Ra, 5 ng recombinant IL-1β was added to each culture well. At 24 hour intervals after IL-1 stimulation, PGE2 and IL-1Ra levels in the conditioned media were measured. PGE2 levels were normalized to IL-1 stimulated HSF that were not treated with IL-1Ra, which were assigned the value of 100%. The bottom graph represents the change in PGE2 levels in the media over time from cells receiving either rIL-1Ra or the tIL-1Ra producing cells. The white bars represent cultures receiving the low dose of protein or cells, the grey bars the medium dose, and the black bars the high dose. The inset above reflects the corresponding IL-1Ra concentrations in the conditioned media for each time point and dose. Experiments were performed in triplicate, and each data point repesents the mean value ± SD. *P < 0.05 versus corresponding IL-1Ra source at 24 hours. HSF, human synovial fibroblast; IL-1Ra, IL-1 receptor antagonist; PGE2, prostaglandin E2.
Figure 4.
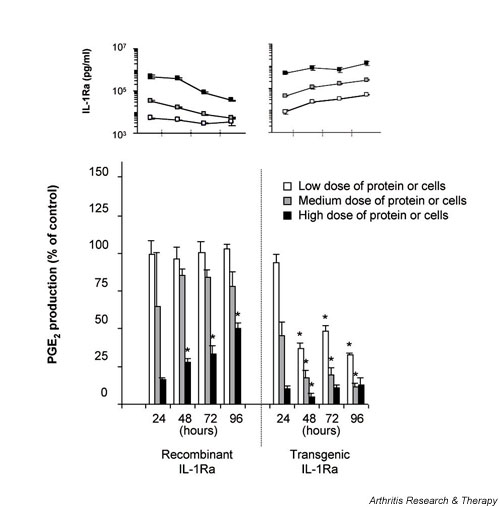
Comparison of the relative inhibitory activity of HIG-82-IL-1Ra+ cells to recombinant IL-1Ra following periodic dilution. Experiments were performed identically to those described for Fig. 3, except that at 24 hour intervals after IL-1 stimulation one half of the conditioned media in each well was removed and replaced with fresh media. As before, the PGE2 and IL-1Ra concentrations in the recovered media were measured using ELISA. Experiments were performed in triplicate, and each data point repesents the mean value ± SD. *P < 0.05 versus corresponding IL-1Ra source at 24 hours. ELISA, enzyme-linked immunosorbent assay; IL-1Ra, IL-1 receptor antagonist; PGE2, prostaglandin E2.
Figure 5.
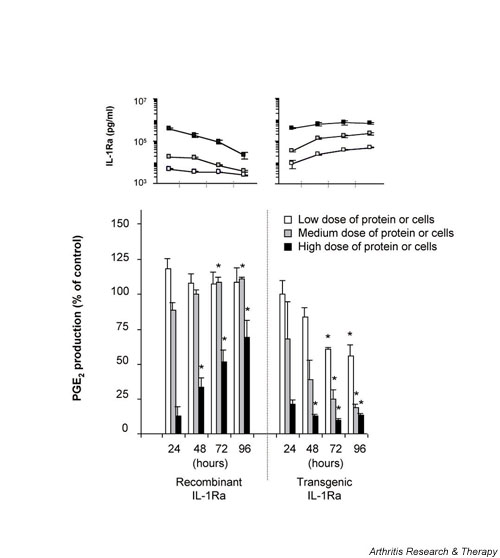
Comparison of the relative inhibitory activity of HIG-82-IL-1Ra+ cells to recombinant IL-1Ra in the presence of chronic IL-1β stimulation. Experiments were performed similar to those described for Fig 4, except that 2 × 104 rat dermal fibroblasts retrovirally transduced to constitutively express and secrete human IL-1β were added to each culture well instead of recombinant IL-1β protein. PGE2 and IL-1Ra concentrations in the conditioned media were measured at 24 hour intervals using ELISA. Experiments were performed in triplicate, and each data point repesents the mean value ± SD. *P < 0.05 versus corresponding IL-1Ra source at 24 hours. ELISA, enzyme-linked immunosorbent assay; IL-1Ra, IL-1 receptorantagonist; PGE2, prostaglandin E2.
Statistical analysis
All results are presented as means ± SD. Statistical analyses were performed using an unpaired Student's t-test, and P < 0.05 was considered statistically significant.
Results
To quantitatively compare the ability of recombinant (r)IL-1Ra and IL-1Ra provided by genetically modified cells (i.e. transgenic [t]IL-1Ra) to inhibit the effects of IL-1β, we performed a series of experiments using both static and dynamic conditions of IL-1 stimulation. Because the synovium is an important contributor to pathogenesis in arthritis, primary cultures of HSFs were used as target cells, and concentrations of PGE2 in media conditioned by the HSFs were used as a measure of IL-1β stimulation.
In our initial experiments we compared the inhibitory activity of rIL-1Ra to that of HIG-82-IL-1Ra+ cells – a cell line engineered to constitutively express human IL-1Ra – when each was added to HSF cultures simultaneously with IL-1β. For this, 5 ng IL-1β was added to 5 × 105 HSFs accompanied by either a range of doses of rIL-1Ra or increasing numbers of HIG-82-IL-1Ra+ cells. Forty-eight hours later, the conditioned media were analyzed for IL-1Ra and PGE2 concentrations. A plot of IL-1Ra concentration versus PGE2 production of the IL-1Ra treated cells, relative to PGE2 levels of control HSFs incubated with IL-1β alone, is shown in Fig. 1. Over a wide range of doses the recombinant and transgenic sources of IL-1Ra were similarly capable of blocking the effects of the added IL-1β. For each source of IL-1Ra, 50% IL-1β inhibition was extrapolated to a concentration of approximately 230 ng/ml IL-1Ra and complete inhibition at approximately 800 ng/ml. This translated to IL-1Ra : IL-1β ratios of approximately 46 : 1 and 160 : 1, respectively. In control experiments, levels of PGE2 produced by IL-1β challenge of co-culture of HSFs with nontransduced HIG-82 cells were found to be identical to those of HSFs alone (data not shown).
Figure 1.
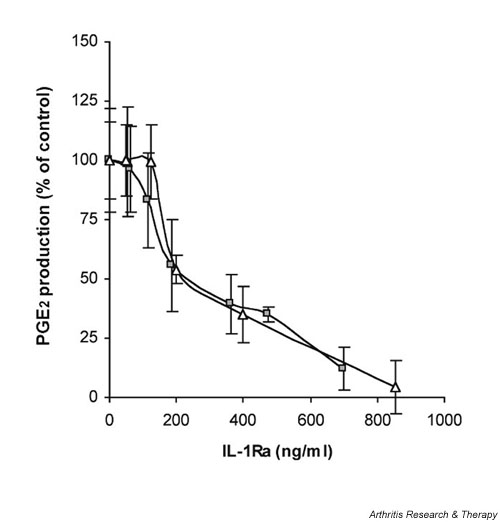
Comparison of the relative inhibitory activity of recombinant IL-1Ra to HIG-82-IL-1Ra+ cells when added to HSF cultures simultaneous to stimulation with IL-1β. Approximately 5 × 105 HSFs were plated in several wells of a 24-well plate. Twenty-four hours later, a range of doses of rIL-1Ra (from 0.13 to 1 μg) or HIG-82-IL-1Ra+ cells (ranging from 4 × 102 to 4 × 105) was added to individual wells accompanied by 5 ng recombinant IL-1β. Forty-eight hours later the conditioned media were harvested and analyzed for IL-1Ra and PGE2 content by ELISA. PGE2 levels were normalized to control, namely IL-1 stimulated HSF cultures that did not receive IL-1Ra. PGE2 levels from these controls were assigned a value of 100%. The relationship between the relative PGE2 production and IL-1Ra concentration for each treatment group is shown in the graph. The gray boxes represent the PGE2/IL-1Ra levels for the rIL-1Ra treated cultures; white triangles represent those from the cultures receiving the IL-1Ra producing cells. Experiments were performed in triplicate, and each data point represents the mean value ± SD. ELISA = enzyme-linked immunosorbent assay; HSF, human synovial fibroblast; IL-1Ra, IL-1 receptor antagonist; PGE2 prostaglandin E2.
Relative to the end-point concentrations of IL-1Ra, there was no apparent difference in the effectiveness of the two sources. By adding the recombinant protein and the modified cells at the same time as IL-1β, the concentration of rIL-1Ra would be at its maximum at the time of initial IL-1β stimulation. That of the tIL-1Ra, however, would be essentially zero, and would not reach its maximal concentration until 48 hours later, at the time of media harvest. Using this rationale, we compared the effectiveness of rIL-1Ra and tIL-1Ra under conditions in which the concentration of each would be similar at the time of IL-1β stimulation. To allow sufficient time for the IL-1Ra producing cells to adhere and begin transgenic expression, we performed experiments similar to that above, but added the IL-1β 24 hours after addition of the range of doses of rIL-1Ra or tIL-1Ra producing cells to the HSF cultures. The conditioned media were analyzed at the time of IL-1β addition and 48 hours later for PGE2 and IL-1Ra content. A plot of the IL-1Ra concentrations versus PGE2 production relative to IL-1β stimulated controls is shown in Fig. 2. To illustrate the change in IL-1Ra concentration over time in the wells receiving the IL-1Ra producing cells, we plotted the final PGE2 concentration versus the IL-1Ra concentration at the time of IL-1β stimulation and at the end of the 48 hour incubation. Because these values represent the starting and end-point IL-1Ra concentrations, the effective tIL-1Ra dose should lie somewhere between and is represented by the shaded area between the two curves.
As might be expected of a competitive inhibitor, preincubation of the HSF with IL-1Ra for 24 hours before IL-1β stimulation significantly reduced the 50% inhibition level for each source of IL-1Ra. For the recombinant protein, 50% IL-1β inhibition was extrapolated to 50 ng/ml IL-1Ra, and for the tIL-1Ra 50% inhibition fell between 10 and 50 ng/ml. For complete inhibition approximately 950 ng/ml rIL-Ra was required, whereas for the tIL-1Ra between 400 and 700 ng/ml was necessary.
The previous experiments provided evidence that the two molecules rIL-1Ra and tIL-1Ra were functionally similar and equally capable of blocking the effects of IL-1β. They also suggested that time was a factor critical to comparing the effectiveness of rIL-1Ra and IL-1Ra constitutively produced by genetically modified cells. Thus, in several additional experiments we monitored the relationship between IL-1Ra and IL-1β stimulation daily over a 96 hour interval. For these experiments, three doses of rIL-1Ra or HIG-82-IL-1Ra+ cells were used, which from Fig. 2 provided either low (approximately 10–15%), medium (approximately 25–50%), or high level (approximately 70–80%) inhibition of IL-1β. HSFs were cultured in the presence of the various doses of cells or protein, followed 24 hours later by the addition of 5 ng IL-1β. At 24 hour intervals, IL-1Ra and PGE2 were measured in the conditioned media. As shown in Fig. 3, under these static culture conditions there was little meaningful change in the levels of PGE2 production over time. At the low and medium doses rIL-1Ra had little protective effect but, relative to the 24 hour time point, a significant increase in IL-1 stimulation was seen in the wells receiving the high dose by day 4. In the wells receiving the HIG-82-IL-1Ra+ cells, protection from IL-1 stimulation was maintained over time. Although the mean levels of PGE2 were reduced over the 4 days of the experiment, this was not statistically significant. Thus, under these conditions there were no dramatic differences between a single dose of rIL-1Ra and the tIL-1Ra producing cells.
It has been shown in vivo that agents injected into the joint space can be cleared from the synovial fluid in as little as 30 min, suggesting a steady egress of solutes from the joint. Thus, to compare the effects of rIL-1Ra and tIL-1Ra producing cells under more dynamic conditions, perhaps closer to those that might be encountered in the joint in vivo, experiments were performed identically to that described for Fig 3 except that one half of the culture media was replaced every 24 hours for 4 days. Analysis of media recovered at each day showed that the medium and low doses of rIL-1Ra provided a marginal level of protection over time, and in wells receiving the highest dose initially high levels of protection were steadily lost (Fig 4). In stark contrast, the HSFs incubated with the IL-1Ra producing cells exhibited a sharp reduction in PGE2 production throughout the course of the experiment. For these groups, HSF cultures receiving even the lowest dose of HIG-82-IL-1Ra+ cells showed approximately 70% inhibition of IL-1β at 96 hours.
The previous experiment was intended to evaluate the effects of IL-1Ra under dynamic conditions following a single stimulus of IL-1β. To establish a situation of chronic IL-1β stimulation as might be encountered in an arthritic joint, experiments were designed like that described for Figure 4, but in this case the source of IL-1β was provided by the addition of dermal fibroblasts genetically modified to constitutively secrete mature human IL-1β. As shown in Fig. 5, using these conditions the single dose of rIL-1Ra was unable to block the effects of persistent IL-1β production. Even at the highest dose, the steady dilution of rIL-1Ra in the presence of constant IL-1β synthesis rapidly lost its protective effects. In this milieu, however, the potency of gene transfer as a method of drug delivery was perhaps most effectively illustrated. The maintenance of and gradual increase in IL-1Ra concentration provided by ongoing synthesis by the genetically modified cells at all doses provided sustained and increased protection from chronic synthesis of IL-1β over time.
Discussion
For the treatment of arthritis, gene therapy has the theoretical advantage over protein therapy of sustained delivery of the therapeutic product to a discrete site. Although its feasibility and effectiveness in animal models are now well established [13,14,16-19,23-25], no direct study of the intrinsic merits of using genetically modified cells as a mechanism for protein delivery has been reported. We postulated several potential benefits that could arise, including natural protein processing and intercellular presentation of the transgene product. Indeed, it is known that anakinra, the recombinant form of human IL-1Ra, differs from its naturally occurring counterpart by possessing an additional amino-terminal methionine residue and lacking glycosylation [26]. In the first part of the present study similar end-point levels of transgenic and recombinant IL-1Ra were necessary to inhibit fully the response of synoviocytes to a single challenge with IL-1β. Furthermore, and in agreement with earlier literature on this subject [4,5,27], both forms of IL-1Ra required a molar excess over IL-1 of at least two orders of magnitude to exercise an inhibition of 100%. Altogether, these data suggest that the biochemical alterations of the recombinant IL-1Ra do not significantly affect its ability to antagonize the responses of synovial fibroblasts to human IL-1β.
From the data presented in Fig. 2, however, a modest increase in the effectiveness of tIL-1Ra is suggested when one considers the relative concentrations of recombinant and transgenic IL-1Ra with time. In experiments in which IL-1β was added at the same time as the source of IL-1Ra, similar end-point concentrations of IL-1Ra were found to provide corresponding inhibitory effects, despite the fact that the recombinant protein was at its maximal concentration at the time of IL-1 stimulation whereas that from the modified cells was zero. In other experiments in which, prior to IL-1 stimulation, time was allowed for the IL-1Ra producing cells to establish concentrations equivalent to that of the recombinant, the inhibitory curve was shifted slightly to the left, indicating a slight increase in the effectiveness of IL-1Ra when provided as a transgene product. This may arise from increased concentrations of the IL-1Ra gene product in the cellular microenvironment where the protein is locally synthesized and secreted.
Interestingly, following extended culture under static conditions, largely unremarkable differences in activity were observed between the constitutively produced tIL-1Ra and rIL-1Ra. Significant differences between the two methods of protein delivery were only found under dynamic conditions in which the concentration of rIL-1Ra was reduced with time. In these situations, such as that reported in Figs 4 and 5, the importance of maintaining the local level of IL-1Ra became dramatically apparent, as were the advantages of gene transfer as a means of protein delivery. In the face of continual dilution, the constitutive production of the IL-1Ra gene product was able to maintain effective protein levels and was able to sustain and increase protection of the HSFs from IL-1 stimulation as time progressed. This was even more pronounced under conditions of continuing IL-1 production, in which the rIL-1Ra was readily overwhelmed. It should be noted that all of the experiments performed in this study used HSFs derived from OA patients. It remains possible that the amplitude of the responses to IL-1 and IL-1Ra may vary somewhat between HSFs from OA, RA, and nondiseased individuals; however, the overall result will probably remain the same.
In rodent models of RA, maximum therapeutic effects are only achieved when pumps are used to maintain a constant supply of large amounts of recombinant IL-1Ra. Under these conditions IL-1Ra has both antierosive and anti-inflammatory effects in collagen-induced arthritis. As discussed by Bendele and coworkers [3], constant serum concentrations of approximately 1 μg IL-1Ra/ml are antierosive, but it is necessary to achieve serum concentrations of approximately 5 μg IL-1Ra/ml before important anti-inflammatory effects are seen. A single, subcutaneous injection of 150 mg recombinant IL-1Ra in humans achieves a peak plasma concentration of only about 1.6 μg IL-1Ra/ml, and concentrations superior or equal to 1 μg IL-1Ra/ml exist only for about 14 hours. Local, intra-articular gene delivery of IL-1Ra could produce enough protein, with only a single injection of vector, to trigger both antierosive and anti-inflammatory local effects. This is of real interest because the clinical response to Kineret is modest and might be explained by these circumstances. Moreover, its antierosive effect is more pronounced than the anti-inflammatory one probably because of the low local concentrations.
As suggested by our results, maintaining higher in vivo concentrations of IL-1Ra in a sustained manner may be key to realizing the full therapeutic potential of this material. Gene delivery may offer the greatest chance of early success. Recent data from our laboratory have shown that the synovial lining is capable of maintaining therapeutic levels of transgene expression for at least 6 months [28], providing increased optimism for the use of gene transfer in the treatment of chronic articular disease. Indeed, IL-1Ra gene therapy has demonstrated impressive efficacy in animal models of RA and OA, and a phase I human trial has recently confirmed that the human IL-1Ra cDNA can be safely transferred to and expressed within human rheumatoid joints [19]. A planned phase II study will determine the efficacy of this procedure.
Conclusion
Recombinant human IL-1Ra and human IL-1Ra synthesized transgenically in mammalian cells are equipotent antagonists of human IL-1β. Our data indicate that the greater efficiency noted for transgenic IL-1Ra in previous animal gene therapy investigations reflects the ability of gene delivery to maintain higher in vivo concentrations of IL-1Ra in a sustained manner. This property was particularly striking under experimental conditions that resemble those found during chronic inflammatory conditions, in which IL-1β is produced continually and the concentration of rIL-1Ra administered as a single bolus progressively falls. These findings are relevant to the clinical use of Kineret and the possible future use of IL-1Ra gene therapy to treat joint diseases.
Competing interests
None declared.
Abbreviations
DMEM = Dulbecco's modified Eagle medium; ELISA = enzyme-linked immunosorbent assay; FBS = fetal bovine serum; HSF = human synovial fibroblast; (r/t)IL-1Ra = (recombinant/transgenic) IL-1 receptor antagonist; OA = osteoarthritis; PBS = phosphate-buffered saline; PGE2 = prostaglandin E2; RA = rheumatoid arthritis.
Acknowledgments
Acknowledgements
This work was supported in part by a grant from the Cambridge-MIT Institute.
References
- Dinarello CA. The interleukin-1 family [IL-1F1, F2] In: Thomson A, Lotze MT, editor. In The Cytokine Handbook. 4. Vol. 2. London, UK; pp. 643–668. [Google Scholar]
- Arend A, Evans CH. The interleukin-1 receptor antagonist [IL-1F3] In: Thomson A, Lotze MT, editor. In The Cytokine Handbook. 4. Vol. 2. London, UK; pp. 669–708. [Google Scholar]
- Bendele AM, Chlipala ES, Scherrer J, Frazier J, Sennello G, Rich WJ, Edwards CK., III Combination benefit of treatment with the cytokine inhibitors interleukin-1 receptor antagonist and PEGylated soluble tumor necrosis factor receptor type I in animal models of rheumatoid arthritis. Arthritis Rheum. 2000;43:2648–2659. doi: 10.1002/1529-0131(200012)43:12<2648::AID-ANR4>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- Arend WP, Welgus HG, Thompson RC, Eisenberg SP. Biological properties of recombinant human monocyte-derived interleukin 1 receptor antagonist. J Clin Invest. 1990;85:1694–1697. doi: 10.1172/JCI114622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith RJ, Chin JE, Sam LM, Justen JM. Biologic effects of an interleukin-1 receptor antagonist protein on interleukin-1-stimulated cartilage erosion and chondrocyte responsiveness. Arthritis Rheum. 1991;34:78–83. doi: 10.1002/art.1780340112. [DOI] [PubMed] [Google Scholar]
- Seckinger P, Klein-Nulend J, Alander C, Thompson RC, Dayer JM, Raisz LG. Natural and recombinant human IL-1 receptor antagonists block the effects of IL-1 on bone resorption and prostaglandin production. J Immunol. 1990;145:4181–4184. [PubMed] [Google Scholar]
- Evans CH, Robbins PD. The interleukin-1 receptor antagonist and its delivery by gene transfer. Receptor. 1994;4:9–15. [PubMed] [Google Scholar]
- Bakker AC, Van De Loo FA, Joosten LA, Arntz OJ, Varley AW, Munford RS, Van Den Berg WB. C3-Tat/HIV-regulated intraarticular human interleukin-1 receptor antagonist gene therapy results in efficient inhibition of collagen-induced arthritis superior to cytomegalovirus-regulated expression of the same transgene. Arthritis Rheum. 2002;46:1661–1670. doi: 10.1002/art.10481. [DOI] [PubMed] [Google Scholar]
- Frisbie DD, Ghivizzani SC, Robbins PD, Evans CH, McIlwraith CW. Treatment of experimental equine osteoarthritis by in vivo delivery of the equine interleukin-1 receptor antagonist gene. Gene Ther. 2002;9:12–20. doi: 10.1038/sj.gt.3301608. [DOI] [PubMed] [Google Scholar]
- Oligino T, Ghivizzani S, Wolfe D, Lechman E, Krisky D, Mi Z, Evans C, Robbins P, Glorioso J. Intra-articular delivery of a herpes simplex virus IL-1Ra gene vector reduces inflammation in a rabbit model of arthritis. Gene Ther. 1999;6:1713–1720. doi: 10.1038/sj.gt.3301014. [DOI] [PubMed] [Google Scholar]
- Fernandes J, Tardif G, Martel-Pelletier J, Lascau-Coman V, Dupuis M, Moldovan F, Sheppard M, Krishnan BR, Pelletier JP. In vivo transfer of interleukin-1 receptor antagonist gene in osteoarthritic rabbit knee joints: prevention of osteoarthritis progression. Am J Pathol. 1999;154:1159–1169. doi: 10.1016/S0002-9440(10)65368-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller-Ladner U, Roberts CR, Franklin BN, Gay RE, Robbins PD, Evans CH, Gay S. Human IL-1Ra gene transfer into human synovial fibroblasts is chondroprotective. J Immunol. 1997;158:3492–3498. [PubMed] [Google Scholar]
- Pelletier JP, Caron JP, Evans C, Robbins PD, Georgescu HI, Jovanovic D, Fernandes JC, Martel-Pelletier J. In vivo suppression of early experimental osteoarthritis by interleukin-1 receptor antagonist using gene therapy. Arthritis Rheum. 1997;40:1012–1019. doi: 10.1002/art.1780400604. [DOI] [PubMed] [Google Scholar]
- Bakker AC, Joosten LA, Arntz OJ, Helsen MM, Bendele AM, van de Loo FA, van den Berg WB. Prevention of murine collagen-induced arthritis in the knee and ipsilateral paw by local expression of human interleukin-1 receptor antagonist protein in the knee. Arthritis Rheum. 1997;40:893–900. doi: 10.1002/art.1780400517. [DOI] [PubMed] [Google Scholar]
- Welling TH, Davidson BL, Zelenock JA, Stanley JC, Gordon D, Roessler BJ, Messina LM. Systemic delivery of the interleukin-1 receptor antagonist protein using a new strategy of direct adenoviral-mediated gene transfer to skeletal muscle capillary endothelium in the isolated rat hindlimb. Hum Gene Ther. 1996;7:1795–1802. doi: 10.1089/hum.1996.7.15-1795. [DOI] [PubMed] [Google Scholar]
- Otani K, Nita I, Macaulay W, Georgescu HI, Robbins PD, Evans CH. Suppression of antigen-induced arthritis in rabbits by ex vivo gene therapy. J Immunol. 1996;156:3558–3562. [PubMed] [Google Scholar]
- Makarov SS, Olsen JC, Johnston WN, Anderle SK, Brown RR, Baldwin AS, Jr, Haskill JS, Schwab JH. Suppression of experimental arthritis by gene transfer of interleukin 1 receptor antagonist cDNA. Proc Natl Acad Sci USA. 1996;93:402–406. doi: 10.1073/pnas.93.1.402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandara G, Mueller GM, Galea-Lauri J, Tindal MH, Georgescu HI, Suchanek MK, Hung GL, Glorioso JC, Robbins PD, Evans CH. Intraarticular expression of biologically active interleukin 1-receptor-antagonist protein by ex vivo gene transfer. Proc Natl Acad Sci USA. 1993;90:10764–10768. doi: 10.1073/pnas.90.22.10764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans CH, Robbins PD, Ghivizzani SC, Herndon JH, Kang R, Bahnson AB, Barranger JA, Elders EM, Gay S, Tomaino MM, Wasko MC, Watkins SC, Whiteside TL, Glorioso JC, Lotze MT, Wright TM. Clinical trial to assess the safety, feasibility, and efficacy of transferring a potentially anti-arthritic cytokine gene to human joints with rheumatoid arthritis. Hum Gene Ther. 1996;7:1261–1280. doi: 10.1089/hum.1996.7.10-1261. [DOI] [PubMed] [Google Scholar]
- Lewthwaite J, Blake S, Thompson RC, Hardingham TE, Henderson B. Antifibrotic action of interleukin-1 receptor antagonist in lapine monoarticular arthritis. Ann Rheum Dis. 1995;54:591–596. doi: 10.1136/ard.54.7.591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghivizzani SC, Kang R, Georgescu HI, Lechman ER, Jaffurs D, Engle JM, Watkins SC, Tindal MH, Suchanek MK, McKenzie LR, Evans CH, Robbins PD. Constitutive intra-articular expression of human IL-1 beta following gene transfer to rabbit synovium produces all major pathologies of human rheumatoid arthritis. J Immunol. 1997;159:3604–3612. [PubMed] [Google Scholar]
- Georgescu HI, Mendelow D, Evans CH. HIG-82: an established cell line from rabbit periarticular soft tissue, which retains the 'activatable' phenotype. In Vitro Cell Dev Biol. 1988;24:1015–1022. doi: 10.1007/BF02620875. [DOI] [PubMed] [Google Scholar]
- Kang R, Marui T, Ghivizzani SC, Nita IM, Georgescu HI, Suh JK, Robbins PD, Evans CH. Ex vivo gene transfer to chondrocytes in full-thickness articular cartilage defects: a feasibility study. Osteoarthritis Cartilage. 1997;5:139–143. doi: 10.1016/s1063-4584(97)80007-6. [DOI] [PubMed] [Google Scholar]
- Baragi VM, Renkiewicz RR, Qiu L, Brammer D, Riley JM, Sigler RE, Frenkel SR, Amin A, Abramson SB, Roessler BJ. Transplantation of adenovirally transduced allogeneic chondrocytes into articular cartilage defects in vivo. Osteoarthritis Cartilage. 1997;5:275–282. doi: 10.1016/s1063-4584(97)80023-4. [DOI] [PubMed] [Google Scholar]
- Ghivizzani SC, Lechman ER, Kang R, Tio C, Kolls J, Evans CH, Robbins PD. Direct adenovirus-mediated gene transfer of interleukin 1 and tumor necrosis factor alpha soluble receptors to rabbit knees with experimental arthritis has local and distal anti-arthritic effects. Proc Natl Acad Sci USA. 1998;95:4613–4618. doi: 10.1073/pnas.95.8.4613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenberg SP, Evans RJ, Arend WP, Verderber E, Brewer MT, Hannum CH, Thompson RC. Primary structure and functional expression from complementary DNA of a human interleukin-1 receptor antagonist. Nature. 1990;343:341–346. doi: 10.1038/343341a0. [DOI] [PubMed] [Google Scholar]
- Seckinger P, Kaufmann MT, Dayer JM. An interleukin 1 inhibitor affects both cell-associated interleukin 1-induced T cell proliferation and PGE2 /collagenase production by human dermal fibroblasts and synovial cells. Immunobiology. 1990;180:316–327. doi: 10.1016/S0171-2985(11)80295-0. [DOI] [PubMed] [Google Scholar]
- Gouze E, Pawliuk R, Pilapil C, Gouze JN, Fleet C, Palmer GD, Evans CH, Leboulch P, Ghivizzani SC. In vivo gene delivery to synovium by lentiviral vectors. Mol Ther. 2002;5:397–404. doi: 10.1006/mthe.2002.0562. [DOI] [PubMed] [Google Scholar]