

Abstract
Stat proteins are SH2 domain-containing transcription factors that are activated in cells by various cytokines and growth factors. In the case of cytokines whose receptors lack protein kinase activity, phosphorylation–activation is mediated by members of the JAK family of tyrosine protein kinases. In the case of growth factors whose receptors have intrinsic tyrosine protein kinase activity, it is thought that Stat proteins can be activated either directly by the receptor or indirectly through JAK proteins. To test the possibility of direct activation, we have used purified Stat3α, Stat3β, and epidermal growth factor receptor kinase produced in recombinant baculovirus-infected Sf9 insect cells. The Stat proteins formed a stable complex with the receptor kinase, and they were phosphorylated on tyrosine by the receptor kinase and activated for binding to DNA, properties shared with Stat proteins purified from Sf9 cells coexpressing JAK1 or JAK2. Both JAK-phosphorylated Stat3β and Stat3β phosphorylated in vitro by the receptor kinase were 20–50 times more active on a molar basis for DNA binding than phosphorylated Stat3α. We conclude that Stat3 isoforms can be directly phosphorylated and thereby activated in vitro by the epidermal growth factor receptor kinase.
Keywords: signal transduction, growth factors, cytokines, JAK proteins
Polypeptide cell signaling molecules such as growth factors and cytokines influence a variety of biological events of the cell through interaction with their specific receptors at the cell surface. Ligand binding initiates a series of molecular interactions involving the cytoplasmic domain of the receptor, eventually leading to the sequential expression of target genes. Many cytokines and growth factors signal through a recently described widespread pathway, the JAK–Stat pathway, first uncovered in studies on interferon-regulated gene expression (for review, see refs. 1, 2, 3).
Stat proteins are SH2 domain-containing transcription factors found in latent form in the cytoplasm in the absence of signaling ligands. To date, six members of the Stat family have been described (Stat1 to Stat6) and in some instances long and short isoforms have been found that are encoded by alternatively spliced mRNAs (4, 5, 6, 7, 8, 9, 10, 11). JAK proteins (JAK1, JAK2, JAK3, and Tyk2) are cytoplasmic tyrosine protein kinases (12, 13). In cytokine transmembrane signaling, one or more JAK proteins associate with the cytoplasmic domain of a ligand-activated receptor, become phosphorylated, and then phosphorylate multiple tyrosine residues of the receptor. A Stat protein is thought to bind to the phosphorylated receptor–JAK complex via its SH2 domain (14, 15) and is phosphorylated by JAK on a tyrosine residue near the C terminus that is conserved in all Stat family members (Tyr-701 in Stat1 and Tyr-705 in Stat3) (16, 17). Phosphorylation of this tyrosine residue leads to homodimerization (or heterodimerization with a different Stat), movement of the activated Stat to the nucleus, binding to a specific DNA response element, and gene activation.
Unlike cytokine receptors, certain growth factor receptors, e.g., epidermal growth factor (EGF) and platelet-derived growth factor (PDGF) receptors, possess intrinsic tyrosine kinase activity that is activated by the ligand (18), raising the possibility that the receptor kinase directly phosphorylates and activates Stat proteins. Evidence in favor of direct activation of Stat proteins by receptor kinase is the finding that cell lines deficient in each of the known JAK family members still show Stat activation in response to EGF or PDGF (19, 20, 25). Moreover, activated EGF receptor kinase present in cellular immunoprecipitates prepared with anti-receptor antibody can activate Stat in vitro (21); however, the possibility that such precipitates have intermediary kinases is difficult to exclude.
In the present study, we set out to determine whether purified EGF receptor kinase produced in recombinant baculovirus-infected cells can activate in vitro the two known isoforms of Stat3. We report herein that the receptor kinase phosphorylates purified Stat3α and Stat3β on Tyr-705, thereby activating the Stat proteins for binding to DNA. In addition, we describe the formation of a receptor kinase-Stat complex and a marked quantitative difference in the DNA-binding activities of tyrosine-phosphorylated forms of Stat3α and Stat3β.
MATERIALS AND METHODS
Expression and Purification of Stat3 Proteins.
A BamHI–HindIII DNA fragment containing either murine Stat3α or Stat3β (11) was cloned into the baculovirus expression vector, pFastBac1 (GIBCO/BRL), and a 10-histidine residue tag was engineered onto the N terminus of Stat3. Baculoviruses encoding murine JAK1 and JAK2 were kindly provided by J. Ihle (St. Jude Children’s Research Hospital, Memphis, TN). For expression of proteins, Sf9 cells (2 × 106 cells per ml) were infected with recombinant virus encoding Stat3 at a multiplicity of infection of 1 and harvested at 3 days after infection. For preparation of phosphorylated Stat3, the cells were infected with Stat3- and JAK-expressing viruses. The cells (6 × 107 cells) were resuspended and lysed in 10 ml of lysis buffer [20 mM Hepes, pH 7.9/100 mM NaCl/0.5% Nonidet P-40/15% glycerol/2 mM 2-mercaptoethanol/1 mM phenylmethylsulfonyl fluoride/leupeptin (10 μg/ml)/1 mM Na3VO4/1 mM Na2MoO4/15 mM imidazole] for 1 hr. Lysates were centrifuged for 10 min at 20,000 × g to remove insoluble material, and incubated with Ni2+ ProBond resin (Invitrogen) for 1 hr. The mixture of lysate and Ni2+ resin was packed in a column and extensively washed with lysis buffer containing 30 mM imidazole. Ni2+-bound proteins were serially eluted with imidazole at the concentrations of 50, 70, 80, and 350 mM. Stat proteins were eluted at 350 mM imidazole. The purified Stat proteins were incubated with 20 mM EDTA, dialyzed against the buffer containing 10 mM Hepes, pH 7.4/100 mM NaCl/0.5 mM dithiothreitol, and stored at −70°C.
Electrophoretic Mobility-Shift Assay (EMSA).
Purified Stat proteins were incubated with 32P-labeled high-affinity c-fos sis-inducible element (hSIE; Santa Cruz Biotechnology) in the presence of 1 μg of poly(dI·dC) in 10 mM Hepes, pH 7.8/50 mM KCl/1 mM EDTA/5 mM MgCl2/10% glycerol/5 mM dithiothreitol/BSA (1 mg/ml)/leupeptin (10 μg/ml)/pepstatin A (5 μg/ml)/0.5 mM phenylmethylsulfonyl fluoride/1 mM Na3VO4. After a 15-min incubation at room temperature, the samples were electrophoresed on 4% polyacrylamide gels.
Immunoblot Analysis.
Purified Stat proteins were electrophoresed in SDS/10% polyacrylamide gels, transferred to nitrocellulose, and detected as reported (11). For quantitation, the blots were probed using anti-Stat3 [Transduction Laboratories (Lexington, KY) catalog no. S21320] or anti-phosphotyrosine (4G10, Upstate Biotechnology) monoclonal antibody and 125I-labeled goat anti-mouse IgG (22). Labeled proteins were visualized and quantitated on a PhosphorImager.
In Vitro EGF Receptor Kinase Reaction.
EGF receptor kinase (EGFRKD) prepared in recombinant baculovirus-infected insect cells and purified by immunoabsorption with anti-phosphotyrosine monoclonal antibody (23) was supplied by D. Cadena and G. Gill or purchased from Stratagene. Results with each preparation were similar. SDS/polyacrylamide gel electrophoresis (PAGE) and staining with silver or Coomassie blue revealed a predominant protein corresponding to 68 kDa in each case, as reported (23). For the kinase reaction, purified Stat3 protein (2–10 μg) was mixed with 0.1 μg of EGFRKD in 40 μl of 12.5 mM Hepes, pH 7.4/50 μM Na3VO4/10 mM MnCl2/20 μM ATP/10 μCi of [γ-32P]ATP (1 Ci = 37 GBq). The mixtures were incubated for 1 hr at room temperature, and the reaction stopped by the addition of SDS/PAGE sampling buffer. The samples were subjected to SDS/PAGE and autoradiography. For use in EMSA, 10 μg of Stat3 proteins and 1 μg of EGFRKD were mixed in the same buffer without [γ-32P]ATP. The reactions were incubated with 10 mM EDTA overnight at 4°C, and aliqouts were analyzed by EMSA. PD157655, an inhibitor of EGF receptor kinase, was kindly provided by D. Fry (Parke Davis).
In Vitro Stat Binding to EGF Receptor Kinase.
Stat proteins and EGFRKD were mixed and phosphorylated as described above. Binding buffer (1 ml), indicated detergents, and Ni2+ matrix (100 μl) were added and incubated for 1 hr at 4°C with constant rotation. The matrix was collected, washed three times with the same buffer, and resuspended in SDS/PAGE sampling buffer. Proteins were separated on a SDS/10% polyacrylamide gel and visualized by autoradiography.
RESULTS
Preparation of Inactive and Active Forms of Stat3α and Stat3β.
In preparation for in vitro studies of activation of Stat3 proteins by EGF receptor kinase, both nonphosphorylated and tyrosine-phosphorylated (activated) recombinant forms of Stat3α and Stat3β containing 10 N-terminal histidine residues were prepared from baculovirus-infected Sf9 cells and assessed for DNA-binding activity. The tyrosine-phosphorylated proteins were prepared from cells coinfected with JAK1- or JAK2-expressing baculovirus (24). Each protein was purified to homogeneity (Fig. 1).
Figure 1.
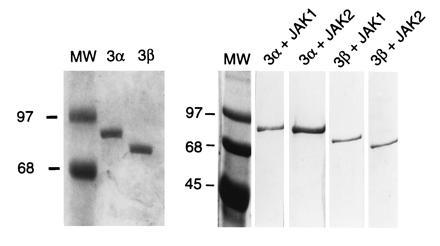
Coomassie blue-stained SDS/PAGE gels of Stat3α and Stat3β purified from insect Sf9 cells infected with recombinant baculovirus. (Left) Nonphosphorylated Stat proteins. (Right) Stat proteins phosphorylated in JAK-containing cells.
As seen in Fig. 2, in the absence of JAK1 or JAK2, the purified Stat3α contained no detectable phosphotyrosine, whereas the Stat3β was tyrosine-phosphorylated at a low level. “Constitutive” phosphorylation of Stat3β, but not Stat3α, had previously been observed in mammalian cells expressing the respective isoform from a transfecting plasmid (11). When the insect cells also contained either JAK1 or JAK2, Stat3α was phosphorylated on tyrosine, and the tyrosine phosphorylation of Stat3β increased markedly. The ratio of phosphotyrosine signal to Stat3 protein was generally found to be higher in the case of Stat3β compared with Stat3α, as illustrated in Fig. 2. Also shown in Fig. 2 is a comparison of JAK-associated tyrosine phosphorylation of wild-type Stat3 isoforms versus Y705F mutant proteins. Tyr-705 is the tyrosine residue that is phosphorylated in cells expressing Stat3α or Stat3β in response to EGF (T.S.S., unpublished work). The mutant proteins had essentially no phosphotyrosine. We conclude from this series of experiments that both Stat3 isoforms appear to be substrates for JAK1 or JAK2 resulting in phosphorylation of Tyr-705, although we cannot exclude the possibility that the JAKs act indirectly in the insect cells.
Figure 2.
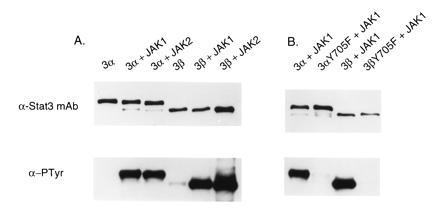
Tyr-705-phosphorylated Stat3α and Stat3β. Stat3 wild-type and Y705F mutant proteins were prepared from Sf9 cells in the presence or absence of JAK-expressing baculovirus. Purified Stat3 proteins were subjected to SDS/PAGE for immunoblot analysis either with anti-Stat3 antibody or with anti-phosphotyrosine antibody.
We next determined whether JAK1- and JAK2-associated phosphorylation of Stat3α and Stat3β leads to activation of the Stat proteins for DNA binding to a high-affinity site (high-affinity c-fos sis-inducible element, ref. 26), as anticipated from prior work with Stat proteins (27, 28). As seen in Fig. 3, Stat3α prepared from cells lacking JAK1 or JAK2 was inactive, and Stat3β (which showed a low level of tyrosine phosphorylation) had low activity. In each case, wild-type protein prepared from cells expressing JAK1 or JAK2 was active, whereas the Y705F protein was not.
Figure 3.
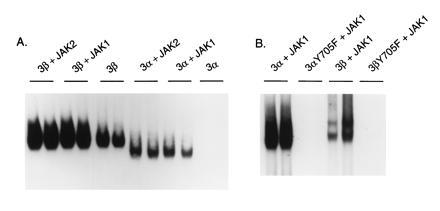
DNA binding activities of Stat3 proteins purified from JAK-containing Sf9 cells. EMSA was performed with two different amounts of each indicated Stat3 protein. (Left) Proteins at 0.1 (right lane of each pair) and 0.2 μg were present. (Right) Amounts of proteins were as follows: Stat3α and 3αY705F, 0.5 (left lane of each pair) and 1 μg, respectively; and Stat3β and 3βY705F, 1 and 2 ng, respectively.
As observed in EGF-treated mammalian cells (T.S.S., unpublished work), Stat3β has greater activity than Stat3α on a molar basis. From the results of several measurements with two different preparations of purified proteins, we estimate that the high-affinity c-fos sis-inducible element binding activity of Tyr-705-phosphorylated Stat3β is approximately 20- to 50-fold greater than that of Tyr-705-phosphorylated Stat3α.
In Vitro Phosphorylation and Activation of Stat3 by EGFRKD.
Having shown that Stat3α and Stat3β are phosphorylated on Tyr-705 in cells containing JAK1 or JAK2 and thereby activated for DNA binding, we wanted to determine if these isoforms could be tyrosine-phosphorylated and activated by signaling protein tyrosine kinases in vitro. In preliminary experiments involving immune complexes of JAK1 or JAK2 prepared from Sf9 cells expressing one or the other of these proteins, we could barely detect phosphorylation of Stat proteins, and we did not pursue this further. However, recombinant EGF receptor kinase purified from baculovirus-infected Sf9 cells (23, 29) was found to phosphorylate both purified Stat3α and Stat3β as well as itself. It should be noted that phosphorylation occurred in the absence of activators (23), and that autophosphorylation was stimulated by Stat (Fig. 4A). This reaction was inhibited by PD157655, a specific inhibitor of EGF receptor kinase. Moreover, there was evidence for the formation of a stable complex between the EGFRKD and the Stat3 protein (Fig. 4B). When the kinase domain and either Stat3α or Stat3β were incubated together in the presence of [γ-32P]ATP and the Stat protein was then adsorbed to a Ni2+- containing matrix in the presence of neutral detergent, both phosphorylated Stat and phosphorylated kinase were retained by the matrix. In the absence of Stat, the kinase was not retained. The complex between Stat and EGF receptor kinase appeared to be stable, requiring high concentration of ionic detergent for dissociation.
Figure 4.
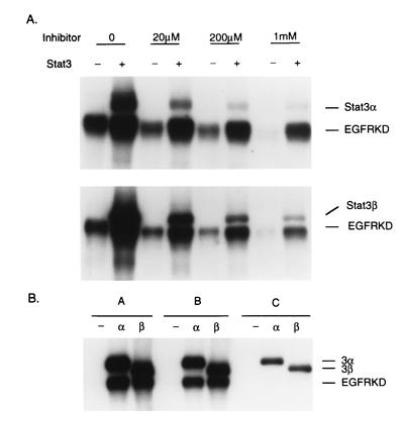
(A) In vitro phosphorylation of Stat3α and Stat3β by EGFRKD. Purified Stat3α (2 μg) and Stat3β (2 μg) were used for the kinase reaction; each reaction tube contained 0.1 μg of kinase. An EGF receptor kinase-specific inhibitor, PD157655, was included in the reactions at the indicated concentrations. The band labeled EGFRKD corresponds to receptor kinase detected by immunoblotting. (B) Formation of a stable complex between Stat3 protein and EGF receptor kinase. The detergents included in binding buffers [50 mM Tris·HCl, pH 8.0/1 mM phenylmethylsulfonyl fluoride/leupeptin (10 μg/ml)/50 mM imidazole] were as follows: A, 1% Triton X-100; B, 1% Nonidet P-40/0.1%SDS/0.5% sodium deoxycholate; C, 1% Nonidet P-40/1% SDS. Every reaction tube contained 0.1 μg of kinase and, where indicated, 2 μg of Stat.
That the phosphorylation of Stat3 isoforms by EGF receptor kinase was accompanied by activation for DNA binding is shown in Fig. 5. Just as in the case of Stat3 isoforms phosphorylated in cells expressing JAK1 or JAK2, both Stat3α and Stat3β were activated by EGF receptor kinase in vitro and the activity of phosphorylated Stat3β was manyfold greater than that of Stat3α. Furthermore, although the Y705F-substituted Stat proteins were still phosphorylated by EGF receptor kinase (in a PD157655-inhibitable reaction; data not shown), the resulting Stat proteins were unable to bind DNA (Fig. 6). We conclude from this series of experiments that EGF receptor kinase can interact directly with Stat3α and Stat3β, phosphorylate Tyr-705, and thereby activate them for DNA binding. The fact that the Y705F proteins are still phosphorylated by EGF receptor kinase suggests that one or more tyrosine residues of the wild-type Stat proteins other than Tyr-705 are also phosphorylated, but this remains to be determined.
Figure 5.
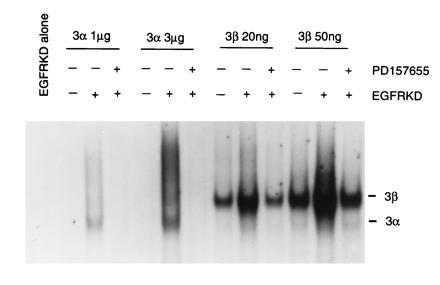
EGF receptor kinase activation of Stat3α and Stat3β for DNA binding. Stat3 proteins (10 μg) were phosphorylated by EGF receptor kinase (1 μg) with nonradioactive ATP in the presence or absence of PD157655 (1 mM). Indicated amounts of in vitro-phosphorylated Stat3 proteins were subjected to EMSA.
Figure 6.
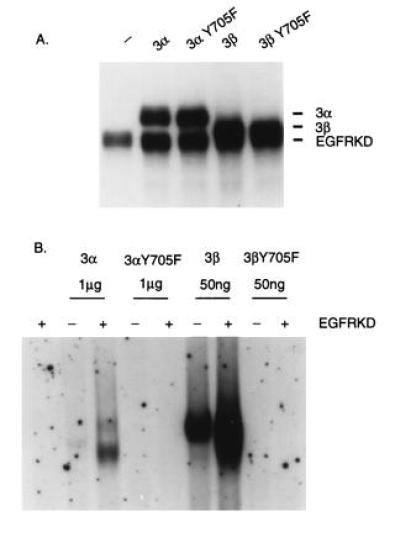
Requirement for Tyr-705 for in vitro activation of Stat3α and Stat3β by EGF receptor kinase. (A) Stat3 wild-type and Y705F mutant proteins (2 μg) were phosphorylated in vitro by EGF receptor kinase (0.1 μg) as described in Fig. 4. (B) In vitro-phosphorylated nonradioactive Stat3 proteins were subjected to EMSA as described in Fig. 5.
DISCUSSION
The main conclusion of this report is that EGF receptor kinase can phosphorylate Stat3α and Stat3β in vitro at Tyr-705, thereby activating the Stat proteins for DNA binding. This conclusion is consistent with the observation that in cell lines lacking individual JAK proteins, EGF still activated Stat (19, 20, 25) and with the results of experiments in which immune complexes of cell extracts prepared with anti-EGF receptor antibody activated Stat (21). In the latter case, and even in our own experiments, it is difficult to exclude the presence of a small amount of an intermediary kinase that copurifies with the receptor kinase. Our results do not imply that activation of Stat proteins in response to ligands that act through receptor kinases is solely or even mainly through direct phosphorylation of Stat by the receptor, since in certain cell lines JAK1 is phosphorylated (activated) in response to EGF (19, 30) and JAK1, JAK2, and Tyk2 are phosphorylated (activated) in response to PDGF (20). Therefore, it is likely that in at least some cells both direct receptor kinase activation and JAK-mediated activation of Stat occur in response to EGF or PDGF.
The significance of the complex formed in vitro between the EGF receptor kinase and Stat remains to be determined. We do not yet know if the complex (or complexes) contains phosphorylated and/or nonphosphorylated Stat (since both are retained by the Ni2+ resin), or if its formation requires phosphorylated EGF receptor kinase, as would be anticipated from an interaction involving the SH2 domain of Stat (31). The complex could be an intermediate in the phosphorylation reaction and/or in the release of phosphorylated Stat from the receptor kinase. These possibilities should be testable with purified components.
There was a marked quantitative difference in the DNA-binding activities of the tyrosine-phosphorylated isoforms, whether phosphorylated by JAK1 or JAK2 in insect cells (Fig. 3) or by EGF receptor kinase in vitro (Fig. 5). Phosphorylated Stat3β had 20- to 50-fold greater activity than Stat3α on a molar basis. Although the reason for the difference is not yet clear, we suspect that phosphorylated Stat3β forms more stable dimers than phosphorylated Stat3α. Whatever underlies the difference, it is likely to be reflected in the transcriptional activities of each isoform during the cellular response to ligands. It may also be generalizable to other pairs of Stat isoforms.
Finally, we note that the quantitative difference between the DNA binding activities of Stat3α and Stat3β reported herein is not the only difference in their transcription-related activities. Stat3β was discovered as a c-Jun-interacting protein (11) and was shown to cooperate with Jun or Jun/Fos in the activation of an interleukin 6-responsive promoter that has binding sites for both Stat3 and Jun. Stat3α, in contrast, showed no transcriptional cooperativity (11). Furthermore, only Stat3β showed constitutive activation in transfected cells incubated in the absence of known cytokines or growth factors, suggesting that high levels of short forms of Stat3 as seen in certain tumor cell lines (T.S.S., unpublished observation), might by-pass dependence on ligand for some of the cellular responses to growth factors and cytokines.
Acknowledgments
We thank L. K. Sanders for expert technical assistance, J. N. Ihle for JAK-expressing baculoviruses, D. Cadena and G. Gill for highly purified EGF receptor kinase, and D. Fry for compound PD157655.
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Abbreviations: EGF, epidermal growth factor; PDGF, platelet-derived growth factor; EMSA, electrophoretic mobility-shift assay; EGFRKD, EGF receptor kinase domain.
References
- 1.Ihle J N. Cell. 1996;84:331–334. doi: 10.1016/s0092-8674(00)81277-5. [DOI] [PubMed] [Google Scholar]
- 2.Schindler C, Darnell J E., Jr Annu Rev Biochem. 1995;64:621–651. doi: 10.1146/annurev.bi.64.070195.003201. [DOI] [PubMed] [Google Scholar]
- 3.Darnell J E, Jr, Kerr I M, Stark G R. Science. 1994;264:1415–1421. doi: 10.1126/science.8197455. [DOI] [PubMed] [Google Scholar]
- 4.Fu X-Y, Schindler C, Improta T, Aebersold R, Darnell J E., Jr Proc Natl Acad Sci USA. 1992;89:7840–7843. doi: 10.1073/pnas.89.16.7840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Veals S A, Schindler C, Leonard D, Fu X-Y, Aebersold R, Darnell J E, Jr, Levy D E. Mol Cell Biol. 1992;12:3315–3324. doi: 10.1128/mcb.12.8.3315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schindler C, Fu X-Y, Improta T, Aebersold R, Darnell J E., Jr Proc Natl Acad Sci USA. 1992;89:7836–7839. doi: 10.1073/pnas.89.16.7836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhong Z, Wen Z, Darnell J E., Jr Proc Natl Acad Sci USA. 1994;91:4806–4810. doi: 10.1073/pnas.91.11.4806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Akira S, Nishio Y, Inoue M, Wang X-J, Wei S, Matsusaka T, Yoshida K, Sudo T, Naruto M, Kishimoto T. Cell. 1994;77:63–71. doi: 10.1016/0092-8674(94)90235-6. [DOI] [PubMed] [Google Scholar]
- 9.Wakao H, Gouilleux F, Groner B. EMBO J. 1994;13:2182–2191. doi: 10.1002/j.1460-2075.1994.tb06495.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hou J, Schindler U, Henzel W J, Ho T C, Brasseur M, McKnight S L. Science. 1994;265:1701–1706. doi: 10.1126/science.8085155. [DOI] [PubMed] [Google Scholar]
- 11.Schaefer T S, Sanders L K, Nathans D. Proc Natl Acad Sci USA. 1995;92:9097–9101. doi: 10.1073/pnas.92.20.9097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Müller M, Briscoe J, Laxton C, Guschin D, Ziemiecki A, Silvennoinen O, Harpur A G, Barbieri G, Witthuhn B A, Schindler C, Pellegrini S, Wilks A F, Ihle J N, Stark G R, Kerr I M. Nature (London) 1993;366:129–136. doi: 10.1038/366129a0. [DOI] [PubMed] [Google Scholar]
- 13.Wilks A F. Proc Natl Acad Sci USA. 1989;86:1603–1607. doi: 10.1073/pnas.86.5.1603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stahl N, Farruggella T J, Boulton T G, Zhong Z, Darnell J E, Jr, Yancopoulos G D. Science. 1995;267:1349–1353. doi: 10.1126/science.7871433. [DOI] [PubMed] [Google Scholar]
- 15.Heim M H, Kerr I M, Stark G R, Darnell J E., Jr Science. 1995;267:1347–1349. doi: 10.1126/science.7871432. [DOI] [PubMed] [Google Scholar]
- 16.Shuai K, Stark G R, Kerr I M, Darnell J E., Jr Science. 1993;261:1744–1746. doi: 10.1126/science.7690989. [DOI] [PubMed] [Google Scholar]
- 17.Kaptein A, Paillard V, Saunders M. J Biol Chem. 1996;271:5961–5964. doi: 10.1074/jbc.271.11.5961. [DOI] [PubMed] [Google Scholar]
- 18.Ullrich A, Schlessinger J. Cell. 1990;61:203–212. doi: 10.1016/0092-8674(90)90801-k. [DOI] [PubMed] [Google Scholar]
- 19.Leaman D W, Pisharody S, Flickinger T W, Commane M A, Schlessinger J, Kerr I M, Levy D E, Stark G R. Mol Cell Biol. 1996;16:369–375. doi: 10.1128/mcb.16.1.369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vignais M-L, Sadowski H B, Watling D, Rogers N C, Gilman M. Mol Cell Biol. 1996;16:1759–1769. doi: 10.1128/mcb.16.4.1759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Quelle F W, Thierfelder W, Witthuhn B A, Tang B, Cohen S, Ihle J N. J Biol Chem. 1995;270:20775–20780. doi: 10.1074/jbc.270.35.20775. [DOI] [PubMed] [Google Scholar]
- 22.Burnette W N. Anal Chem. 1981;112:195–203. doi: 10.1016/0003-2697(81)90281-5. [DOI] [PubMed] [Google Scholar]
- 23.Wedegaertner P B, Gill G N. J Biol Chem. 1989;264:11346–11353. [PubMed] [Google Scholar]
- 24.Schindler U, Wu P, Rothe M, Brasseur M, McKnight S L. Immunity. 1995;2:689–697. doi: 10.1016/1074-7613(95)90013-6. [DOI] [PubMed] [Google Scholar]
- 25.David M, Wong L, Flavell R, Thompson S A, Wells A, Larner A C, Johnson G R. J Biol Chem. 1996;271:9185–9188. doi: 10.1074/jbc.271.16.9185. [DOI] [PubMed] [Google Scholar]
- 26.Wagner B J, Hayes T E, Hoban C J, Cochran B H. EMBO J. 1990;9:4477–4484. doi: 10.1002/j.1460-2075.1990.tb07898.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sadowski H B, Shuai K, Darnell J E, Jr, Gilman M Z. Science. 1993;261:1739–1744. doi: 10.1126/science.8397445. [DOI] [PubMed] [Google Scholar]
- 28.Zhong Z, Wen Z, Darnell J E., Jr Science. 1994;264:95–98. doi: 10.1126/science.8140422. [DOI] [PubMed] [Google Scholar]
- 29.Wedegaertner P B, Gill G N. Arch Biochem Biophys. 1992;292:273–280. doi: 10.1016/0003-9861(92)90079-c. [DOI] [PubMed] [Google Scholar]
- 30.Shuai K, Ziemiecki A, Wilks A F, Harpur A G, Sadowski H B, Gilman M Z, Darnell J E., Jr Nature (London) 1993;366:580–583. doi: 10.1038/366580a0. [DOI] [PubMed] [Google Scholar]
- 31.Fu X-Y, Zhang J-J. Cell. 1993;74:1135–1145. doi: 10.1016/0092-8674(93)90734-8. [DOI] [PubMed] [Google Scholar]