

Abstract
Saccharomyces telomeres consist of ≈300 bp of C1–3A/TG1–3 DNA. Cells lacking the activity of the essential gene CDC13 display a cell cycle arrest mediated by the DNA damage sensing, RAD9 cell cycle checkpoint, presumably because they exhibit strand-specific loss of telomeric and telomere-adjacent DNA [Garvik, B., Carson, M. & Hartwell, L. (1995) Mol. Cell. Biol. 15, 6128–6138]. Cdc13p expressed in Escherichia coli or overexpressed in yeast bound specifically to single-strand TG1–3 DNA. The specificity of binding displayed by Cdc13p in vitro indicates that in vivo it could bind to both the short, constitutive single-strand TG1–3 tails thought to be present at telomeres at most times in the cell cycle as well as to the long single-strand TG1–3 tails that are intermediates in telomere replication. Genes located near yeast telomeres are transcriptionally repressed, a phenomenon known as telomere position effect. Cells overexpressing a mutant form of Cdc13p had reduced telomere position effect at high temperatures. These data suggest that Cdc13p functions by binding directly to telomeric DNA, thereby limiting its accessibility to degradation and transcription as well as masking it from factors that detect damaged DNA.
Telomeric DNA in virtually all eukaryotes consists of a stretch of repeated DNA in which the strand running 5′ to 3′ from the center of the chromosome to its end has clusters of G residues (reviewed in ref. 1). For example, Saccharomyces chromosomes end in ≈300 bp of a heterogeneous sequence, C1–3A/TG1–3 (2). In ciliated protozoa, the G strand at each telomere is extended to form a 12- to 16-base single-strand (ss) G tail that is present during all or most of the cell cycle (3, 4, 5). In yeast, transient ss TG1–3 tails >30 bases in length are detected on plasmid and chromosomal telomeres late in the S phase (6).
Yeast chromosomes without telomeres are unstable (7). Telomeric DNA is essential for both the replication and protection of chromosome ends and helps the RAD9 DNA damage sensing, cell cycle checkpoint distinguish intact from broken chromosomes. In addition, in yeast, the transcription of telomere-linked genes is repressed (8), a phenomenon known as telomere position effect (TPE).
Yeast telomeric DNA is assembled into a nonnucleosomal chromatin structure, the telosome (9). The major telosomal protein in yeast is Rap1p (10, 11). Rap1p is a sequence-specific, duplex DNA-binding protein (12, 13). Although Rap1p also binds to ss TG1–3 DNA, it does so with about three orders of magnitude less affinity than to duplex telomeric DNA (14). Rap1p binding is not limited to telomeres because there are many internal binding sites for Rap1p in vivo (e.g., see ref. 15).
In contrast to Rap1p, the major telomere binding proteins in ciliated protozoa such as Oxytricha (16) and Euplotes (17) are telomere-limited. These proteins are restricted to telomeres because their binding requires the ss G tail found at each telomere. Thus, these telomere-limited binding proteins could theoretically help cells distinguish intact from broken DNA molecules. In vitro, the ciliate proteins protect telomeres from exonucleolytic degradation (16, 18) and mediate telomere–telomere interactions (19). Genetic (20) and biochemical (21) data suggest that other organisms, including yeast, also have telomere-limited binding proteins.
Replication of chromosomal DNA molecules is initiated by 8- to 12-base RNA primers. The following model for telomere replication in yeast is based on studies with linear plasmids. Telomeric DNA replicates late in the S phase (22). After semiconservative replication of telomeric DNA occurs, both telomeres of a given DNA molecule acquire a ≈50- to 100-base ss TG1–3 tail by a telomerase independent mechanism (23). These ss TG1–3 tails are probably generated by cell cycle-regulated degradation of the C1–3A strand. The long ss TG1–3 tails are converted to duplex DNA before mitosis (22). Although conventional replication activities can synthesize the complementary C1–3A strand, removal of the terminal RNA primers will generate an 8- to 12-base ss TG1–3 tail at each telomere. Thus, short ss TG1–3 tails, potential substrates for a ss G tail binding protein, should occur at both telomeres of an individual chromosome.
Cells limited for the essential protein Cdc13p arrest in the G2 phase of the cell cycle by a RAD9-dependent process (24). When cdc13-1 cells are grown at semipermissive temperature, they show an increase in mitotic recombination in telomere-adjacent regions of the chromosome (25). Moreover, cdc13-1 cells arrested by growth at restrictive temperatures contain ss telomeric and telomere-adjacent DNA, with the strand running 5′ to 3′ from the end toward the center of the chromosome, the C1–3A strand, being preferentially degraded. In rad9 cdc13-1 cells, this strand-specific degradation extends for as much as 30 kb from the end of the chromosome (25). These data suggest that Cdc13p limits the C-strand degradation that occurs at the end of the S phase (23).
Here we show that both Escherichia coli- and Saccharomyces-produced Cdc13p binds ss TG1–3 DNA in vitro. The binding properties of Cdc13p are consistent with its being able to bind both a short ss TG1–3 tail, probably present during most of the cell cycle, and the long ss TG1–3 tails generated at the end of S phase. Cells overexpressing the cdc13-1 mutant form of the protein had defects in TPE at high temperature. Together with previous information (25), these data suggest that Cdc13p is a telosomal protein in vivo.
MATERIALS AND METHODS
Oligonucleotides, Competitor Nucleic Acids, and Electrophoretic Mobility-Shift Assays.
Oligonucleotides (DNAgency, Malvern, PA) are listed in Table 1. The sequence of TG22 was based on the sequence of a cloned yeast telomere (2). Oligonucleotides were 32P-labeled by T4 polynucleotide kinase (New England Biolabs) and purified from a 12% sequencing gel. For ease of comparison, in the competition experiments, concentration for competitors containing ss TG1–3 DNA was expressed in terms of the number of bases of TG1–3 DNA. For example, if the concentration of the oligonucleotide TG22 was 1 molecule per ml, a solution of 3′-Tel, a 42-base oligonucleotide that also had 22 bases of TG1–3 DNA, of equal concentration would be 1 molecule per ml, whereas TG43 and ss pTG270 would be, respectively, 0.51 and 0.08 molecule per ml. To make tailed duplex or fully duplex substrates, labeled TG19 was annealed with an excess of the CAS9, CAS14, or CAS19 oligonucleotide, and the products were purified from a 12% polyacrylamide gel. Total yeast RNA was from BDH. To generate pTG270, 276 bp of TG1–3 DNA from a cloned telomere (26) was inserted into pYES2 (Invitrogen). ss pTG270 was prepared by superinfection of XL1-blue cells carrying pTG270 with M13-CO8 followed by precipitation of phagemids by PEG. Phagemids were extracted with phenol/chloroform and ethanol-precipitated. ss pTG270 was sheared by sonication and sized by agarose gel electrophoresis. Duplex pTG270 was prepared by a standard plasmid preparation from the same strain but without M13-C08 infection.
Table 1.
Oligonucleotides used in this study
| Name | Sequence |
|---|---|
| TG22 | 5′-GTGGTGGGTGGGTGTGTGTGGG-3′ |
| TG43 | 5′-GTGGTGGGTGGGTGTGTGTGGGTGTGGTGGGTGTGTGGGTGTG-3′ |
| TG19 | 5′-GCGGATCCCGGGTGTGGTG-3′ |
| CAS9 | 5′-GGGATCCGC-3′ |
| CAS14 | 5′-CACCCGGGATCCGC-3′ |
| CAS19 | 5′-CACCACACCCGGGATCCGC-3′ |
| 3′ Tel | 5′-GCCATGGCCACCGGGATCCCGTGGTGGGTGGGTGTGTGTGGG-3′ |
| Internal Tel | 5′-GCCATGGCCAGTGGTGGGTGGGTGTGTGTGGGCCGGGATCCC-3′ |
| 5′ Tel | 5′-GTGGTGGGTGGGTGTGTGTGGGGCCATGGCCACCGGGATCCC-3′ |
| CDC135Δ | 5′-TGTTTCTCTTTGGATACGAATGACCGTGGAAACTATCGCCTCGTTCAGAATGACACG-3′ |
| CDC133Δ | 5′-GCAATTTGGCACCGCCGCGTTGGGCTGCGCGGATCATGTCCTCTTGGCCTCCTCTAG-3′ |
| CDC135 | 5′-GCTCTAGAGCCCGGGCCATGGATACCTTAGAAG-3′ |
| CDC133 | 5′-AACTGCATACTAGTCGACTCTTGCTTCTTACC-3′ |
| CDC13ts5 | 5′-AGCTCTAGATGCATGGATACCTTAGAAGAGCCTGAG-3′ |
| CDC13ts3 | 5′-AGGCTCGCAGTCGACTTGGGCTGCGCGGATCATGTC-3′ |
Total yeast or E. coli extracts for the gel mobility-shift assays were prepared from 5-ml cultures as described (27). The concentration of protein in the yeast and E. coli extracts was ≈1-2 mg/ml or ≈2-4 mg/ml, respectively. Cell extracts were mixed with 1.0 ng of 32P-labeled TG22 in a 15-μl reaction mixture containing 50 mM Tris·HCl (pH 7.5), 1 mM EDTA, 50 mM NaCl, 1 mM DTT, and 1 μg of ss poly(dI·dC) and incubated at room temperature for 10 min. Three microliters of 80% glycerol was added and the reaction mixtures were loaded on an 8% nondenaturing polyacrylamide gel. Before loading, gels were run at 120 V for 10 min. Electrophoresis was in TBE (89 mM Tris·borate/2 mM EDTA) at 120 V for 2.5 hr after loading the samples. The gel was dried and autoradiographed. For competition analysis, 1.0 ng of 32P-labeled TG22 was mixed with varying amounts of cold competitors before addition of the cell extracts. Binding activity was quantified with a Molecular Dynamics PhosphorImager.
Expression of Cdc13p in Bacteria and Yeast.
To construct plasmids that overexpressed Cdc13p in S. cerevisiae, oligonucleotides CDC135 and CDC133 were mixed with YEP24-CDC13-161-4, a plasmid containing the CDC13 gene (25), and PCR-amplified using the Expand High Fidelity PCR system (Boehringer Mannheim). The PCR product was digested with NcoI and SalI and the ≈3-kb fragment was gel-purified and inserted into NcoI–SalI-digested pTHA to generate pTHA-CDC13. pTHA is a modified form of the 2-μm LEU2 plasmid pRS425 (28). To generate pTHA, a PGK1 promoter, a tri-HA tag, and a poly(A) tail were inserted into the SpeI, SmaI, and ApaI sites, respectively, of pRS425 (a generous gift from C. Lin, Fred Hutchinson Cancer Research Center). The resulting plasmid was digested with PstI, made blunt-ended with T4 DNA polymerase, and ligated to an NcoI linker. In pTHA-CDC13, the complete CDC13 ORF was fused in frame with three HA tags at its amino terminus and expressed from the PGK1 promoter. Other than the HA tags, the Cdc13p produced from pTHA-CDC13 was identical to the wild-type protein.
To express Cdc13p in E. coli, the PCR products obtained using oligonucleotides CDC135 and CDC133 were digested with NcoI and PstI and ligated to NcoI–PstI-digested pKK233-2 (Pharmacia) to generate pKKCDC13. In this plasmid, CDC13 was under the control of the isopropyl β-d-thiogalactoside-inducible E. coli trc promoter from the first AUG to ≈100 bp past the CDC13 termination codon.
To clone cdc13-1, genomic DNA from strain 2758–8-4b (MATa cdc13-1 his7 leu2-3, 112 ura3-52 trp1-289; provided by L. Hartwell, University of Washington) was PCR-amplified with oligonucleotides CDC13ts5 and CDC13ts3 using the Expand High Fidelity PCR system (Boehringer Mannheim). The PCR product was cloned into pCRII provided in the TA Cloning Kit (Invitrogen) to generate pTA-cdc13-1. To express the cdc13-1 gene in yeast, pTA-cdc13-1 was digested with SalI and XhoI, and the 2.8-kbp SalI–XhoI fragment containing the gene was isolated and used to replace the comparable SalI–XhoI fragment of CDC13 in pTHA-CDC13 to generate a plasmid in which all but the 5′ most 55 bp of the gene were derived from cdc13-1. The following criteria established that pTHA-cdc13-1 contained the correct gene: (i) For 11 of 12 restriction enzymes, cdc13-1 had the same restriction map as CDC13. (ii) Cloned cdc13-1 had an EcoRI site not present in CDC13 that was present in genomic DNA from the cdc13-1 strain 2758–8-4b. Although this polymorphism was not sequenced, we infer from the position of the mutation and examination of the CDC13 sequence that it generated a proline-to-serine change at amino acid 371. (iii) pTHA-cdc13-1 rescued the lethality of cdc13Δ strains at 25°C and 30°C, but these cells had growth defects at higher temperatures and did not grow at temperatures >37°C (data not shown).
Yeast Methods.
All yeast strains were constructed in the YPH499 background (MATa ura3-52 lys3-5 ade2-10 trp1-Δ63 his3-Δ200 leu2-Δ1) (28) but with URA3 and ADE2 inserted near the telomeres of chromosomes VII-L and V-R, respectively, as described (8). A null mutant of CDC13 was constructed in an otherwise homozygous diploid of YPH499 that was heterozygous for the telomere insertions as described (29) using oligonucleotides CDC135Δ and CDC133Δ. This procedure precisely deleted the CDC13 ORF and replaced it with HIS3 to create YJL401-UTAT. Spore products from YJL401-UTAT showed 2:2 segregation for viability, with the viable spores being His−. Plasmids were introduced by transformation into YJL401-UTAT, tetrads were dissected, and strains with the appropriate genotype were identified.
For TPE assays, strains were streaked on yeast complete (YC)-Leu plates and grown for 5 days at 25°C. Colonies were resuspended in water, and aliquots of different dilutions were plated on YC-Leu plates or 5-fluoroorotic acid (FOA)-Leu plates. For cells grown at 25°C, colonies were counted after 6 days; for cells grown at 30°C, colonies were counted after 3 days for YC-Leu and after 6 days for FOA-Leu plates; for YC-Leu cells grown at 37°C, colonies were counted after 6 days for cdc13Δ/pTHA-cdc13-1 cells and after 3 days for other strains. Cells grown at 37°C on FOA-Leu plates were counted after 14 days. The frequency of TPE is the median frequency of FOA resistant (FOAR) cells from 5 to 10 colonies for each strain.
RESULTS
E. coli- or Saccharomyces-Synthesized Cdc13p Is a ss TG1–3 Binding Protein in Vitro.
The sequence of the CDC13 gene predicts a 924-amino acid protein with a molecular mass of 104,903 Da (25). Extracts were prepared from E. coli expressing full-length Cdc13p (Fig. 1A, lanes 4 and 5) or from cells carrying vector alone (Fig. 1A, lanes 2 and 3), mixed with a 32P-labeled 22-base TG1–3 oligonucleotide (Table 1, TG22), and assayed for DNA binding using a gel mobility-shift assay. E. coli cells carrying vector alone had proteins that bound ss TG1–3 DNA, but these activities appeared to be RNA-binding proteins (Fig. 2A, lanes 15–17). In addition, cells expressing Cdc13p had a new protein–TG22 complex (Fig. 1, indicated by arrows) that was absent in the extract from cells carrying the vector alone.
Figure 1.
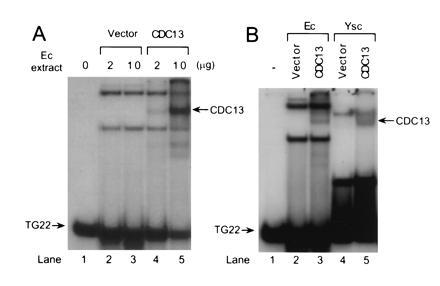
Cdc13p is a ss TG1–3 binding protein. 32P-labeled TG22, a 22-base oligonucleotide of ss TG1–3 DNA (Table 1), was used in gel mobility-shift assays using total extracts from either bacteria (Ec) or yeast (Ysc). All gel mobility-shift assays reported here were done in the presence of ss poly(dI·dC). E. coli extracts were prepared from cells expressing Cdc13p or carrying the vector alone. Yeast extracts were prepared from a CDC13 strain carrying vector alone or pTHA-CDC13 for overexpression of Cdc13p. The arrows point to a TG22-protein complex that was seen only in E. coli cells expressing Cdc13p or yeast cells overexpressing Cdc13p. For both panels, the first lane had no extract. (A) The amount of extract used is indicated. (B) Two micrograms of extract was used.
Figure 2.
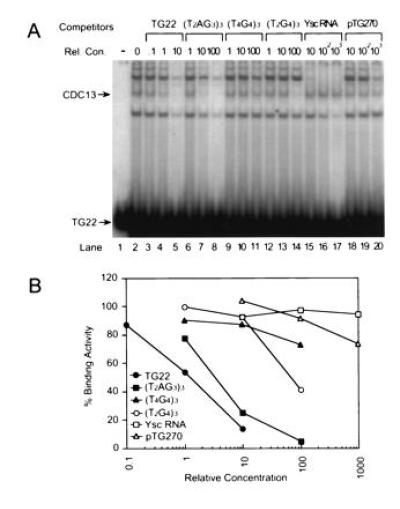
Cdc13p binding is specific. (A) Gel mobility-shift assays were carried out using 1 ng of 32P-labeled TG22 and 10 μg of E. coli extract from cells expressing Cdc13p. Prior to loading the gel, the TG22 was mixed with unlabeled ss oligonucleotides of the sequence and at the relative concentration (Rel. Con.) indicated above each lane or with total yeast RNA or pTG270, a 6300-bp plasmid containing 270 bp of C1–3A/TG1–3 DNA. The Cdc13p–TG22 complex is indicated by the arrow. Lane 1 had no extract. (B) The amount of gel shift activity in the Cdc13p-TG22 complex was quantified for each oligonucleotide at each concentration. Relative concentration of competitor nucleic acids in all gels was expressed as the ratios of the amount of ss TG1–3 DNA in the competitor to the amount of ss TG1–3 DNA in TG22 (see Materials and Methods); for competitors that did not have ss TG1–3 DNA (e.g., Ysc RNA), it was expressed as the ratio of nanograms of competitor to nanograms of TG22. In this and subsequent gels, the amount of gel shifted complex in TG22 with no competitor was defined as 100%. The values presented are the average of two independent experiments; for each point, the two values were within 14% of each other.
To study Cdc13p binding activity in yeast, the CDC13 ORF was inserted into pTHA, a multicopy yeast vector in which the transcription of inserted genes is under the control of the constitutive, efficient PGK1 promoter. Thus, cells carrying pTHA-CDC13 are expected to produce more Cdc13p than wild-type cells. At 25°C, 30°C, and 37°C, wild-type or cdc13Δ cells carrying pTHA-CDC13 grew at a rate comparable to that of wild-type cells with pTHA alone, indicating that the cloned gene complemented a cdc13Δ strain and that moderate overexpression of Cdc13p caused no major growth defects (data not shown).
Extracts prepared from wild-type yeast cells contain multiple, abundant ss TG1–3 binding activities as detected by a gel mobility shift assay, although most of these activities are probably due to RNA binding proteins (27). Extracts prepared from wild-type cells carrying vector alone and analyzed by a gel mobility-shift assay using TG22 had no detectable protein–DNA complex with the mobility of the E. coli-generated Cdc13p–TG22 complex (Fig. 1B, lane 4). However, when extract from cells overexpressing Cdc13p was mixed with labeled TG22, a complex with the mobility expected for Cdc13p-TG22 was detected (Fig. 1B, lane 5). Together with the E. coli results, these data indicate that yeast-generated Cdc13p is a ss TG1–3 binding protein in vitro but that wild-type cells express too little of it to allow its detection by the gel mobility shift assay.
Characterization of the Cdc13p ss TG1–3 Binding Activity.
To determine the specificity of the Cdc13p binding activity, extracts prepared from E. coli expressing Cdc13p were mixed with 32P-labeled TG22 and varying amounts of unlabeled competitor nucleic acids and assayed by the gel mobility-shift assay. As expected, unlabeled TG22 competed very efficiently with TG22, such that binding was reduced by 50% when labeled and competitor TG22 were present at equal concentrations (Fig. 2A, lanes 3–5). An oligonucleotide with the sequence of the G strand of vertebrate telomeric DNA, (T2AG3)3, also competed for Cdc13p binding (Fig. 2A, lanes 6–8), although ≈10 times as much (T2AG3)3 was needed to obtain the same level of competition as that obtained with TG22 (Fig. 2B). In contrast, two other G-strand telomeric DNAs, (T4G4)3, the Oxytricha telomeric sequence, and (T2G4)3, the Tetrahymena telomeric sequence, were poor competitors for Cdc13p binding (Fig. 2A, lanes 9–14). In addition, total yeast RNA (Fig. 2A, lanes 15–17) and duplex C1–3A/TG1–3 DNA (pTG270; Fig. 2A, lanes 18–20; see also Fig. 3, lane 12) did not compete for Cdc13p binding.
Figure 3.
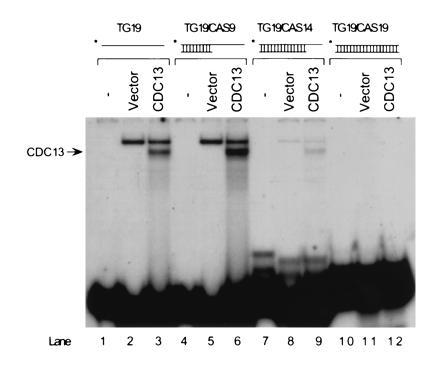
Cdc13p binds tailed duplex molecules. Gel mobility-shift assays were carried out using 10 μg of extract from E. coli cells expressing Cdc13p or carrying vector alone. 32P-labeled TG19 was annealed to various oligonucleotides to generate substrates with 9 bp of duplex DNA and a 10-base ss TG1–3 tail (TG19/CAS9), with 14 bp of duplex DNA and a 5-base ss TG1–3 tail (TG19/CAS14), or duplex TG19 (TG19/CAS19). The arrow indicates the position of the Cdc13p-DNA complex. The complex migrating more slowly than the Cdc13p-DNA complex in the vector-alone lanes was present in reduced amounts in lane 8, compared with lanes 2 and 5, and absent in lane 11 because it was due to an E. coli-encoded ss TG1–3 DNA binding activity, and there was less (lane 8) or no (lane 11) ss TG1–3 DNA in the DNA substrates used in those lanes.
To determine if ss TG1–3 needs to be at the very end of the oligonucleotide for Cdc13p binding, 42-base oligonucleotides (Table 1) having TG22 sequences at the 3′ end (3′ Tel), the 5′ end (5′ Tel), or in the middle (internal Tel) of the oligonucleotide were constructed. Each oligonucleotide was tested as a competitor in a gel mobility-shift assay using TG22 and E. coli-expressed Cdc13p (Fig. 4A). Because internal Tel and 3′ Tel both competed as well as TG22 and 5′ Tel competed almost as well, ss TG1–3 DNA need not be at the very end of the molecule to support Cdc13p binding.
Figure 4.
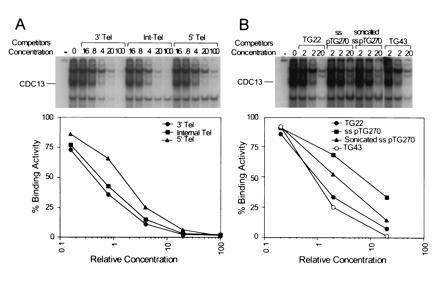
Cdc13p binding does not require an end. Gel mobility-shift assays were carried out using 1 ng of 32P-labeled TG22 and 10 μg of extract from E. coli expressing Cdc13p. TG22 was mixed with unlabeled competitor DNA, extract was added, and the samples were examined by electrophoresis. The % binding activity was determined as described in Fig. 2. In the interest of space, only the top portion of each gel is shown. (A) The competitor DNAs were unlabeled 42-base ss oligonucleotides having TG22 at its 3′ end (3′ Tel), in the middle of the oligonucleotide (Internal Tel), or at its 5′ end (5′ Tel). (B) The competitor DNA was unlabeled TG22, ss pTG270, a circular 6300-base ss phagemid containing 276 bases of ss TG1–3 DNA, ss pTG270 sheared to an average size of 1000 bases, or TG43, an oligonucleotide having 43 bases of ss TG1–3 DNA.
Cdc13p Has the Appropriate Specificity to Bind to the Long TG1–3 Tails Present at the End of the S Phase and to Nonreplicating Telomeres.
At the end of the S phase, yeast telomeres transiently acquire ss TG1–3 tails that are >30 bases in length (6). To determine if longer ss TG1–3 tails are substrates for Cdc13p binding in vitro, TG22 binding to E. coli-generated Cdc13p was examined using a 43-base TG1–3 oligonucleotide (TG43; Table 1) as competitor (Fig. 4B). TG43 competed efficiently in this assay such that one molecule of TG43 was about as effective as two molecules of TG22. Thus, Cdc13p also binds efficiently in vitro to oligonucleotides of about the length of the transient ss TG1–3 tails seen at the end of the S phase.
To determine if Cdc13p requires an end for binding, Cdc13p binding to TG22 was examined using phagemid pTG270 as competitor (Fig. 4B) This ≈6300-base circular ss DNA contains a 270-base stretch of ss TG1–3 DNA. The phagemid DNA was used intact or after shearing to fragments of ≈1000 bases. Although both DNAs were effective competitors, sheared ss pTG270 was better than intact ss pTG270. However, the difference was relatively small, with three times as much intact ss pTG270 needed to obtain the same level of competition as sheared ss pTG270. Thus, Cdc13p may have a modest preference for, but does not require, an end for binding.
By analogy to ciliated protozoa, telomeres are thought to consist of duplex telomeric DNA with a short 3′ ss G tail, hereafter called a “tailed duplex.” To determine if Cdc13p binds tailed duplex molecules in vitro, a 19-base oligonucleotide, TG19, containing 10 bases of TG1–3 DNA and 9 bases of nontelomeric DNA was mixed with complementary oligonucleotides of various lengths (Table 1). Tailed duplex substrates were generated that had either 9 bp of duplex DNA with a 10-base 3′ ss TG1–3 tail or 14 bp of duplex DNA with a 5-base 3′ ss TG1–3 tail. These substrates were mixed with an E. coli extract containing Cdc13p and examined by the gel mobility-shift assay (Fig. 3). As controls, the binding of Cdc13p to the TG19 ss oligonucleotide or to duplex TG19 was examined.
Cdc13p bound to TG19, demonstrating that 10 bases of ss TG1–3 are sufficient for Cdc13p binding (Fig. 3, lane 3), but not to duplex TG19 (Fig. 3, lane 12). Cdc13p bound efficiently to the tailed duplex containing 10 bases of ss TG1–3 DNA (Fig. 3, lane 6) and very weakly to the tailed duplex with a 5-base ss TG1–3 tail (Fig. 3, lane 9). Because the binding of Cdc13p to the tailed duplex was, if anything, better than the binding to ss TG19 (Fig. 3, compare lanes 3 and 6), Cdc13p has the appropriate binding specificity to be associated with telomeres in vivo.
Cells Expressing cdc13-1p Have Reduced TPE at High Temperature.
Genetic evidence suggests that yeast has a telomere-limited binding protein that is important for TPE (20). TPE is typically measured by determining the fraction of cells able to grow on plates containing FOA (8), a compound that kills cells expressing Ura3p (30), in a strain that has URA3 next to a telomere. Because CDC13 is essential, TPE cannot be measured in a cdc13Δ strain. The only available conditional allele of CDC13 is cdc13-1. We did not detect defects in TPE in a cdc13-1 strain. However, the maximum permissive temperature at which cdc13-1 cells grow is 25°C (25), and these cells accumulate suppressor mutations at permissive temperatures. Moreover, individual colonies from both wild-type and mutant strains yielded widely varying frequencies of TPE when assayed at 25°C (Fig. 5). Finally, the isogenic wild-type strain had very low TPE, such that only ≈0.1% of its cells were FOAR at 25°C, whereas other wild-type strains typically have ≈30-50% FOAR cells when analyzed in the same way (8). As an alternative to studying TPE in a cdc13-1 strain, we studied TPE in a cdc13Δ strain carrying pTHA-cdc13-1.
Figure 5.
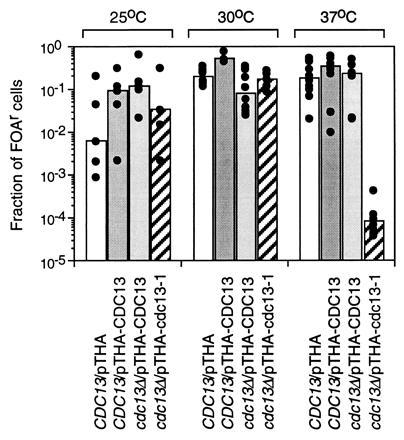
TPE is reduced at high temperature in cells overexpressing Cdc13-1p. CDC13 or cdc13Δ cells with URA3 next to the telomere of chromosome VII-L and carrying the plasmids indicated below the bars were spread on plates containing or lacking FOA and incubated at the indicated temperatures. The top of the bar is the median value for the fraction of FOAR cells, with dots indicating values for independent colonies. Independent colonies with identical values are represented by a single dot.
The cdc13-1 gene was cloned by high-fidelity PCR (see Materials and Methods) from DNA from a cdc13-1 strain, inserted into pTHA, and introduced into a cdc13Δ strain with URA3 at the telomere of chromosome VII-L. The cdc13Δ cells carrying pTHA-cdc13-1 grew about as well as the same strain carrying pTHA-CDC13 at 25°C and 30°C, but had slower growth at higher temperatures (data not shown). At 37°C, cdc13Δ/pTHA-cdc13-1 took 6–7 days to form a robust colony on complete medium lacking leucine, whereas the same strain carrying pTHA-CDC13 took 3 days. The ability of cdc13Δ/pTHA-cdc13-1 cells to grow at higher temperatures than cdc13-1 cells presumably was due to overexpression of the mutant protein.
The fraction of FOAR colonies for cdc13Δ/pTHA-cdc13-1 and for three control strains, cdc13Δ/pTHA-CDC13, CDC13/pTHA, and CDC13/pTHA-CDC13, was determined at 25°C, 30°C, and 37°C (Fig. 5). At both 25°C and 30°C, cdc13Δ/pTHA-cdc13-1 produced a similar fraction of FOAR colonies as the three controls, although, as noted previously, values for 25°C cells varied considerably among individual colonies. However, at 37°C, the fraction of FOAR cells for cdc13Δ/pTHA-cdc13-1 was 10−4, ≈1000-fold less than for the three controls. The C1–3A strand of telomeric DNA, which is the same strand from which URA3 is transcribed, is degraded in cdc13-1 cells arrested by growth at restrictive temperatures (25). However, the loss of TPE seen in cells grown at semipermissive temperatures cannot be attributed to degradation of the telomere-linked URA3 gene because cells in which URA3 is degraded will die, experience cell cycle arrest, or yield FOAR colonies, whereas loss of TPE generates FOA-sensitive colonies. However, C-strand degradation may explain why only 36% of cdc13Δ/pTHA-cdc13-1 produced colonies at 37°C on complete medium lacking leucine.
DISCUSSION
These experiments demonstrate that the essential Saccharomyces protein Cdc13p binds ss TG1–3 DNA in vitro (Fig. 1). There are two potential telomeric substrates for Cdc13p. Throughout most of the cell cycle, chromosomal telomeres are postulated to have short ss TG1–3 tails. Cdc13p bound efficiently to molecules with 9 bp of duplex DNA and a 10-base ss TG1–3 tail (Fig. 3), a substrate that mimics the expected structure of telomeres during most of the cell cycle. In addition, after conventional, semiconservative replication of telomeric DNA, yeast telomeres transiently acquire long ss TG1–3 tails (6). Cdc13p binding to a 22-base ss TG1–3 oligonucleotide was competed efficiently by a 43-base ss TG1–3 oligonucleotide and by a circular phagemid containing 270 bases of ss TG1–3 DNA (Fig. 4B), suggesting that Cdc13p can also bind the transient ss TG1–3 tails generated during telomere replication.
Although there is no direct evidence that Cdc13p binds telomeres in vivo, there are genetic data consistent with this possibility. Cells overexpressing a mutant form of Cdc13p, Cdc13-1p, had normal TPE at 25°C and 30°C (Fig. 5) and essentially wild-type growth rates (data not shown). However, at 37°C, the highest temperature at which cells overexpressing Cdc13-1p continued to grow, TPE was reduced (Fig. 5). Together with its binding specificity, these data suggest that Cdc13p binds to the constitutive ss TG1–3 tails thought to be present at most times in the cell cycle. Genetic data suggest that yeast has a protein important for TPE whose binding is restricted to telomeres and that is present in limiting amounts (20). Because Cdc13p binding activity was detected in yeast extracts only when it was overproduced (Fig. 1B, lane 5), Cdc13p, like this hypothetical telomere-limited binding protein, is probably not an abundant protein. Although its binding specificity suggests that Cdc13p may be the functional counterpart of the ciliate telomere binding proteins (16), it had no obvious sequence similarity to the ciliate proteins (31, 32) and, unlike the ciliate proteins, its binding to telomeric DNA was not stable to high salt (unpublished results).
Cells lacking Cdc13p sustain degradation of the C strand of telomeric DNA, which generates ss regions much more extensive than those detected in wild-type cells (25). These data suggest that in vivo Cdc13p limits the C-strand degradation that occurs at the end of the S phase (23), consistent with the possibility that Cdc13p also binds the transient ss TG1–3 tails generated as intermediates in telomere replication. If Cdc13p binding to the transient ss TG1–3 tails limits C-strand degradation, a plausible model is that Cdc13p must dissociate from telomeres or alter its conformation to allow the limited C-strand degradation that occurs after conventional replication is complete. Cdc13p has eight potential recognition sites for the major yeast cyclin-dependent kinase, CDC28 (A. Smith, unpublished results), raising the possibility that cell cycle-regulated phosphorylation controls Cdc13p binding to telomeric DNA.
Because a cell that loses a single telomere arrests via the RAD9 DNA damage-sensing cell cycle checkpoint, telomeres help the cell distinguish intact from damaged DNA (7). Single-stranded DNA has been proposed to be the signal to which the RAD9 checkpoint responds (25). Despite the likely presence of a constitutive ss tail, telomeres are not detected as damaged DNA (7). We speculate that Cdc13p masks the ss TG1–3 tails from the RAD9 pathway. This interpretation suggests that loss of Cdc13p could trigger a RAD9-mediated arrest in two ways, by unmasking the short, constitutive ss TG1–3 tails or by allowing extensive degradation of C-strand DNA after telomere replication.
Acknowledgments
We thank B. Garvik from L. Hartwell’s laboratory for strains, plasmids, and advice, and C. Lin from R. Reeder’s laboratory for pTHA. We also thank members of our laboratory for sequence analysis of CDC13 (A. Smith), for help with the figures (J. Mangahas and C. Lewis), and for their careful reading of the manuscript. This work was supported by grants from the National Institutes of Health.
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Abbreviations: ss, single-strand; TPE, telomere position effect; HA, hemagglutinin; YC, yeast complete; FOA, 5-fluoroorotic acid, FOAR, FOA resistant.
References
- 1.Zakian V A. Science. 1995;270:1601–1607. doi: 10.1126/science.270.5242.1601. [DOI] [PubMed] [Google Scholar]
- 2.Shampay J, Szostak J W, Blackburn E H. Nature (London) 1984;310:154–157. doi: 10.1038/310154a0. [DOI] [PubMed] [Google Scholar]
- 3.Oka Y, Shiota S, Nakai S, Nishida Y, Okubo S. Gene. 1980;10:301–306. doi: 10.1016/0378-1119(80)90150-x. [DOI] [PubMed] [Google Scholar]
- 4.Klobutcher L A, Swanton M T, Donini P, Prescott D M. Proc Natl Acad Sci USA. 1981;78:3015–3019. doi: 10.1073/pnas.78.5.3015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Henderson E R, Blackburn E H. Mol Cell Biol. 1989;9:345–348. doi: 10.1128/mcb.9.1.345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wellinger R J, Wolf A J, Zakian V A. Cell. 1993;72:51–60. doi: 10.1016/0092-8674(93)90049-v. [DOI] [PubMed] [Google Scholar]
- 7.Sandell L L, Zakian V A. Cell. 1993;75:729–739. doi: 10.1016/0092-8674(93)90493-a. [DOI] [PubMed] [Google Scholar]
- 8.Gottschling D E, Aparicio O M, Billington B L, Zakian V A. Cell. 1990;63:751–762. doi: 10.1016/0092-8674(90)90141-z. [DOI] [PubMed] [Google Scholar]
- 9.Wright J H, Gottschling D E, Zakian V A. Genes Dev. 1992;6:197–210. doi: 10.1101/gad.6.2.197. [DOI] [PubMed] [Google Scholar]
- 10.Conrad M N, Wright J H, Wolf A J, Zakian V A. Cell. 1990;63:739–750. doi: 10.1016/0092-8674(90)90140-a. [DOI] [PubMed] [Google Scholar]
- 11.Wright J H, Zakian V A. Nucleic Acids Res. 1995;23:1454–1460. doi: 10.1093/nar/23.9.1454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shore D, Nasmyth K. Cell. 1987;51:721–732. doi: 10.1016/0092-8674(87)90095-x. [DOI] [PubMed] [Google Scholar]
- 13.Buchman A R, Kimmerly W J, Rine J, Kornberg R D. Mol Cell Biol. 1988;8:210–225. doi: 10.1128/mcb.8.1.210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Giraldo R, Rhodes D. EMBO J. 1994;13:2411–2420. doi: 10.1002/j.1460-2075.1994.tb06526.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kurtz S, Shore D. Genes Dev. 1991;5:616–628. doi: 10.1101/gad.5.4.616. [DOI] [PubMed] [Google Scholar]
- 16.Gottschling D E, Zakian V A. Cell. 1986;47:195–205. doi: 10.1016/0092-8674(86)90442-3. [DOI] [PubMed] [Google Scholar]
- 17.Price C M. Mol Cell Biol. 1990;10:3421–3431. doi: 10.1128/mcb.10.7.3421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Price C M, Cech T R. Genes Dev. 1987;1:783–793. doi: 10.1101/gad.1.8.783. [DOI] [PubMed] [Google Scholar]
- 19.Lipps H J. Proc Natl Acad Sci USA. 1980;77:4104–4107. doi: 10.1073/pnas.77.7.4104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wiley E, Zakian V A. Genetics. 1995;139:67–79. doi: 10.1093/genetics/139.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cardenas M E, Bianchi A, de Lange T. Genes Dev. 1993;7:870–882. doi: 10.1101/gad.7.5.883. [DOI] [PubMed] [Google Scholar]
- 22.Wellinger R J, Wolf A J, Zakian V A. Mol Cell Biol. 1993;13:4057–4065. doi: 10.1128/mcb.13.7.4057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wellinger R J, Ethier K, Labrecque P, Zakian V A. Cell. 1996;85:423–433. doi: 10.1016/s0092-8674(00)81120-4. [DOI] [PubMed] [Google Scholar]
- 24.Weinert T A, Hartwell L H. Genetics. 1993;134:63–80. doi: 10.1093/genetics/134.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Garvik B, Carson M, Hartwell L. Mol Cell Biol. 1995;15:6128–6138. doi: 10.1128/mcb.15.11.6128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Runge K W, Zakian V A. Mol Cell Biol. 1989;9:1488–1497. doi: 10.1128/mcb.9.4.1488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lin J J, Zakian V A. Nucleic Acids Res. 1994;22:4906–4913. doi: 10.1093/nar/22.23.4906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sikorski R S, Hieter P. Genetics. 1989;122:19–27. doi: 10.1093/genetics/122.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Baudin A, Ozier-Kalogeropoulos O, Denouel A, Lacroute F, Cullin C. Nucleic Acids Res. 1993;21:3329–3330. doi: 10.1093/nar/21.14.3329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Boeke J D, LaCroute F, Fink G R. Mol Gen Genet. 1984;197:345–346. doi: 10.1007/BF00330984. [DOI] [PubMed] [Google Scholar]
- 31.Gray J T, Celander D W, Price C M, Cech T R. Cell. 1991;67:807–814. doi: 10.1016/0092-8674(91)90075-a. [DOI] [PubMed] [Google Scholar]
- 32.Hicke B J, Celander D W, MacDonald G H, Price C M, Cech T R. Proc Natl Acad Sci USA. 1990;87:1481–1485. doi: 10.1073/pnas.87.4.1481. [DOI] [PMC free article] [PubMed] [Google Scholar]