

Abstract
We show that an electric field can drive single-stranded RNA and DNA molecules through a 2.6-nm diameter ion channel in a lipid bilayer membrane. Because the channel diameter can accommodate only a single strand of RNA or DNA, each polymer traverses the membrane as an extended chain that partially blocks the channel. The passage of each molecule is detected as a transient decrease of ionic current whose duration is proportional to polymer length. Channel blockades can therefore be used to measure polynucleotide length. With further improvements, the method could in principle provide direct, high-speed detection of the sequence of bases in single molecules of DNA or RNA.
Measurement of ionic current passing through single ion channels in biological membranes or planar lipid bilayers is routine in neurobiology and biophysics (1, 2). Although most such channels undergo voltage- or ligand-dependent gating (3, 4), several relatively large ion channels, including Staphylococcus aureus α-hemolysin (5, 6) and the mitochondrial voltage-dependent anion channel (7, 8), can remain open for extended periods, thereby allowing continuous ionic current to flow across a lipid bilayer (9, 10). We reasoned that a transmembrane voltage applied across a continuously open channel of appropriate size should draw polyanionic DNA or RNA molecules through the channel as extended linear chains whose presence would detectably reduce or block normal ionic flow. Such blockages should make it possible to use single channel recordings to characterize the length and, possibly, other characteristics of the polymer.
Random coil polymers have previously been used to investigate channel pore dimensions (8) and single channel current recordings show that small poly(ethylene glycol) polymers partition into channels formed by alamethicin (11) or S. aureus α-hemolysin (12, 13). Recent investigations show that proteins can also traverse a lipid bilayer by moving through protein translocating channels, presumably as unfolded, extended chains (14, 15, 16, 17, 18). While translocating a protein, such channels are electrically silent; they regain their ion permeability only after the translocating polypeptide has been cleared from the channel (18). The ionic conductance of large nuclear pore complexes is similarly reduced during translocation of transcription factors, although in this case the macromolecules are assumed to move through the nuclear pore in a folded state (19, 20).
To determine whether nucleic acid polymers can be detected by single channel measurements, S. aureus α-hemolysin was used to form a single channel across a lipid bilayer separating two buffer-filled compartments (21). The α-hemolysin monomers spontaneously insert into lipid bilayers (9) and assemble to form heptameric transmembrane channels (6) that are 2.6 nm in diameter (L. Song, M. R. Hobaugh, C. Shustak, S. Cheley, H. Bayley, and J. E. Gouaux, personal communication). Channels with this dimension should be sufficiently large to accommodate the diameter of an extended, single-stranded polynucleotide.
MATERIALS AND METHODS
Conductance Measurements.
A solvent-free bilayer membrane of diphytanoyl-PC (Avanti Polar Lipids) was formed across a ≈0.1-mm-diameter orifice in a 25-μm thick Teflon partition separating two buffer filled compartments (ref. 23, patterned after ref. 21) each containing 1.5 ml of 1 M KCl, 5 mM Hepes (pH 7.5). α-Hemolysin channels were reconstituted into the membrane by adding less than 1 μg of α-hemolysin to the compartment we designated as cis. A single channel usually formed within 5 min. To prevent further channel incorporation, the cis compartment was flushed extensively with fresh buffer solution. The current was converted to voltage and amplified by either an Axopatch 200A (Axon Instruments, Foster City, CA) or a Dagan 3900A patch clamp amplifier (Dagan Instruments, Minneapolis). The signal was filtered at less than 3/8 the sampling rate, which was typically 64,000 samples/sec, using a Frequency Devices (Haverhill, MA) low pass Bessel filter. Unless stated otherwise, the data was digitized using a National Instruments (Austin, TX) AT-MIO-16-X 16-bit board using programs written in lab windows/CVI (National Instruments). The bilayer apparatus was shielded from electric and magnetic noise sources using a μ-metal box (Amuneal, Philadelphia).
Polynucleotides.
For most of our experiments we used polyuridylic acid (poly[U]) because this homo-polynucleotide has minimal secondary structure or base pairing (24) that might interfere with translocation of the polymer. Fractions of the poly[U] (Sigma), prepared with polynucleotide phosphorylase, were size selected by elution from agarose after fractionation by electrophoresis in the presence of formaldehyde. All DNA was synthesized using standard phosphoramidite methods. Although the sequence of the 150-nt DNA (CTCACCTATC CTTCCACTCA TTTTCCTTAA CCATTTCATT CACCCATCTC TTCACTCCAT CTATCACCTC CATACATACC CTCCATATTA CACTCCCATC ACTATCATTA TCTACATCCA TTACATCACT ACTCCTCACA CTACCATACC) was designed to minimize base pairing expected in the nondenaturing conditions we used, this polymer exhibited a larger proportion of long-lived blockades than did the homopolymers. Many of these blockades required clearing by brief polarity reversal. The 120-nt DNA for competitive PCR contained the same sequence as the 150-nt molecule but lacked residues 97–126. Double-stranded DNA, designed to be amplified by PCR using the same primers as the 150-nt single-stranded DNA, was made by mixing and annealing a synthetic 100-nt sense strand with its 100-nt antisense strand. The 100-nt sense strand contained the same sequence as the 150-nt molecule but lacked residues 49–98.
PCR Analysis and Blockade Counts.
Competitive PCR analysis (25) was performed after amplification (26) optimized to avoid the plateau phase. To prevent DNA loss by adsorption to the trans chamber walls, 55 μg/ml of carrier poly[U] was added to the trans chamber when DNA was to be harvested for PCR analysis. The PCR products were resolved on a 15% (total monomer) acrylamide gel and stained with SYBR Green I (Molecular Probes) for densitometry. In experiments with 1–8 channels, the number of blockades in peak 2 + peak 3 was determined by counting individual blockades during several 5-min sampling periods throughout the experiment; in the experiment with 40 channels, the number of blockades was estimated by multiplying (peak 2 + peak 3 rate observed with one channel) × (number of channels in the membrane) × (time of experimental voltage application).
RESULTS AND DISCUSSION
In the absence of polymer, applying a potential of −120 mV (cis side negative) resulted in single channel currents that were free of transients (Fig. 1). Following the addition of poly[U] to the cis chamber, numerous short-lived current blockades occurred. Typically, the magnitude of the blockades was large (the current was reduced by 85–100%) and, depending on polymer length, the blockades lasted from several hundred to several thousand microseconds. For a given polymer size, the total number of transient blockades was directly proportional to polymer molar concentration (not shown). Virtually no blockades were observed in the absence of poly[U] (Fig. 1) or if the potential across the bilayer was reversed (not shown). Occasionally, after adding polymer, long-lived blockades of several seconds or more occurred which could be cleared by briefly reducing the voltage to 0, or by reversing the polarity of the potential. We assume that these indefinitely long blockages were produced when a polynucleotide entered but could not completely traverse the channel because of entangling secondary structure. The millisecond time scale blockades were also seen in similar experiments with poly[A], poly[C], poly[dT], poly[dC], as well as with single-stranded synthetic DNA composed of 150 nt of poly[dA, dT, dC] (see Materials and Methods).
Figure 1.
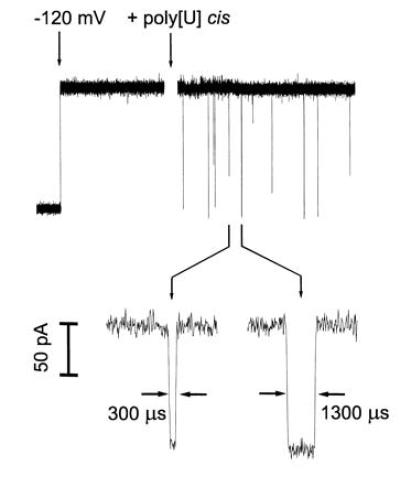
Oligomers of poly[U] caused transient blockades in the α-hemolysin single channel current. At the first arrow, a potential of −120 mV was applied across the membrane (cis side negative). This voltage caused a continuous current of −120 pA to flow. At the second arrow, poly[U] of mean length 210 bases was stirred into the cis compartment to a final concentration of 0.1 mg/ml. The polynucleotides caused short-lived current blockades. The Inset (expanded time scale) shows two typical blockades with lifetimes of 300 and 1300 μs. For the purposes of illustration, the low time resolution current recordings (a total of 4 sec is shown here) were digitally filtered using an exponential smoothing moving average algorithm.
If the blockades were caused by the transport of extended linear polynucleotides through the channel, the blockade lifetimes should be proportional to the length of the polymer and inversely proportional to the applied potential. A histogram of the current blockade lifetimes for 210-nt-long poly[U] shows that the blockades cluster into three distinct peaks (Fig. 2). Similar histograms for other size selected poly[U]s demonstrated that the mean lifetimes for peaks 2 and 3 were related to polymer length and inversely related to the applied potential (Fig. 3). On the other hand, the numerous fast blockades in peak 1 were independent of polymer length (Fig. 3a) or applied potential. We attribute these fast peak 1 blockades to polymers that collided with the channel, or partially entered but failed to traverse the channel. Thus, only the blockades in peaks 2 and 3, whose lifetimes were proportional to polymer length, appeared to be the result of polymers that traversed the channel.
Figure 2.
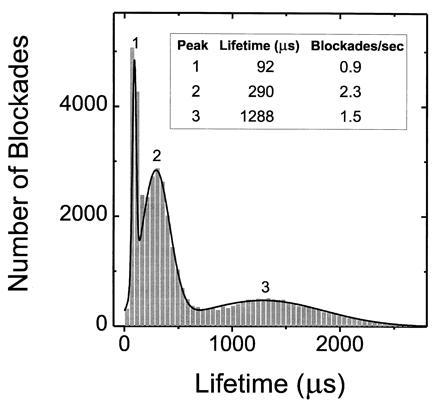
The characteristic lifetimes of channel blockades caused by poly[U] (0.1 mg/ml; mean length, 210 nt) fell within three well-defined peaks. The mean lifetimes corresponding to the three peaks were determined by fitting the sum of three Gaussians to the data.
Figure 3.
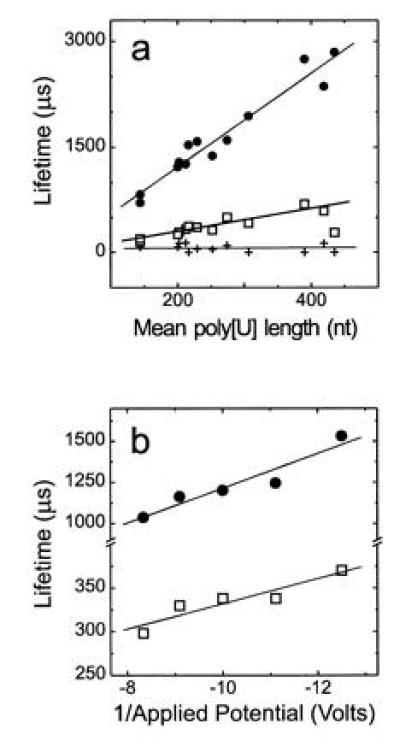
Poly[U]-induced channel blockade lifetimes were proportional to (a) mean polymer length and (b) inversely proportional to applied voltage. The plots show lifetimes for (a) peaks 1 (+), 2 (□), and 3 (•) in experiments using V = −120 mV with 13 different size selected poly[U]s and (b) for peaks 2 (□), and 3 (•) with poly[U] of mean length 215 nt at the indicated voltages. Although the peak 1 lifetime appeared to be independent of the applied voltage (data not shown), because this lifetime (≈100 μs) was barely a factor of 2 greater than the time resolution of our system, a slight voltage dependence could not be ruled out.
Although the length dependency for peaks 2 and 3 was consistent with the notion that a polynucleotide traversed the channel as an extended linear chain, we were surprised that passage through the channel of relatively monodisperse, unbranched polymers would produce blockades whose lifetimes fell under two distinct peaks that contained approximately equal numbers of events. An explanation for the presence of these two peaks is that the characteristic times for polymer transit in the 3′ to 5′ direction may differ from that in the 5′ to 3′ direction. If true, the physical basis for such a difference remains to be explained.
The 2.6-nm diameter of the α-hemolysin channel is apparently too narrow to allow passage of double-stranded polynucleotides because blunt-ended double-stranded DNA produced only the rapid blockades similar to peak 1 events. We also tested mixtures containing equal concentrations of diverse length poly[A] with poly[U] or diverse length poly[dC] with poly[dG]. Such preparations would contain many base paired regions surrounded by unpaired single-stranded ends that might enter the channels but stop at a region of paired bases. As expected, these preparations produced indefinitely long blockades that required voltage polarity reversal to be cleared.
To demonstrate that blockades were attributable to DNA traversing the channels, we performed experiments to compare the number of blockades observed in peaks 2 and 3 with the number of single-stranded molecules that actually traversed the bilayer from the cis to the trans chamber. Taking advantage of our observation that double-stranded DNA could not traverse the α-hemolysin channel, we used double-stranded DNA as an internal control for inadvertent contamination of the trans chamber. The results (Table 1 and Fig. 4) showed that the amount of single-stranded DNA in the trans chamber approximated the amount we estimated should have accumulated. The single-stranded DNA found in the trans chamber could not be the result of contamination by aerosol or leaks from the cis chamber because such contamination would have yielded approximately the same ratio of single-stranded 150-nt DNA to double-stranded 100-nt DNA in the trans chamber as in the cis chamber. This was clearly not the case (Fig. 4, lanes 1, 2). Because the number of single-stranded molecules that accumulated in the trans chamber depended on the number of channels in the membrane and was proportional to the total number of peak 2 and peak 3 blockades, significant movement of single-stranded DNA through the lipid bilayer itself, rather than through the channels, could be ruled out.
Table 1.
DNA transport through α-hemolysin channels
| No. of channels | Blockades/min | Molecules/min |
|---|---|---|
| 35 | 1300 | 1250 |
| 8 | 220 | 280 |
| 6 | 150 | 80 |
The number of blockades/min is compared with the number of molecules/min that moved to the trans chamber during experiments in which the cis chamber contained 0.27 mg/ml of 150-nt single-stranded DNA and an equivalent number of 100-nt strands of double-stranded DNA. Multiple channels were used in these experiments to show the proportionality between channel number and DNA movement. Because frequent current reversals were required during the experiments (see Materials and Methods), the precision of the blockade rate calculation is limited to ±30%. The number of molecules transported to the trans chamber per minute was calculated from the total number of molecules found in the trans chamber at the end of each experiment, as measured by competitive PCR analysis which can reliably discriminate 2-fold differences in copy number (25).
Figure 4.
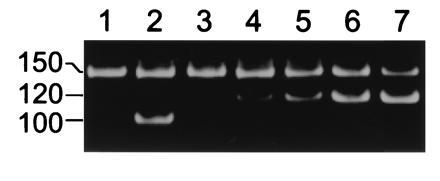
Single-stranded DNA traversed the α-hemolysin pore. The PCR-amplified product (lane 1) of a 40 μl sample of the trans chamber compared with (lane 2) the similarly amplified product of a 109-fold diluted sample of the cis chamber. All samples were taken following an 8 channel experiment in which a mixture of 150-nt single-stranded DNA (see Materials and Methods) and 100-nt double-stranded DNA was added to the cis chamber. Material in the trans chamber sample reflects accumulation during application of −120 mV (cis chamber negative) for 42 min. Lanes 3-7: example of competitive PCR analysis (25) to show accumulation of single-stranded molecules in the trans chamber. The results show that the 40-μl sample used here contained ≈400 of the 150-nt single-stranded molecules. To each of five 40-μl samples of the trans chamber were mixed (lanes 3–7) 0, 100, 200, 400, or 800 molecules of a competing 120-nt DNA that was PCR amplified using the same primers as the 150-nt DNA. The position of the amplified 150-, 120-, and 100-nt polymers is shown at left. Lanes 1 and 3 show independently amplified 40-μl samples of the same trans chamber.
While we have not yet established the quantitative relation between polymer molar concentration and blockade rate for a wide range of polymer sizes, we have observed that the number of blockades/minute produced by short (≈200 nt) polymers was significantly greater than the number produced by equimolar concentrations of longer (≈1000 nt) polymers. These observations suggested that the blockade rate could be used as an especially sensitive monitor of polynucleotide hydrolysis. For example, hydrolysis of long polymers by ribonuclease would increase blockade rate by simultaneously increasing the polymer molar concentration and decreasing the polymers’ sizes. Indeed, upon addition of ribonuclease A to a cis compartment containing poly[U] there was a striking increase in blockade rate until the polymer fragment sizes fell below the temporal resolution of the instrumentation, whereupon the detected blockade rate decreased until no further blockades were observed (Fig. 5). Upon addition of fresh poly[U] to the cis chamber, a burst of blockades was again observed (not shown), demonstrating that the channel was still capable of detecting polymer induced blockades and that active ribonuclease remained in the cis chamber. Ribonuclease had no effect when poly[U] was replaced by poly[A], which is not a substrate for ribonuclease A. Thus, the initial increase in blockade rate cannot be attributed to the action of RNase on the bilayer or the channel. We conclude that analysis of blockade rate can provide a novel and rapid measure of enzymatic activity.
Figure 5.
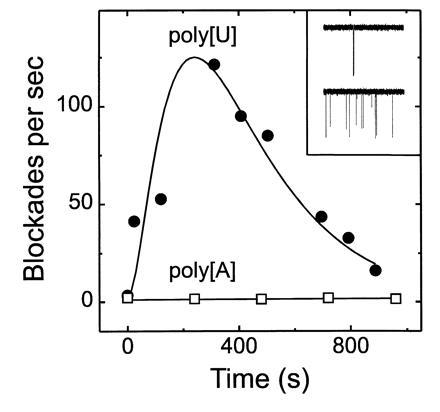
Enzymatic hydrolysis of poly[U] caused a transient increase in the rate of oligonucleotide-induced channel blockades. In two separate experiments, at time 0, ribonuclease A (bovine pancreatic RNase Type XII-A; Sigma or Worthington) was added to the cis chamber containing either 0.13 mg/ml poly[U] (•) (mean length, 1100 nt) or 0.27 mg/ml poly[A] (□) (mean length, 20,000 nt; Fluka). The final ribonuclease concentrations were 0.04 mg/ml (poly[U] experiment) or 0.13 mg/ml (poly[A] experiment). Blockade rates were calculated from single channel recordings. The Inset shows (null)/1;3-sec sample traces for the poly[U] experiment before (Upper) and ≈30 sec after (Lower) addition of ribonuclease. Similar experiments with a poly[U] sample of greater length (3000 nt) yielded results comparable to those obtained with the 1100-nt poly[U]. The current was digitized using a SONY/Dagan DAT recorder at 48,000 points/sec.
The discovery that a polynucleotide can easily be drawn through a single, well-defined channel provides a simple system for measuring the driven motion of polymers through constraints, a basic problem underpinning many biological processes, as well as gel electrophoresis (27). Recent theories of polymer translocation through a single membrane pore (28) and through multiple pores (22) could easily be tested in this system.
The realization that single channel measurements can be used to determine the lengths of individual RNA or DNA chains suggests that other characteristics of a polynucleotide could also be measured as the molecule traverses the channel. In principle, a single purine or pyrimidine nucleotide passing through the limiting aperture of a channel should block the ionic current in a manner that reflects the molecular size and chemical properties of each nucleotide in a polynucleotide. If so, it may be possible to use single channel recordings to directly determine the sequence of a polynucleotide. For this to be possible, at least five conditions must be met: each nucleotide must produce a characteristic transient current blockage,¶ the limiting aperture of the channel must have appropriate dimensions to reflect the presence of only one nucleotide at a time, the time resolution of ionic flow measurement must exceed the rate of nucleotide movement through the channel, backward movement of the polynucleotide must be minimal, and the channel and membrane must be sufficiently robust to withstand whatever temperature and chemical treatments are required to eliminate interference from polynucleotide secondary structure.
Acknowledgments
We thank H. Bayley for the generous gift of α-hemolysin and H. Berg, J. Golovchenko, and D. Nelson for their encouragement and suggestions. We also thank A. Schroder, R. Li, and S. Lee for technical assistance. This research was supported by grants from the National Aeronautics and Space Administration (D.W.D.), the National Science Foundation (D.B.), the National Institutes of Health (D.W.D.), and a National Academy of Sciences/National Research Council Research Associateship (J.J.K.).
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
In closely related experiments, F. Roth, R. Baldarelli, and G. M. Church have modeled the movement of DNA through the pore formed by the LamB receptor. Their model shows that DNA can cause conductance changes that should be sensitive to the structure of the different bases (personal communication).
References
- 1.Neher E, Sakmann B. Nature (London) 1976;260:799–802. doi: 10.1038/260799a0. [DOI] [PubMed] [Google Scholar]
- 2.Hille B. Ionic Channels of Excitable Membranes. 2nd Ed. Sunderland, MA: Sinauer; 1992. [Google Scholar]
- 3.Sigworth F J. Q Rev Biophys. 1994;27:1–40. doi: 10.1017/s0033583500002894. [DOI] [PubMed] [Google Scholar]
- 4.Keynes R D. Q Rev Biophys. 1994;27:339–434. doi: 10.1017/s0033583500003097. [DOI] [PubMed] [Google Scholar]
- 5.Bhakdi S, Tranum-Jensen J. Microbiol Rev. 1991;55:733–751. doi: 10.1128/mr.55.4.733-751.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gouaux J E, Braha O, Hobaugh M R, Song L, Cheley S, Shustak C, Bayley H. Proc Natl Acad Sci USA. 1994;91:12828–12831. doi: 10.1073/pnas.91.26.12828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schein S J, Colombini M, Finkelstein A. J Membr Biol. 1976;30:99–120. doi: 10.1007/BF01869662. [DOI] [PubMed] [Google Scholar]
- 8.Colombini M. J Membr Biol. 1980;53:79–84. [Google Scholar]
- 9.Menestrina G. J Membr Biol. 1986;90:177–190. doi: 10.1007/BF01869935. [DOI] [PubMed] [Google Scholar]
- 10.Colombini M. J Membr Biol. 1989;111:103–111. doi: 10.1007/BF01871775. [DOI] [PubMed] [Google Scholar]
- 11.Bezrukov S M, Vodyanoy I, Parsegian V A. Nature (London) 1994;370:279–281. doi: 10.1038/370279a0. [DOI] [PubMed] [Google Scholar]
- 12.Krasilnikov O V, Sabirov R Z, Ternovsky V I, Merzliak P G, Muratkodjaev J N. FEMS Microbiol Immun. 1992;105:93–100. doi: 10.1111/j.1574-6968.1992.tb05891.x. [DOI] [PubMed] [Google Scholar]
- 13.Korchev Y E, Bashford C L, Alder G M, Kasianowicz J J, Pasternak C A. J Membr Biol. 1995;147:233–239. doi: 10.1007/BF00234521. [DOI] [PubMed] [Google Scholar]
- 14.Henry J P, Chich J F, Goldschmidt D, Thieffry M. J Membr Biol. 1989;112:139–147. doi: 10.1007/BF01871275. [DOI] [PubMed] [Google Scholar]
- 15.Brundage L, Hendrick J P, Schiebel E, Driessen A J M, Wickner W. Cell. 1990;62:649–657. doi: 10.1016/0092-8674(90)90111-q. [DOI] [PubMed] [Google Scholar]
- 16.Akimaru J, Matsuyama S I, Tokuda H, Mizushima S. Proc Natl Acad Sci USA. 1991;88:6545–6549. doi: 10.1073/pnas.88.15.6545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goerlich D, Rapoport T A. Cell. 1993;75:615–630. doi: 10.1016/0092-8674(93)90483-7. [DOI] [PubMed] [Google Scholar]
- 18.Simon S M, Blobel G. Cell. 1991;65:371–380. doi: 10.1016/0092-8674(91)90455-8. [DOI] [PubMed] [Google Scholar]
- 19.Bustamante J O, Hanover J A, Leipins A. J Membr Biol. 1995;146:239–251. doi: 10.1007/BF00233944. [DOI] [PubMed] [Google Scholar]
- 20.Bustamante J O, Oberleithner H, Hanover J A, Liepins A. J Membr Biol. 1995;146:253–261. doi: 10.1007/BF00233945. [DOI] [PubMed] [Google Scholar]
- 21.Montal M, Mueller P. Proc Natl Acad Sci USA. 1972;69:3561–3566. doi: 10.1073/pnas.69.12.3561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lee N, Obukhov S. J Phys II. 1996;6:195–204. [Google Scholar]
- 23.Kasianowicz J J, Bezrukov S M. Biophys J. 1995;69:94–105. doi: 10.1016/S0006-3495(95)79879-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cantor C R, Schimmel P R. Biophysical Chemistry: The Behavior of Biological Macromolecules. New York: Freeman; 1995. Part 3. [Google Scholar]
- 25.Gilliland G, Perrin S, Bunn H F. In: PCR Protocols. Innis M A, Gelfand D H, Sninsky J J, White T J, editors. San Diego: Academic; 1990. pp. 60–69. [Google Scholar]
- 26.Ruano G, Fenton W, Kidd K K. Nucleic Acids Res. 1989;17:5407. doi: 10.1093/nar/17.13.5407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zimm B H, Levene S D. Q Rev Biophys. 1992;25:171–204. doi: 10.1017/s0033583500004662. [DOI] [PubMed] [Google Scholar]
- 28.Sung W, Park P J. Phys Rev Lett. 1996;77:783–786. doi: 10.1103/PhysRevLett.77.783. [DOI] [PubMed] [Google Scholar]