

Abstract
Little is known about the signals that govern the network of meristem and organ identity genes that control flower development. In Arabidopsis, we can induce a heterochronic switch from flower to shoot development, a process known as floral meristem reversion, by manipulating photoperiod in the floral homeotic mutant agamous and in plants heterozygous for the meristem identity gene leafy. The transformation from flower to shoot meristem is suppressed by hy1, a mutation blocking phytochrome activity, by spindly, a mutation that activates basal gibberellin signal transduction in a hormone independent manner, or by the exogenous application of gibberellins. We propose that LFY and AG play an important role in the maintenance of flower meristem identity and that floral meristem reversion in heterozygous lfy and in ag flowers is regulated by a phytochrome and gibberellin signal transduction cascade.
Keywords: gibberellins, phytochrome, floral reversion
Plant growth and morphogenesis is controlled by meristems, organized tissues containing pluripotent stem cells whose identities and activities are regulated by intrinsic and environmental signals. In Arabidopsis, the shoot apical meristem undergoes two phases, vegetative and inflorescence; both phases are characterized by reiterative and indeterminate patterns of growth and organogenesis (1). The vegetative meristem produces a compact rosette consisting of a short stem and a variable number of leaves. By contrast, the inflorescence meristem produces an elongated stem punctuated by narrow cauline leaves, lateral secondary shoots, and flowers that are derived from the flanks of the inflorescence meristem. Although closely related spatially and by cell lineage to the inflorescence meristem, the floral meristem proceeds along a determinate developmental pathway producing four compact whorls of organs (four sepals, four petals, six stamens, and two carpels). At the completion of organogenesis, the floral meristem is thought to be depleted or its activity is suppressed (2, 3). The transition from vegetative to inflorescence shoot meristem is controlled by environmental signals including photoperiod and temperature (4, 5, 6), by intrinsic growth regulators such as the gibberellins (6, 7, 8), and by a system of flowering time genes (9). The inflorescence meristem in turn produces an indeterminate number of floral meristems. Genetic and molecular studies have shown that the establishment of floral meristem identity is governed by a network of genes including APETALA1 (AP1), APETALA2 (AP2), CAULIFLOWER (CAL), CLAVATA1 (CLV1), CLAVATA3 (CLV3), and LEAFY (LFY) (3, 10, 11, 12, 13, 14, 15, 16, 17, 18), and several models have been proposed to explain how these genes function together (17, 19, 20). By comparison, much less is known about the signals and genes required for the maintenance of flower meristem identity.
Recently several genes have been implicated in the maintenance of flower meristem identity including LFY and AGAMOUS (AG) (3, 15, 17, 21, 22, 23). The LFY gene encodes a novel polypeptide that is reported to have DNA binding activity in vitro (20). AG encodes a protein that belongs to the evolutionarily conserved MADS domain family of eukaryotic transcription factors (24, 25). However, little is known about how these proteins govern meristem identity at the cellular and molecular levels. Previous studies suggest that the maintenance of flower meristem identity in lfy and in ag mutants is controlled by photoperiod and by one class of plant growth regulators, the gibberellins (12, 15, 21). Under short day (SD) photoperiod heterozygous lfy-6 and homozygous ag-1 flowers display a heterochronic transformation from flower to inflorescence shoot meristem—a novel developmental switch called floral meristem reversion (21). We have used the floral meristem reversion process to begin exploring the photo and hormonal control of flower meristem identity in Arabidopsis.
MATERIALS AND METHODS
Plant Material.
Landsberg-erecta (L-er) was used as wild-type flower control. ag-1 (L-er) and hy1-1 (L-er) were provided by Maarten Koornneef (Wageningen Agricultural University, Wageningen, The Netherlands). lfy-6 (L-er) was provided by Detlef Weigel (Salk Institute, La Jolla, CA). The lfy-6 allele is a well-characterized null mutant (14, 16). spy-2 and spy-3 (Col) (26) were provided by Neil Olszewski (University of Minnesota, Minneapolis).
Plant Growth Conditions.
Plants were grown under a mixture of cool white fluorescent (Sylvania CW/VHO) and incandescent lights (Phillips) in a Conviron E15 chamber (Controlled Environments, Asheville, NC) in a 1:1:1 mixture containing vermiculite/perlite/peat moss. Long day (LD) growth conditions consist of 16 h light, 150–180 μmol·m−2·s−1, and 8 h darkness. SD conditions were 9 h light, 150–180 μmol·m−2·s−1, and 15 h darkness. SD double incident light [SD(D)] conditions consist of 9 h light, 300 μmol·m−2·s−1. Plants were watered with a 1/4 strength Peter’s solution (Grace-Sierra, Milpitas, CA). For night-break experiments, red light was produced by using Sylvania cool white fluorescent lamps wrapped in a Lee HT026 filter (Lee Colortran, Burbank, CA). The transmission of light at wavelengths less than 580 nm was less than 0.5%. For gibberellin spray experiments, exogenous application of gibberellin A3 (GA3) (Sigma) and gibberellin A4+7 (GA4+7) was performed as described by Wilson et al. (8). GA4+7 was kindly provided by Phil Grau (Abbott Laboratories).
Analysis of Floral Reversion.
Plants segregating for lfy-6 and ag-1 were grown under SD(D) light conditions as described previously and were scored for the presence of ectopic shoots or swollen carpels by position from basipetal to acropetal on the primary inflorescence. For lfy-6, 31 of 60 plants displayed a strong floral reversion phenotype. All 31 plants were confirmed heterozygous by F1 analysis. Thirteen plants were confirmed as homozygous wild type for LFY. The remaining 17 plants were homozygous lfy-6 and did not produce flowers or seed. For ag-1, 16 plants were homozygous ag-1, and all produced flowers displaying floral meristem reversion. To generate lfy-6 spy mutants, homozygous lfy-6 was crossed with spy-2 and spy-3. F2 seedlings homozygous for spy were selected by germination on 1.2 × 10−4 M paclobutrazol (26), washed extensively with H2O, transplanted to soil, and grown under SD(D) conditions.
Structural Analysis Using Light Microscopy.
Flowers were fixed overnight in FAA (3.7% formaldehyde/5% acetic acid/50% ethanol), dehydrated in a graded ethanol series, infiltrated, and embedded in plastic resin. Serial 2 μm longitudinal sections were stained with toluidine blue-basic fuchsin. Images were obtained by using bright-field optics.
In Situ Hybridization Analysis.
In situ hybridizations were performed by using a digoxygenin-labeled AG RNA probe (24) as described (27) with the exception that the acetylation step was omitted. Hybridization was detected using a modified Genius (Boehringer Mannheim) procedure (G. N. Drews, personal communication). Images were photographed using dark-field optics.
Image Processing.
All images were scanned and digitized using a Polaroid Sprintscan 35 (Polaroid, Cambridge, MA). Color balance and brightness were adjusted using photoshop 3.0.1 (Adobe Systems, Mountain View, CA). Printed images were generated using a Codonics NP1600 printer (Codonics, Middleburg Heights, OH).
RESULTS
Transforming Flowers into Shoots.
Homozygous lfy mutants are characterized by a partial and conditional block in the establishment of the flower meristem (12, 15, 16). Under LD photoperiod, early-arising basipetal flowers are indeterminate and shoot-like, whereas the later-arising acropetal flowers are determinate. As shown in Fig. 1A, a homozygous acropetal lfy-6 flower is distinguished by the production of sepals and carpels and by the absence of petals and stamens when grown under LDs. By contrast, lfy-6 flower development is strongly repressed under SD conditions. Determinate floral buds are replaced by indeterminate shoots that continuously produce leaf-like organs (Fig. 1B).
Figure 1.

Photoperiod controls floral meristem activity in lfy-6 and ag-1 flowers. (A) LD homozygous lfy-6 flower. lfy-6 flowers show a basipetal to acropetal gradient of phenotypes (16). A typical acropetal LD lfy-6 flower is characterized by the production of sepals (s) and carpels (c) and the absence of petals and stamens. (B) SD homozygous lfy-6 flower. lfy flower development is suppressed under SDs (12, 15). A typical SD “flower” produces an indeterminate number of leaf-like organs (l). (C–F) SD heterozygous lfy-6 flower development. Heterozygous lfy-6 flowers produce a normal complement of organs (C) and are initially indistinguishable from wild type. Later, basipetal flowers are distinguished by a dramatically swollen ovary (so) (D). The swollen ovary often ruptures and a single flower-bearing ectopic shoot (es) emerges (E). The resulting shoot (es) produces several leaf-like organs, numerous flowers, displays internode elongation, and a spiral pattern of phyllotaxis (F). Flowers produced by the ectopic shoot appear either wild type or resemble ap2 mutant flowers. (G) LD ag-1 flower. ag-1 flowers are characterized by the transformation of stamens into petals and an indeterminate pattern of floral organogenesis (sepal, petal, petal)n. (H and I) SD ag-1 flower development. Basipetal SD ag-1 flowers initially produce several whorls of sepals and petals (H). Organogenesis then ceases and is replaced by the production of an ectopic shoot (es) that bears numerous flowers and displays internode elongation (I). (J–L) Ectopic shoot formation in basipetal SD heterozygous lfy-6 flowers. Longitudinal sections (2 μm) were prepared as described in Materials and Methods. An ectopic shoot (es) arises directly from the apical dome of the floral meristem (J). A magnified view of the ectopic shoot is shown in K. At this stage of development the ectopic shoot produces leaf-like organs (l). The ectopic shoot continues to grow and eventually fills the ovary (L). ov, The ovary wall. (Bar = 0.1 mm.) (M) Wild-type flower. Shown is a longitudinal section (2 μm) through a SD wild-type flower. Floral meristem activity is suppressed after carpel primordia are established. (Bar = 0.1 mm.) (N and O) Ectopic shoot formation in SD ag-1 flowers. A 2-μm longitudinal section through a SD basipetal ag-1 flower shows floral buds (fb) emerging from the apex of the floral meristem (N). A LD ag-1 flower is shown for comparison (O). (Bar = 0.1 mm.) (P–R) AG gene expression in SD heterozygous lfy-6 flowers. The spatial pattern of AG gene expression in SD heterozygous lfy-6 (P and Q) and wild-type flowers (R) was determined by in situ hybridization as described. The presence of AG transcripts is revealed by the orange staining. The flowers in P, Q, and R are at stages 3, 8, and 10, respectively (28). es, The ectopic shoot primordium. (Bar = 0.1 mm.)
We previously suggested that SD photoperiod dramatically alters flower development in heterozygous lfy-6 plants (21). SDs can induce heterozygous lfy-6 flowers to undergo a heterochronic transformation from flower to shoot development. Fig. 1C shows that initially the heterozygous lfy-6 floral meristem produces the normal complement of floral organs—four sepals, four petals, six stamens, and two carpels. Later, the ovary of the first 15–25 flowers along the primary inflorescence becomes swollen (Fig. 1D) and ruptures (Fig. 1E). Emerging from the center of the ruptured ovary is a single ectopic flower-bearing shoot (Fig. 1F), suggesting that the flower meristem has been reprogrammed and has undergone floral meristem reversion. The ectopic inflorescence is similar to a normal inflorescence, producing both leaf-like organs and flowers, displaying internode elongation, and a spiral pattern of phyllotaxis. Table 1 shows that this SD-induced switch in flower meristem identity is photoperiod dependent and not due to the decrease in total light per day because a compensatory increase in SD light intensity did not suppress the frequency of meristem transformations. By contrast, LD-grown heterozygous lfy-6 flowers remain phenotypically wild type (Table 1). These results show that lfy is haplo-insufficient under SDs and that the heterochronic switch from flower to shoot development is controlled by photoperiod.
Table 1.
Photo and hormonal control of floral reversion
| Genotype | Light and hormone treatment | Flowers per plant showing floral reversion* | No. of plants scored† |
|---|---|---|---|
| WT | LD | 0 | 14 |
| WT | SD | 0.4 (0.7) | 9 |
| lfy-6(+/−) | LD | 0 | 16 |
| lfy-6(+/−) | SD | 12 (2.2) | 16 |
| lfy-6(+/−) | SD(D) | 15 (6.5) | 31 |
| lfy-6(+/−) hy1-1 | SD(D) | 1.9 (2.7) | 30 |
| lfy-6(+/−) | SD + 0.5 hr red NB | 4.0 (5.4) | 24 |
| lfy-6(+/−) | SD + 2 hr red NB | 0.1 (0.3) | 16 |
| lfy-6(+/−) | SD(D) + 10−5 M GA3 | 0.1 (0.5) | 15 |
| lfy-6(+/−) spy-2 | SD(D) | 0 | 17 |
| lfy-6(+/−) spy-3 | SD(D) | 0 | 24 |
| ag-1 | LD | 0 | 15 |
| ag-1 | SD(D) | 16 (4.5) | 16 |
| ag-1 hy1-1 | SD(D) | 0 | 15 |
| ag-1 | SD + 0.5 hr red NB | 0.2 (0.4) | 6 |
| ag-1 | SD + 2 hr red NB | 0 | 4 |
| ag-1 | SD(D) + 10−5 M GA3 | 0 | 14 |
Growth conditions and light regimes are described in Materials and Methods. NB, night break (29).
Average number of flowers per primary inflorescence per plant displaying ectopic shoot development or swollen ovaries. Standard deviation values are given in parentheses. No floral meristem reversion was observed in hy1, spy-2, spy-3, or WT + 10−5 M GA3 control plants grown under SD conditions.
Heterozygous lfy-6 plants were confirmed by F1 analysis.
Photoperiod plays a similar role in controlling ag flower meristem identity (21). As shown in Fig. 1G, LD ag-1 flowers are distinguished by the homeotic transformation of stamens into petals and the replacement of carpels with a new flower primordium (30, 31). This pattern of organogenesis is repeated indeterminately, resulting in a (sepal, petal, petal)n pattern of development. By contrast, under SDs the ag-1 floral meristem can produce several whorls of floral organs and then revert to an inflorescence meristem and flower production (Fig. 1 H and 1I). This phenotype is distinct from those described for weak loss-of-function ag mutants or for transgenic AG antisense plants that display floral meristem indeterminacy but do not change identity (22, 23). As in the case of heterozygous lfy-6 flowers, the maintenance of ag-1 flower meristem identity is not influenced by light intensity but is controlled by photoperiod (Table 1). Thus both AG and LFY play key roles in the maintenance of floral meristem identity under SD conditions.
Ectopic Shoot Meristem Development.
Longitudinal sections through SD heterozygous lfy-6 flowers show that the ectopic shoot originates from cells located within the ovary at the apex of the floral meristem. Fig. 1 J–L shows that the cells in this region continue to proliferate despite the presence of a structurally normal septum, placenta, and ovules. At this level, the structure of the ectopic shoot meristem is indistinguishable from that of a normal shoot apex. A section through a wild-type flower is shown for comparison (Fig. 1M). Similarly, longitudinal sections through a SD ag-1 flower reveal that the ectopic shoot also emerges from the apex of the ag-1 floral meristem (Fig. 1N). A section through a LD ag-1 flower is shown for reference (Fig. 1O). One interpretation of these results is that under SDs the heterozygous lfy and the ag-1 flower meristems are reprogrammed during flower development. Alternatively, the ectopic shoots may arise from a cryptic population of cells that were never committed to flower development when the floral meristem was initially formed.
We carried out an AG in situ hybridization experiment to determine whether cells that give rise to the ectopic shoot in SD heterozygous lfy-6 flowers were initially floral in identity. In LD wild-type flowers, AG gene transcripts are undetectable in vegetative and inflorescence meristems and are restricted to developing stamens and carpels (24, 32). Fig. 1P shows that in young SD heterozygous lfy-6 flowers AG gene transcripts are detectable throughout the stamen and carpel primordia. AG transcripts are detectable continuously at the floral apex through later stages of flower development (K.D.J., data not shown) and are even detectable in the nascent ectopic shoot meristem (Fig. 1Q) but not at later stages of ectopic shoot development (K.D.J., data not shown). The distribution of AG gene transcripts in a SD wild-type flower is shown in Fig. 1R for comparison and is similar to that observed in LD flowers. We conclude that the ectopic shoot meristem is formed from cells that were originally floral in identity, and that the heterozygous lfy-6 floral meristem is not irreversibly determined but retains a photoperiod-dependent capacity for indeterminate shoot development.
Spatial Control of Ectopic Shoot Formation.
Previous studies have shown that homozygous lfy flower development varies by the position of each flower on the inflorescence stem (15, 16, 33). The strongest lfy phenotypes are found in early-arising basipetal flowers and decrease in severity acropetally or toward the apex of the inflorescence. Similarly, we found that the frequency of floral meristem reversion in SD heterozygous lfy-6 and in SD ag-1 plants is highest in basipetal flowers. Fig. 2A shows that 80–100% of heterozygous lfy-6 flowers from positions 1–15 on the primary inflorescence stem display swollen ovaries and produce ectopic shoots. The frequency of flower meristem reversion gradually declines to zero from positions 15–30. Fig. 2B shows a similar pattern for ag-1 flowers. Together these results show that floral meristem reversion is induced in both mutants in a significant number of flowers and with a high degree of certainty. Moreover, these results suggest that there is a progressive acropetal decrease in the requirement for LFY and AG activities for the maintenance of floral meristem identity.
Figure 2.
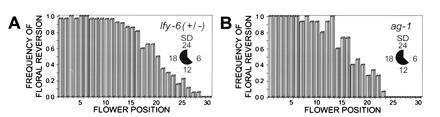
Frequency of floral meristem reversion exhibits a basipetal-acropetal gradient. (A) Floral meristem reversion in SD heterozygous lfy-6 flowers. (B) Floral meristem reversion in SD ag-1 flowers.
We also observed that floral meristem reversion occurs at a low frequency in SD-grown control plants. Table 1 shows that the first 1–2 flowers produced by the primary inflorescence in Landsberg-erecta produce an ectopic shoot at a frequency of two flowers per every five plants. A similar reversion can be induced in several ecotypes including Dijon, Wein, Muhlen, and Moscow (A. Murphy, L. Taiz, and J.K.O., unpublished results). Based on these results we propose that the increased frequency of floral meristem reversion observed in heterozygous lfy-6 and in ag-1 flowers may be due to a decreased sensitivity of the floral meristem for a signal required for the maintenance of flower meristem identity, or alternatively, to an enhanced sensitivity to a signal that promotes shoot development.
Control of Ectopic Shoot Formation by Phytochrome.
The observation that flower meristem reversion is photoperiod-dependent suggested to us that meristem identity may be governed in part by the phytochrome system of photoreceptors. To explore this hypothesis we first used a night-break strategy similar to that used to analyze the role of phytochrome in the photoperiodic control of flowering time in Arabidopsis (29). Table 1 shows that a 0.5-h night-break pulse of red light was sufficient to suppress floral reversion in ag-1 flowers. However, the same treatment was only 87% effective on heterozygous lfy-6. By contrast, a 2-h night-break was sufficient to suppress floral meristem reversion in both mutants (Table 1).
Although the night-break results could be construed as being consistent with a phytochrome hypothesis one could argue that night-break treatments suppress floral meristem reversion by simply promoting LD growth. Thus we adopted a genetic approach to test whether phytochrome activity is required for SD floral meristem reversion. There are at least five phytochrome genes in Arabidopsis (PHYA-E) (34, 35). The activities of all five genes are strongly suppressed by the hy1 mutation that blocks phytochrome chromophore biosynthesis (36). We used the hy1-1 allele (37) to generate ag-1 hy1-1 and lfy-6(+/−) hy1-1 mutants and grew them under SDs. Table 1 shows that hy1-1 completely suppressed SD floral meristem reversion in ag-1 flowers. Meristem reversion was suppressed by 87% in heterozygous lfy-6. Together, these results support the proposal that floral meristem identity in heterozygous lfy and ag flowers is controlled in part by phytochrome.
Gibberellin Signal Transduction Promotes the Maintenance of Flower Meristem Identity.
How does the perception of photoperiod by phytochrome control flower development? First, previous studies showed that gibberellin synthesis or activity is governed by photoperiod in many plant species including Arabidopsis (38, 39, 40, 41). In tobacco, gibberellin levels in the leaf can be reduced by the overexpression of the oat phytochrome A gene (42). Second, the gibberellins are one of at least two signals that can promote flowering in Arabidopsis (6, 7, 8). Under SD conditions the gibberellin biosynthetic mutant ga1-3 does not flower but grows vegetatively for an indefinite period of time (8). Moreover, SD-grown ga1-3 mutants can be rescued by the application of exogenous gibberellins (8). Thus, gibberellins are absolutely required to initiate flowering under SD conditions. Therefore, we reasoned that floral meristem reversion may be a programmed response to decreased gibberellin levels or gibberellin activity.
To test the gibberellin hypothesis we treated SD heterozygous lfy-6 and SD ag-1 plants with 10−5 M GA3 and with 10−5 M GA4+7. Both GA3 and GA4+7 have been shown to be biologically active in Arabidopsis (7, 43). Table 1 shows that GA3 completely suppressed floral reversion in both mutants. There was no detectable difference between LD and GA3-treated SD flower morphology. GA4+7 was equally effective in suppressing floral reversion (K.D.J., C.L.-P., and J.K.O., data not shown). Thus exogenous gibberellins promote the maintenance of flower meristem identity over shoot meristem identity.
To determine genetically whether a gibberellin signal response can suppress floral meristem reversion we used the SPINDLY (SPY) gene mutations spy-2 and spy-3 (26). Homozygous mutations in SPY activate a basal level of gibberellin signaling in a hormone independent manner but exhibit no dramatic effects on floral meristem identity. Table 1 shows that both spy-2 and spy-3 completely suppressed floral meristem reversion in SD heterozygous lfy-6 flowers. Together, these results show that a gibberellin signal can promote the maintenance of floral meristem identity in SD heterozygous lfy and in ag mutant flowers and that a decrease in gibberellin activity may be responsible for SD changes in flower development.
DISCUSSION
We have begun to explore the signals and genes that control the maintenance of flower meristem identity in Arabidopsis using a novel process called floral meristem reversion. Our results show that flower meristem identity and determinacy must be actively maintained in Arabidopsis, even after the initiation of all four types of floral organs, and that the maintenance of flower meristem identity involves LFY and AG, as well as phytochrome and gibberellin signal transduction.
The frequent occurrence of floral meristem reversion observed in SD heterozygous lfy-6 flowers (Fig. 2A) provides two important insights into LFY gene function. First, LFY is important for the maintenance of flower meristem identity. Second, LFY is haplo-insufficient for the maintenance of flower meristem identity under SDs. Since ectopic shoot formation is detectable in stage 8 heterozygous lfy-6 flowers (Fig. 1Q), and since LFY gene activity is also required to initiate flower development (12, 15, 16, 18), we hypothesize that LFY is active continuously from initiation through at least the middle of flower development and organogenesis. Consistent with this hypothesis, LFY gene transcripts are detectable during early flower development, in flower anlage to the immature carpels of stage 9 flowers (16).
We observed that AG is also required for the maintenance of flower meristem identity under SDs. Unlike LFY, however, AG is haplo-sufficient for the maintenance of meristem identity. One important question for understanding the contribution of LFY and AG to the maintenance of meristem identity is whether the effect of heterozygous LFY on flower meristem identity is due, in part, to the role of LFY as a regulator of AG gene expression. Weigel and Meyerowitz (14) showed that the appearance of AG transcripts is delayed in young LD homozygous lfy flowers. They also showed, however, that AG transcript levels recovered as mutant flowers matured (14). Our results suggest that AG gene expression is not affected at the RNA level in SD heterozygous lfy-6 flowers since AG transcript levels (Fig. 1P) are equal to if not greater than that observed in LD wild-type flowers (K.D.J., data not shown). Thus, it does not appear that floral meristem reversion in heterozygous lfy-6 flowers is the result of the down regulation of AG gene expression.
Do LFY and AG function together to maintain flower meristem identity? LFY represents a novel class of evolutionarily conserved nuclear proteins (16) that reportedly can bind to DNA in vitro (20). By contrast, AG belongs to the MADS domain family of transcription factors (24, 25) including the flower control genes AP1, AP3, and PI (44, 45, 46). The AG protein is capable of forming both homo- and heterodimers in vitro (47). Although it is not known whether LFY and AG interact physically in cells, it has been reported that lfy-1 ag-1 flowers display novel inflorescence-like traits such as branching and the production of axillary floral buds under LDs (15), suggesting that LFY interacts genetically with AG. LFY has also been shown to interact genetically with AP1 (15, 16) and with AP2 (12, 13, 15) to promote flower meristem identity and to positively regulate the expression of genes that specify flower organ identity including AP3, PI, and AG (14). By contrast, LFY and AG may function together to maintain flower meristem identity, in part, by negatively regulating the expression of genes that promote shoot development such as TFL (48, 49), EMF (50), or STM (51). Consistent with this hypothesis, we have detected the ectopic expression of a putative Arabidopsis homolog of the snapdragon shoot apical meristem gene CENTRORADIALIS (52) in ectopic shoots produced by SD heterozygous lfy-6 flowers (W.S. and J.K.O., data not shown).
How is flower meristem identity governed by photoperiod? SD-induced floral meristem reversion in heterozygous lfy-6 and in ag-1 plants was strongly suppressed by either a 30-min or a 120-min red light night-break treatment (Table 1), suggesting that phytochrome might be involved in the control of flower meristem identity. However, it could be argued that the long night-break treatment suppresses floral meristem reversion by simply inducing a LD phenotype rather than by a photochemical change in phytochrome activity. Our double mutant studies using hy1-1 provide genetic support for the role of phytochrome in controlling flower meristem identity (Table 1). hy1-1 completely suppressed floral meristem reversion in ag-1 mutant flowers (Table 1). hy1-1 also strongly suppressed meristem reversion in heterozygous lfy-6 flowers. However, a low but significant level of residual flower meristem reversion was observed in heterozygous lfy-6 hy1-1 flowers (Table 1). One interpretation of the residual flower reversion is that a second signal, perhaps from another pigment system, may promote ectopic shoot development. Alternatively, residual flower meristem reversion may be attributable to the presence of a low level of physiologically active and biochemically detectable phytochrome in hy1-1 plants (53, 54).
The frequency of floral meristem reversion in SD heterozygous lfy-6 and in ag-1 flowers forms a basipetal–acropetal gradient along the inflorescence stem (Fig. 2), suggesting that the concentration, activity, or perception of one or more intrinsic signals may vary with time or distance during inflorescence development. Similarly, the effects of homozygous lfy, ap1, and ap2 mutations on floral meristem activity are position dependent and sensitive to photoperiod (10, 11, 12, 15, 16, 33, 55, 56). We hypothesize that the position-dependent change in the establishment and maintenance of flower meristem identity in these mutants can be explained, in part, by changes in gibberellin signal transduction which in turn is controlled by phytochrome. Consistent with this hypothesis we showed that meristem reversion is suppressed in both heterozygous lfy-6 and in ag-1 flowers by exogenously applied gibberellins and genetically by mutations in spy (Table 1). Moreover, the basipetal–acropetal gradient of flower phenotypes exhibited by ap1 and weak ap2 mutant flowers (10, 11, 12, 55, 56) is also suppressed by gibberellins (K.D.J., C.L.-P., and J.K.O., data not shown) and by spy (C. Brown and J.K.O., unpublished results). Taken together, these results suggest that gibberellin signal transduction is important to the control of Arabidopsis floral meristem identity.
Our observation that cells that initiate floral meristem reversion are separated, temporally and spatially, from those that initiate flower development at the shoot apex (Fig. 1) suggests that the establishment and maintenance of flower meristem identity are distinct processes. We can propose a speculative model for the maintenance of flower meristem identity that begins to integrate phytochrome and gibberellin signal transduction together with AG and LFY gene activity. As shown in Fig. 3, the maintenance of flower meristem identity under SDs is controlled, in part, by a putative signal transduction cascade involving phytochrome, gibberellins, SPY, AG, and LFY. One key feature of this model is that floral meristem identity is positively regulated by gibberellins and that gibberellin synthesis or activity is downregulated by phytochrome under SDs, as it is in many plant species including Arabidopsis (38, 39, 40, 41). Fig. 3 also shows that gibberellin signal transduction is negatively regulated by SPY as shown by Jacobsen and Olszewski (26). In contrast to wild type, both heterozygous lfy and ag plants are competent for the establishment of flower meristem identity under SDs, but a gibberellin-mediated signal is required for the maintenance of meristem identity. Floral meristem reversion is not observed in homozygous lfy-6 plants because the requirement for LFY in SD flower development is absolute (12, 15).
Figure 3.
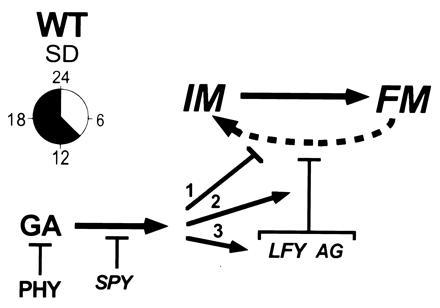
A speculative model for the photo and hormonal control of the maintenance of floral meristem identity by LFY and AG. In wild-type plants cells produced by the inflorescence meristem (IM) are committed to flower meristem development (FM) by a network of genes including AP1, AP2, CAL, CLV1, CLV3, LFY, and others. Under SDs floral meristem reversion, represented by the broken arrow from FM to IM, is strongly but not totally repressed by LFY and AG (see Table 1). GA signal transduction is negatively controlled by SPY. GA synthesis or activity is negatively regulated by phytochrome (PHY). This model suggests three possible pathways for the maintenance of flower meristem identity by gibberellin (GA) signal transduction as shown by arrows 1–3 (see text).
According to the model, there are three possible pathways for the maintenance of flower meristem identity by a gibberellin-mediated phytochrome signal transduction cascade in wild-type flowers under SDs (Fig. 3). First, gibberellins may promote the maintenance of flower meristem identity by a pathway independent of LFY and AG gene activity (arrow 1). For example, floral meristem reversion has been reported at very low frequencies in both ap1-1 and clv1 flowers (3, 11). It is not known, however, whether the frequency of floral reversion in these mutants is significantly greater than that which we observed in wild-type flowers (Table 1). It has also been reported that the frequency of floral meristem reversion is higher in ap1-1 ag-1 and ap1-1 clv1 flowers (3, 11), and in ap1-1 ap2-2 ag-1 flowers (11). However this could be attributed to the presence of ag in these mutants, or to the role of CLV1 as a positive regulator of AG gene expression (3). Second, gibberellins may promote the activity of genes located genetically downstream from LFY and AG (arrow 2) or genes that function together with LFY to promote determinate flower development such as AP1/CAL (18), CLV1 (3), CLV3 (17), or the flowering time gene CONSTANS (CO) (57). Finally, gibberellins may repress floral meristem reversion by promoting LFY gene activity directly or indirectly (arrow 3). We reasoned that LFY is a likely target for gibberellin signaling because heterozygous lfy-6 mutants exhibit meristem reversion and respond to exogenous gibberellins, whereas homozygous lfy-6 mutants remain blocked in flower development. By contrast, AG is not a likely target of gibberellins because ag-1 is a null mutant (24) yet displays strong floral meristem reversion and responds to exogenous gibberellins. Alternatively, gibberellins may suppress the activity of genes in the floral meristem that may promote shoot development such as TFL, EMF, or STM (48, 49, 50, 51).
The capacity of the floral meristem, normally a determinate structure, to be developmentally reprogrammed and switch to indeterminate shoot development has been observed in many plant species. Floral meristem reversions were described for roses and carnations in 1790 by Johann von Goethe (58) and have been observed in monocotyledonous and dicotyledonous plant species (59). The combination of phytochrome and gibberellin signal transduction mutants together with lfy and ag provide a novel opportunity to integrate plant signal transduction with the genetic and molecular systems controlling plant reproduction.
Acknowledgments
We thank Covington Brown for generating the lfy-6 spy and ag-1 spy double mutants, Dr. Gary Drews for assistance with the in situ hybridization procedure, and Dr. Jon Krupp for assistance with microscopy and image processing. We also thank Drs. John Harada and Bob Fischer for critical reading of this manuscript. Finally we wish to thank Drs. Winslow Briggs, Bernard Phinney, Jane Silverthorne, and Lincoln Taiz for their encouragement. This study was supported by National Institutes of Health Grant GM46309 to J.K.O. B.G.W.d.B. was supported by a European Community predoctoral fellowship. C.L.-P. was supported by a University of California–Systemwide Biotechnology Training Grant. W.S. was supported by National Institutes of Health–Minority Biomedical Research Support Program Grant RR08132.
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Abbreviations: GA3, gibberellin A3; GA4+7, gibberellin A4+7; LD, long day; SD, short day; [SD(D)], SD double incident light.
References
- 1.Medford J I, Callos J D, Behringer F J, Link B M. In: Arabidopsis. Meyerowitz E M, Somerville C R, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1994. pp. 355–378. [Google Scholar]
- 2.Sussex I M. Cell. 1989;56:225–229. doi: 10.1016/0092-8674(89)90895-7. [DOI] [PubMed] [Google Scholar]
- 3.Clark S E, Running M P, Meyerowitz E M. Development (Cambridge, UK) 1993;119:397–418. doi: 10.1242/dev.119.2.397. [DOI] [PubMed] [Google Scholar]
- 4.Laibach F. Beitr Biol Pflanz. 1951;28:173–210. [Google Scholar]
- 5.Gregory F G, Hussey G G. Proc Linn Soc London. 1953;164:137–139. [Google Scholar]
- 6.Napp-Zinn K. In: The Induction of Flowering: Some Case Histories. Evans L T, editor. Melbourne, Australia: Macmillan; 1969. pp. 291–304. [Google Scholar]
- 7.Langridge J. Nature (London) 1957;180:36–37. [Google Scholar]
- 8.Wilson R N, Heckman J W, Somerville C R. Plant Physiol. 1992;100:403–408. doi: 10.1104/pp.100.1.403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Martínez-Zapater J, Coupland G, Dean C, Koornneef M. In: Arabidopsis. Meyerowitz E M, Somerville C R, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1994. pp. 403–433. [Google Scholar]
- 10.Irish V F, Sussex I M. Plant Cell. 1990;2:741–753. doi: 10.1105/tpc.2.8.741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bowman J L, Alvarez J, Weigel D, Meyerowitz E M, Smyth D R. Development (Cambridge, UK) 1993;119:721–743. [Google Scholar]
- 12.Schultz E A, Haughn G W. Development (Cambridge, UK) 1993;119:745–765. [Google Scholar]
- 13.Shannon S, Meeks-Wagner D R. Plant Cell. 1993;5:639–655. doi: 10.1105/tpc.5.6.639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Weigel D, Meyerowitz E M. Science. 1993;261:1723–1726. doi: 10.1126/science.261.5129.1723. [DOI] [PubMed] [Google Scholar]
- 15.Huala E, Sussex I M. Plant Cell. 1992;4:901–913. doi: 10.1105/tpc.4.8.901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Weigel D, Alvarez J, Smyth D R, Yanofsky M F, Meyerowitz E M. Cell. 1992;69:843–859. doi: 10.1016/0092-8674(92)90295-n. [DOI] [PubMed] [Google Scholar]
- 17.Clark S E, Running M P, Meyerowitz E M. Development (Cambridge, UK) 1995;121:2057–2067. [Google Scholar]
- 18.Weigel D, Nilsson O. Nature (London) 1995;377:495–500. doi: 10.1038/377495a0. [DOI] [PubMed] [Google Scholar]
- 19.Haughn G W, Schultz E A, Martínez-Zapater J M. Can J Bot. 1995;73:959–981. [Google Scholar]
- 20.Weigel D. Annu Rev Genet. 1995;29:19–39. doi: 10.1146/annurev.ge.29.120195.000315. [DOI] [PubMed] [Google Scholar]
- 21.Okamuro J K, den Boer B G W, Jofuku K D. Plant Cell. 1993;5:1183–1193. doi: 10.1105/tpc.5.10.1183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mizukami Y, Ma H. Plant Mol Biol. 1995;28:767–784. doi: 10.1007/BF00042064. [DOI] [PubMed] [Google Scholar]
- 23.Sieburth L E, Running M P, Meyerowitz E M. Plant Cell. 1995;7:1249–1258. doi: 10.1105/tpc.7.8.1249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yanofsky M F, Ma H, Bowman J L, Drews G N, Feldmann K A, Meyerowitz E M. Nature (London) 1990;346:35–38. doi: 10.1038/346035a0. [DOI] [PubMed] [Google Scholar]
- 25.Schwarz-Sommer Z, Huijser P, Nacken W, Saedler H, Sommer H. Science. 1990;250:931–936. doi: 10.1126/science.250.4983.931. [DOI] [PubMed] [Google Scholar]
- 26.Jacobsen S E, Olszewski N E. Plant Cell. 1993;5:887–896. doi: 10.1105/tpc.5.8.887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jofuku K D, den Boer B G W, Van Montagu M, Okamuro J K. Plant Cell. 1994;6:1211–1225. doi: 10.1105/tpc.6.9.1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Smyth D R, Bowman J L, Meyerowitz E M. Plant Cell. 1990;2:755–767. doi: 10.1105/tpc.2.8.755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Goto N, Kumagai T, Koornneef M. Physiol Plant. 1991;83:209–215. [Google Scholar]
- 30.Haughn G W, Somerville C R. Dev Genet. 1988;9:73–89. [Google Scholar]
- 31.Bowman J L, Smyth D R, Meyerowitz E M. Plant Cell. 1989;1:37–52. doi: 10.1105/tpc.1.1.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Drews G N, Bowman J L, Meyerowitz E M. Cell. 1991;65:991–1002. doi: 10.1016/0092-8674(91)90551-9. [DOI] [PubMed] [Google Scholar]
- 33.Schultz E A, Haughn G W. Plant Cell. 1991;3:771–781. doi: 10.1105/tpc.3.8.771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sharrock R A, Quail P H. Genes Dev. 1989;3:1745–1757. doi: 10.1101/gad.3.11.1745. [DOI] [PubMed] [Google Scholar]
- 35.Clack T, Mathews S, Sharrock R A. Plant Mol Biol. 1994;25:413–427. doi: 10.1007/BF00043870. [DOI] [PubMed] [Google Scholar]
- 36.Parks B M, Quail P H. Plant Cell. 1991;3:1177–1186. doi: 10.1105/tpc.3.11.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Koornneef M, Rolff E, Spruit C J P. Z Pflanzenphysiol. 1980;100S:147–160. [Google Scholar]
- 38.Gianfagna T, Zeevaart J A D, Lusk W J. Plant Physiol. 1983;72:86–89. doi: 10.1104/pp.72.1.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pharis R P, Evans L T, King R W, Mander L N. Plant Physiol. 1987;84:1132–1138. doi: 10.1104/pp.84.4.1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Talón M, Zeevaart J A D. Planta. 1992;188:457–461. doi: 10.1007/BF00197035. [DOI] [PubMed] [Google Scholar]
- 41.Xu Y-L, Li L, Wu K, Peeters A J M, Gage D A, Zeevaart J A D. Proc Natl Acad Sci USA. 1995;92:6640–6644. doi: 10.1073/pnas.92.14.6640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jordan E T, Hatfield P M, Hondred D, Talón M, Zeevaart J A D, Vierstra R D. Plant Physiol. 1995;107:797–805. doi: 10.1104/pp.107.3.797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kobayashi M, Gaskin P, Spray C R, Suzuki Y, Phinney B O, MacMillan J. Plant Physiol. 1993;102:379–386. doi: 10.1104/pp.102.2.379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mandel M A, Gustafson-Brown C, Savidge B, Yanofsky M F. Nature (London) 1992;360:273–277. doi: 10.1038/360273a0. [DOI] [PubMed] [Google Scholar]
- 45.Jack T, Brockman L L, Meyerowitz E M. Cell. 1992;68:683–697. doi: 10.1016/0092-8674(92)90144-2. [DOI] [PubMed] [Google Scholar]
- 46.Goto K, Meyerowitz E M. Genes Dev. 1994;8:1548–1560. doi: 10.1101/gad.8.13.1548. [DOI] [PubMed] [Google Scholar]
- 47.Reichmann J L, Krizek B A, Meyerowitz E M. Proc Natl Acad Sci USA. 1996;93:4793–4798. doi: 10.1073/pnas.93.10.4793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Alvarez J, Guli C L, Yu X-H, Smyth D R. Plant J. 1992;2:103–116. [Google Scholar]
- 49.Shannon S, Meeks-Wagner D R. Plant Cell. 1991;3:877–892. doi: 10.1105/tpc.3.9.877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sung Z R, Belachew A, Shunong B, Bertrand-Garcia R. Science. 1992;258:1645–1647. doi: 10.1126/science.258.5088.1645. [DOI] [PubMed] [Google Scholar]
- 51.Barton M K, Poethig R S. Development (Cambridge, UK) 1993;119:823–831. [Google Scholar]
- 52.Bradley D, Carpenter R, Copsey L, Vincent C, Rothstein S, Coen E. Nature (London) 1996;379:791–797. doi: 10.1038/379791a0. [DOI] [PubMed] [Google Scholar]
- 53.Cone J W, Kendrick R E. Planta. 1985;163:43–54. doi: 10.1007/BF00395896. [DOI] [PubMed] [Google Scholar]
- 54.Parks B M, Shanklin J, Koornneef M, Kendrick R E, Quail P H. Plant Mol Biol. 1989;12:425–437. doi: 10.1007/BF00017582. [DOI] [PubMed] [Google Scholar]
- 55.Komaki M K, Okada K, Nishino E, Shimura Y. Development (Cambridge, UK) 1988;104:195–203. [Google Scholar]
- 56.Kunst L, Klenz J E, Martínez-Zapater J, Haughn G W. Plant Cell. 1989;1:1195–1208. doi: 10.1105/tpc.1.12.1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Putterill J, Robson F, Lee K, Simon R, Coupland G. Cell. 1995;80:847–857. doi: 10.1016/0092-8674(95)90288-0. [DOI] [PubMed] [Google Scholar]
- 58.von Goethe J W. Versuch die Metamorphose der Pflanzen zu erklaren. Gotha, Germany: Carl Wilhem Ettinger; 1790. [Google Scholar]
- 59.Battey N H, Lyndon R F. Bot Rev. 1990;56:162–189. [Google Scholar]