

Abstract
In vertebrates, inheritance of mitochondria is thought to be predominantly maternal, and mitochondrial DNA analysis has become a standard taxonomic tool. In accordance with the prevailing view of strict maternal inheritance, many sources assert that during fertilization, the sperm tail, with its mitochondria, gets excluded from the embryo. This is incorrect. In the majority of mammals—including humans—the midpiece mitochondria can be identified in the embryo even though their ultimate fate is unknown. The “missing mitochondria” story seems to have survived—and proliferated—unchallenged in a time of contention between hypotheses of human origins, because it supports the “African Eve” model of recent radiation of Homo sapiens out of Africa. We will discuss the infiltration of this mistake into concepts of mitochondrial inheritance and human evolution.
Keywords: paternal mitochondria, mammalian fertilization, evolutionary theory
Mitochondrial DNA
Mitochondria carry out oxidative phosphorylation and mediate most of the energy in eukaryotic organisms. In plant cells, the equivalent organelles are plastids or chloroplasts. Both mitochondria and chloroplasts are thought to have resulted from ancestral endosymbiotic relationships between nucleated cells and free living bacteria capable of exploiting oxygen (1). In mammals, mitochondria have retained a restricted set of independent circular genomes of 16.6 kilobases each coding for 13 polypeptides, part of the oxidative phosphorylation pathway. The remaining approximately 80 control genes in the somatic nuclei are Mendelian inherited. Mitochondria, along with their mitochondrial DNA (mtDNA) are semiautonomous and have the capacity to replicate, divide, and fuse independent of the somatic nuclear division cycle (2). Most, but not all, eukaryotic organisms show predominant uniparental inheritance of mtDNA, an adaptation perhaps necessary to avoid lethal genome conflict between subservient organelles (3). Because the mitochondrial genome replicates frequently, is deficient in DNA proofreading and repair mechanisms (4), and is vulnerable to cumulative damage from free radicals generated by the electron transport chain, the mitochondrial genome mutates rapidly. Sequestration of the dangerous process of oxidative phosphorylation to an extranuclear site may have evolved to protect the somatic genome (5). In addition, there are close links between alterations to the mitochondrial genome, bioenergetic diseases, and aging (6, 7).
The combination of mainly uniparental inheritance and frequent mutation invites great interest in mtDNA as an indicator of evolutionary relationships (8). However, as we will demonstrate here, the interest in maternal inheritance that began as scientific curiosity has given rise to an absolute dogma in the school of human evolutionary studies based on molecular biology. In this process, a basic error in fertilization biology has emerged.
Mitochondria and Mammalian Fertilization
“Maternal inheritance” of mtDNA in mammals is of interest to reproductive biologists; as in almost all species the entire sperm, including the midpiece mitochondrial sheath, enters the egg at fertilization.
Subsequently, tail and midpiece structures can be traced for several division cycles (9, 10). The only known exception to this is the Chinese hamster, Cricetulus griseus (11, 12). Here the tail and midpiece of the giant sperm (the largest known among Eutheria) (13) remains outside the oocyte after fertilization. Partial or delayed incorporation of the tail may also occur in some Insectivora (14). The typical mammalian sperm midpiece contains approximately 50–75 mitochondria with one copy of mtDNA in each. This represents an 8- to 10-fold decline in copy number during spermiogenesis (15). In contrast, the mammalian oocyte contains around 100,000 (105) to 100,000,000 (108) mitochondria (16), and the human oocyte in particular is estimated to contain 100,000 (105) copies of mtDNA (17). Thus the oocyte’s mtDNA copy number exceeds that of the sperm by a factor of at least 1000 (103).
As the sperm takes several weeks to form and mature within the male tract before ejaculation and the mtDNA may well be degraded during this period, the simplest explanation for so called maternal inheritance is that the paternal contribution is diluted beyond the limits of detection using conventional restriction enzyme analysis. While selective inactivation or destruction of the paternal mtDNA by differential methylation has been suggested (a form of genomic imprinting), there is no evidence to support this (15).
Intriguingly, the oocyte’s mtDNA derives from a very small pool (perhaps as few as five) precursor mitochondrial genomes during oogenesis. This selective pressure may serve as a selective genetic filter against defective mitochondrial genomes, just as it may explain why some intergenerational changes may occur so rapidly (18, 19).
There is a wide variety of patterns of inheritance of cytoplasmic genes and organelles among multicellular organisms. Some are strictly maternal, some paternal, and some are mixed (3). It is quite possible that between mammalian species, cytoplasmic factors, such as the paternal mtDNA and the sperm centrosome, function differently once they are inside the ovum. In humans, some of these may contribute to infertility though arrested or altered syngamy (20). However, there can be no doubt that at fertilization paternal mitochondria do enter into the ovum (Fig. 1). We do not know what really happens to them in subsequent development in most vertebrate species.
Figure 1.
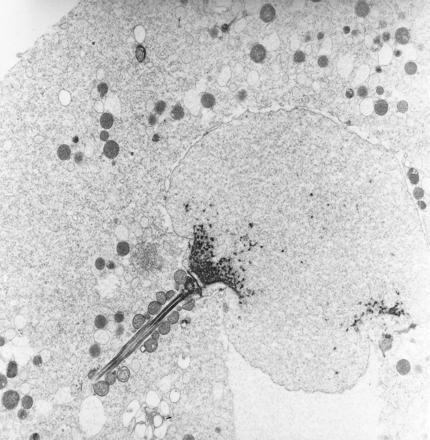
Human sperm within the egg. The partially decondensed sperm head is still attached to its midpiece circa 6 h after penetration (circa × 22,000). [Reproduced with permission from Sathananthan et al. (24) (Copyright 1986, Wiley, New York).]
Recently the case for obligatory maternal inheritance of mitochondrial DNA was strengthened by Kaneda et al. (21), who found specific elimination of paternal mitochondrial DNA (detected by double round PCR with nested primers) in intraspecific hybrids of Mus musculus, but not in interspecific crosses between M. musculus and Mus spretus. Moreover, paternal mtDNA was also eliminated from those embryos derived from M. musculus oocytes fertilized by sperm of the congenic strain B6.mtspr, which carries M. spretus mitochondrial DNA on a background of M. musculus nuclear genes.
They concluded that the most likely explanation was some form of interaction between cytoplasmic factors and nuclear encoded proteins of the sperm midpiece such as the sperm-specific microtubules. This would be consistent with ultra structural evidence of midpiece dissolution in association with multivesicular bodies in hamster oocytes (22). However, we should be cautious in extrapolating these observations from rodents to humans. Mice show unusual patterns of cytoplasmic inheritance. The centrosome, for example, is maternally inherited in mice, while in humans and many other vertebrates (where known) it is derived from the sperm centrosome.
Thus the elimination of paternal cytoplasmic inheritance may be a recent evolutionary development confined to the rapid radiation of the Rodentia (23). Certainly in human embryos intact sperm midpiece mitochondria can be identified after fertilization (24, 25) and later up to the morula stage (A. H. Sathananthan, personal communication), so it is not possible at present to be sure that they are ultimately eliminated.
Fate of Sperm Mitochondria and Other Organelles
Most early studies assumed that the sperm mitochondria participated fully in early embryonic development. However, in 1965, Szollosi (26), reporting the fate of sperm midpiece mitochondria in rats, observed that they remain associated with axonemal structures and did not distribute evenly between blastomeres. Szollosi (26) found that these mitochondria swelled and appeared to disintegrate by the eight-cell stage. This differed from previous observations in the mouse (27). Szollosi (26) concluded, “If it is true that sperm mitochondria disintegrate, it could be postulated that only maternal type mitochondria would be found in the embryo. If further experiments confirm these observations, this system would provide an excellent opportunity for studying cytoplasmatic inheritance in higher animals.”
Is this how the story of strict maternal mtDNA inheritance began? Probably not, as the roots go back at least to a description of a “throw away” sperm tail by Wells et al. (ref. 28, p. 149) in 1935. They wrote, “After the spermatozoon has activated the ovum it does something further. Its head burrows actually into the substance of the ovum, leaving the tail outside to wriggle for a while and then perish.” Wells had apparently been unfamiliar with Lillie’s 1923 Problems of Fertilization (29) that clearly set out accurate observations on sperm entry dating back into the 19th century. Lillie did indeed observe loss of sperm tails during fertilization in the Annelid Nereis and considered this to be so unusual as to warrant special comment (30).
Twenty-eight years after Szollosi’s study, Simerly et al. (31) studied the incorporation of the sperm tail into early embryos of mice and concluded: “… Fertilization involves contributions of various and different components by sperm and egg… ” suggesting that “… This hypothesis of maternal inheritance may need modification.” Thus, Simerly et al. (31) reject the tentative hypothesis of maternal inheritance at least on ultrastructural grounds. It appears that spermatozoa may indeed contribute a number of other cytoplasmic factors that modulate early embryogenesis, including the centrosome, unique sperm proteins, and cell cycle modulators (32, 33, 34, 35).
Mitochondrial Inheritance in Studying Human Evolution
Most human anatomy textbooks correctly report that in mammals the entire sperm cell, including the mitochondria and flagellum, enters the egg at fertilization (36, 37, 38). However, these facts appear to have been ignored by some scholars of anthropology and human evolution.
There have been many faulty iterations that the sperm tail (including the midpiece) plays no part in development (refs. 39, 40, 41, to mention only three). This error has even crept into standard texts on biochemistry (42). The misconception that all mtDNA inheritance must be maternal threatens future research. For example, Cann et al. (43) give several reasons to establish the suitability of mtDNA for their study, one being, “… because mtDNA is inherited maternally and does not recombine, it is a tool for relating individuals to one another.” They claim “Molecular biology is now a major source of quantitative and objective information about the evolutionary history of the human species.”
This error of obligatory maternal inheritance of mtDNA in humans appears to arise entirely from approaches using restriction fragment analysis (44, 45). However, Gyllensten et al. (46), using the PCR, detected paternally inherited mtDNA molecules in mice at a frequency of 1 in 10,000 (104), relative to the maternal contributions. They concluded: “Paternal inheritance of mtDNA also means that mtDNA phylogenies are not exclusively matriarchal.” [Note that the paternal inheritance rate of 1 in 10,000 (104) is that to be expected by simple dilution.]
The Myth of the Nonparticipating Midpiece
The myth of the missing midpiece seems to have been well established by the mid 1980s. In a review of the evolution of mtDNA in primates Spuhler (ref. 47, p. 20) declares: “Most models for the evolutionary genetics of mtDNA in mammalia assume that inheritance is strictly maternal (Wilson et al., 1985). Although the sperm of vertebrates may contain hundreds of mitochondria in the middle-piece, usually that structure does not enter the fertilized egg, and if paternal mitochondria do enter, seemingly few, if any, survive the first equational cell division.” This is clearly incorrect. Indeed, the paper by Wilson et al. (48) to which Spuhler refers neither provides concrete information nor gives any reason to explain why mtDNA inheritance should be strictly maternal.
Examples of reports about the wayward sperm tail are widespread, like a Dawkinsian “meme” [a unit of cultural transmission, not necessarily correct, that survives through psychological appeal (49)]. More recently the idea of the discarded sperm tail seems to serve as a support for the African Eve hypothesis of recent human evolution and radiation from Africa about 150,000 years ago (43). Regardless whether the African Eve hypothesis is correct or not (44), it cannot not be supported by the assumption of strictly matrilineal mtDNA evolution.
In 1992, Wilson and Cann (50) restated their conviction that the molecular approach, not fossil evidence, provides the most direct documentation to unravel human evolution. They also published a cartoon of an egg and a sperm that illustrates how “All the mitochondria and the DNA they contain, however, derive from the egg.” The sperm in the cartoon contains a single stylized mitochondrion in the head. In the article’s text they state “… unlike nuclear DNA, mitochondrial DNA is inherited from the mother alone, unchanged except for chance mutations.” They also suggest two “evolutionary trees” that illustrate strictly maternal genetic lineages of descent. Responding in the same issue, two anthropologists [Thorne and Wolpoff (51)] while rejecting the mtDNA molecular clock that Wilson and Cann advocate repeatedly reiterate the “meme” about obligatory maternal inheritance of mtDNA. Indeed, Wolpoff (52) has recently reasserted that mtDNA is only passed from mother to offspring and explains that all humans get their cytoplasm and its contents from the oocyte, not the sperm. Furthermore, general texts such as Lewin’s (53) illustrate the loss of the entire sperm tail with a detailed caricature showing and describing the “Discarded sperm, still carrying mitochondria.” (Fig. 2). Lewin concludes: “… Unlike an individual’s nuclear genes, which are a combination of genes from both parents, the mitochondrial genome comes only from the mother (except under unusual circumstances). Because of this maternal mode of inheritance, there is no recombination of maternal and paternal genes, which sometimes blurs the history of the genome as read by geneticists. Potentially, therefore mitochondrial DNA offers a powerful way of inferring population history, unhindered by the genetic fog of recombination.” A similar colorful—and inaccurate— cartoon can be found in Whitfield’s 1995 “The Human Body Explained. A Guide to Understanding the Incredible Living Machine” (54). Also books addressing a general informed audience, such as Shipman (55) reiterate the same misinformation. There are many others. Even the Cambridge Encyclopedia of Human Evolution (56) states on p. 320: “… In addition, mtDNA is mainly maternally inherited: it passes down the female line. This is because the sperm provides almost no cytoplasm (and hence no mitochondria) to the fertilized egg. The egg has thousands of mitochondria and it is these that are passed to the developing embryo.”
Figure 2.
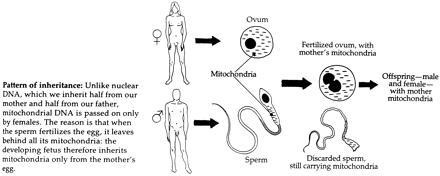
One of the examples of the erroneous loss of the sperm tail depicted in a cartoon. [From Lewin (53) (Reproduced by permission of Blackwell Scientific, Oxford).]
Perhaps the most egregious mistatement appears in Dawkins’ “The Blind Watchmaker” (ref. 57, p. 176): “All the mitochondria in you are descended from the small population that traveled from your mother in her egg. Sperms are too small to contain mitochondria (our italics), so mitochondria travel exclusively down the female line, and male bodies are dead ends as far as mitochondrial reproduction is concerned. Incidentally, this means that we can use mitochondria to trace our ancestry strictly down the female line.”
In one of his most recent books, Dawkins (58) concedes, “Sperms are too small to contain more than a few mitochondria … these mitochondria are cast away with the tail when the sperm head is absorbed in the egg at fertilization.” Having acknowledged almost grudgingly that sperm may contain mitochondria, he elaborates on the significance of the total loss of paternal mtDNA for more than 12 pages. Thus, we see a progression of error from the position that paternal inheritance of mtDNA is insignificant to another where such a phenomenon is declared impossible because the sperm supposedly lacks mitochondria. The common thread in all of this is that many authors have come to accept the obligatory “loss” of paternal mtDNA because it does not conflict with their belief in the African Eve hypothesis (43) and its dependence upon strict maternal inheritance. Indeed the supposed loss of the sperm’s contribution has progressed in step with the intensity of the debate between the “Recent African Genesis” (50) and the “Multiregional Evolution” models of human origin (51). In primatology, the mtDNA genome is even treated as “haplotype” (59, 60). Perhaps the major irony in this story of the wayward sperm midpiece is that Dawkins, who invented the idea of the meme, its transmission, and inherent changeability (49), has unwittingly contributed to the story’s climax. We hope that this article will lead to extinction of this particular meme.
Conclusions
Mistakes in science should be corrected. If they are being perpetrated by scientists who are copying from one another without checking the facts, a serious dilemma is created. Trustingly naive misstatements concerning the total loss of the paternal mtDNA at fertilization, based on the false assumption that the sperm tail is discarded, must not be used to support theories of evolutionary relationships based on obligatory maternal inheritance of mitochondria. We simply do not yet know what happens to the paternal contribution of mtDNA in humans, but the simplest explanation is that it is diluted beyond recognition by researchers using relatively low resolution techniques of molecular biology. We do know that paternal mtDNA enters the oocyte at fertilization. Models of human evolution using mtDNA analysis must take these facts into account. All theories of the timing of human evolution that depend on the premise that the sperm midpiece does not contribute to the embryo must be reevaluated or rejected.
Acknowledgments
We thank Dr. Pat Saling for making the connection between F.A.S. in Durham, NC, and J.M.C. in Australia. She also provided useful references. Special thanks to A. H. Sathananthan who provided Fig. 1. Our thanks go to C. V. Simons, E. L. Simons, J. Jernvall, J. T. Laitman, and T. Rasmussen who read the manuscript and helped to clarify the text; to J. M. Bedford, R. Yanagimachi, and G. Schatten for advice; and last not least to K. Richardson as he established the link back to Lillie. We also thank John T. Maltsberger for excellent editorial advice.
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Abbreviation: mtDNA, mitochondrial DNA.
References
- 1.Margulis L, Sagan D. Origins of Sex: Three Billion Years of Genetic Recombination. New Haven, CT: Yale Univ. Press; 1986. [Google Scholar]
- 2.Bereiter-Hahn M Microsc Res Technique. 1994;27:198–219. doi: 10.1002/jemt.1070270303. [DOI] [PubMed] [Google Scholar]
- 3.Hurst L D. Proc R Soc London B Biol Sci. 1992;248:135–140. [Google Scholar]
- 4.Clayton D A. Trends Biochem Sci. 1991;16:107–111. doi: 10.1016/0968-0004(91)90043-u. [DOI] [PubMed] [Google Scholar]
- 5.Lindahl T. Nature (London) 1993;362:709–725. doi: 10.1038/362709a0. [DOI] [PubMed] [Google Scholar]
- 6.Wallace D C, Shoffner J M, Trounce I, Brown M D, Ballinger S W, Corral-Debrinski M, Horton T, Jun A S, Lott M T. Biochim Biophys Acta. 1995;1271:141–151. doi: 10.1016/0925-4439(95)00021-u. [DOI] [PubMed] [Google Scholar]
- 7.Shoffner J M, Wallace D C. Adv Hum Genet. 1990;19:267–330. [PubMed] [Google Scholar]
- 8.Brown W M, George M J, Wilson A C. Proc Natl Acad Sci USA. 1979;76:1967–1971. doi: 10.1073/pnas.76.4.1967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fleming A D, Cummins J M, Kuehl T J, Seidel G E, Yanagimachi R. J Exp Zool. 1986;237:383–390. doi: 10.1002/jez.1402370311. [DOI] [PubMed] [Google Scholar]
- 10.Shalgi R, Magnus A, Jones R, Phillips D M. Mol Reprod Dev. 1994;37:264–271. doi: 10.1002/mrd.1080370304. [DOI] [PubMed] [Google Scholar]
- 11.Pickworth S, Chang M C. J Reprod Fertil. 1969;19:371–374. doi: 10.1530/jrf.0.0190371. [DOI] [PubMed] [Google Scholar]
- 12.Yanagimachi R, Kamiguchi Y, Sugawara S, Mikamo K. Gamete Res. 1983;8:97–117. [Google Scholar]
- 13.Cummins J M, Woodall P F W. J Reprod Fertil. 1985;75:153–175. doi: 10.1530/jrf.0.0750153. [DOI] [PubMed] [Google Scholar]
- 14.Bedford J M, Cooper G W, Phillips D M, Dryden G L. Biol Reprod. 1994;50:820–834. doi: 10.1095/biolreprod50.4.820. [DOI] [PubMed] [Google Scholar]
- 15.Hecht N B, Liem H, Kleene K C, Distel R J, Ho S M. Dev Biol. 1984;102:452–461. doi: 10.1016/0012-1606(84)90210-0. [DOI] [PubMed] [Google Scholar]
- 16.Piko L, Matsumoto L. Dev Biol. 1976;4:1–10. doi: 10.1016/0012-1606(76)90253-0. [DOI] [PubMed] [Google Scholar]
- 17.Chen X, Prosser R, Simonetti S, Sadlock J, Jagiello G, Schon E A. Neurology. 1994;44:A336. [PMC free article] [PubMed] [Google Scholar]
- 18.Nagley P, Zhang C, Martinus R D, Vaillant F, Linnane A W. In: Mitochondrial DNA in Human Pathology. DiMauro S, Wallace D, editors. New York: Raven; 1993. pp. 137–157. [Google Scholar]
- 19.Hauswirth W W, Laipis P J. In: Achievements and Perspectives of Mitochondrial Research. Quagliariello E, Slater E C, Pamieri F, Saccone C, Kroon A M, editors. Amsterdam: Elsevier; 1985. pp. 49–59. [Google Scholar]
- 20.Asch R, Simerly C, Ord T, Ord V A, Schatten G. Hum Reprod. 1995;10:1897–1906. doi: 10.1093/oxfordjournals.humrep.a136204. [DOI] [PubMed] [Google Scholar]
- 21.Kaneda H, Hayashi J I, Takahama S, Taya C, Lindahl K F, Yonekawa H. Proc Natl Acad Sci USA. 1995;92:4542–4546. doi: 10.1073/pnas.92.10.4542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hiraoka J, Hirao Y. Gamete Res. 1988;19:369–380. doi: 10.1002/mrd.1120190408. [DOI] [PubMed] [Google Scholar]
- 23.Schatten G. Dev Biol. 1994;165:299–235. doi: 10.1006/dbio.1994.1256. [DOI] [PubMed] [Google Scholar]
- 24.Sathananthan A H, Ng S C, Edirisinghe R, Ratnam S S, Wong P C. Gamete Res. 1986;15:317–326. [Google Scholar]
- 25.Sathananthan A H, Ratnam S S, Ng S C, Tarin J J, Gianarolli L, Trounson A. Hum Reprod. 1996;11:346–356. doi: 10.1093/humrep/11.2.345. [DOI] [PubMed] [Google Scholar]
- 26.Szollosi D. J Exp Zool. 1965;159:367–378. doi: 10.1002/jez.1401590309. [DOI] [PubMed] [Google Scholar]
- 27.Gresson R A R. Nature (London) 1940;145:425. [Google Scholar]
- 28.Wells H G, Huxley J S, Wells G P. The Science of Life. Doran, and Company, Garden City, NY: Doubleday; 1935. [Google Scholar]
- 29.Lillie F R. Problems of Fertilization. Chicago: Univ. of Chicago Press; 1923. [Google Scholar]
- 30.Wilson E B. The Cell in Development and Heredity. New York: Macmillan; 1928. [Google Scholar]
- 31.Simerly C R, Hecht N B, Goldberg E, Schatten G. Dev Biol. 1993;158:536–548. doi: 10.1006/dbio.1993.1211. [DOI] [PubMed] [Google Scholar]
- 32.Navara C S, First N L, Schatten G. Dev Biol. 1994;162:29–40. doi: 10.1006/dbio.1994.1064. [DOI] [PubMed] [Google Scholar]
- 33.Schatten G. Dev Biol. 1994;165:299–235. doi: 10.1006/dbio.1994.1256. [DOI] [PubMed] [Google Scholar]
- 34.Simerly C, Wu G J, Zoran S, Ord T, Rawlins R, Jones J, Navara C, Gerrity M, Rinehart J, Binor Z, Asch R, Schatten G. Nat Med. 1995;1:47–52. doi: 10.1038/nm0195-47. [DOI] [PubMed] [Google Scholar]
- 35.Tesarik J, Souza M. Fertil Steril. 1994;62:1197–1204. doi: 10.1016/s0015-0282(16)57185-4. [DOI] [PubMed] [Google Scholar]
- 36.Crouch J E. Functional Human Anatomy. Philadelphia: Lea and Febiger; 1965. [Google Scholar]
- 37.Gray H. Gray’s Anatomy of the Human Body. Philadelphia: Lea and Febiger; 1995. [Google Scholar]
- 38.Curtis H, Barnes N S. Biology. New York: Worth; 1989. [Google Scholar]
- 39.Harrison G A, Tanner J M, Pilbeam D R, Baker P T. Human Biology. An Introduction to Human Evolution, Variation, Growth, and Adaptability. Oxford: Oxford Univ. Press; 1988. [Google Scholar]
- 40.Brown M H. Have Scientists Found the Mother of us all? The Search for Eve. New York: Harper and Row; 1990. [Google Scholar]
- 41.Rushton J P. Race, Evolution, and Behavior. A Life History Perspective. New Brunswick, London: Transaction; 1995. [Google Scholar]
- 42.Lehniger A L, Nelson D L, Cox M M. Principles of Biochemistry. New York: Worth Publishers; 1993. [Google Scholar]
- 43.Cann R L, Stoneking M, Wilson A. Nature (London) 1987;325:31–36. doi: 10.1038/325031a0. [DOI] [PubMed] [Google Scholar]
- 44.Templeton A R. Am Anthropol. 1993;95:51–72. [Google Scholar]
- 45.Giles R E, Blanc H, Cann H M, Wallace D C. Proc Natl Acad Sci USA. 1980;77:6715–6719. doi: 10.1073/pnas.77.11.6715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gyllensten U, Wharton D, Josefsson A, Wilson A C. Nature (London) 1991;352:255–257. doi: 10.1038/352255a0. [DOI] [PubMed] [Google Scholar]
- 47.Spuhler J N. Yearbook Phys Anthropol. 1988;31:15–48. [Google Scholar]
- 48.Wilson A C, Cann R L, Carr S M, Matthew G, Gyllensten U B, Helm-Bychowski K M, Higuchi G, Palumbi S R, Prager E M, Sage R D, Stoneking M. Biol J Linnean Soc. 1985;26:375–400. [Google Scholar]
- 49.Dawkins R. The Selfish Gene. Oxford: Oxford Univ. Press; 1989. [Google Scholar]
- 50.Wilson A C, Cann R L. Sci Am. 1992;266:22–27. doi: 10.1038/scientificamerican0492-68. [DOI] [PubMed] [Google Scholar]
- 51.Thorne A G, Wolpoff M H. Sci Am. 1992;266:28–33. doi: 10.1038/scientificamerican0492-76. [DOI] [PubMed] [Google Scholar]
- 52.Wolpoff M H. Human Evolution. New York: McGraw–Hill; 1996. [Google Scholar]
- 53.Lewin R. Human Evolution. An Illustrated Introduction. Oxford: Blackwell; 1993. [Google Scholar]
- 54.Whitfield P. The Human Body Explained. A Guide to Understanding the Incredible Living Machine. New York: Henry Holt; 1995. [Google Scholar]
- 55.Shipman P. Human Differences and the Use and Abuse of Science. The Evolution of Racism. New York: Simon & Schuster; 1995. [Google Scholar]
- 56.Jones S, Martin R, Pilbeam D. The Cambridge Encyclopedia of Human Evolution. Cambridge, U.K.: Cambridge University Press; 1992. [Google Scholar]
- 57.Dawkins R. The Blind Watchmaker. New York: W. W. Norton; 1986. [Google Scholar]
- 58.Dawkins R. River out of Eden, A Darwinian View of Life. New York: Basic Books; 1995. [Google Scholar]
- 59.Melnick D J, Hoelzer G A. Evol Anthropol. 1993;2:2–10. doi: 10.1093/oxfordjournals.molbev.a040004. [DOI] [PubMed] [Google Scholar]
- 60.Cann R L, Rickards O, Lum J K. In: Origins of Anatomically Modern Humans. Nitecki M H, Nitecki D V, editors. New York: Plenum; 1993. pp. 135–149. [Google Scholar]