

Abstract
Tularemia is a zoonosis of humans caused by infection with the facultative intracellular bacterium Francisella tularensis. Interest in F. tularensis has increased markedly in the past few years because of its potential use as an agent of bioterrorism. Five subspecies of this organism are found in the Northern hemisphere, but only F. tularensis subsp. tularensis and subsp. holarctica cause disease in humans. This review summarizes what is known about the pathogenesis of tularemia with a focus on bacterial surface components such as lipopolysaccharide and capsule as well as information obtained from the F. tularensis subsp. tularensis SCHU S4 genome. In particular, the mechanisms of action of recently identified virulence factors are discussed in the context of bacterial replication in macrophages and manipulation of the host inflammatory response. Throughout this report shared and unique features of F. tularensis subsp. tularensis, subsp. holarctica, and subsp. novicida are discussed.
Keywords: lipopolysaccharide, genome, virulence factor, macrophage, cytokines
INTRODUCTION
The anthrax attacks that occurred during the fall of 2001 dramatically alerted Americans to the potential deadly effects of bioterrorism and prompted the government of the United States to support research into the development of diagnostic, preventive, and therapeutic measures to combat organisms with biowarfare potential. The use of biological agents as weapons has a long history that predates our understanding of the microbial basis of infectious disease (12, 29, 64, 80, 114). As early as 400 B.C., Scythian archers dipped their arrowheads in feces or decaying cadavers, and during the Mongol siege of Kaffa in 1347 plague-infected bodies were hurled into the city by the Tartars. In 1650, a Polish artillery officer suggested that projectiles filled with the saliva of rabid dogs could be used to cause epidemics, and during the American Revolutionary War blankets contaminated with variola were given to Native Americans. Before World War II, Japan, France, Germany, the Soviet Union, and Great Britain all had active biological weapons programs and the United States began similar activities in the early 1940s.
Pursuant to an executive order, the United States terminated its biological weapons program in 1970 and all stockpiles of infectious agents were destroyed by 1973 (29). Consequently, there was hope that this form of warfare would be abandoned worldwide. Indeed, the Biological Weapons Convention was implemented in 1972 and biowarfare was renounced by 140 nations (29). Nevertheless, the number of countries or other entities that possess toxic biological agents is increasing (80). This finding, together with the potential severity of natural infections, necessitates an understanding of mechanisms of pathogenesis and the human immune response in order to develop appropriate treatments, preventive vaccines, and other defensive measures.
TAXONOMY AND GEOGRAPHIC DISTRIBUTION
In 1911, an outbreak of plague-like illness in ground squirrels in Tulare County, California, led to the discovery of a new pathogen by McCoy & Chapin (88). Shortly thereafter, it was appreciated that these hardy, nonspore-forming bacteria could persist in the environment for long periods and cause disease in humans. In honor of Edward Francis, these small, nonmotile, gram-negative cocco-bacilli are now called Francisella tularensis.
Francisella is the only genus within the family Francisellaceae and, on the basis of small subunit RNA sequences, is a member of the γ-subclass of proteobacteria (49). The organism most closely related to Francisella is Wolbachia persica, a tick endosymbiont (49). As defined by DNA similarity and fatty acid composition, there are two species within the Francisella genus: tularensis and philomiragia (49, 67). In addition, there are five subspecies of F. tularensis: tularensis (also called F. tularensis type A), novicida, mediasiatica, holarctica (F. tularensis type B), and a variant of holarctica found in Japan (113) (Figure 1). Of these, only F. tularensis subsp. tularensis and subsp. holarctica cause disease in humans; and highly virulent type A organisms are evolutionarily older than moderately virulent type B bacteria (113). The live vaccine strain (LVS) is an attenuated variant of F. tularensis subsp. holarctica and F. philomiragia is a muskrat pathogen.
Figure 1.

Evolutionary schema based on loss of regions of difference and extensive nucleotide variation. Adapted from Svennson et al. (113).
F. tularensis is widely distributed in the Northern hemisphere and is capable of infecting hundreds of different vertebrates and invertebrates (97). Recent studies have shown that F. tularensis is resistant to killing by the free-living amoebae, Acanthamoeba castellanii (1). In Norway, environmental infection of Acanthamoeba with F. tularensis may be a major source of outbreaks of gastrointestinal and respiratory tularemia in humans (16). The most important mammalian species involved in human infection include lagomorphs, voles, mice, squirrels, muskrats, and beavers. Infectious transfer to humans and other mammals can occur through aerosolization of infected material, the bite of an insect (ticks), and contact with infected animal products. F. tularensis is present throughout the continental United States, and infections caused by this organism were common prior to World War II but have declined dramatically since the 1950s (97). The current case rate is approximately 0.05 reported infections per 100,000 population, with 60% of all cases occurring in Midwestern states, such as Arkansas, Missouri, South Dakota, and Oklahoma (23a).
All F. tularensis subspecies and other Francisellaceae can be differentiated biochemically (97) and genetically by polymerase chain reaction (PCR) amplification of regions of difference (21). Recent studies have further divided F. tularensis subsp. tularensis into two genetically distinct clades, type A1 and type A2 (47). F. tularensis subsp. tularensis and subsp. holarctica also differ in geographic distribution (47). Type A1 organisms are found predominately in the Midwest, California, and Massachusetts, and type A2 bacteria have been detected in California and in the Mountain states. In marked contrast, F. tularensis subsp. holarctica is less common in North America and is distributed across Europe and Asia.
POTENTIAL FOR USE IN BIOWARFARE
F. tularensis subsp. tularensis is considered a potential bioweapon because of its extreme virulence, low infectious dose, ease of aerosol dissemination, and capacity to cause severe illness and death. Indeed, inhalation of as few as 10 colony-forming units (CFU) is sufficient to cause disease in humans and 30% to 60% of untreated infections can be fatal (109, 110). The highly infectious nature of F. tularensis has been appreciated since the mid-twentieth century owing to water-borne outbreaks in Europe and the Soviet Union, laboratory accidents, and epizootic cases in the United States (52, 69, 70, 115). The largest naturally occurring outbreak of airborne tularemia occurred in Sweden in 1966–1967, during which more than 600 farmers were infected with F. tularensis type B (41).
Japanese germ warfare research units studied F. tularensis subsp. tularensis, and this organism may have been used against Chinese civilians, Russian troops, and American prisoners of war between 1932 and 1945 (65). In addition, the appearance of tularemia in thousands of Russian and German troops at the siege of Stalingrad may have been the result of deliberate use by the Soviets (3). However, a natural cause for this outbreak has not been eliminated, and military personnel may have acquired F. tularensis from mice and rats whose numbers multiplied owing to the widespread disruption of sanitation and hygiene during battle (39). During the Cold War both the Soviet Union and the United States prepared and stockpiled tons of infectious agents for potential use against the civilian populations of their enemies (29). In 1969, a World Health Organization expert committee estimated that aerosol dispersal of 50 kg of virulent F. tularensis bacteria over a metropolitan area with five million inhabitants would result in 250,000 casualties requiring extensive medical care and 19,000 deaths (42, 104a).
BACTERIAL STRUCTURE
Lipopolysaccharide
Lipopolysaccharide (LPS, or endotoxin) is the major component of the outer membrane of gram-negative bacteria (103), and the interaction of LPS/MD-2 complexes with Toll-like receptor 4 (TLR4) on macrophages and endothelial cells activates a signaling cascade that results in the release of proinflammatory cytokines (23, 56, 103). Escherichia coli LPS consists of a hexaacylated glucosamine disaccharide phosphorylated at the 1- and 4′-positions (lipid A) that is modified further by the addition of core oligosaccharides and a variable number of additional sugars that form the O-antigen (103). Alterations in LPS structure, including changes in phosphorylation or fatty acid chain length, position, and number, significantly affect endotoxin bioactivity (103, 116). In this regard, it is noteworthy that the LPS of F. tularensis is 1000-fold less potent than the LPS of enteric bacteria (4, 106) and does not activate cells via TLR4 (106, 117).
Recent studies have begun to define unique structural features of Francisella LPS that may account for its low bioactivity. The LPS of both F. tularensis subsp. holarctica and LVS is a tetraacylated structure that lacks fatty acids at the O-3 position of hexosamine II in the lipid A backbone (Figure 2), and compared with E. coli, the fatty acids that are present are unusually long (98, 125). LVS lipid A is not phosphorylated at either the 1- or 4′-position (125). Dephosphorylation of the 1-position is catalyzed by LpxE and requires prior LPS transport across the inner membrane by ValA, a homolog of the ABC transporter MsbA (89, 90, 127). In contrast to LVS, F. tularensis subsp. holarctica LPS contains a galactosamine-1-phosphate linked to the reducing terminus of the lipid A disaccharide (Figure 2) (98). A homolog of the Salmonella enterica serovar Typhimurium arabinosamine transferase, ArnT, is present in the SCHU S4 genome and may encode a galactosamine transferase (98). Modification of Salmonella lipid A with arabinose confers resistance to polymyxin B and cationic antimicrobial peptides (103), but whether galactosamine has a similar effect on Francisella LPS is unclear.
Figure 2.
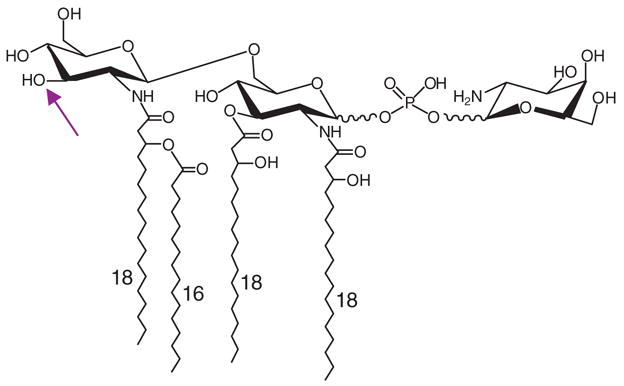
The lipid A of F. tularensis subsp. holarctica strain 1547. Note the asymmetrical lipid A structure and the absence of O-acylation on hexosamine II (arrow). The phosphogalactosamine substitution on the 4′-position of this hexosamine is also unique to Francisella.
Structural analysis indicates that the core region of F. tularensis LPS contains a single 3-deoxy-D-manno-octulosonic acid and lacks heptose (125). Specifically, the structure of the core region sugars is
The O-antigens of F. tularensis subsp. tularensis and subsp. holarctica LPS are identical, but the O-antigen of F. tularensis subsp. novicida is distinct (101, 119, 124, 126). The repeating subunits of the F. tularensis subsp. tularensis and subsp. novicida O-antigens are (124)
F. tularensis subsp. tularensis
F. tularensis subsp. novicida
The O-antigen gene cluster in the SCHU S4 genome contains an insertion element (IS-Ftu1) that is flanked by transposase genes, and a similar insertion element in Shigella sonnei is essential for virulence (77, 101). ISFtu1 may be responsible for Francisella LPS phase variation, which in turn modulates bacterial fate in murine macrophages (37, 72).
Capsule
Wild strains of F. tularensis and LVS possess a surface exopolysaccharide capsule (68, 105, 112), and our data demonstrate the presence of immunogenic surface capsular material on F. tularensis subsp. holarctica strain 1547 (Figure 3). It has been known for some time that the F. tularensis capsule protects bacteria from killing by serum complement (105, 112) and a cluster of genes putatively involved in polysaccharide biosynthesis is present in the SCHU S4 genome (77). Nevertheless, a detailed analysis of the capsule has not been performed and our understanding of its composition is incomplete. In the 1970s biochemical studies found that capsular extracts of SCHU S4 contain carbohydrates (including mannose, rhamnose, and two unidentified dideoxy sugars), as well as amino acids and α-OH 14:0 and 16:0 fatty acids (68). A role for capsule in virulence is suggested by the finding that bacteria from which the capsular material has been extracted do not cause disease in mice or guinea pigs (68). Whether capsule synthesis is regulated in vivo or in vitro is unknown. However, continued growth on synthetic media can restore encapsulation and virulence to a relatively avirulent, unencapsulated strain of LVS (28), and recent data suggest that capsule may be downregulated by LVS inside macrophages (57).
Figure 3.
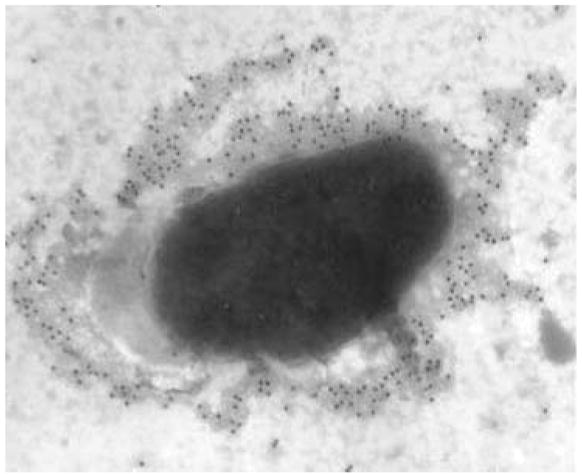
A whole mount electron micrograph of F. tularensis subsp. holarctica strain 1547 fixed in glutaraldehyde with ruthenium red and stained with monoclonal antibody XE8 and secondary antibodies conjugated to colloidal gold to show the putative surface capsule. Magnification: ×15,000.
Pilus
Orthologs of genes required for type IV pilus expression are present in the F. tularensis subsp. tularensis SCHU S4 genome (55, 77), and microscopy data demonstrate the presence of structures that resemble type IV pili on the surface of LVS (55). In other organisms type IV pili mediate bacterial attachment to host cells, DNA uptake, twitching motility, and biofilm formation (22, 24, 95). Nevertheless, the role of pili in Francisella physiology and virulence has not been determined.
GENOME
The genome of F. tularensis subsp. tularensis SCHU S4 is small, approximately 1.9 Mb, and contains 1804 predicted coding sequences, of which 302 are unique to Francisella (77). Five of the predicted coding sequences unique to strain SCHU S4 form a new protein family of unknown function (77). Because no obvious virulence factors were predicted by the genome, the relatively large number of Francisella-specific predicted coding sequences might provide the bacterium with novel uncharacterized factors responsible for the high virulence of this organism. Thus, it is of interest that F. tularensis subsp. tularensis does not have either of the plasmids found in LVS or F. tularensis subsp. novicida (77, 96, 100).
A large proportion of inactivated genes characterize the SCHU S4 genome. As a result of insertions, deletions, and substitution mutations, more than 10% of all genes are pseudogenes or gene fragments; and 14% of all pseudogenes can be accounted for by the presence of five insertion elements (IS-Ftu 1–5) scattered throughout the genome (77, 120). The decaying genome of F. tularensis has disrupted more than half of the predicted metabolic pathways for which genes are present and explains the fastidious growth requirements of the organism (77). Fourteen compounds are essential to sustain growth of Francisella because of absent or incomplete enzymatic pathways.
A unique 33.9-kb region of DNA is duplicated in the SCHU S4 genome (77). The 25 genes encoded within this region have no known bacterial homologs and may encode as yet unidentified virulence determinants. However, the presence of this genomic island in LVS as well as F. tularensis subsp. novicida indicates that these genes alone cannot account for the high virulence of F. tularensis subsp. tularensis (91). Because disruption of genes within this region generates mutant bacteria that are attenuated for survival in macrophages, this region of the chromosome has been called the Francisella pathogenicity island (FPI) (58, 61, 75, 91).
Compared with other bacteria, F. tularensis has relatively few transcriptional regulators (77). MglA is the only regulatory protein described to date. Encoding proteins homologous to the E. coli stationary-phase regulators SspA and SspB, the mglAB operon is required for F. tularensis subsp. novicida intracellular growth and intra-amoeba survival (14, 78). MglA regulates expression of several FPI genes, including iglC, iglA, pdpA, and pdpD (78).
F. tularensis does not appear to secrete toxins, and genes encoding type III, IV, or V secretion systems have not been identified in the SCHU S4 genome. However, homologs of pilin and pseudopilin genes associated with type II secretion are present (55), along with 15 potential ATP-binding cassette systems that may be involved in type I secretion (77).
Although iron is an essential nutrient for F. tularensis, genes encoding TonB and receptors for siderophores, transferrin, lactoferrin, or other iron-containing compounds are not present in the SCHU S4 genome (50, 77). Nevertheless, a gene encoding the ferric uptake protein, Fur, and genes regulated by Fur (FTT0030) are present, indicating that expression of some genes may be iron dependent (77).
Genetic Tools for Manipulation of Francisella tularensis
Efforts to understand F. tularensis pathogenesis have been hindered by of a lack of reliable methods for genetic manipulation of the virulent Francisella subspecies and by the inability to make targeted mutations. Because F. tularensis subsp. novicida is more genetically amenable by standard transformation techniques, most genetic manipulation has been conducted in this bacterium. However, recent efforts of several researchers have provided the Francisella community with tools to introduce DNA into the other subspecies of Francisella and to make specific mutations. These methods have been successfully employed in A- and B-type strains, creating mutations in many genes including those encoded within the FPI.
pFNL10, a cryptic plasmid isolated from F. tularensis subsp. novicida strain F6168, is the basis for all F. tularensis–replicating plasmids (96). The origin of Francisella replication in this plasmid was determined by deletion and sequence analysis (96, 100). Using pFNL10, E. coli–F. tularensis shuttle vectors were created, but these plasmids are not stably maintained within F. tularensis (73, 86, 94). A stable Francisella plasmid, pFNLTP1, was created by spontaneous deletion during passage through LVS (86). A conditionally replicating derivative of pFNLTP1, pFNLTP9, was created as a vector for allelic exchange or transposon delivery (86).
Conjugation, transformation, electroporation, and cryotransformation have been used to introduce DNA into F. tularensis. Conjugation has been performed in both the SCHU S4 strain and LVS to create mutations (60, 122). In LVS, transfer is most efficient within 18 h of mating and at 25°C (60). Chemical transformation of DNA has been used successfully only in F. tularensis subsp. novicida, and efficiency of DNA uptake depends on the plasmid construct and the length and source of DNA Moreover, Francisella DNA is transformed with higher efficiency than non-Francisella DNA (10, 123). Attempts to transform LVS using this method were unsuccessful (10). Electroporation is an efficient method of introducing DNA into type A Francisella spp., LVS, and F. tularensis subsp. novicida (10, 86, 100). Transformation efficiency of electroporated DNA is approximately 3–6 × 107 CFU μg−1 of DNA in LVS and 2 × 103 CFU μg−1 of DNA in F. tularensis subsp. novicida (86). Transformation efficiency of F. tularensis subsp. novicida is 2 × 108 when novicida DNA is transformed, suggesting the presence of a restriction modification system; no such system has been observed in LVS (86). Subspecies and strains of Francisella transform with different efficiencies and the presence of an exopolysaccharide capsule may affect DNA uptake (10, 13, 86, 100). Cryotransformation is an efficient and reliable method of introducing DNA into F. tularensis subsp. novicida and subsp. holarctica, and it is a fast procedure because competent cells do not need to be made (75, 96).
Mutational analysis is critical to elucidating the pathogenesis of F. tularensis. Francisella mutants were initially created using random transposon mutagenesis of F. tularensis DNA in E. coli, followed by transformation into F. tularensis and allelic exchange (10, 14, 17, 36, 61). More recently, stable random mutants have been constructed in LVS using the Epicentre EZ::TN transposon mutagenesis system (71). Allelic replacement using the suicide plasmid pPV was used to mutate iglC in LVS and FTT0918 in strain SCHU S4; both genes are required for intracellular survival (60, 122).
IMMUNOPATHOGENESIS
Infection of Macrophages
F. tularensis can infect macrophages of humans, mice, rats, rabbits, and guinea pigs, and it is generally believed that macrophages are the major reservoir of Francisella in vivo. In the past few years several research groups have begun to define the niche occupied by this organism inside host cells and to delineate how bacterial growth and survival are modulated by specific virulence factors.
Receptors and phagocytosis
Internalization of microbes and particles via phagocytosis requires specific receptor-ligand interactions and is driven by local actin polymerization (2). The binding interactions that confer uptake of Francisella by phagocytes are only beginning to be defined. Of note in this regard is the fact that cells of the commonly used murine macrophage cell line J774 are remarkably resistant to F. tularensis (in standard tissue culture media), and high multiplicities of infection (MOI of 100:1 to 500:1) are needed achieve uptake of one or two bacteria per cell (74, 76, 117). Phagocytosis of F. tularensis by primary murine bone marrow–derived macrophages and human monocytes is also inefficient (18, 31). These data contrast sharply with the notion that macrophages are the major intracellular reservoir of F. tularensis in vivo and the fact that as few as 10 organisms can cause severe disease. Therefore, in vitro model systems may not be optimized. Indeed, recent data indicate that infection of human monocyte-derived macrophages (MDM) or monocyte-like THP-1 cells by a clinical F. tularensis isolate or LVS is enhanced markedly by fresh serum containing active complement factors (30, 31). Under these conditions uptake of opsonized bacteria is mediated in part by complement receptor 3 (CD11b/CD18) and occurs via the extension of large asymmetric pseudopod loops that project from the macrophage surface to encircle attached bacteria (31).
Taken together, the data suggest that serum opsonins are required for optimal uptake of F. tularensis by mononuclear phagocytes. Nevertheless, in future studies it will be important to compare directly the ability of F. tularensis subspecies to infect different types of mononuclear phagocytes including alveolar macrophages in the presence and absence of opsonins and to assess whether infection efficiency is modulated by surface virulence factors such as capsule or LPS. In this regard it is of interest that both capsule-deficient organisms and LPS-phase variants are serum sensitive (36, 66, 105, 112). Moreover, diminished O-antigen content impairs virulence in mice and reduces survival in macrophages in vitro (36, 66).
Phagosome escape and replication in the cytosol
Under normal circumstances nascent phagosomes undergo a maturation process during which the composition of this compartment is modified by sequential interactions with early endosomes, late endosomes, and lysosomes (2). In the mature phagolysosome, oxidants, cationic antimicrobial peptides, and lysosomal acid hydrolases act in concert to kill and degrade ingested bacteria. Pathogens that replicate inside macrophages must either evade or withstand this hostile microenvironment. It has been known for some time that LVS phagosomes do not fuse with thorium-loaded lysosomes in J774 cells (7) and that ingested bacteria begin to replicate after a lag of approximately 6 h (7, 50). The composition of the Francisella phagosome was not determined in these early studies. However, phagosome neutralization inhibits LVS replication, which suggested that F. tularensis might reside in an acidic vacuole that allows access to iron and other essential nutrients (50). By contrast, recent ultrastructural analyses demonstrate that both clinical F. tularensis isolates and LVS escape the phagosome in primary human and murine macrophages (as well as transformed macrophage cell lines) and replicate in the cytosol (30, 57). Confocal microscopy studies have begun to define the nature of the compartment from which F. tularensis escapes. Nascent phagosomes containing live clinical F. tularensis isolates, LVS, or F. tularensis subsp. novicida associate transiently with early endosomes and thereafter accumulate the late-endosome markers lamp-1, lamp-2, and CD63 (30, 57, 107). However, further maturation of the F. tularensis phagosome does not occur; these compartments are only moderately acidified (pH 6.7) and do not acquire lysosomal hydrolases (30, 107). A progressive decline in phagosome lamp-1 and CD63 beginning 3 to 4 h into infection correlates directly with disruption of the phagosome membrane and with the presence of bacteria in the cytosol (30, 57).
That all F. tularensis isolates studied to date escape the phagosome and replicate in the cytosol of human and murine macrophages suggests that this process is controlled by conserved virulence determinants. Nevertheless, how the phagosome membrane is breached is unknown. Virulence factors with homology to listerolysin O have not been described in F. tularensis, and synthesis of relatively few proteins is affected during intracellular infection (58). Transposon mutagenesis of F. tularensis subsp. novicida demonstrated that iglB and iglC are essential for intramacrophage growth (61). IglC is a 23-kDa protein that is upregulated inside macrophages (58), and the intracellular growth locus, iglABCD, defines an operon that is a major component of the FPI (91). Expression of genes in the FPI is controlled by the transcriptional regulators MglA and MglB (macrophage growth locus A and B) (78). Disruption of mglA, mglB, iglB, or iglC inhibits phagosome escape, intramacrophage growth, and virulence in mice (14, 61, 75, 78, 108). Other FPI genes controlled by MglA include pdpA, pdpB, pdpC, and pdpD (78, 91). The function of the Pdp proteins is unknown; however, it is of interest that pdpD is present in type A organisms, altered in F. tularensis subsp. novicida, and absent in type B strains including LVS (91).
Additional proteins required for intracellular survival are alanine racemase, glutamine phosphoribosylpyrophosphate amido-transferase, ClpB heat-shock protease, and a 58-kDa protein of unknown function (61, 122). Finally, the role of capsule and LPS in intramacrophage survival has not been defined precisely. Strains with rough LPS survive poorly in mice and macrophages (36, 37, 66), but whether phagosome maturation is affected directly has not been determined. The reduced fitness of these organisms may be indirect and may reflect membrane perturbations introduced by the membrane attack complex of the complement cascade.
Apoptosis
Replication of LVS in J774 cells culminates in cell death after approximately 24 h (7, 58). Cytotoxicity requires bacterial replication and does not occur if phagocytosis is blocked or if ingested organisms are killed with ciprofloxacin (74). Dying cells exhibit features of apoptosis including DNA laddering and surface exposure of annexin V (74, 75). Moreover, apoptosis is triggered by bacteria that reach the cytosol, as judged by the sustained viability of J774 cells infected with ΔiglC LVS or the phenotype of thioglycolate-elicited peritoneal macrophages that ingested F. tularensis subsp. novicida mutants ΔmglA or ΔpdpA (75, 87).
Pathogen-induced macrophage apoptosis can favor either the host or the microbe (54). With regard to Francisella, two distinct mechanisms of apoptosis have been described (76, 87). LVS infection of J774 cells (MOI 500:1) activates the intrinsic apoptosis pathway. Mitochondrial cytochrome c release, activation of caspases -9 and -3, and cleavage of poly-ADP ribose polymerase peak at ~18 h and viable bacteria are released from dying cells. Conversely, infection of thioglycolate-elicited peritoneal macrophages with opsonized F. tularensis subsp. novicida (MOI 30:1) triggers rapid apoptosis (90% death within 5 h) via the caspase-1 inflammasome pathway. This mechanism of cell death is an element of innate host defense, and deletion of caspase-1 markedly increases F. tularensis subsp. novicida virulence in mice during the first day post infection. The reasons for these divergent outcomes are unknown; differences in macrophage type and activation state, unique features of different F. tularensis subspecies, or triggering of distinct signaling pathways by opsonized and unopsonized bacteria could account for this discordance.
Role of Neutrophils
Neutrophils (also called polymorphonuclear leukocytes, or PMN) are understudied in the context of tularemia and whether they contribute to pathogenesis or host defense is controversial. The results of one study demonstrated that depletion of granulocytes with monoclonal antibodies markedly increases the lethality of LVS administered to mice via the dermis or the peritoneal cavity (111). Conversely, neutropenia does not exacerbate disease course when LVS is administered via the aerosol route (32). In this model, PMN depletion has no significant effect on bacterial burden in the lung and spleen, and the LVS load in the liver is only moderately elevated. Importantly, this host defense defect was not due to an absence of PMN in the alveoli of control animals, and as such the data suggest that neutrophils may not control F. tularensis in the microenvironment of the lung.
Relatively few studies have examined the interactions of F. tularensis with neutrophils isolated from the peripheral blood of humans or rhesus monkeys. Like most bacteria, binding of F. tularensis subsp. holarctica or LVS to neutrophils and their subsequent phagocytosis requires complement components in fresh serum (83, 102). A central aspect of the killing arsenal of the neutrophil is the NAPDH oxidase, which is a multicomponent enzyme that catalyzes the conversion of molecular oxygen into superoxide anions (93). Superoxide anions rapidly dismutate to form hydrogen peroxide, and in the presence of myeloperoxidase released from primary granules, hydrogen peroxide is converted into highly toxic hypochlorous acid (HOCl) (92). Three studies performed more than 20 years ago examined whether a strain of F. tularensis subsp. holarctica or LVS activated neutrophils as judged by degranulation and synthesis of oxidants (83, 84, 102). The data indicate that F. tularensis opsonized with immune serum is a more potent stimulus than bacteria preincubated with normal human serum or specific antibodies alone. Moreover, normal serum does not support killing of F. tularensis by PMNs and >95% of ingested bacteria remain viable. In contrast, in the presence of immune serum approximately one third of virulent bacteria and nearly all attenuated organisms are eliminated. Furthermore, LVS is more susceptible than wild F. tularensis to superoxide and HOCl in a cell-free system (85). Taken together, the data suggest that oxidants contribute to Francisella control by neutrophils, that wild F. tularensis may be more resistant to reactive oxygen species than attenuated bacteria, and, most importantly, that oxidative mechanisms of host defense may not be effective during primary infection.
Virulence factors that modulate the phagocyte respiratory burst are largely undefined. A purified acid phosphatase (AcpA) inhibits the oxidative burst of porcine neutrophils stimulated with formyl peptides or phorbol esters (104). However, acpA-null mutants of F. tularensis subsp. novicida are not attenuated in mice (15). Thus, resistance to toxic oxidants may be multifactorial. In support of this notion, capsule-deficient variants of LVS are more resistant to killing by PMNs despite increased serum sensitivity (105), and inactivation of minD enhances F. tularensis subsp. novicida sensitivity to superoxide anions in vitro and impairs virulence in mice (8).
Cytokines and Phagocyte Activation in Host Defense
Interferon (IFN)-γ, tumor necrosis factor-α, and interleukin (IL)-12 play important roles in murine resistance to LVS and F. tularensis subsp. novicida (9, 43, 45, 46, 51, 59, 79, 81). IL-12 is a potent stimulus for production of IFN-γ, which in turn induces nitric oxidase synthase (iNOS) and synthesis of NO by macrophages (11, 38, 51, 62, 63). NO is bacteriostatic (82) and studies of iNOS-null animals confirm a key role for reactive nitrogen intermediates in Francisella control (82). IFN-γ also limits bacterial growth inside murine alveolar macrophages by an NO-independent mechanism (99). In contrast to mouse cells, human macrophages generate NO under a limited set of conditions (121), and as such specific mechanisms of control relevant to human tularemia are not well defined. However, recent data indicate that IFN-γ enhances maturation of F. tularensis subsp. novicida phagosomes in human MDMs and impairs phagosome escape (107).
On the other hand, LVS disrupts secretion of proinflammatory cytokines by human mononuclear phagocytes (18, 117, 118) and human umbilical vein endothelial cells (48) and uncouples antigen presentation from cytokine production in murine pulmonary den-dritic cells (19). Determining the mechanism by which Francisella alters cytokine production is an area of active investigation. IglC impairs intracellular signaling pathways required for cytokine synthesis (117). In addition, the unusual structure of F. tularensis LPS likely accounts for its low bioactivity and may prevent cell activation via TLRs (106, 117). Indeed, TLR4 has no significant role in host defense against F. tularensis subsp. tularensis in the lung or against LVS in the dermis (25, 26). These data, together with the finding that Francisella LPS does not act as an antagonist (4), suggest that the ability of Francisella LPS to interact with one or more of the host endotoxin-binding molecules that confer cell activation (LPS binding protein, CD14, or TLR4/MD-2) may be impaired. A role for LPS in modulating macrophage activation during the immune response is also of interest given the higher bioactivity of LPS in F. tularensis subsp. novicida organisms (72).
Late control and ultimate resolution of F. tularensis infection require T-lymphocytes (34, 35, 46, 128). Both CD4+ and CD8+ cells are required to combat SCHU S4, and the ability of F. tularensis subsp. tularensis to cause thymic atrophy and thymocyte depletion may account in part for the marked virulence of this F. tularensis subspecies (27, 53). Conversely, the role of B-lymphocytes in primary tularemia is less clear. In humans antibody levels do not correlate with protection against infection (115) and B-cell-deficient mice do not succumb to sublethal doses of LVS (44). Nevertheless, B-cell-derived cytokines appear to modulate neutrophil influx and killing of infected hepatocytes by mechanisms independent of antibody production (33, 40), and as noted above, specific antibodies may be important as opsonins that favor killing of F. tularensis by neutrophils in the immune host. In support of this notion, B-cell-deficient mice exhibit profound neutrophilia and die secondary to PMN degranulation, tissue damage, and shock in response to a second dose of LVS (20).
CONCLUSIONS
Our understanding of Francisella pathogenesis is advancing rapidly. Sequencing of the genome and the development of tools for genetic manipulation of the organism have enabled researchers to identify novel virulence factors that impair macrophage function and disrupt the host immune response. Other unique features of F. tularensis were identified by structural analysis of LPS and other surface determinants. The wealth of information obtained in the past few years is substantial and the fast pace of new discoveries in this field will likely continue for some time.
Despite recent advances, many important questions have not yet been answered. For example, the features of F. tularensis subsp. tularensis and the aspects of the lung microenvironment that account for the ability of as few as 10 organisms to cause severe pneumonic tularemia in humans have not been defined. Also lacking is a clear understanding of the fate of F. tularensis in cell types other than macrophages. The extent to which F. tularensis interacts with and alters the function of endothelial cells, epithelial cells, neutrophils, and dendritic cells is largely unknown. Given the ability of F. tularensis to cause a wide spectrum of diseases, it is tempting to speculate that virulence factor expression may be affected by route of infection or bacterial residence in different tissues including the lung, skin, spleen, and liver. It is likely that answers to these questions (and many others) will be obtained in the near future and that the insights gained will advance our understanding of the pathogenesis of tularemia and provide information essential for the development of new vaccines.
SUMMARY POINTS
Francisella subspecies differ in geographic distribution and only F. tularensis subsp. holarctica and subsp. tularensis cause disease in humans.
The F. tularensis subsp. tularensis SCHU S4 genome is notable for its small size, the presence of a pathogenicity island, and a large number of pseudogenes that disrupt metabolic pathways.
Francisella tularensis subsp. holarctica LPS has an unusual structure and exhibits low bioactivity.
Opsonized Francisella enter macrophages via complement receptor 3 and escape the phagosome, and bacterial replication in the cytosol culminates in macrophage apoptosis.
LPS, surface capsule, and genes in the pathogenicity island are required for virulence in mice and replication in human and murine macrophages in vitro.
Resolution of infection requires T-cells and associated cytokines. B-cells and antibodies may modulate neutrophil function in the immune host.
Unique features of Francisella that account for the pronounced virulence of F. tularensis subsp. tularensis in humans, particularly in the context of pulmonary tularemia, remain obscure.
Acknowledgments
We thank Dr. Willliam Nauseef for his helpful comments. Research on this topic in the authors’ laboratories is supported by funds from the National Institutes of Health (P01-AI044642 to M.A. and L.A.) and funds from the Midwest Regional Center of Excellence for Biodefense and Emerging Infectious Disease Research (to L.A.).
- Pathogen
a microorganism that causes disease
- Live vaccine strain (LVS)
an attenuated strain of F. tularensis subsp. holarctica previously used as a live vaccine
- CFU
colony-forming unit
- Lipopolysaccharide (LPS, or endotoxin)
the major surface component of gram-negative bacteria
- TLR
Toll-like receptor
- SCHU S4
genome strain of F. tularensis subsp. tularensis
- D-GalNacAN
2-acetamido-2-deoxy-2-galacturonamide
- Capsule
a sugar-rich structure on the surface of some bacteria that can inhibit phagocytosis or prevent killing by serum complement
- Virulence factor
a component of a microorganism that is important for its ability to damage a host and cause disease
- Francisella pathogenicity island (FPI)
a distinct region of the genome containing genes that encode virulence factors
- Igl
intracellular growth locus
- Mgl
macrophage growth locus
- Phagocytosis
a specialized form of endocytosis that allows macrophages and neutrophils to engulf microorganisms
- MOI
multiplicity of infection
- MDM
human monocyte-derived macrophage
- Opsonize
binding of opsonins (serum complement factors and/or antibodies) to the surface of microbes to facilitate phagocytosis
- PMN
polymorphonuclear leukocyte, neutrophil
- iNOS
inducible nitric oxide synthase
LITERATURE CITED
- 1.Abd H, Johansson T, Golovliov I, Sandstrom G, Forsman M. Survival and growth of Francisella tularensis in Acanthamoeba castellanii. Appl Environ Microbiol. 2003;69:600–6. doi: 10.1128/AEM.69.1.600-606.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593–23. doi: 10.1146/annurev.immunol.17.1.593. [DOI] [PubMed] [Google Scholar]
- 3.Alibek K. Biohazard: The Chilling True Story of the Largest Covert Biological Weapons Program in the World, Told From the Inside by the Man Who Ran It. New York: Random House; 1999. [Google Scholar]
- 4.Ancuta P, Pedron T, Girard R, Sandstrom G, Chaby R. Inability of the Francisella tularensis lipopolysaccharide to mimic or to antagonize the induction of cell activation by endotoxins. Infect Immun. 1996;64:2041–46. doi: 10.1128/iai.64.6.2041-2046.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Deleted in proof
- 6.Deleted in proof
- 7.Anthony LD, Burke RD, Nano FE. Growth of Francisella spp. in rodent macrophages. Infect Immun. 1991;59:3291–96. doi: 10.1128/iai.59.9.3291-3296.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Anthony LS, Cowley SC, Mdluli KE, Nano FE. Isolation of a Francisella tularensis mutant that is sensitive to serum and oxidative killing and is avirulent in mice: correlation with the loss of MinD homologue expression. FEMS Microbiol Lett. 1994;124:157–65. doi: 10.1016/0378-1097(94)90243-7. [DOI] [PubMed] [Google Scholar]
- 9.Anthony LS, Ghadirian E, Nestel FP, Kongshavn PA. The requirement for gamma interferon in resistance of mice to experimental tularemia. Microb Pathog. 1989;7:421–28. doi: 10.1016/0882-4010(89)90022-3. [DOI] [PubMed] [Google Scholar]
- 10.Anthony LS, Gu MZ, Cowley SC, Leung WW, Nano FE. Transformation and allelic replacement in Francisella spp. J Gen Microbiol. 1991;137:2697–703. doi: 10.1099/00221287-137-12-2697. [DOI] [PubMed] [Google Scholar]
- 11.Anthony LS, Morrissey PJ, Nano FE. Growth inhibition of Francisella tularensis live vaccine strain by IFN-gamma-activated macrophages is mediated by reactive nitrogen intermediates derived from l-arginine metabolism. J Immunol. 1992;148:1829–34. [PubMed] [Google Scholar]
- 12.Bailey KC. Directors Series on Proliferation. Springfield, VA: Lawrence Livermore Natl. Lab; 1994. [Google Scholar]
- 13.Baron GS, Myltseva SV, Nano FE. Electroporation of Francisella tularensis. Methods Mol Biol. 1995;47:149–54. doi: 10.1385/0-89603-310-4:149. [DOI] [PubMed] [Google Scholar]
- 14.Baron GS, Nano FE. MglA and MglB are required for the intramacrophage growth of Francisella novicida. Mol Microbiol. 1998;29:247–59. doi: 10.1046/j.1365-2958.1998.00926.x. [DOI] [PubMed] [Google Scholar]
- 15.Baron GS, Reilly TJ, Nano FE. The respiratory burst-inhibiting acid phosphatase AcpA is not essential for the intramacrophage growth or virulence of Francisella novicida. FEMS Microbiol Lett. 1999;176:85–90. doi: 10.1111/j.1574-6968.1999.tb13646.x. [DOI] [PubMed] [Google Scholar]
- 16.Berdal BP, Mehl R, Meidell NK, Lorentzen-Styr AM, Scheel O. Field investigations of tularemia in Norway. FEMS Immunol Med Microbiol. 1996;13:191–95. doi: 10.1111/j.1574-695X.1996.tb00235.x. [DOI] [PubMed] [Google Scholar]
- 17.Berg JM, Mdluli KE, Nano FE. Molecular cloning of the recA gene and construction of a recA strain of Francisella novicida. Infect Immun. 1992;60:690–93. doi: 10.1128/iai.60.2.690-693.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bolger CE, Forestal CA, Italo JK, Benach JL, Furie MB. The live vaccine strain of Francisella tularensis replicates in human and murine macrophages but induces only the human cells to secrete proinflammatory cytokines. J Leukoc Biol. 2005;77:893–97. doi: 10.1189/jlb.1104637. [DOI] [PubMed] [Google Scholar]
- 19.Bosio CM, Dow SW. Francisella tularensis induces aberrant activation of pulmonary dendritic cells. J Immunol. 2005;175:6792–801. doi: 10.4049/jimmunol.175.10.6792. [DOI] [PubMed] [Google Scholar]
- 20.Bosio CM, Elkins KL. Susceptibility to secondary Francisella tularensis live vaccine strain infection in B-cell-deficient mice is associated with neutrophilia but not with defects in specific T-cell-mediated immunity. Infect Immun. 2001;69:194–203. doi: 10.1128/IAI.69.1.194-203.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Broekhuijsen M, Larsson P, Johansson A, Bystrom M, Eriksson U, et al. Genome-wide DNA microarray analysis of Francisella tularensis strains demonstrates extensive genetic conservation within the species but identifies regions that are unique to the highly virulent F. tularensis subsp. tularensis. J Clin Microbiol. 2003;41:2924–31. doi: 10.1128/JCM.41.7.2924-2931.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Burrows LL. Weapons of mass retraction. Mol Microbiol. 2005;57:878–88. doi: 10.1111/j.1365-2958.2005.04703.x. [DOI] [PubMed] [Google Scholar]
- 23.Caroff M, Karibian D. Structure of bacterial lipopolysaccharides. Carbohydr Res. 2003;338:2431–47. doi: 10.1016/j.carres.2003.07.010. [DOI] [PubMed] [Google Scholar]
- 23a.Centers for Disease Control. Summary of notifiable diseases, United States, 2001. MMWR Morb Mortal Wkly Rep. 2001;50:1–108. [PubMed] [Google Scholar]
- 24.Chen I, Dubnau D. DNA transformation during transformation. Front Biosci. 2003;8:s544–56. doi: 10.2741/1047. [DOI] [PubMed] [Google Scholar]
- 25.Chen W, KuoLee R, Shen H, Busa M, Conlan JW. Toll-like receptor 4 (TLR4) does not confer a resistance advantage on mice against low-dose aerosol infection with virulent type A Francisella tularensis. Microb Pathog. 2004;37:185–91. doi: 10.1016/j.micpath.2004.06.010. [DOI] [PubMed] [Google Scholar]
- 26.Chen W, Kuolee R, Shen H, Busa M, Conlan JW. Toll-like receptor 4 (TLR4) plays a relatively minor role in murine defense against primary intradermal infection with Francisella tularensis LVS. Immunol Lett. 2005;97:151–54. doi: 10.1016/j.imlet.2004.10.001. [DOI] [PubMed] [Google Scholar]
- 27.Chen W, Kuolee RK, Austin JW, Shen H, Che Y, Conlan JW. Low dose aerosol infection of mice with virulent type A Francisella tularensis induces severe thymus atrophy and CD4+ CD8+ thymocyte depletion. Microb Pathog. 2005;39:189–96. doi: 10.1016/j.micpath.2005.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cherwonogrodzky JW, Knodel MH, Spence MR. Increased encapsulation and virulence of Francisella tularensis live vaccine strain (LVS) by subculturing on synthetic medium. Vaccine. 1994;12:773–75. doi: 10.1016/0264-410x(94)90284-4. [DOI] [PubMed] [Google Scholar]
- 29.Christopher GW, Cieslak TJ, Pavlin JA, Eitzen EMJ. Biological warfare. A historical perspective. JAMA. 1997;278:412–17. [PubMed] [Google Scholar]
- 30.Clemens DL, Lee BY, Horwitz MA. Virulent and avirulent strains of Francisella tularensis prevent acidification and maturation of their phagosomes and escape into the cytoplasm in human macrophages. Infect Immun. 2004;72:3204–17. doi: 10.1128/IAI.72.6.3204-3217.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Clemens DL, Lee BY, Horwitz MA. Francisella tularensis enters macrophages via a novel process involving pseudopod loops. Infect Immun. 2005;73:5892–902. doi: 10.1128/IAI.73.9.5892-5902.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Conlan JW, KuoLee R, Shen H, Webb A. Different host defences are required to protect mice from primary systemic vs pulmonary infection with the facultative intracellular bacterial pathogen, Francisella tularensis LVS. Microb Pathog. 2002;32:127–34. doi: 10.1006/mpat.2001.0489. [DOI] [PubMed] [Google Scholar]
- 33.Conlan JW, North RJ. Early pathogenesis of infection in the liver with the facultative intracellular bacteria Listeria monocytogenes, Francisella tularensis, and Salmonella typhimurium involves lysis of infected hepatocytes by leukocytes. Infect Immun. 1992;60:5164–71. doi: 10.1128/iai.60.12.5164-5171.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Conlan JW, Sjostedt A, North RJ. CD4+ and CD8+ T-cell-dependent and -independent host defense mechanisms can operate to control and resolve primary and secondary Francisella tularensis LVS infection in mice. Infect Immun. 1994;62:5603–7. doi: 10.1128/iai.62.12.5603-5607.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cowley SC, Elkins KL. Multiple T cell subsets control Francisella tularensis LVS intracellular growth without stimulation through macrophage interferon gamma receptors. J Exp Med. 2003;198:379–89. doi: 10.1084/jem.20030687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cowley SC, Gray CJ, Nano FE. Isolation and characterization of Francisella novicida mutants defective in lipopolysaccharide biosynthesis. FEMS Microbiol Lett. 2000;182:63–67. doi: 10.1111/j.1574-6968.2000.tb08874.x. [DOI] [PubMed] [Google Scholar]
- 37.Cowley SC, Myltseva SV, Nano FE. Phase variation in Francisella tularensis affecting intracellular growth, lipopolysaccharide antigenicity and nitric oxide production. Mol Microbiol. 1996;20:867–74. doi: 10.1111/j.1365-2958.1996.tb02524.x. [DOI] [PubMed] [Google Scholar]
- 38.Cowley SC, Myltseva SV, Nano FE. Suppression of Francisella tularensis growth in the rat by coinfection with F. novicida. FEMS Microbiol Lett. 1997;153:71–74. doi: 10.1111/j.1574-6968.1997.tb10465.x. [DOI] [PubMed] [Google Scholar]
- 39.Croddy E, Krcalova S. Tularemia, biological warfare, and the battle for Stalingrad (1942–1943) Mil Med. 2001;166:837–38. [PubMed] [Google Scholar]
- 40.Culkin SJ, Rhinehart-Jones T, Elkins KL. A novel role for B cells in early protective immunity to an intracellular pathogen, Francisella tularensis strain LVS. J Immunol. 1997;158:3277–84. [PubMed] [Google Scholar]
- 41.Dahlstrand S, Ringertz O, Zetterberg B. Airborne tularemia in Sweden. Scand J Infect Dis. 1971;3:7–16. doi: 10.3109/inf.1971.3.issue-1.02. [DOI] [PubMed] [Google Scholar]
- 42.Dennis DT, Inglesby TV, Henderson DA, Bartlett JG, Ascher MS, et al. Tularemia as a biological weapon: medical and public health management. JAMA. 2001;285:2763–73. doi: 10.1001/jama.285.21.2763. [DOI] [PubMed] [Google Scholar]
- 43.Duckett NS, Olmos S, Durrant DM, Metzger DW. Intranasal interleukin-12 treatment for protection against respiratory infection with the Francisella tularensis live vaccine strain. Infect Immun. 2005;73:2306–11. doi: 10.1128/IAI.73.4.2306-2311.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Elkins KL, Bosio CM, Rhinehart-Jones TR. Importance of B cells, but not specific antibodies, in primary and secondary protective immunity to the intracellular bacterium Francisella tularensis live vaccine strain. Infect Immun. 1999;67:6002–7. doi: 10.1128/iai.67.11.6002-6007.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Elkins KL, Cooper A, Colombini SM, Cowley SC, Kieffer TL. In vivo clearance of an intracellular bacterium, Francisella tularensis LVS, is dependent on the p40 subunit of interleukin-12 (IL-12) but not on IL-12 p70. Infect Immun. 2002;70:1936–48. doi: 10.1128/IAI.70.4.1936-1948.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Elkins KL, Rhinehart-Jones TR, Culkin SJ, Yee D, Winegar RK. Minimal requirements for murine resistance to infection with Francisella tularensis LVS. Infect Immun. 1996;64:3288–93. doi: 10.1128/iai.64.8.3288-3293.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Farlow J, Smith KL, Wong J, Abrams M, Lytle M, Keim P. Francisella tularensis strain typing using multiple-locus, variable-number tandem repeat analysis. J Clin Microbiol. 2001;39:3186–92. doi: 10.1128/JCM.39.9.3186-3192.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Forestal CA, Benach JL, Carbonara C, Italo JK, Lisinski TJ, Furie MB. Francisella tularensis selectively induces proinflammatory changes in endothelial cells. J Immunol. 2003;171:2563–70. doi: 10.4049/jimmunol.171.5.2563. [DOI] [PubMed] [Google Scholar]
- 49.Forsman M, Sandstrom G, Sjostedt A. Analysis of 16S ribosomal DNA sequences of Francisella strains and utilization for determination of the phylogeny of the genus and for identification of strains by PCR. Int J Syst Bacteriol. 1994;44:38–46. doi: 10.1099/00207713-44-1-38. [DOI] [PubMed] [Google Scholar]
- 50.Fortier AH, Leiby DA, Narayanan RB, Asafoadjei E, Crawford RM, et al. Growth of Francisella tularensis LVS in macrophages: The acidic intracellular compartment provides essential iron required for growth. Infect Immun. 1995;63:1478–83. doi: 10.1128/iai.63.4.1478-1483.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fortier AH, Polsinelli T, Green SJ, Nacy CA. Activation of macrophages for destruction of Francisella tularensis: identification of cytokines, effector cells, and effector molecules. Infect Immun. 1992;60:817–25. doi: 10.1128/iai.60.3.817-825.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Francis E. Sources of infection and seasonal incidence of tularemia in man. Public Health Rep. 1937;52:103–13. [Google Scholar]
- 53.Fulop M, Mastroeni P, Green M, Titball RW. Role of antibody to lipopolysaccharide in protection against low- and high-virulence strains of Francisella tularensis. Vaccine. 2001;19:4465–72. doi: 10.1016/s0264-410x(01)00189-x. [DOI] [PubMed] [Google Scholar]
- 54.Gao LY, Abu Kwaik Y. Hijacking of apoptotic pathways by bacterial pathogens. Microbes Infect. 2000;2:1705–19. doi: 10.1016/s1286-4579(00)01326-5. [DOI] [PubMed] [Google Scholar]
- 55.Gil H, Benach JL, Thanassi DG. Presence of pili on the surface of Francisella tularensis. Infect Immun. 2004;72:3042–47. doi: 10.1128/IAI.72.5.3042-3047.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gioannini TL, Teghanemt A, Zhang DS, Coussens NP, Dockstader W, et al. Isolation of an endotoxin-MD-2 complex that produces Toll-like receptor 4-dependent cell activation at picomolar concentrations. Proc Natl Acad Sci USA. 2004;101:4186–91. doi: 10.1073/pnas.0306906101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Golovliov I, Baranov V, Krocova Z, Kovarova H, Sjostedt A. An attenuated strain of the facultative intracellular bacterium Francisella tularensis can escape the phagosome of monocytic cells. Infect Immun. 2003;71:5940–50. doi: 10.1128/IAI.71.10.5940-5950.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Golovliov I, Ericsson M, Sandstrom G, Tarnvik A, Sjostedt A. Identification of proteins of Francisella tularensis induced during growth in macrophages and cloning of the gene encoding a prominently induced 23-kDa protein. Infect Immun. 1997;65:2183–89. doi: 10.1128/iai.65.6.2183-2189.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Golovliov I, Sandstrom G, Ericsson M, Sjostedt A, Tarnvik A. Cytokine expression in the liver during the early phase of murine tularemia. Infect Immun. 1995;63:534–38. doi: 10.1128/iai.63.2.534-538.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Golovliov I, Sjostedt A, Mokrievich A, Pavlov V. A method for allelic replacement in Francisella tularensis. FEMS Microbiol Lett. 2003;222:273–80. doi: 10.1016/S0378-1097(03)00313-6. [DOI] [PubMed] [Google Scholar]
- 61.Gray CG, Cowley SC, Cheung KK, Nano FE. The identification of five genetic loci of Francisella novicida associated with intracellular growth. FEMS Microbiol Lett. 2002;215:53–56. doi: 10.1111/j.1574-6968.2002.tb11369.x. [DOI] [PubMed] [Google Scholar]
- 62.Green SJ, Nacy CA, Schreiber RD, Granger DL, Crawford RM, et al. Neutralization of gamma interferon and tumor necrosis factor alpha blocks in vivo synthesis of nitrogen oxides from l-arginine and protection against Francisella tularensis infection in Mycobacterium bovis BCG-treated mice. Infect Immun. 1993;61:689–98. doi: 10.1128/iai.61.2.689-698.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Green SJ, Scheller LF, Marletta MA, Seguin MC, Klotz FW, et al. Nitric oxide: cytokine-regulation of nitric oxide in host resistance to intracellular pathogens. Immunol Lett. 1994;43:87–94. doi: 10.1016/0165-2478(94)00158-8. [DOI] [PubMed] [Google Scholar]
- 64.Harris R, Paxman J. A Higher Form of Killing. New York: Hill & Wang; 1982. p. 74. [Google Scholar]
- 65.Harris S. Japanese biological warfare research on humans: a case study of microbiology and ethics. Ann N Y Acad Sci. 1992;666:21–52. doi: 10.1111/j.1749-6632.1992.tb38021.x. [DOI] [PubMed] [Google Scholar]
- 66.Hartley G, Taylor R, Prior R, Newstead S, Hitchen PG, et al. Grey variants of the live vaccine strain of Francisella tularensis lack lipopolysaccharide O-antigen, show reduced ability to survive in macrophages and do not induce protective immunity in mice. Vaccine. 2006;24:989–96. doi: 10.1016/j.vaccine.2005.08.075. [DOI] [PubMed] [Google Scholar]
- 67.Hollis DG, Weaver RE, Steigerwalt AG, Wenger JD, Moss CW, Brenner DJ. Francisella philomiragia comb. nov (formerly Yersinia philomiragia) and Francisella tularensis biogroup novicida (formerly Francisella novicida) associated with human disease . J Clin Microbiol. 1989;27:1601–8. doi: 10.1128/jcm.27.7.1601-1608.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hood AM. Virulence factors of Francisella tularensis. J Hyg. 1977;79:47–60. doi: 10.1017/s0022172400052840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Jellison WL, Kohls GM. Tularemia in sheep and sheep industry workers in western United States. Public Health Serv. 1955:17. doi: 10.5962/bhl.title.117381. [DOI] [PubMed] [Google Scholar]
- 70.Karpov SP. The spread of tularemia through water as a new factor in its epidemiology. J Bacteriol. 1936;32:243–58. doi: 10.1128/jb.32.3.243-258.1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kawula TH, Hall JD, Fuller JR, Craven RR. Use of transposon-transposase complexes to create stable insertion mutant strains of Francisella tularensis LVS. Appl Environ Microbiol. 2004;70:6901–4. doi: 10.1128/AEM.70.11.6901-6904.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kieffer TL, Cowley S, Nano FE, Elkins KL. Francisella novicida LPS has greater immunobiological activity in mice than F. tularensis LPS, and contributes to F. novicida murine pathogenesis. Microbes Infect. 2003;5:397–403. doi: 10.1016/s1286-4579(03)00052-2. [DOI] [PubMed] [Google Scholar]
- 73.Kuoppa K, Forsberg A, Norqvist A. Construction of a reporter plasmid for screening in vivo promoter activity in Francisella tularensis. FEMS Microbiol Lett. 2001;205:77–81. doi: 10.1111/j.1574-6968.2001.tb10928.x. [DOI] [PubMed] [Google Scholar]
- 74.Lai XH, Golovliov I, Sjostedt A. Francisella tularensis induces cytopathogenicity and apoptosis in murine macrophages via a mechanism that requires intracellular bacterial multiplication. Infect Immun. 2001;69:4691–94. doi: 10.1128/IAI.69.7.4691-4694.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Lai XH, Golovliov I, Sjostedt A. Expression of IglC is necessary for intracellular growth and induction of apoptosis in murine macrophages by Francisella tularensis. Microb Pathog. 2004;37:225–30. doi: 10.1016/j.micpath.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 76.Lai XH, Sjostedt A. Delineation of the molecular mechanisms of Francisella tularensis-induced apoptosis in murine macrophages. Infect Immun. 2003;71:4642–46. doi: 10.1128/IAI.71.8.4642-4646.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Larsson P, Oyston PC, Chain P, Chu MC, Duffield M, et al. The complete genome sequence of Francisella tularensis, the causative agent of tularemia. Nat Genet. 2005;37:153–59. doi: 10.1038/ng1499. [DOI] [PubMed] [Google Scholar]
- 78.Lauriano CM, Barker JR, Yoon SS, Nano FE, Arulanandam BP, et al. MglA regulates transcription of virulence factors necessary for Francisella tularensis intraamoebae and intramacrophage survival. Proc Natl Acad Sci USA. 2004;101:4246–49. doi: 10.1073/pnas.0307690101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Leiby DA, Fortier AH, Crawford RM, Schreiber RD, Nacy CA. In vivo modulation of the murine immune response to Francisella tularensis LVS by administration of anticytokine antibodies. Infect Immun. 1992;60:84–89. doi: 10.1128/iai.60.1.84-89.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lesho E, Dorsey D, Bunner D. Feces, dead horses, and fleas: evolution of the hostile use of biological agents. West J Med. 1998;168:512–16. [PMC free article] [PubMed] [Google Scholar]
- 81.Lindgren H, Golovliov I, Baranov V, Ernst RK, Telepnev M, Sjostedt A. Factors affecting the escape of Francisella tularensis from the phagolysosome. J Med Microbiol. 2004;53:953–58. doi: 10.1099/jmm.0.45685-0. [DOI] [PubMed] [Google Scholar]
- 82.Lindgren H, Stenmark S, Chen W, Tarnvik A, Sjostedt A. Distinct roles of reactive nitrogen and oxygen species to control infection with the facultative intracellular bacterium Francisella tularensis. Infect Immun. 2004;72:7172–82. doi: 10.1128/IAI.72.12.7172-7182.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lofgren S, Tarnvik A, Bloom GD, Sjoberg W. Phagocytosis and killing of Francisella tularensis by human polymorphonuclear leukocytes. Infect Immun. 1983;39:715–20. doi: 10.1128/iai.39.2.715-720.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lofgren S, Tarnvik A, Carlsson J. Demonstration of opsonizing antibodies to Francisella tularensis by leukocyte chemiluminescence. Infect Immun. 1980;29:329–34. doi: 10.1128/iai.29.2.329-334.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Lofgren S, Tarnvik A, Thore M, Carlsson J. A wild and an attenuated strain of Francisella tularensis differ in susceptibility to hypochlorous acid: a possible explanation of their different handling by polymorphonuclear leukocytes. Infect Immun. 1984;43:730–34. doi: 10.1128/iai.43.2.730-734.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Maier TM, Havig A, Casey M, Nano FE, Frank DW, Zahrt TC. Construction and characterization of a highly efficient Francisella shuttle plasmid. Appl Environ Microbiol. 2004;70:7511–19. doi: 10.1128/AEM.70.12.7511-7519.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Mariathasan S, Weiss DS, Dixit VM, Monack DM. Innate immunity against Francisella tularensis is dependent on the ASC/caspase-1 axis. J Exp Med. 2005;202:1043–49. doi: 10.1084/jem.20050977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.McCoy GW, Chapin CW. Further observations on a plague-like diseases of rodents with a preliminary note on the causative agent, Bacterium tularensis. J Infect Dis. 1912;10:61–72. [Google Scholar]
- 89.McDonald MK, Cowley SC, Nano FE. Temperature-sensitive lesions in the Francisella novicida valA gene cloned into an Escherichia coli msbA lpxK mutant affecting deoxycholate resistance and lipopolysaccharide assembly at the restrictive temperature. J Bacteriol. 1997;179:7638–43. doi: 10.1128/jb.179.24.7638-7643.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Mdluli KE, Anthony LS, Baron GS, McDonald MK, Myltseva SV, Nano FE. Serum-sensitive mutation of Francisella novicida: association with an ABC transporter gene. Microbiology. 1994;140:3309–18. doi: 10.1099/13500872-140-12-3309. [DOI] [PubMed] [Google Scholar]
- 91.Nano FE, Zhang N, Cowley SC, Klose KE, Cheung KK, et al. A Francisella tularensis pathogenicity island required for intramacrophage growth. J Bacteriol. 2004;186:6430–36. doi: 10.1128/JB.186.19.6430-6436.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Nauseef WM. Contributions of myeloperoxidase to proinflammatory events: more than an antimicrobial system. Int J Hematol. 2001;74:125–33. doi: 10.1007/BF02981994. [DOI] [PubMed] [Google Scholar]
- 93.Nauseef WM. Assembly of the phagocyte NADPH oxidase. Histochem Cell Biol. 2004;122:277–91. doi: 10.1007/s00418-004-0679-8. [DOI] [PubMed] [Google Scholar]
- 94.Norqvist A, Kuoppa K, Sandstrom G. Construction of a shuttle vector for use in Francisella tularensis. FEMS Immunol Med Microbiol. 1996;13:257–60. doi: 10.1111/j.1574-695X.1996.tb00248.x. [DOI] [PubMed] [Google Scholar]
- 95.Nudleman E, Kaiser D. Pulling together with type IV pili. J Mol Microbiol Biotechnol. 2004;7:52–62. doi: 10.1159/000077869. [DOI] [PubMed] [Google Scholar]
- 96.Pavlov VM, Mokrievich AN, Volkovoy K. Cryptic plasmid pFNL10 from Francisella novicida-like F6168: the base of plasmid vectors for Francisella tularensis. FEMS Immunol Med Microbiol. 1996;13:253–56. doi: 10.1111/j.1574-695X.1996.tb00247.x. [DOI] [PubMed] [Google Scholar]
- 97.Penn RL. Francisella tularensis (tularemia) In: Mandell GL, Bennett JE, Dolin R, editors. Principles and Practice of Infectious Diseases. 6 New York: Churchill Livingstone; 2005. pp. 2674–85. [Google Scholar]
- 98.Phillips NJ, Schilling B, McLendon MK, Apicella MA, Gibson BW. Novel modification of lipid A of Francisella tularensis. Infect Immun. 2004;72:5340–48. doi: 10.1128/IAI.72.9.5340-5348.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Polsinelli T, Meltzer MS, Fortier AH. Nitric oxide-independent killing of Francisella tularensis by IFN-gamma-stimulated murine alveolar macrophages. J Immunol. 1994;153:1238–45. [PubMed] [Google Scholar]
- 100.Pomerantsev AP, Obuchi M, Ohara Y. Nucleotide sequence, structural organization, and functional characterization of the small recombinant plasmid pOM1 that is specific for Francisella tularensis. Plasmid. 2001;46:86–94. doi: 10.1006/plas.2001.1538. [DOI] [PubMed] [Google Scholar]
- 101.Prior JL, Prior RG, Hitchen PG, Diaper H, Griffin KF, et al. Characterization of the O antigen gene cluster and structural analysis of the O antigen of Francisella tularensis subsp tularensis. J Med Microbiol. 2003;52:845–51. doi: 10.1099/jmm.0.05184-0. [DOI] [PubMed] [Google Scholar]
- 102.Proctor RA, White JD, Ayala E, Canonico PG. Phagocytosis of Francisella tularensis by Rhesus monkey peripheral leukocytes. Infect Immun. 1975;11:146–51. doi: 10.1128/iai.11.1.146-151.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Raetz CR, Whitfield C. Lipopolysaccharide endotoxins. Annu Rev Biochem. 2002;71:635–700. doi: 10.1146/annurev.biochem.71.110601.135414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Reilly TJ, Baron GS, Nano FE, Kuhlenschmidt MS. Characterization and sequencing of a respiratory burst-inhibiting acid phosphatase from Francisella tularensis. J Biol Chem. 1996;271:10973–83. doi: 10.1074/jbc.271.18.10973. [DOI] [PubMed] [Google Scholar]
- 104a.Report of a WHO Group of Consultants. Health Aspects of Chemical and Biological Weapons. Geneva, Switzerland: World Health Org; 1970. p. 132. [Google Scholar]
- 105.Sandstrom G, Lofgren S, Tarnvik A. A capsule-deficient mutant of Francisella tularensis LVS exhibits enhanced sensitivity to killing by serum but diminished sensitivity to killing by polymorphonuclear leukocytes. Infect Immun. 1988;56:1194–202. doi: 10.1128/iai.56.5.1194-1202.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Sandstrom G, Sjostedt A, Johansson T, Kuoppa K, Williams JC. Immunogenicity and toxicity of lipopolysaccharide from Francisella tularensis LVS. FEMS Microbiol Immunol. 1992;5:201–10. doi: 10.1111/j.1574-6968.1992.tb05902.x. [DOI] [PubMed] [Google Scholar]
- 107.Santic M, Molmeret M, Abu Kwail Y. Modulation of biogenesis of the Francisella tularensis subsp novicida-containing phagosome in quiescent human macrophages and its maturation into a phagolysosome upon activation by IFN-γ. Cell Microbiol. 2005;7:957–67. doi: 10.1111/j.1462-5822.2005.00529.x. [DOI] [PubMed] [Google Scholar]
- 108.Santic M, Molmeret M, Klose KE, Jones S, Abu Kwail Y. The Francisella tularensis pathogenicity island protein IglC and its regulator MglA are essential for modulating phagosome biogenesis and subsequent bacterial escape into the cytoplasm. Cell Microbiol. 2005;7:969–79. doi: 10.1111/j.1462-5822.2005.00526.x. [DOI] [PubMed] [Google Scholar]
- 109.Saslaw S, Eigelsbach HT, Prior JA, Wilson HE, Carhart S. Tularemia vaccine study. II Respiratory challenge. Arch Intern Med. 1961;107:702–14. doi: 10.1001/archinte.1961.03620050068007. [DOI] [PubMed] [Google Scholar]
- 110.Saslaw S, Eigelsbach HT, Wilson HE, Prior JA, Carhart S. Tularemia vaccine study. I Intracutaneous challenge. Arch Intern Med. 1961;107:689–701. doi: 10.1001/archinte.1961.03620050055006. [DOI] [PubMed] [Google Scholar]
- 111.Sjostedt A, Conlan JW, North RJ. Neutrophils are critical for host defense against primary infection with the facultative intracellular bacterium Francisella tularensis in mice and participate in defense against reinfection. Infect Immun. 1994;62:2779–83. doi: 10.1128/iai.62.7.2779-2783.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Sorokin VM, Pavlovich NV, Prozorova LA. Francisella tularensis resistance to bactericidal action of normal human serum. FEMS Immunol Med Microbiol. 1996;13:249–52. doi: 10.1111/j.1574-695X.1996.tb00246.x. [DOI] [PubMed] [Google Scholar]
- 113.Svensson K, Larsson P, Johansson D, Bystrom M, Forsman M, Johansson A. Evolution of subspecies of Francisella tularensis. J Bacteriol. 2005;187:3903–8. doi: 10.1128/JB.187.11.3903-3908.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Takafuji ET. Biological Weapons and Modern Warfare. Washington, DC: Ind. Coll. Armed Forces, Natl. Def. Univ., Ft. McNair; 1991. [Google Scholar]
- 115.Tarnvik A. Nature of protective immunity to Francisella tularensis. Rev Infect Dis. 1989;11:440–51. [PubMed] [Google Scholar]
- 116.Teghanemt A, Zhang DS, Levis EN, Weiss JP, Gioannini TL. Molecular basis of reduced potency of underacylated endotoxins. J Immunol. 2005;175:4669–76. doi: 10.4049/jimmunol.175.7.4669. [DOI] [PubMed] [Google Scholar]
- 117.Telepnev M, Golovliov I, Grundstrom T, Tarnvik A, Sjostedt A. Francisella tularensis inhibits Toll-like receptor-mediated activation of intracellular signaling and secretion of TNF-alpha and IL-1 from murine macrophages. Cell Microbiol. 2003;5:41–51. doi: 10.1046/j.1462-5822.2003.00251.x. [DOI] [PubMed] [Google Scholar]
- 118.Telepnev M, Golovilov I, Sjostedt A. Francisella tularensis LVS initially activates but subsequently down-regulates intracellular signaling and cytokine secretion in mouse monocytic and human peripheral blood mononuclear cells. Microb Pathog. 2005;38:239–47. doi: 10.1016/j.micpath.2005.02.003. [DOI] [PubMed] [Google Scholar]
- 119.Thirumalapura NR, Goad DW, Mort A, Morton RJ, Clarke J, Malayer J. Structural analysis of the O-antigen of Francisella tularensis subspecies tularensis strain OSU 10. J Med Microbiol. 2005;54:693–95. doi: 10.1099/jmm.0.45931-0. [DOI] [PubMed] [Google Scholar]
- 120.Thomas R, Johansson A, Neeson B, Isherwood K, Sjostedt A, et al. Discrimination of human pathogenic subspecies of Francisella tularensis by using restriction fragment length polymorphism. J Clin Microbiol. 2003;41:50–57. doi: 10.1128/JCM.41.1.50-57.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Thomassen MJ, Kavuru MS. Human alveolar macrophages and monocytes as a source and target for nitric oxide. Int Immunopathol. 2001;1:1479–90. doi: 10.1016/s1567-5769(01)00092-3. [DOI] [PubMed] [Google Scholar]
- 122.Twine S, Bystrom M, Chen W, Forsman M, Golovliov I, et al. A mutant of Francisella tularensis strain SCHU S4 lacking the ability to express a 58-kilodalton protein is attenuated for virulence and is an effective live vaccine. Infect Immun. 2005;73:8345–52. doi: 10.1128/IAI.73.12.8345-8352.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Tyeryar FJJ, Lawton WD. Transformation of Pasteurella novicida. J Bacteriol. 1969;100:1112–13. doi: 10.1128/jb.100.2.1112-1113.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Vinogradov E, Perry MB. Characterization of the core part of the lipopolysaccharide O-antigen of Francisella novicida (U112) Carbohydr Res. 2004;339:1643–48. doi: 10.1016/j.carres.2004.04.013. [DOI] [PubMed] [Google Scholar]
- 125.Vinogradov E, Perry MB, Conlan JW. Structural analysis of Francisella tularensis lipopolysaccharide. Eur J Biochem. 2002;269:6112–18. doi: 10.1046/j.1432-1033.2002.03321.x. [DOI] [PubMed] [Google Scholar]
- 126.Vinogradov EV, Shashkov AS, Knirel YA, Kochetkov NK, Tochtamysheva NV, et al. Structure of the O-antigen of Francisella tularensis strain 15. Carbohydr Res. 1991;214:289–97. doi: 10.1016/0008-6215(91)80036-m. [DOI] [PubMed] [Google Scholar]
- 127.Wang X, Karbarz MJ, McGrath SC, Cotter RJ, Raetz CR. MsbA transporter-dependent lipid A 1-dephosphorylation on the periplasmic surface of the inner membrane: topography of Francisella novicida LpxE expressed in Escherichia coli. J Biol Chem. 2004;279:49470–78. doi: 10.1074/jbc.M409078200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Yee D, Rhinehart-Jones TR, Elkins KL. Loss of either CD4+ or CD8+ T cells does not affect the magnitude of protective immunity to an intracellular pathogen, Francisella tularensis strain LVS. J Immunol. 1996;157:5042–48. [PubMed] [Google Scholar]