

Abstract
The 5′ flanking sequences and the intron of the mouse Hoxa-7 gene were searched for regulatory elements that can function in Drosophila. Only the intron is able to activate a lacZ fusion gene in various tissues of Drosophila embryos. This enhancer function requires a cluster of three homeodomain binding sites (HB1-element) that are also found in the introns of other Hox genes as well as in a putative autoregulatory element of Ultrabithorax (Ubx), the Drosophila homolog of Hoxa-7. If a single binding site in the autoregulatory element of fushi tarazu (ftz) is replaced by the HB1-element of Hoxa-7, the expression pattern is altered and newly controlled by the homeotic gene caudal (cad). These data suggest that HB1 is a potential target for different homeodomain proteins of both vertebrates and invertebrates.
Homeotic genes of vertebrates and invertebrates determine cell fates along the anterior-posterior body axis (1). The homeobox encodes a conserved DNA binding domain, the homeodomain (2–16), which is required for the function of the homeotic proteins as transcription factors in vivo (17–27). The high degree of evolutionary conservation of the homeodomain suggests that the DNA binding sites recognized by the homeodomains might also be conserved and that some of the regulatory circuits may be the same in both mammals and insects. It has been shown that the mammalian proteins can mimic the function of their Drosophila counterparts and produce the same phenotypes when expressed ectopically in flies (28–32). The mouse Hoxb-6 gene, the homolog of Antennapedia (Antp), can induce antenna to leg transformations similar to Antp (28), whereas the mouse Pax-6 gene, the homolog of the eyeless (ey) gene, is able to induce ectopic eye formation in the fly (32). In addition, Hoxd-4 control regions were shown to exhibit enhancer activity in Drosophila similar to Deformed (Dfd), whereas a Dfd autoregulatory element was active in the expression domain of the paralog group 4 genes in the mouse, suggesting that this pathway may be conserved between vertebrates and invertebrates (33, 34). Since some of the homeobox genes are autoregulated (Ubx; ref. 35); Dfd (36); ftz (22); and crossregulated (Antp by Ubx; refs. 37 and 38), Ubx by abd-A (39, 40), the control regions of the mouse Hox genes were expected, by analogy to their Drosophila homologs, to contain functional homeodomain binding sites. Therefore, the genomic region of the mouse Hoxa-7 gene (41, 42), the homolog of the Drosophila Ubx gene, was analyzed. It has been shown that 7 kb of genomic region are sufficient in transgenic mice for the early expression in the spinal cord and the somites (43, 44). The primary control elements for activation are located within a 469-bp fragment 1.6 kb upstream of the transcription start site, whereas the intron is needed for repression in sclerotomal cells in the somites (43–45). Thus, we tested whether the upstream and intronic regions exhibit enhancer activity in Drosophila. A 680-bp and a larger 4-kb upstream fragment as well as an intron fragment were cloned in front of a lacZ reporter gene under the control of an hsp70 minimal promoter in the transformation vector Hz50pl (48), and lacZ expression was analyzed in transformed embryos.
MATERIALS AND METHODS
Fly Strains.
The rosy506-strain (ry) was used as recipient for injections. Antpw10, Ubx1, cad1, cad2, and cad3 mutant alleles were used in this study. The following heat shock strains were used: HS-AntpH4 (17), HS-UbxIa22/TM6B (18), HTcad-1 (46), and HS-ftzT1 (U. Grossniklaus and K. Cadigan, personal communication).
Plasmid Constructs.
Further details of the cloning procedures are available on request (47). All constructs were cloned in the same orientation into Hz50pl (48) except HA1, which was cloned in RS27 (47). p40 contains a 4-kb PstI fragment, p7 a 680-bp AluI–Sau3A fragment, and PB6 a PstI–BamHI fragment from pm6Δ (43).
β-Galactosidase (β-gal) activity staining was performed as described by Freeman et al. (49).
Antibody stainings were performed as described by Frasch et al. (50) using the Vectastain kit (Vector Laboratories). A monoclonal serum against β-gal was used from Promega. The antiserum against the engrailed (en) gene product was a gift from N. Patel. The antiserum against CAD was described by Mlodzik et al. (51).
Heat Shocks.
Virgins of the heat shock lines were crossed with males homozygous for the lacZ transgene. Two- to 6-h-old embryos were collected and heat-shocked for 20 min at 37°C, incubated at 18°C for 7 h, and fixed.
Mobility-Shift Assays.
ANTP and FTZ shifts were described previously (2, 52). The samples were electrophoresed at 4°C on native, 6% polyacrylamide gels. ANTP and FTZ homeodomains were purified from bacterial extracts (2, 52). CAD protein was a gift of R. Rivera-Pomar and H. Jäckle (53). The sequence of the HB1 oligonucleotides were as follows: HB1-5, 5′-TTGCTCATAAAACTTTTTATGGCCCAATTAATGGGTTC-3′; and SP3, 5′-GAACCCATTAATTGGGCCATAAAAAGTTTTATGA-3′.
RESULTS
The Hoxa-7 5′ Control Region Is Not Functional in Drosophila.
We found that the fragments from the upstream region (p7 and p40, Fig. 1A) inserted into the Hz50pl vector only exhibited enhancer activity within epidermal cells in the maxillary segment at the time of head involution (Fig. 2A). This activity, although on average weaker, resembles that attributed to the Hox d-4 upstream element that was previously studied (33). Analyzing transformants of the Hz50pl vector without any insert revealed that almost all lines (29/33) showed the same expression pattern (Fig. 2B). This result shows that this enhancer activity depends on vector sequences alone and suggests that the Hoxa-7 upstream enhancers are not functional in Drosophila.
Figure 1.
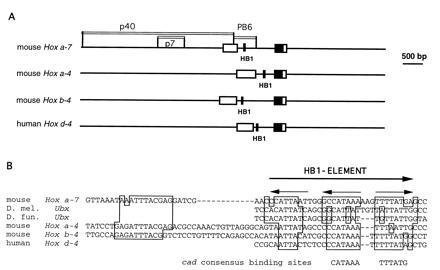
A conserved cluster of homeodomain binding sites in vertebrate and invertebrate homeobox genes. (A) Location of the p40, p7, and PB6 fragments and the HB1 elements. The ORFs of the Hox genes are indicated by boxes, the homeoboxes are black. (B) Three conserved homeodomain binding sites (HB1-element) are present in the introns of the mouse Hoxa-7, Hoxa-4, Hoxb-4, and the human Hoxd-4 genes as well as in the Ubx gene of two Drosophila species. The sequence of Hoxa-7 is shown in the opposite orientation. An additional conserved element is similar to the in vitro consensus site for the Pax-6 paired domain (61).
Figure 2.
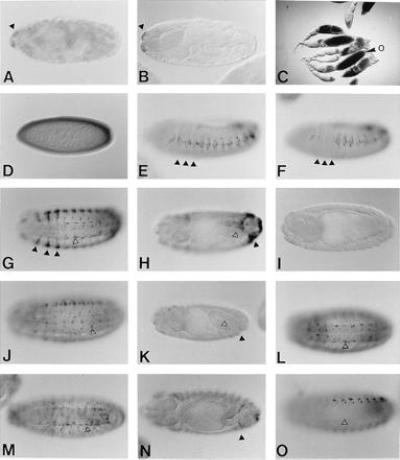
Enhancer activity of the Hoxa-7 intron in Drosophila. Stainings of embryos with antiserum against β-gal, if not indicated differently. (A) Stage 17 embryo of p40 showing epidermal expression in the maxillary segment (arrowhead). (B) Embryo of a control Hz50pl line without insert shows the same expression as A. (C) β-Gal activity staining of PB6 ovaries. Strong staining is obtained in the nurse cells from at stage 9 on and later in the oocyte (o). (D) Stage 5 PB6 embryo showing maternal expression. (E–N) Stage 13–15 embryos (E) PS4 embryo showing expression in the lateral epidermis. The thoracic segments are indicated by arrowheads. (F) PS4 embryo in a presumptive Antpw10 mutant background. Expression in the lateral epidermis in the three thoracic segments is missing (arrowheads). (G and H) XS3 embryos showing ventrolateral expression (G) and expression in the posterior segments (H). The zygotic staining in PS4, XS3, and FS2 lines is identical to PB6. (I) HB1 embryos show no expression. (J and K) XO3 embryos showing staining in cells of the ventral and peripheral nervous system and in some lateral epidermal cells (arrowhead) (J), but not in the cells around the posterior midgut (open arrowhead) and in the posterior segments (solid arrowhead) (K). (L) P1 embryo showing staining in the ventral nervous system. (M and N) ON3 embryo showing the same staining as P1 (M) as well as weak staining in the posterior segments (N). (O) HA1 embryo showing similar (nuclear) staining as HS1, but the P1 pattern (L) in the ventral nervous system is absent (arrowhead).
The Intron of Hoxa-7 Directs Expression in Drosophila Embryos.
The enhancer activity of a 600-bp intron fragment of Hoxa-7, containing clusters of putative homeodomain binding sites, was analyzed in Drosophila (PB6, Fig. 1A). Many of the PB6 transgenic lines (6/11) showed maternal expression; early expression in the nurse cells subsequently in the oocytes (Fig. 2C) and in early embryos (Fig. 2D). All the lines showed zygotic expression starting shortly after germ band retraction. Strong staining appeared in lateral and dorsolateral cells in the epidermis and in the peripheral nervous system with enhanced expression in the three thoracic segments (Fig. 2 E and G). Strong staining was also observed in the posterior abdominal segments in epidermal cells and in mesodermal cells around the invaginating hindgut (Fig. 2H).
To identify the sequences that can function as enhancer elements, a deletion analysis of the PB6 fragment was performed (Fig. 3). The enhancer activity of the maternal and the zygotic expression was found to be located in different regions. None of the 3′ deletion constructs PS4, XS3, and FS2 showed maternal enhancer activity, whereas the zygotic activity was identical to PB6. However, XO3, a 3′ deletion of 40 bp from XS3 including a cluster of three putative homeodomain binding sites, the HB1-element, resulted in the loss of the strong zygotic activity (compare Fig. 2 G and H with J and K). XO3 showed activity only in the ventral nervous system and in some cells of the lateral ectoderm (Fig. 2J).
Figure 3.
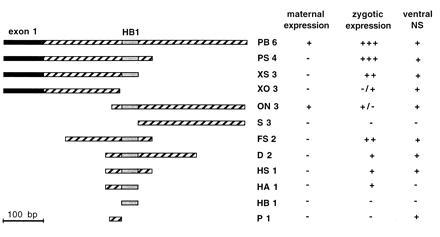
Enhancer activity of deletion constructs. ▪, Exon 1 part; ▨, intron; ░⃞, HB1-element. PS4, FS2, and HS1 have the same 3′ ends, whereas D2, HS1, and HA1 have the same 5′ ends.
On the other hand, several (4/10) of the ON3 lines that contained a 5′ deletion construct showed expression in the nurse cells, whereas only very weak zygotic expression was observed (Fig. 2 M and N). S3, a 5′ deletion lacking the HB1-element, showed neither maternal nor zygotic activity (data not shown). These results suggested that the HB1-element is required for both maternal and zygotic expression. But the HB1-element by itself was not sufficient to activate lacZ expression in any Drosophila tissue (Fig. 2I).
Finally, it was noted that both XO3 and ON3 showed enhancer activity in the ventral nervous system. The common sequence of ON3 and XO3, P1, was found to be sufficient for this activity (Fig. 2L). However, the construct HA1 in which the P1 sequence was cloned in front of an hsp27 promoter, did not show this expression pattern (Fig. 2O). Therefore, we conclude that P1 is functional in the context of the hsp70 promoter and inactive in the combination with the hsp27 promoter.
Mobility-shift assays showed that three molecules of purified ANTP homeodomain are able to bind to the HB1-element in vitro (Fig. 4A). Therefore, we analyzed the zygotic expression of PS4 lines in the mutant background of homeotic selector genes. We found that the early expression in the lateral epidermis was missing in all three thoracic segments in Antpw10 mutant embryos (compare Fig. 2 E with F). However, staining in this region was observed at later stages. In Ubx1 mutant background, expression in the lateral epidermis was only slightly decreased, suggesting that some other activators are present within the Ubx expression domain (data not shown). The staining in the posterior segments that resembles the expression of the homeobox gene caudal (cad, 46, 51) was, however, not altered in cad mutant background. Overexpression of Antp, Ubx, and cad by heat shock on the other hand did not show any significant change in the expression pattern.
Figure 4.
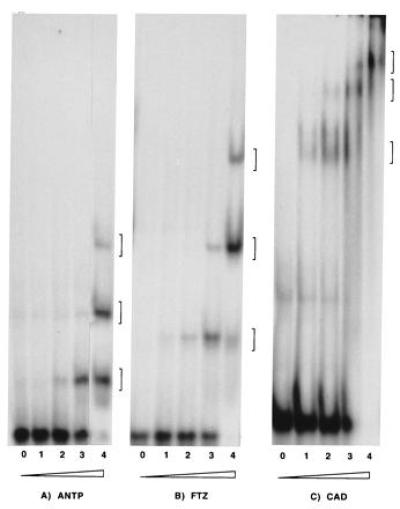
Mobility-shift assays of homeodomain-DNA binding. (A) Binding of a 68-amino acid long (8 kDa) ANTP homeodomain to the HB1 sequence. (B) Binding of a 70 amino acid long FTZ homeodomain to HB1. (C) Binding of a 46 kDa CAD protein to HB1. Lanes 0 contain no protein. Concentration of the ANTP and FTZ homeodomains in lanes 1 is 0.8 nM, that of CAD is 100 nM. The concentration of ANTP and FTZ homeodomain increase with each lane by a factor of 4, that of CAD by a factor of 3. Three complexes are observed with all three proteins.
Elements Similar to HB1 Are Present in the Introns of other Hox Genes as well as in a Putative Ubx Autoregulatory Element.
Sequences related to HB1 were found in the introns of Hoxa-4, b-4, and d-4. (Fig. 1B; refs. 54–56). They are located in the intron approximately 200 bp downstream of the 5′ splice site (Fig. 6B), but the HB1-element of Hoxa-7 is in the opposite orientation. An HB1 related sequence was also found in the control region of Ubx, the Drosophila homolog of Hoxa-7. This sequence is located downstream of the transcription start site and is conserved between Drosophila melanogaster and D. funebris (57). This sequence was shown to be required for the expression of Ubx in parasegment 7 in the visceral mesoderm (35). The strong sequence conservation of HB1 suggests an important functional role of this element.
Figure 6.
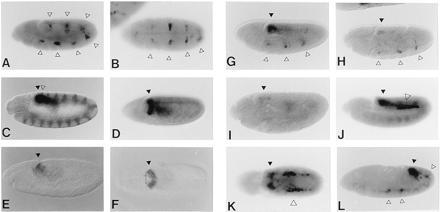
Expression patterns of AE20A, 4A7ftz and 3A7ftz lines in Drosophila embryos. β-Gal activity stainings (A–D and G–L) and stainings with an antiserum against EN (C) and CAD (E and F). Lateral view (A) and ventral view (B) of a stage 11 AE20A embryo showing staining in alternate segments (arrowheads). Lateral view (C) and dorsal view (D) of an early stage 11 4A7ftz embryo. (C) The β-gal staining (solid arrowhead) appears more posteriorly of the last en-stripe (open arrowhead). (E and F) Stage 10 (E) and stage 11 (F) embryo stained with antiserum against the cad gene product (arrowheads). (G) Early stage 11 3A7ftz embryo showing ftz-like (open arrowheads) and cad-like (solid arrowhead) expression. (H) Early stage 11 3A7ftz embryo in a presumptive cad1 mutant background. The cad-like expression (solid arrowhead) is almost absent, whereas the ftz-like expression (open arrowheads) is not affected. (I) 4A7ftz embryo in a presumptive cad3 mutant background showing reduced staining (solid arrowhead). (J) Lateral view and (K) dorsal view. Staining of a late stage 11 4A7ftz embryo after ubiquitous expression of cad. Ectopic expression appears in lateral mesodermal cells (open arrowhead). (L) Early stage 12 4A7ftz embryo after ubiquitous expression of ftz showing ftz-like expression (open arrowheads).
Functional Analysis of the HB1-Element in the Context of the ftz Autoregulatory Element.
To find out whether the HB1-element is a direct target for homeodomain proteins, it was analyzed in the context of the ftz autoregulatory element. The ftz gene is expressed in seven stripes in alternate parasegments. In a second-site suppression experiment, direct interaction between the FTZ protein and the ftz autoregulatory element has been demonstrated (22). This ftz enhancer contains redundant FTZ binding sites located in four regions (58). We defined a minimal element, AE20A (Fig. 5B), which exhibits enhancer activity in alternate segments in the mesoderm (Fig. 6 A and B). This 154 bp long fragment contains only one FTZ binding site as previously shown by footprint analysis (59).
Figure 5.
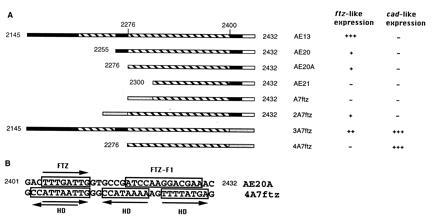
Analysis of the HB1-element in the context of the ftz enhancer. (A) Deletion constructs of the ftz autoregulatory element and fusion constructs to HB1. ▪, FTZ binding sites; ░⃞, Hoxa-7 HB1-elements. AE13 contains several FTZ binding sites, AE20A a single binding site. Both exhibit enhancer activity in alternate segments in the mesoderm. AE21 is not functional. In 4A7ftz, the FTZ binding site and a binding site for FTZ-F1 (□) has been replaced by the HB1-element. (B) Sequence comparison of the ftz sequence in AE20A and the homeodomain binding sites in 4A7ftz. Binding sites for FTZ and FTZ-F1 as well as the homeodomain binding sites (HD) are indicated.
Mobility-shift assays indicate that three molecules of purified FTZ-homeodomain can bind to the HB1-element in vitro (Fig. 4B). Thus, the single FTZ binding site and a putative binding site for FTZ-F1 gene product (22, 58, 60) were replaced in AE20A by the three ftz binding sites of HB1 (4A7ftz, Fig. 5 A and B). If the HB1 binding sites were recognized by FTZ in vivo, expression in alternate segments as in AE20A would have been expected.
However, the 4A7ftz lines showed no ftz-like expression. Instead, at the stage of complete germ band extension, staining was seen in the last abdominal segment (Fig. 6 C and D). This pattern resembles the expression of the homeobox gene cad (Fig. 6 E and F) but the expression is restricted to the mesoderm. In 3A7 ftz lines (Fig. 5A) the same expression pattern was superimposed on a ftz-like expression pattern (Fig. 6G). This enhancer contains additional FTZ binding sites and was therefore expected to activate lacZ in seven stripes. If HB1 is located 5′ of AE21 or AE20A, replacing the 5′ FTZ binding site of AE20 (Fig. 5A; A7ftz and 2A7ftz), no cad-like expression is observed (data not shown), suggesting that the position of the HB1 element is important.
A mobility-shift assay showed that three molecules of CAD protein are able to bind to the HB1-element in vitro (Fig. 4C). To demonstrate interaction in vivo, 3A7ftz and 4A7ftz lines were analyzed in cad zygotic mutant embryos (cad1, cad2, and cad3). In this background, the expression in 3A7ftz and 4A7ftz embryos is almost absent in the cad expression domain (Fig. 6 H and I). In contrast, ubiquitous expression of cad by heat shock (pHTcad-1), induced ectopic expression in lateral mesodermal cells in 4A7ftz (Fig. 6 J and K) but not in AE20A embryos (data not shown). This ectopic activation was not observed in control experiments using AntpH4 or UbxIa22 heat shock lines (data not shown).
The expression of the 4A7ftz embryos suggested that FTZ was not able to activate the transgene in these lines. When FTZ protein was overexpressed by heat shock induction in 4A7ftz lines (HS-ftzT1), ftz-like expression was observed in addition to the cad-dependent expression (Fig. 6L). Therefore, we conclude that FTZ is still able to activate the transgene in 4A7ftz lines, but activation requires high levels of FTZ protein. Thus, the HB1-element contains potential direct target sites for both CAD and FTZ proteins.
DISCUSSION
The HB1-Element Is a Potential Target Motif for CAD and FTZ.
The HB1-element is an evolutionarily conserved DNA sequence in the intron of Hoxa-7, in the leader of its Drosophila homolog Ubx, and in the introns of the paralog group 4 Hox genes. The analysis of the HB1-element in the Ubx gene revealed that the first two binding sites of HB1 are required for Ubx expression in the visceral mesoderm and it was suggested that they may function as an autoregulatory element (35). We have shown that three molecules of ANTP, FTZ, or CAD homeodomains can bind to the HB1-element in vitro (Fig. 4) and that ftz regulates HB1 in the context of the 120-bp ftz autoregulatory sequence (Fig. 6L). It has been previously shown that the ftz autoregulatory enhancer is directly controlled by FTZ (16). Direct FTZ interaction with the binding site in AE20A is very likely, although it has not been demonstrated in vivo. Since there are no other binding sites present in 4A7ftz as previously shown by footprint analysis (59), we can assume that FTZ binds directly to the HB1-element. However, we found that high levels of FTZ are required to obtain ftz-like expression. Since a FTZ-F1 binding site was replaced in 4A7ftz by two homeodomain binding sites of HB1 (Fig. 5B), the FTZ-F1 binding site seems to be required for activation at physiological FTZ concentrations, suggesting that FTZ-F1 might serve as cofactor of FTZ to select binding sites in vivo.
We have shown that the 4A7ftz enhancer is regulated by cad. In previous studies analyzing the ftz zebra element, an expression pattern similar to the one of 4A7ftz was found and it was demonstrated that this activity requires two in vitro binding sites (TTTATG) for CAD (6). Two of these CAD binding motifs are also present in the HB1-element (Fig. 1B). Therefore, it is likely that CAD also binds directly to the HB1-element.
The Function of the HB1-Element in the Intron of Hoxa-7 in Drosophila and the Mouse.
The upstream region of Hoxa-7 (p40 and p7), which contains functional elements regulating expression in the mouse embryo, was found not to confer any specific enhancer activity in Drosophila. The only activity of these constructs, similar to that of the conserved human Hoxd-4 upstream enhancer (33), is also observed in the control experiment using the Hz50pl vector without insert. The intron of Hoxa-7 on the other hand can function as enhancer in Drosophila. The observed expression pattern in Drosophila was responding to Antp, but only weakly to Ubx and not to cad as shown in mutants of these genes. In the mouse however, Cdx-1, the homolog of cad, is known to be a regulator of Hoxa-7 (ref. 62; P. Gruss, personal communication), and our results with the 4A7ftz construct suggest that the HB1-element may be a potential target site for Cdx-1. Therefore, the HB1-element may be a conserved target for various homeodomain proteins of both vertebrates and invertebrates.
Acknowledgments
We thank P. Gruss and collaborators for the Hoxa-7 genomic clones, R. Rivera-Pomar and H. Jäckle for providing the CAD protein, as well as U. Grossniklaus and K. Cadigan for the HS-ftzT1 stock. This work was supported by the Kantons Basel, grants from the Swiss National Science Foundation, and by the Human Frontier Science Program.
Footnotes
Abbreviations: β-gal, β-galactosidase; HB1-element, homeodomain binding sites.
References
- 1.McGinnis W, Krumlauf R. Cell. 1992;68:283–302. doi: 10.1016/0092-8674(92)90471-n. [DOI] [PubMed] [Google Scholar]
- 2.Müller M, Affolter M, Leupin W, Otting G, Wüthrich K, Gehring W J. EMBO J. 1988;7:4299–4304. doi: 10.1002/j.1460-2075.1988.tb03328.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Beachy P A, Krasnow M A, Gavis E R, Hogness D S. Cell. 1988;55:1069–1081. doi: 10.1016/0092-8674(88)90251-6. [DOI] [PubMed] [Google Scholar]
- 4.Desplan C, Theis J, O’Farrell P H. Cell. 1988;54:1081–1090. doi: 10.1016/0092-8674(88)90123-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hoey T, Levine M S. Nature (London) 1988;332:858–861. doi: 10.1038/332858a0. [DOI] [PubMed] [Google Scholar]
- 6.Dearolf C R, Topol J, Parker C S. Nature (London) 1989;341:341–343. doi: 10.1038/341340a0. [DOI] [PubMed] [Google Scholar]
- 7.Affolter M, Percival-Smith A, Müller M, Leupin W, Gehring W J. Proc Natl Acad Sci USA. 1990;87:4093–4097. doi: 10.1073/pnas.87.11.4093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ekker S C, Young K E, von Kessler D P, Beachy D. EMBO J. 1991;10:1179–1186. doi: 10.1002/j.1460-2075.1991.tb08058.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Regulski M, Dessain S, McGinnis N, McGinnis W. Genes Dev. 1991;5:278–286. doi: 10.1101/gad.5.2.278. [DOI] [PubMed] [Google Scholar]
- 10.Ekker S C, von Kessler D P, Beachy P A. EMBO J. 1992;11:4059–4072. doi: 10.1002/j.1460-2075.1992.tb05499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Otting G, Qian Y Q, Billeter M, Müller M, Affolter M, Gehring W J, Wüthrich K. EMBO J. 1990;9:3085–3092. doi: 10.1002/j.1460-2075.1990.tb07505.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Qian Y Q, Otting G, Billeter M, Müller M, Gehring W, Wüthrich K. J Mol Biol. 1993;234:1070–1083. doi: 10.1006/jmbi.1993.1660. [DOI] [PubMed] [Google Scholar]
- 13.Billeter M, Qian Y Q, Otting G, Müller M, Gehring W, Wüthrich K. J Mol Biol. 1993;234:1084–1097. doi: 10.1006/jmbi.1993.1661. [DOI] [PubMed] [Google Scholar]
- 14.Kissinger C R, Liu B, Martin-Blanco E, Kornberg T B, Pabo C O. Cell. 1990;63:579–590. doi: 10.1016/0092-8674(90)90453-l. [DOI] [PubMed] [Google Scholar]
- 15.Wolberger C, Vershon A K, Liu B, Johnson A D, Pabo C O. Cell. 1991;67:517–528. doi: 10.1016/0092-8674(91)90526-5. [DOI] [PubMed] [Google Scholar]
- 16.Klemm J D, Rould M A, Aurora R, Herr W, Pabo C O. Cell. 1994;77:21–32. doi: 10.1016/0092-8674(94)90231-3. [DOI] [PubMed] [Google Scholar]
- 17.Schneuwly S, Klemenz R, Gehring W J. Nature (London) 1987;325:816–818. doi: 10.1038/325816a0. [DOI] [PubMed] [Google Scholar]
- 18.Mann R S, Hogness D S. Cell. 1990;60:597–610. doi: 10.1016/0092-8674(90)90663-y. [DOI] [PubMed] [Google Scholar]
- 19.Gibson G, Schier A, LeMotte P, Gehring W J. Cell. 1990;62:1087–1103. doi: 10.1016/0092-8674(90)90386-s. [DOI] [PubMed] [Google Scholar]
- 20.Gibson G, Gehring W J. Development (Cambridge, UK) 1988;102:657–675. [Google Scholar]
- 21.Kuziora M A, McGinnis W. Mech Dev. 1991;33:83–94. doi: 10.1016/0925-4773(90)90137-b. [DOI] [PubMed] [Google Scholar]
- 22.Schier A F, Gehring W J. Nature (London) 1992;356:804–807. doi: 10.1038/356804a0. [DOI] [PubMed] [Google Scholar]
- 23.Lin L, McGinnis W. Genes Dev. 1992;6:1071–1081. doi: 10.1101/gad.6.6.1071. [DOI] [PubMed] [Google Scholar]
- 24.Chan S-K, Mann R, S. Genes Dev. 1993;7:796–811. doi: 10.1101/gad.7.5.796. [DOI] [PubMed] [Google Scholar]
- 25.Capovilla M, Brandt M, Botas J. Proc Natl Acad Sci USA. 1994;76:4464–4468. [Google Scholar]
- 26.Sun B, Hursh D A, Jackson D, Beachy P A. EMBO J. 1995;14:520–535. doi: 10.1002/j.1460-2075.1995.tb07028.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jones, McGinnis W. Genes Dev. 1993;7:229–240. doi: 10.1101/gad.7.2.229. [DOI] [PubMed] [Google Scholar]
- 28.Malicki J, Schughart K, McGinnis W. Cell. 1990;63:961–967. doi: 10.1016/0092-8674(90)90499-5. [DOI] [PubMed] [Google Scholar]
- 29.McGinnis N, Kuziora M A, McGinnis W. Cell. 1990;63:969–976. doi: 10.1016/0092-8674(90)90500-e. [DOI] [PubMed] [Google Scholar]
- 30.Zhao J J, Lazzarini R A, Pick L. Genes Dev. 1993;7:343–354. doi: 10.1101/gad.7.3.343. [DOI] [PubMed] [Google Scholar]
- 31.Bachiller D, Macias A, Duboule D, Morata G. EMBO J. 1994;13:1930–1941. doi: 10.1002/j.1460-2075.1994.tb06462.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Halder G, Callaerts P, Gehring W J. Science. 1995;267:1788–1792. doi: 10.1126/science.7892602. [DOI] [PubMed] [Google Scholar]
- 33.Malicki J, Cianetti L C, Peschle C, McGinnis W. Nature (London) 1992;358:345–347. doi: 10.1038/358345a0. [DOI] [PubMed] [Google Scholar]
- 34.Awgulewitsch A, Jacobs D. Nature (London) 1992;358:341–344. doi: 10.1038/358341a0. [DOI] [PubMed] [Google Scholar]
- 35.Müller J, Thüringer F, Biggin M, Züst B, Bienz M. EMBO J. 1989;8:4143–4151. doi: 10.1002/j.1460-2075.1989.tb08599.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bergson C, McGinnis W. EMBO J. 1990;9:4287–4297. doi: 10.1002/j.1460-2075.1990.tb07877.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Carroll S B, Laymon R A, McCuthcheon M A, Riley P D, Scott M P. Cell. 1986;47:113–122. doi: 10.1016/0092-8674(86)90372-7. [DOI] [PubMed] [Google Scholar]
- 38.Hafen E, Levine M, Gehring W J. Nature (London) 1984;307:287–289. doi: 10.1038/307287a0. [DOI] [PubMed] [Google Scholar]
- 39.Bienz M, Tremml G. Nature (London) 1988;333:576–578. doi: 10.1038/333576a0. [DOI] [PubMed] [Google Scholar]
- 40.Bienz M, Saari G, Tremml G, Müller J, Züst B, Lawrence P A. Cell. 1988;53:567–576. doi: 10.1016/0092-8674(88)90573-9. [DOI] [PubMed] [Google Scholar]
- 41.Colberg-Poley A M, Voss S D, Chowdhury K, Gruss P. Nature (London) 1985;314:713–718. doi: 10.1038/314713a0. [DOI] [PubMed] [Google Scholar]
- 42.Kessel M, Schulze F, Fibi M, Gruss P. Proc Natl Acad Sci USA. 1987;84:5306–5310. doi: 10.1073/pnas.84.15.5306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Püschel A W, Balling R, Gruss P. Development (Cambridge, UK) 1990;108:435–442. doi: 10.1242/dev.108.3.435. [DOI] [PubMed] [Google Scholar]
- 44.Püschel A W, Balling R, Gruss P. Development (Cambridge, UK) 1991;112:279–287. doi: 10.1242/dev.112.1.279. [DOI] [PubMed] [Google Scholar]
- 45.Knittel T, Kessel M, Kim M H, Gruss P. Development (Cambridge, UK) 1995;121:1077–1088. doi: 10.1242/dev.121.4.1077. [DOI] [PubMed] [Google Scholar]
- 46.Mlodzik M, Gehring W J. Cell. 1987;48:465–478. doi: 10.1016/0092-8674(87)90197-8. [DOI] [PubMed] [Google Scholar]
- 47.Haerry T. Ph.D. thesis. Basel, Switzerland: Univ. of Basel; 1994. [Google Scholar]
- 48.Hiromi Y, Gehring W J. Cell. 1987;50:963–974. doi: 10.1016/0092-8674(87)90523-x. [DOI] [PubMed] [Google Scholar]
- 49.Freeman M, Nüsslein-Volhard C, Glover D M. Cell. 1986;46:457–468. doi: 10.1016/0092-8674(86)90666-5. [DOI] [PubMed] [Google Scholar]
- 50.Frasch M, Hoey T, Rushlow C, Doyle H, Levine M. EMBO J. 1987;6:749–759. doi: 10.1002/j.1460-2075.1987.tb04817.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mlodzik M, Gibson G, Gehring W J. Development (Cambridge, UK) 1990;109:271–277. doi: 10.1242/dev.109.2.271. [DOI] [PubMed] [Google Scholar]
- 52.Qian Y Q, Resendez-Perez D, Gehring W J, Wüthrich K. Proc Natl Acad Sci USA. 1994;91:4091–4095. doi: 10.1073/pnas.91.9.4091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rivera-Pomar R, Lu X, Perrimon N, Taubert H, Jäckle H. Nature (London) 1995;376:253–256. doi: 10.1038/376253a0. [DOI] [PubMed] [Google Scholar]
- 54.Gaillot B, Dollé P, Vigneron M, Featherstone M S, Baron A, Duboule D. Development (Cambridge, UK) 1989;107:343–359. doi: 10.1242/dev.107.2.343. [DOI] [PubMed] [Google Scholar]
- 55.Morrison A, Chaudhuri Ch, McNaughton L, Kuroiwa A, Krumlauf R. Mech Dev. 1995;53:47–59. doi: 10.1016/0925-4773(95)00423-8. [DOI] [PubMed] [Google Scholar]
- 56.Cianetti L, Di Cristofaro A, Zappavigna V, Bottero L, Boccoli G, Testa U, Russo G, Boncinelli E, Peschle C. Nucleic Acids Res. 1990;18:4361–4368. doi: 10.1093/nar/18.15.4361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wilde D, Akam M. EMBO J. 1987;6:1393–1401. doi: 10.1002/j.1460-2075.1987.tb02380.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Schier A F, Gehring W J. EMBO J. 1993;12:1111–1119. doi: 10.1002/j.1460-2075.1993.tb05752.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pick L, Schier A, Affolter M, Schmidt-Glenewinkel T, Gehring W J. Genes Dev. 1990;4:1224–1239. doi: 10.1101/gad.4.7.1224. [DOI] [PubMed] [Google Scholar]
- 60.Han W, Yu Y, Altan N, Pick L. Mol Cell Biol. 1993;13:5549–5559. doi: 10.1128/mcb.13.9.5549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Epstein J, Cai J, Glaser T, Jepeal L, Maas R. J Biol Chem. 1994b;269:8355–8361. [PubMed] [Google Scholar]
- 62.Subraminian V, Meyer B I, Gruss P. Cell. 1995;83:641–653. doi: 10.1016/0092-8674(95)90104-3. [DOI] [PubMed] [Google Scholar]