

Abstract
Separate event-related brain potential (ERP) components have been hypothesized to index familiarity and recollection processes that support recognition memory. A 300–500 ms mid-frontal FN400 old/new difference has been related to familiarity, whereas a 500–800 ms parietal old/new difference has been related to recollection. Other recent work has cast doubt on the FN400 familiarity hypothesis, especially its application to familiarity-based recognition of conceptually impoverished stimuli such as novel faces. Here we show that FN400 old/new differences can be observed with novel faces, and as predicted by the familiarity hypothesis, these differences are observed regardless of whether or not recognition is accompanied by the recollection of specific details from the study episode. Furthermore, FN400 differentiation between hits and misses is more consistent with an explicit familiarity process than an implicit memory process.
Dual-process theorists propose that recollection and familiarity are dissociable processes contributing to recognition memory (e.g., Hintzman & Curran, 1994; Jacoby, 1991; Mandler, 1980; Norman & O’Reilly, 2003; Rugg & Yonelinas, 2003; Yonelinas, 2002). Recollection is thought to involve processes by which specific details can be accurately recalled when something is recognized. Familiarity is thought to underlie a general sense that a stimulus has been previously encountered without the ability to remember specific details.
A wealth of evidence has supported the dual-process perspective (for reviews, see, Rugg & Yonelinas, 2003; Yonelinas, 2002), including research measuring event-related brain potentials (for reviews see, Curran, Tepe, & Piatt, 2006b; Friedman & Johnson, 2000; Mecklinger, 2000; Wilding & Sharpe, 2003). Familiarity has been associated with negative-going electrical activity recorded over frontal scalp locations between 300–500 ms, called the FN400 or mid-frontal old/new effect. Recollection has been associated with positive-going activity recorded over posterior parietal locations between 500–800 ms, often called the parietal old/new effect. For example, Curran (Curran, 2000; Curran & Cleary, 2003) performed a series of experiments in which subjects studied words or pictures. ERPs were recorded during recognition tests including studied items, non-studied lures that were similar to studied items (e.g., plurality-reversed words or orientation-reversed pictures), and new items. The FN400 discriminated between familiar (studied and similar lures) versus unfamiliar (new) items, whereas the parietal effect discriminated between recollected (hits to studied items) and non-recollected items (false alarms to similar lures or correctly rejected new items). Similar results have been obtained with semantically similar lures (Nessler, Mecklinger, & Penney, 2001, 2005).
A recent experiment by Yovel and Paller (2004) has cast some doubt on the proposed relationship between the FN400 and familiarity. The experiment used face stimuli to explore the “butcher-on-the-bus phenomenon,” or the intuitive feeling experienced when a familiar person is encountered in an unfamiliar situation (such as seeing one’s butcher on a bus). Twelve participants studied previously unfamiliar faces paired with occupations that were presented verbally by a male voice. Faces were monochromatic photographs with minimal background information, depicting young Caucasian adults without distinctive facial features such as facial hair or glasses (from Endl, Walla, Lindinger, Lalouschek, Barth, Deecke, & Lang, 1998). During ten 24-item study lists, participants judged whether each face and occupation fit together or not. Each test list included 24 old/studied faces and 12 new/unstudied faces without occupations. First, participants gave standard old/new recognition judgments. If the participant responded “new,” the trial ended. If they responded “old,” they were presented with three subsequent choices: “occupation”, “other specifics”, and “no specifics”. If the participant chose the “occupation” response, she or he said the occupation aloud. If the participant responded “other specifics”, this meant that they recollected information from the study phase other than occupation. Participants responded “no specifics” if they knew the face was seen in the study phase, without recollection of occupation or other details. The latter responses were interpreted as indicative of familiarity-based responses.
Yovel and Paller’s results were inconsistent with the hypothesis that FN400 old/new differences are related to familiarity: Rather than differentiating correctly classified old and new faces regardless of whether or not occupations or other details were recollected, the 300–500 ms FN400 old/new differences were only observed when occupations were recollected. Later, 500–700 ms amplitudes corresponding to the “parietal old/new effect” showed a graded pattern such that amplitudes when participants did not recollect any details fell in between amplitudes to new trials and trials in which occupations were recollected. Rather than indexing familiarity, the authors proposed that 300–500 ms old/new differences may be related to conceptual priming, as has been supported by subsequent research showing a positive correlation between FN400 effects and reaction time differences between old and new items on a conceptual priming task (Voss & Paller, 2006).
Given the theoretical importance of Yovel and Paller’s (2004) experiment, it is critical to determine the replicability and generalizability of the results. The present experiment followed Yovel and Paller’s design in most respects except for two key differences. First, we sought to maximize familiarity-based discrimination of old and new items. Most familiarity-based models of recognition memory suggest that familiarity reflects an assessment of the global similarity between a test item and all previously studied information (Dennis & Humphreys, 2001; Gillund & Shiffrin, 1984; Hintzman, 1988; Norman & O’Reilly, 2003; Shiffrin & Steyvers, 1997). Thus, familiarity does not provide a sufficiently accurate basis for discriminating studied items from similar lures (e.g., Hintzman & Curran, 1994; Hintzman & Curran, 1995; Hintzman, Curran, & Oppy, 1992). Indeed, previous research has shown that the FN400 differentiates old items from dissimilar lures, but does not differentiate old items from similar lures (Curran, 2000; Curran & Cleary, 2003; Nessler & Mecklinger, 2003; Nessler et al., 2001). Thus, rather than using relatively homogeneous faces with a high degree of inter-item similarity like those of Yovel and Paller (e.g., monochromatic, young, Caucasian, no facial hair or glasses), we chose a more heterogeneous set including color photographs of people who varied in ethnicity (or race), age, facial hair, and glasses. We surmised that a more heterogeneous face set would reduce the similarity between old and new items, and thereby increase familiarity-based discrimination. Second, we noticed that Yovel and Paller’s subjects showed a markedly conservative response bias such that correct rejection rates (88%) were much higher than hit rates (65%)— possibly caused by requiring follow-up responses to “old” judgments, but not to “new” judgments. Participants may have been biased to choose the easier and quicker of the two alternatives. To obtain more hits for ERP analyses, we sought to reduce this response bias by requiring follow-up responses after both “old” and “new” judgments. In particular, subjects were asked to give confidence ratings and to guess the person’s occupation on a randomly selected third of faces judged “new”.
If the FN400 is not related to familiarity, then Yovel and Paller’s results should be replicated under the present conditions that maximize the ability for familiarity to contribute to face recognition. Specifically, the 300–500 ms FN400 old/new difference should be observed only when comparing trials with occupation recollection to new trials. If the FN400 is related to familiarity, then FN400 old/new differences should be similar for words recognized with or without occupation recollection.
Method
Participants
Thirty-three right-handed students at the University of Colorado participated in the experiment for payment of $15 per hour. All participants gave informed consent. Data from nine participants were discarded because of excessive eye-movement artifacts (n = 2), low trial counts (less than 20 trials/condition, n = 4), low accuracy (n = 1), or an excessive number of bad channels (n = 2). Of the remaining 24 subjects included in the analyses (mean age = 21 years; range = 18–27 years), 14 were female.
Stimuli
Stimuli were 360 faces and 240 occupations. Face stimuli were taken from the color FERET database of photographs (Phillips, Moon, Rizvi, & Rauss, 2000). Face stimuli consisted of 126 Caucasian women, 54 non-Caucasian women, 111 Caucasian men, and 69 non-Caucasian men. The racial and ethnic groups of non-Caucasian faces included African American, Asian, Southeast Asian, Middle Eastern, Pacific Islander, Hispanic, and Native American. The photographs depicted 24 individuals wearing glasses, 29 men with moustaches, and 25 men with beards. Face stimuli were shown in color, including shoulders with clothing and some jewelry on the women. The occupations were mostly the same as those used by Yovel and Paller, but we replaced some unusual occupations (e.g., drug dealer) with ones that were more typical, as well as replacing some that were too confusable (e.g., civil engineer and electrical engineer; see Supplementary Materials). The occupations were presented directly below the faces in white on a black background on an LCD computer monitor. All face stimuli subtended a visual angle of 2.46° horizontally, 3.84° vertically.
Design
The experiment consisted of 10 blocks and lasted approximately 136 minutes. In each block, participants studied 24 unique face-occupation pairings. Approximately 0.5 minutes later, the participants were tested with those 24 faces plus 12 new faces. For each subject separately, faces were randomly assigned to old (studied) or new (non-studied) conditions and old faces were randomly paired with occupations. Responses were registered using a four key response pad. EEG was collected only during each test phase.
Procedure
Participants completed a short practice to familiarize them with the various procedures of the experiment. A Geodesic Sensor Net was then applied.
Each study trial began with a 1000 ms gray fixation cross at eye level on a black background followed by a 2000 ms face-occupation presentation. Occupation names appeared visually below targets— a minor departure from Yovel and Paller’s auditory presentation of occupations. To enhance encoding, participants were asked to judge the fit of each face/occupation pair. Participants pressed one of two keys on each trial according to whether they believed the face and occupation fit together or not. Participants were also told to try to remember the face-occupation pairings because they would appear on a subsequent memory test.
During the test phase, the faces appeared without occupations and participants were asked to judge whether they believed the face had appeared in the study phase or not (i.e., “old” or “new”). Each test trial consisted of a pre-stimulus fixation varying between 1250–1750 ms, a face that appeared for 1000 ms, followed by another fixation until the subject made a response. We chose a 1000 ms presentation, rather than 500 ms (Yovel and Paller, 2004), to remove possible offset potentials from the recording epoch. Subjects were instructed to withhold their response until the face disappeared, to lessen response-related ERP effects. If an “old” response was detected, the fixation cross was replaced after a 600 ms delay and the participant was asked, “Do you: 1) Remember the person’s occupation, (occupation), 2) Remember specific details about the person (other specific details), or 3) Not remember any specific details (no specific details)?” If an “occupation” response was detected, this signified that the associated occupation could be recalled, and it was then spoken aloud. If an “other specifics” response was detected, this signified that details of the study-phase episode (other than occupation) could be recalled, but no verbal response was given. If a “no specifics” response was detected, this signified that no details from the study-phase episode could be remembered, and no verbal response was given. If a “new” response was detected, the fixation cross was replaced after a 600 ms delay with the following choices for the next response: “very confident” (high) or “not confident” (low). After this response, the participants were then asked to generate an occupation for the face for randomly selected trials following 33% of “new” responses. This additional requirement made the “new” condition more equitable with the “old” condition in regards to difficulty and time commitment. The experimenter then initiated the next trial after EEG artifacts had subsided. Throughout each test block participants were instructed to refrain from blinking insofar as they were able.
To motivate participants to perform adequately on the task, they received one point for correctly identifying each face as either “old” or “new.” They were told that the highest scoring participant would receive a $25 bonus once all participants had completed the experiment.
EEG/ERP Methods
Scalp voltages were collected during each test list with a 128 channel Geodesic Sensor Net™ connected to an AC-coupled, 128 channel, high-input impedance amplifier (200 MΩ, Net Amps™, Electrical Geodesics Inc., Eugene, OR). Amplified analog voltages (0.1–100 Hz bandpass, -3 dB) were digitized at 250 Hz. Individual sensors were adjusted until impedances were less than 50 kΩ. ERPs were baseline corrected to a 100-ms pre-stimulus recording interval and digitally low-pass filtered at 40 Hz. Trials were discarded from analysis if they contained eye movements (EOG over 70μV), or more than 20% of channels were bad (average amplitude over 100 μV or transit amplitude over 50 μV). EEG was measured with respect to a vertex reference (Cz). An average-reference transformation was used for the primary analyses to more accurately estimate the scalp topography of the measured electrical fields and to minimize the effects of reference-site activity (Dien, 1998). Subsidiary analyses used an average-mastoid reference to facilitate comparison with Yovel and Paller (2004).
Results
Yovel and Paller’s (2004) primary analyses focused on correct rejections, familiar hits (“no specific details”), and hits associated with recollection of the correct occupation. They reported that ERPs were similar for correct rejection and misses, but did not formally report misses. We include misses here because they are potentially relevant for differentiating processes related to familiarity vs. priming, which is particularly relevant to the FN400 interpretation.i Other conditions were considered in the Supplementary Materials. For Analysis of Variance (ANOVA) involving both reaction time (RT) and ERPs, all p values associated with more than 1 degree of freedom have been corrected according to the conservative Geisser-Greenhouse procedure for sphericity violations (Winer, 1971).
Behavioral Results
Accuracy
Responses reflecting accurate recognition memory (“hit rates”) averaged 81% for old faces (range 67–94%). Correct rejections of new stimuli averaged 90% (68–99%). When participants correctly identified faces as old, they recalled occupations 30% (15–60%) of the time, identified “other specific details” 34% (0–68%) of the time, and reported “no details” 36% (9–79%) of the time. The accuracy of verbally reported occupations was 96% (84–100%).
Reaction Time
Reaction times (RT) were slowed artificially by the requirement to withhold responses until the test face disappeared (1 sec after face onset), so RTs are unlikely to accurately reflect decision times. However, we observed RT differences among the four primary conditions used for ERP analyses: occupation hits (MN = 1582 ms), familiar hits (1658 ms), misses (1615 ms), and correct rejections (1429 ms), F(3, 23) = 5.65, MSE = 42315, p < .01. Pairwise comparisons indicated that correct rejections were faster than all other responses involving old faces (all p < .02), but none of the responses to old faces differed among themselves.
ERP Results
Initial analysis focused on spatial regions of interest (ROIs) based upon our previous research (Curran, 2000, 2004; Curran & Cleary, 2003; Curran, DeBuse, & Leynes, in press; Curran, DeBuse, Woroch, & Hirshman, 2006a; Curran & Dien, 2003; Curran & Friedman, 2004; Curran, Tanaka, & Weiskopf, 2002). The FN400 was analyzed over two anterior, superior channels groups located near the standard F3 and F4 sites. These are labeled left and right anterior/superior regions (LAS and RAS, respectively, see Figure 1). The parietal old/new effect was analyzed over two posterior, superior channels groups that included the standard P3/P4 locations in addition to more anterior channels. These are labeled left and right posterior/superior regions (LPS and RPS, respectively, see Figure 1). ERP waveforms created by averaging the ERPs within each region and across subjects are shown in Figure 2. The number of correct, artifact-free trials entered into each ERP/subject averaged 99 for correct rejections, 63 for familiar hits, 44 for occupation hits, and 42 for misses. Following Yovel and Paller (2004), the FN400 was analyzed from 300–500 ms whereas the parietal effects were analyzed from 500–700 ms.
Figure 1.
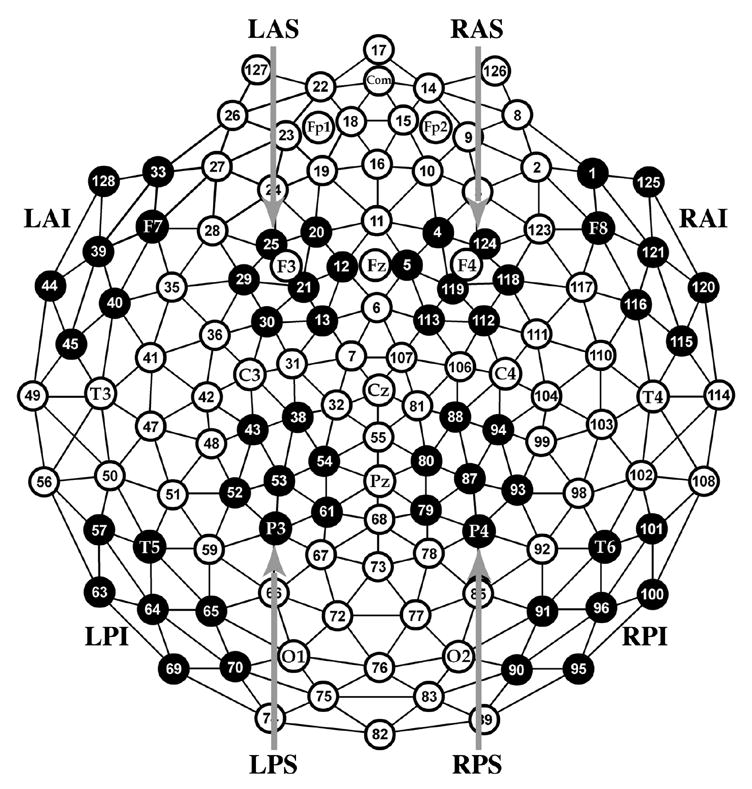
Geodesic Sensor Net Layout. Electrode sites are number along with selected 10-10 positions. Black clusters are regions included in the topographic analyses. L = left, R = right, A = anterior, P = posterior, I = inferior, S = superior.
Figure 2.
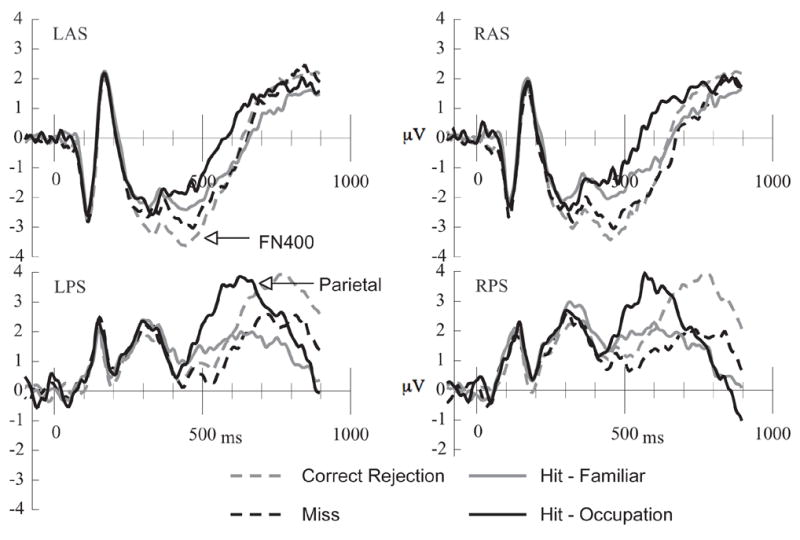
Average-referenced, grand averaged ERPs in each of the 4 regions depicted in Figure 1. L = left, R = right, A = anterior, P = posterior, I = inferior, S = superior.
FN400 Results (300–500 ms)
If the FN400 is related to familiarity, then it should be more negative for correct rejections than hits, as is normally observed, and it should not differ between occupation hits and familiar hits. The FN400 to misses should represent an intermediate level of familiarity, falling in between hits and correct rejections. Differences between hits and misses should be larger than differences between false alarms and misses because the latter two familiarity distributions should have more overlap (both falling to the left of the new/old criterion) than hits and misses (falling on opposite sides of the new/old criterion). If the FN400 is related to implicit priming, on the other hand, it might be expected to differentiate old and new items regardless of explicit recognition accuracy, so hits and misses would be similar and both would differ from correct rejections (e.g., Rugg, Mark, Walla, Schloerscheidt, Birch, & Allan, 1998).
Mean LAS and RAS amplitude between 300 and 500 ms was the dependent measure. A condition (occupation hit, familiar hit, correct rejection, miss) × hemisphere ANOVA indicated differences between conditions, F(3,69) = 7.06, MSE = 2.66, p < .001 (see top of Figure 2 and Table 1A). Pairwise comparisons confirmed that the difference in amplitudes between familiar hits and correct rejections was significant, F(1,23) = 13.61, p < .001, as was the difference in amplitudes between the occupation hits and correction rejections, F(1,23)=19.01, p<.001. Amplitudes for hits associated with familiarity versus occupation recollection did not differ significantly, F(1,23) = .45. Misses were more negative than occupation hits (F(1,23) = 6.53, p < .05), marginally more negative than familiar hits (F(1,23) = 3.69, p = .06), and not different from correct rejections (F(1,23) = 2.47, p = .12).
Table 1.
Amplitude means and (standard errors).
| Latency (ms) | Correct Rejection | Miss | Familiarity Hit | Occupation Hit |
|---|---|---|---|---|
| 300–500 | −3.2 (0.4) | −2.6 (0.4) | −2.0 (0.5) | −1.8 (0.4) |
| 500–700 | 2.2 (0.5) | 1.3 (0.5) | 1.9 (0.5) | 3.3 (0.5) |
Note. 300–500 ms amplitudes were calculated from the average of left and right anterior, superior regions (LAS & RAS). 500–700 ms amplitudes were calculated from the average of left and right posterior, superior regions (LPS & RPS).
Because the pattern of FN400 results was qualitatively different from those reported by Yovel and Paller (2004), we wanted to verify that disparities could not be attributed to analytic approach. Therefore, following Yovel and Paller (2004), we re-referenced the ERPs to the average of the two mastoid electrodes (57 & 101) and focused on a standard series of mid-line channels (see Figure 3). Mean mastoid-referenced amplitudes between 300 and 500 ms were analyzed in a condition (occupation-hit, familiar-hit, correct rejection, miss) × channel (Fpz [16], Fz [6], Cz [62], Pz [62], Oz [76]) ANOVA. All effects were significant: condition (F[3, 69] = 5.20, MSE = 6.18, p < .01), channel (F[4, 92] = 12.44, MSE = 41.81, p < .001), and condition x channel (F[12, 276] = 4.21, MSE = 1.43, p < .001). The condition x location interaction captured the fact that condition effects were larger over frontal than posterior electrodes. Pairwise comparisons focusing on the Fz location, where differences were largest, indicated that amplitudes to correct rejections (MN = −3.84 μV) were more negative than both familiar hits (MN = −2.35 μV; F = 18.72, p < .01) and occupation hits (MN = −1.92 μV; F = 30.99, p < .001), but the two categories of hits did not significantly differ (F = 1.54). Misses (MN = −3.40 μV) were more negative than occupation hits (F(1,23) = 18.46, p < .001), more negative than familiar hits (F(1,23) = 9.34, p < .05), and not different from correct rejections (F(1,23) = 1.61).
Figure 3.
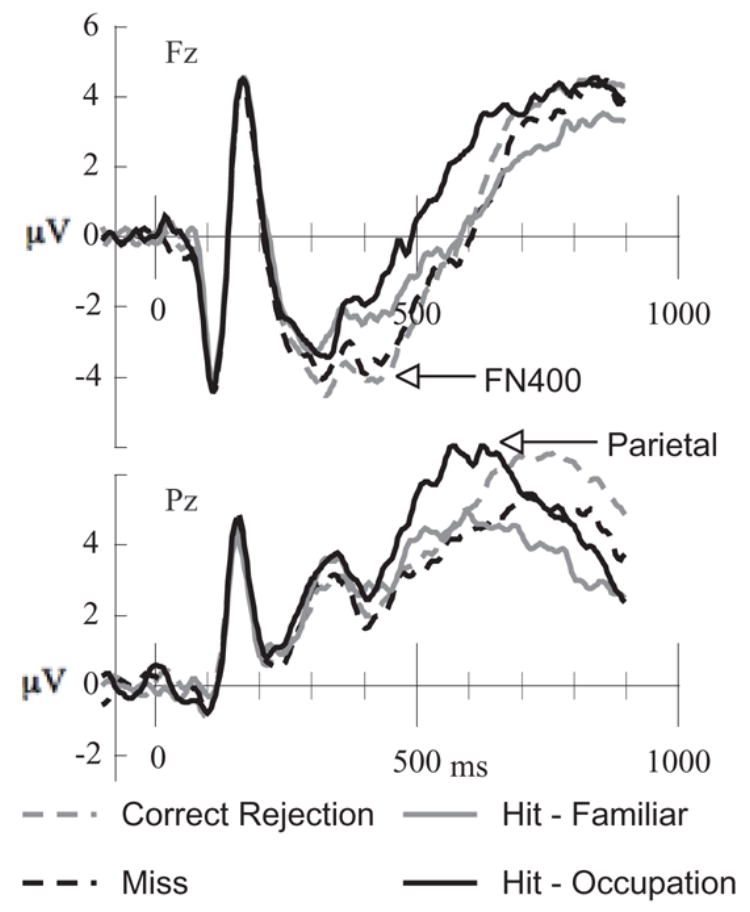
Mastoid-referenced, grand averaged ERPs from frontal (Fz) and parietal (Cz) sensor locations.
Overall, the pattern of FN400 differences is consistent with the hypothesis that this component is related to familiarity: occupation hits = familiar hits > misses = correct rejections. The analysis of mastoid-reference mid-line channels was entirely consistent with our initial average-referenced analysis of the LAS and RAS ROIs, except that the difference between familiar hits and misses was significant with the mastoid reference (p < .05), but only marginally significant for the average reference (p = .06).
Parietal Results (500–700 ms)
Mean LPS and RPS amplitude between 500 and 700 ms was the dependent measure. A condition (occupation hit, familiarity hit, correct rejection, miss) × hemisphere ANOVA indicated that the 500–700 ms effect varied significantly by condition, F(3,69) = 9.48, MSE = 3.62, p < .001 (see bottom of Figure 2 and Table 1A).ii Pairwise comparisons revealed that the difference between occupation hits and correct rejections was significant, F(1,23) = 10.72, p < .01, as was the difference between the occupation hits and familiar hits, F(1,23) = 13.10, p < .002. The difference between the correct rejections and the familiar hits was not significant F(1,23) < 1. Amplitudes for misses were lower than for occupation hits (F(1, 23) = 26.88, p < .001) and correct rejections (F(1, 23) = 4.99, p < .05), but were not different from familiar hits (F(1, 23) = 2.28).
Range Normalized Difference Scores
To consider the broader topography of the results, we computed mean amplitude within each of the 8 electrode clusters depicted in Figure 1 (following, Curran, 2000, 2004; Curran & Cleary, 2003; Curran et al., 2006a; Curran et al., 2002). Separate difference scores were intended to be associated with recognition with recollection (occupation hits minus correct rejections, OH-CR; Figure 4B) and without recollection (familiar hits minus correct rejections, FH-CR; Figure 4A). The difference scores were scaled using a range normalization method (McCarthy & Wood, 1985). iii Qualitatively different scalp topographies might reflect either (a) different neuronal sources or (b) the same source with different relative strengths (see also, Alain, Achim, & Woods, 1999; Picton, Bentin, Berg, Donchin, Hillyard, Johnson, Miller, Ritter, Ruchkin, Rugg, & Taylor, 2000). Thus, condition-by-location interactions are consistent with separate sources, but do not necessarily demand separate sources. Furthermore, we chose this approach to maintain consistency with Yovel and Paller (2004) who also considered scalp topography with rescaled amplitudes.
Figure 4.
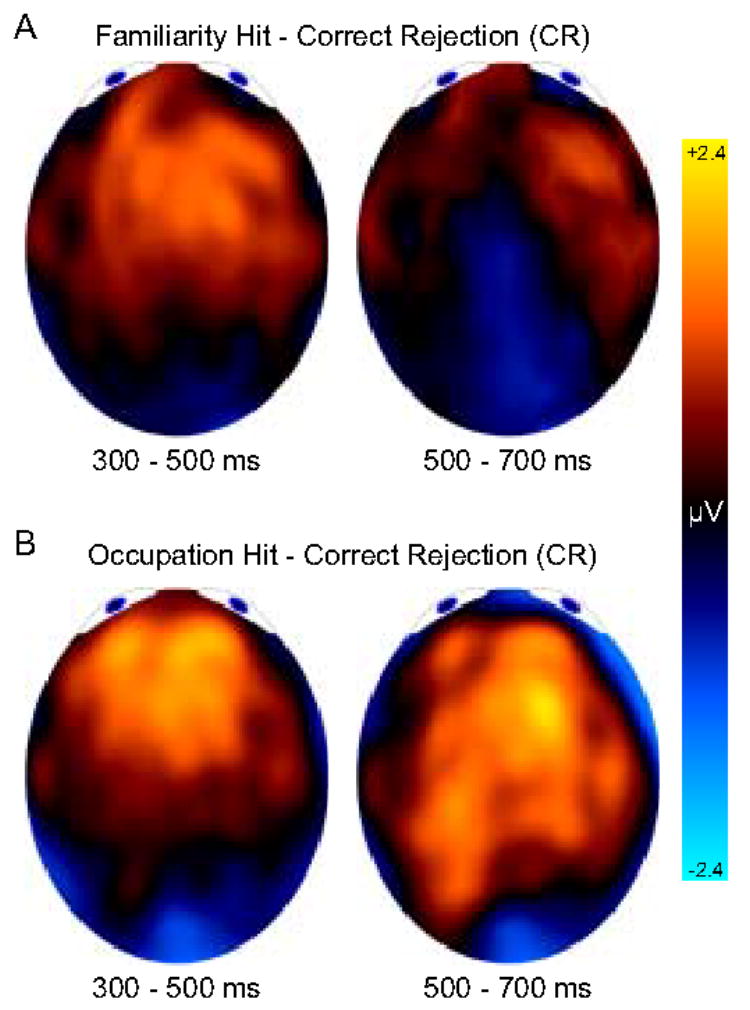
Topographic maps comparing familiarity and recollection. (A) Familiarity-related differences at the time of the FN400 (300 – 500 ms) and parietal effects (500 – 700 ms). (B) Recollection-related differences at the time of the FN400 (300 – 500 ms) and parietal effects (500 – 700 ms).
The range-normalized difference scores were analyzed in a time (300–500, 500–700) × condition (FH-CR, OH-CR) × hemisphere × anterior/posterior (AP) × inferior/superior (IS) ANOVA. As is the purpose of rescaling, the condition main effect was removed, F < 1. Although many time and/or location effects were significant, we will only report interactions with condition that are of primary interest. The time x condition x IS interaction, F(1,23) = 6.39, MSE = 1.03, p = .02, captured the observation that all old/new effects were characterized by positive superior differences together with negative inferior differences, except for the 500–700 FH-CR difference which was relatively flat across superior and inferior regions. This merely reflects the absence of significant old/new effects associated with familiar hits in the later time period (Figure 4A, right). The time x condition x AP interaction, F(1,23) = 8.15, MSE = .68, p = .009, indicated that posterior parietal old/new differences were largest from 500 to 700 ms when occupation was recollected (Figure 4B, right).
Discussion
As predicted by the hypothesis that 300–500 ms mid-frontal FN400 old/new effects are related to familiarity, we found that the FN400 did not differ according to subject’s ability to recollect occupations that had been previously associated with correctly recognized faces. Critically, FN400 old/new differences were significant even when responses were presumably familiarity-based (i.e., “no details”), despite a previous report that familiar faces are not differentiated from new faces until a later 500–700 ms time frame associated with the parietal old/new effect (Yovel & Paller, 2004). Overall, the pattern of mean amplitudes observed for the FN400 was consistent with a familiarity-based process such that amplitudes corresponding to hits, with or without recollection, were more positive than both correct rejections and misses. Conversely, the 500 – 700 ms parietal old/new differences were only observed when occupations were correctly remembered, supporting the association between recollection and the parietal effect (see Supplementary Materials for analyses contrasting recollection and confidence).
Why do the present results differ from those of Yovel and Paller (2004)? We predicted that using a more heterogeneous set of stimuli should have enhanced familiarity-based discrimination between old and new faces, as would be expected if familiarity reflects an assessment of the global similarity between a test item and all previously studied information (Dennis & Humphreys, 2001; Gillund & Shiffrin, 1984; Hintzman, 1988; Norman & O’Reilly, 2003; Shiffrin & Steyvers, 1997). Other research has previously shown that the FN400 discriminates old from dissimilar lures, but not from similar lures (Curran, 2000; Curran & Cleary, 2003; Nessler & Mecklinger, 2003; Nessler et al., 2001). In essence, highly similar materials potentially cause more interference than dissimilar materials, leading to an inability to effectively use familiarity to discriminate between old and new items. Other recent research has shown that the FN400 old/new effect observed with faces is susceptible to interference caused by manipulations of list length (Norman, Tepe, Nyhus, & Curran, submitted).
As detailed in Table 2, overall face recognition accuracy was higher in the present experiment (d′ = 2.16 ) than Yovel and Paller’s experiment (d′ = 1.56). To compare discrimination accuracy within the various response categories while minimizing the influence of response bias, d′ was computed within each category. To compute d′ (Table 2D), results originally reported as the proportion of hits within each response category (Table 2B) were transformed to the proportion of overall old trials in each response category (Table 2C) to compare with the overall false alarm rates. In each and every category d′ was higher in the present experiment, including familiarity-based responses. As previously argued, we believe familiarity-based discrimination was enhanced by the use of a more heterogeneous face set.
Table 2.
Accuracy in Comparison with Yovel and Paller (2004).
| Curran & Hancock | Yovel & Paller | |
|---|---|---|
| A. Overall Recognition Performance | ||
| Hits | 0.81 | 0.65 |
| False Alarms | 0.10 | 0.12 |
| d′ | 2.16 | 1.56 |
| B. Proportion of Hits in Each Response Category | ||
| Occupation | 0.30 | 0.29 |
| Other Details | 0.34 | 0.24 |
| No Details | 0.36 | 0.47 |
| C. Proportion of Old Trials in Each Response Category | ||
| Occupation | 0.24 | 0.19 |
| Other Details | 0.28 | 0.16 |
| No Details | 0.29 | 0.30 |
| D. d′ in Each Response Category | ||
| Occupation | 0.58 | 0.29 |
| Other Details | 0.68 | 0.16 |
| No Details | 0.73 | 0.65 |
Other minor differences between the present experiment and that of Yovel and Paller (2004) are less likely to have influenced the FN400 results. First, an increase in test stimulus duration from 500 to 1000 ms cannot influence 300–500 ms ERPs. Second, changes made to the occupations (visual rather than auditory study, less confusable, more typical) may influence occupation recollection, but should have no bearing on the familiarity of the faces themselves.
The present results add to a growing body of evidence questioning the sufficiency of the hypothesis that FN400 old/new effects are related to conceptual priming. Like the present results, other research has shown FN400 old/new differences using previously unfamiliar faces that are unlikely to have conceptual representations (Johansson, Mecklinger, & Treese, 2004; Nessler et al., 2005; Norman et al., submitted). Other stimuli possibly lacking conceptual representation for which FN400 old/new differences have been observed include novel dimensional shapes (Curran et al., 2002; Groh-Bordin, Zimmer, & Ecker, 2006). Furthermore, if conceptual priming is thought to be implicit (e.g. Voss & Paller, 2006; Yovel & Paller, 2004), one might expect to observe FN400 old/new differences regardless of recognition memory accuracy (e.g., hits = misses > correct rejection, Rugg et al., 1998), but no such implicit influences were observed in the present experiment. Although conceptual representations alone seem insufficient to explain all reported FN400 old/new differences, this does not necessarily rule out a relationship between the FN400 and conceptual priming in some circumstances. Conceptual features are likely to contribute to familiarity under some circumstances, such as when perceptual features vary between study and test (Curran & Dien, 2003), and these same features may contribute to conceptual priming.
Conclusions
Faces associated with correct rejection or misses showed a significantly more negative FN400 (300 – 500 ms) than faces that were correctly recognized as old, regardless of whether or not subjects could also recollect the associated occupation from the study episode. This result is consistent with the hypothesis that the FN400 is related to familiarity, and that the underlying mechanism applies to novel faces as well as to other previously examined stimuli such as words, identifiable pictures, and novel shapes. Later, 500–700 ms, parietal old/new effects were only observed when subjects could recall the associated occupation, as would be expected from the hypothesis that parietal old/new effects are associated with recollection. These results support the dual-process perspective that separate familiarity and recollection processes contribute to recognition memory, and these processes are indexed by separate ERP components.
Supplementary Material
Acknowledgments
This research was funded by NIMH Grant R01-MH64812 and a University of Colorado Summer Undergraduate Research Fellowship. The experiment was part of Jane Hancock’s Senior Honors Thesis, and we would like to thank the other thesis committee members: Mark Whisman and Claudia Van Gerven. We also thank Brion Woroch and Casey DeBuse for programming and research assistance, and Nicole Speer and Erika Nyhus for helpful comments. Portions of the research in this paper use the Color FERET database of facial images collected under the FERET program(Phillips, Moon, Rizvi, & Rauss, 2000).
Footnotes
False alarm rates were not high enough to obtain sufficient observations for ERP analyses.
As for the FN400 results, parietal results were qualitatively similar with analysis of mastoid-referenced mid-line channels.
Recent debates about the use of rescaled amplitudes concern the vector length method rather than the range normalization method used here (Dien & Santuzzi, 2005; Urbach & Kutas, 2002, 2006; Wilding, 2006).
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Alain C, Achim A, Woods DL. Separate memory-related processing for auditory frequency and patterns. Psychophysiology. 1999;36:737–744. [PubMed] [Google Scholar]
- Curran T. Brain potentials of recollection and familiarity. Memory & Cognition. 2000;28:923–938. doi: 10.3758/bf03209340. [DOI] [PubMed] [Google Scholar]
- Curran T. Effects of attention and confidence on the hypothesized ERP correlates of recollection and familiarity. Neuropsychologia. 2004;42:1088–1106. doi: 10.1016/j.neuropsychologia.2003.12.011. [DOI] [PubMed] [Google Scholar]
- Curran T, Cleary AM. Using ERPs to dissociate recollection from familiarity in picture recognition. Cognitive Brain Research. 2003;15:191–205. doi: 10.1016/s0926-6410(02)00192-1. [DOI] [PubMed] [Google Scholar]
- Curran T, DeBuse C, Leynes PA. Conflict and criterion setting in recognition memory. Journal of Experimental Psychology: Learning, Memory, and Cognition. doi: 10.1037/0278-7393.33.1.2. in press. [DOI] [PubMed] [Google Scholar]
- Curran T, DeBuse C, Woroch B, Hirshman E. Combined pharmacological and electrophysiological dissociation of familiarity and recollection. Journal of Neuroscience. 2006a;26:1979–1985. doi: 10.1523/JNEUROSCI.5370-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curran T, Dien J. Differentiating amodal familiarity from modality-specific memory processes: An ERP study. Psychophysiology. 2003;40:979–988. doi: 10.1111/1469-8986.00116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curran T, Friedman WJ. ERP old/new effects at different retention intervals in recency discrimination tasks. Cognitive Brain Research. 2004;8:107–120. doi: 10.1016/j.cogbrainres.2003.09.006. [DOI] [PubMed] [Google Scholar]
- Curran T, Tanaka JW, Weiskopf DM. An electrophysiological comparison of visual categorization and recognition memory. Cognitive, Affective, & Behavioral Neuroscience. 2002;2:1–18. doi: 10.3758/cabn.2.1.1. [DOI] [PubMed] [Google Scholar]
- Curran T, Tepe KL, Piatt C. ERP explorations of dual processes in recognition memory. In: Zimmer HD, Mecklinger A, Lindenberger U, editors. Binding in Human Memory: A Neurocognitive Approach. Oxford: Oxford University Press; 2006b. pp. 467–492. [Google Scholar]
- Dennis S, Humphreys MS. A context noise model of episodic word recognition. Psychological Review. 2001;108:452–478. doi: 10.1037/0033-295x.108.2.452. [DOI] [PubMed] [Google Scholar]
- Dien J. Issues in the application of the average reference: Review, critiques, and recommendations. Behavior Research Methods, Instruments and Computers. 1998;30:34–43. [Google Scholar]
- Dien J, Santuzzi AM. Application of repeated measures ANOVA to high-density ERP datasets: A review and tutorial. In: Handy TC, editor. Event-related potentials: A methods handbook. Cambridge, MA: MIT Press; 2005. pp. 57–82. [Google Scholar]
- Endl W, Walla P, Lindinger G, Lalouschek W, Barth FG, Deecke L, et al. Early cortical activation indicates preparation for retrieval of memory for faces: an event-related potential study. Neurosci Lett. 1998;240:58–60. doi: 10.1016/s0304-3940(97)00920-8. [DOI] [PubMed] [Google Scholar]
- Friedman D, Johnson R., Jr Event-related potential (ERP) studies of memory encoding and retrieval: A selective review. Microscopy Research and Technique. 2000;51:6–28. doi: 10.1002/1097-0029(20001001)51:1<6::AID-JEMT2>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- Gillund G, Shiffrin RM. A retrieval model for both recognition and recall. Psychological Review. 1984;91:1–67. [PubMed] [Google Scholar]
- Groh-Bordin C, Zimmer HD, Ecker UK. Has the butcher on the bus dyed his hair? When color changes modulate ERP correlates of familiarity and recollection. Neuroimage. 2006 doi: 10.1016/j.neuroimage.2006.04.215. [DOI] [PubMed] [Google Scholar]
- Hintzman DL. Judgments of frequency and recognition memory in a multiple-trace memory model. Psychological Review. 1988;95:528–551. [Google Scholar]
- Hintzman DL, Curran T. Retrieval dynamics of recognition and frequency judgments: Evidence for separate processes of familiarity and recall. Journal of Memory and Language. 1994;33:1–18. [Google Scholar]
- Hintzman DL, Curran T. When encoding fails: Instructions, feedback, and registration without learning. Memory & Cognition. 1995;23:213–226. doi: 10.3758/bf03197223. [DOI] [PubMed] [Google Scholar]
- Hintzman DL, Curran T, Oppy B. Effects of similarity and repetition on memory: Registration without learning? Journal of Experimental Psychology: Learning, Memory, and Cognition. 1992;18:667–680. doi: 10.1037//0278-7393.18.4.667. [DOI] [PubMed] [Google Scholar]
- Jacoby LL. A process dissociation framework: Separating automatic from intentional uses of memory. Journal of Memory and Language. 1991;30:513–541. [Google Scholar]
- Johansson M, Mecklinger A, Treese AC. Recognition memory for emotional and neutral faces: an event-related potential study. J Cogn Neurosci. 2004;16:1840–1853. doi: 10.1162/0898929042947883. [DOI] [PubMed] [Google Scholar]
- Mandler G. Recognizing: The judgment of previous occurrence. Psychological Review. 1980;87:252–271. [Google Scholar]
- McCarthy G, Wood CC. Scalp distributions of event-related potentials: An ambiguity associated with analysis of variance models. Electroencepholography and Clinical Neurophysiology. 1985;62:203–208. doi: 10.1016/0168-5597(85)90015-2. [DOI] [PubMed] [Google Scholar]
- Mecklinger A. Interfacing mind and brain: a neurocognitive model of recognition memory. Psychophysiology. 2000;37:565–582. [PubMed] [Google Scholar]
- Nessler D, Mecklinger A. ERP correlates of true and false recognition after different retention delays: stimulus- and response-related processes. Psychophysiology. 2003;40:146–159. doi: 10.1111/1469-8986.00015. [DOI] [PubMed] [Google Scholar]
- Nessler D, Mecklinger A, Penney TB. Event related brain potentials and illusory memories: the effects of differential encoding. Cognitive Brain Research. 2001;10:283–301. doi: 10.1016/s0926-6410(00)00049-5. [DOI] [PubMed] [Google Scholar]
- Nessler D, Mecklinger A, Penney TB. Perceptual fluency, semantic familiarity and recognition-related familiarity: an electrophysiological exploration. Cognitive Brain Research. 2005;22:265–288. doi: 10.1016/j.cogbrainres.2004.03.023. [DOI] [PubMed] [Google Scholar]
- Norman KA, O’Reilly RC. Modeling hippocampal and neocortical contributions to recognition memory: A complementary learning systems approach. Psychological Review. 2003;110:611–646. doi: 10.1037/0033-295X.110.4.611. [DOI] [PubMed] [Google Scholar]
- Norman KA, Tepe KL, Nyhus E, Curran T. Event-related potential correlates of interference effects on recognition memory. submitted. [DOI] [PubMed] [Google Scholar]
- Phillips PJ, Moon H, Rizvi SA, Rauss PJ. The FERET evaluation methodology for face recognition algorithms. IEEE Transactions on Pattern Analysis and Machine Intelligence. 2000;22:1090–1104. [Google Scholar]
- Picton TW, Bentin S, Berg P, Donchin E, Hillyard SA, Johnson R, Jr, et al. Guidelines for using human event-related potentials to study cognition: recording standards and publication criteria. Psychophysiology. 2000;37:127–152. [PubMed] [Google Scholar]
- Rugg MD, Mark RE, Walla P, Schloerscheidt AM, Birch CS, Allan K. Dissociation of the neural correlates of implicit and explicit memory. Nature. 1998;392:595–598. doi: 10.1038/33396. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Yonelinas AP. Human recognition memory: a cognitive neuroscience perspective. Trends in Cognitive Sciences. 2003;7:313–319. doi: 10.1016/s1364-6613(03)00131-1. [DOI] [PubMed] [Google Scholar]
- Shiffrin RM, Steyvers M. A model of recognition memory: REM—Retrieving effectively from memory. Psychological Bulletin and Review. 1997;4:145–166. doi: 10.3758/BF03209391. [DOI] [PubMed] [Google Scholar]
- Urbach TP, Kutas M. The intractability of scaling scalp distributions to infer neuroelectric sources. Psychophysiology. 2002;39:791–808. doi: 10.1111/1469-8986.3960791. [DOI] [PubMed] [Google Scholar]
- Urbach TP, Kutas M. Interpreting event-related brain potential (ERP) distributions: implications of baseline potentials and variability with application to amplitude normalization by vector scaling. Biol Psychol. 2006;72:333–343. doi: 10.1016/j.biopsycho.2005.11.012. [DOI] [PubMed] [Google Scholar]
- Voss JL, Paller KA. Fluent conceptual processing and explicit memory for faces are electrophysiologically distinct. Journal of Neuroscience. 2006;26:926–933. doi: 10.1523/JNEUROSCI.3931-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilding EL. The practice of rescaling scalp-recorded event-related potentials. Biol Psychol. 2006;72:325–332. doi: 10.1016/j.biopsycho.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Wilding EL, Sharpe H. Episodic memory encoding and retreival: Recent insights from event-related potentials. In: Zani A, Proverbio AM, editors. The cognitive electrophysiology of mind and brain. San Diego, CA: Academic Press; 2003. pp. 169–196. [Google Scholar]
- Winer BJ. Statistical principles in experimental design. 2. New York: McGraw-Hill; 1971. [Google Scholar]
- Yonelinas AP. The nature of recollection and familiarity: A review of 30 years of research. Journal of Memory and Language. 2002;46:441–517. [Google Scholar]
- Yovel G, Paller KA. The neural basis of the butcher-on-the-bus phenomenon: when a face seems familiar but is not remembered. NeuroImage. 2004;21:789–800. doi: 10.1016/j.neuroimage.2003.09.034. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.