

Abstract
Host responses to biomaterials control the biological performance of implanted medical devices. Upon implantation, synthetic materials adsorb biomolecules which trigger an inflammatory cascade comprising coagulation, leukocyte recruitment/adhesion, and foreign body reaction. The foreign body reaction and ensuing fibrous encapsulation severely limit the in vivo performance of numerous biomedical devices. While it is well established that plasma fibrinogen and secreted cytokines modulate leukocyte recruitment and maturation into foreign body giant cells, mediators of chronic inflammation and fibrous encapsulation of implanted biomaterials remain poorly understood. Using plasma fibronectin conditional knock-out mice, we demonstrate that plasma fibronectin modulates the foreign body response to polyethylene terephthalate discs implanted subcutaneously. Fibrous collagenous capsules were two-fold thicker in mice depleted of plasma fibronectin compared to controls. In contrast, deletion of plasma fibronectin did not alter acute leukocyte recruitment to the biomaterial, indicating that plasma fibronectin modulates chronic fibrotic responses. The number of foreign body giant cells associated with the implant was three times higher in the absence of plasma fibronectin while macrophage numbers were not different, suggesting that plasma fibronectin regulates the formation of biomaterial-associated foreign body giant cells. Interestingly, cellular fibronectin was present in the capsules of both normal and plasma fibronectin-depleted mice, suggesting that cellular fibronectin could not compensate for the loss of plasma fibronectin. These results implicate plasma fibronectin in the host response to implanted materials and identify a potential target for therapeutic intervention to enhance the biological performance of biomedical devices.
INTRODUCTION
Host responses to biomaterials control the biological performance of implanted medical devices, tissue-engineered constructs, and delivery vehicles for therapeutics [1,2]. Upon implantation, synthetic materials dynamically adsorb proteins and other biomolecules which trigger an inflammatory cascade comprising blood coagulation, leukocyte recruitment and adhesion, foreign body reaction, and fibrous encapsulation [1,2]. The foreign body reaction and ensuing fibrous encapsulation result in a physicochemical barrier that severely limits device integration and the in vivo performance of numerous devices, including chemical biosensors, electrical leads/electrodes, therapeutic delivery systems, and orthopaedic and cardiovascular prostheses [3-6]. Extensive research has identified mechanisms governing acute inflammatory responses to implanted synthetic materials. Adsorption of fibrinogen and complement fragments from plasma onto biomaterial surfaces results in integrin receptor-mediated leukocyte recruitment and adhesion [7-9]. Adherent monocytes/macrophages secrete cytokines and growth factors and mature into foreign body giant cells that coordinate the recruitment and activities of other cell types [10,11], leading to neovascularization and connective tissue formation [2]. Despite our understanding of acute inflammation to implanted synthetic materials, mediators of chronic inflammation and fibrous encapsulation of implanted biomaterials remain poorly understood.
Fibronectins (FNs) are widely expressed, cell-adhesive glycoproteins present as soluble forms in body fluids (e.g., plasma FN, pFN) and insoluble fibrils in extracellular matrices (cellular FN, cFN) [12]. FNs are generated from a single gene, but alternative splicing gives rise to different isoforms [12]. Deletion of the FN gene is embryonically lethal due to defects in mesoderm, neural tube and vascular development [13]. FN is also required for cleft formation during epithelial branching morphogenesis [14]. Extensive in vitro analyses have demonstrated that FNs promote cell adhesion and regulate the survival, cell cycle progression, and expression of differentiated phenotypes in various cell types. Despite the vast amounts of studies on the role of FN in cellular functions, the role of the pFN isoform in adult physiology and pathology has been difficult to analyze because of the embryonic lethality of the FN gene deletion. Recent studies with FN conditional knock-out mice have shown that pFN promotes thrombus growth and stability in injured arterioles and supports neuronal survival following cerebral ischemia, but is not essential to skin-wound healing, likely due to contributions from cFN [15,16]. In the present analysis, we used pFN conditional knock-out mice to examine the contributions of pFN to host responses to implanted biomaterials.
MATERIALS AND METHODS
pFN conditional knockout mice
pFN conditional knock-out mice based on the Cre-loxP system were previously developed by Erickson and Fässler [15] and rederived by Hynes [16]. The Cre-loxP system provides a genetic tool to control site-specific recombination events in genomic DNA, thereby affording a mechanism for deleting a specific gene in response to a stimulus that results in Cre recombinase expression. These mice have the FN gene flanked by loxP sites and express Cre recombinase under control of the interferon- and polyinosinic-polycytidic acid (pI-pC)-inducible Mx promoter. Deletion of the FN gene is induced by intraperitoneal injections of pI-pC and results in complete and stable deletion of FN in the liver (hepatocytes produce pFN) for at least eight months [15]. Breeding pairs of mice containing a floxed (fl; loxP-site containing) fibronectin allele and Mx-Cre were kindly provided by H.P. Erickson and R.O. Hynes. Floxed mice were crossed with Mx-Cre mice to generate FN (fl/fl) animals carrying the Mx-Cre transgene. The presence of the transgene was verified by PCR of DNA extracted from tail tissue with Cre-specific primers. Deletion of the FN gene was induced in 8-12 week-old female mice by 3 intraperitoneal injections of pI-pC (250 μg) at 2 day intervals. Blood samples were drawn from the saphenous vein from intraperitoneal space at least 3 days following the last pI-pC injection to assess pFN levels by Western blotting using a rabbit polyclonal antibody against human fibronectin (Sigma; this antibody cross-reacts with mouse fibronectin). Lavage from intraperitoneal space was obtained from animals at the time of disk explant. All experiments were conducted under IACUC-approved protocols.
Biomaterial implantation and analysis
Discs (9.5 mm diameter, 0.5 mm thick) were cut from PET sheets, washed, and sterilized in ethanol. Samples had endotoxin levels below the recommended maximum FDA level (0.5 EU/ml) as determined by the LAL chromogenic assay. Discs were implanted either intraperitoneally (n = 8 samples/group) or subcutaneously (n = 4-5 samples/group) following IACUC-approved procedures. For intraperitoneal implants, discs were explanted at 16 hours. Adherent cells were trypsinized and either analyzed in a Coulter counter (for total leukocyte cell counts) or fixed and stained with May-Grunwald-Geimsa for differential cell counts in cytospins. For subcutaneous implants, PET discs were explanted at 14 days, formalin-fixed and paraffin-embedded. Histological sections (5 μm thick) were stained with Verhoeff-van Geisson stain for nuclei (dark blue) and collagen (pink). For immunohistochemical staining, sections were incubated in rabbit polyclonal antibody against fibronectin (Sigma, St. Louis, MO) or rat monoclonal antibody against the F4/80 antigen of resident murine tissue macrophages (clone BM8, Accurate Chemical & Scientific), biotinylated secondary antibodies, and an avidin-linked alkaline phosphatase-based reagent (Vector Labs, Burlingame, CA), and counterstained with hematoxylin. For cell macrophage/FBGC counting, high magnification (60X oil objective) images were blindly scored for total nuclei, F4/80+ cells with one nucleus, and F4/80+ multinucleated cells.
Statistical Analysis
Results were analyzed by ANOVA using SYSTAT 8.0 (SPSS Inc., Chicago, IL). For macrophage/FBGC, a nested one-way ANOVA design (animal nested within treatment) was used to account for the variance across subjects [33]. Pair-wise comparisons were performed using Tukey post-hoc test with a 95% confidence level considered significant.
RESULTS
We used pFN conditional knock-out mice based on the Cre-loxP system [15,16] to examine the role of pFN in host responses to implanted biomaterials. Deletion of the FN gene in these mice is induced by intraperitoneal injections of pI-pC and results in complete and stable deletion of FN in the liver (hepatocytes produce pFN) for at least eight months [15]. The extent of FN deletion in other tissues is variable, and some cell types retain the ability to produce cFN. Importantly, these mice express normal levels of other extracellular matrix components such as collagen I, laminin-1, and tenascin. Following pI-pC induction, these mice exhibit less than 2% of pFN in normal mouse plasma (Fig. 1a) and display no overt phenotype under standard laboratory conditions. As expected, deletion of pFN also eliminated FN in the intraperitoneal fluid (Fig. 1b). We refer to these mice as pFN-null mice throughout this paper. No differences between untreated (no pI-pC treatment) mice carrying the Mx-Cre transgene and pI-pC-treated wild-type littermates were observed in any assay, and we refer to these mice as control throughout this manuscript.
Fig. 1.
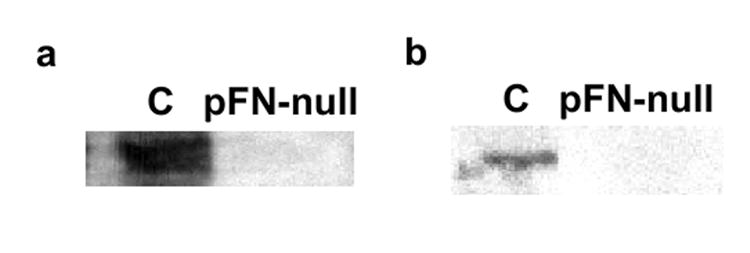
pI-pc induction effectively deletes FN from (a) plasma and (b) intraperitoneal fluid in mice carrying the Mx-Cre transgene (pFN-null) but not control (C) animals. Western blot analysis with polyclonal antibody against fibronectin demonstrated approximately 98% reduction of FN levels.
To investigate the role of pFN on leukocyte recruitment during the acute inflammatory response to biomaterials, polyethylene terephthalate (PET) discs were implanted in the intraperitoneal space for 16 hours and then explanted for analysis of leukocyte recruitment and adhesion. This model has been extensively used to analyze leukocyte recruitment to biomaterials [8,17,18]. PET is a widely used biomaterial which elicits a moderately strong inflammatory response [8]. The knitted form of this material, Dacron®, is widely used in vascular grafts. No differences were observed in total leukocyte counts of cells attached to implanted discs between pFN-null and control mice (p = 0.79) (Fig. 2a). Furthermore, differential cell counts revealed no differences in lymphocyte, neutrophil, and monocyte adhesion to PET discs between pFN-null and control mice (p = 0.19) (Fig. 2b). These results indicate that pFN does not play a major role in the recruitment and adhesion of leukocytes to implanted materials during the acute inflammatory response.
Fig. 2.
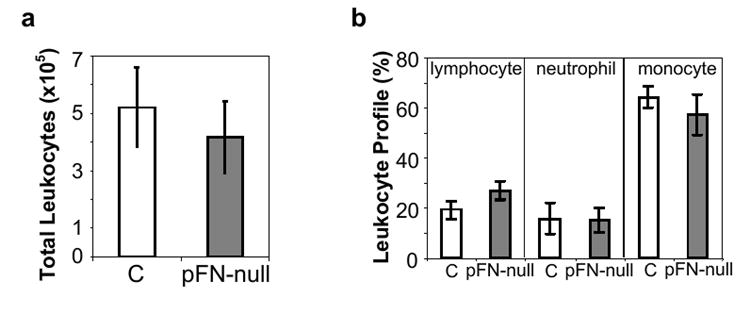
pFN does not influence acute leukocyte recruitment and adhesion to implanted biomaterials. PET discs were implanted in the intraperitoneal cavity in mice with pFN deleted (pFN-null) and control (C) animals. No differences were observed in adherent (a) total or (b) differential leukocyte cell counts (n = 8 animals/group).
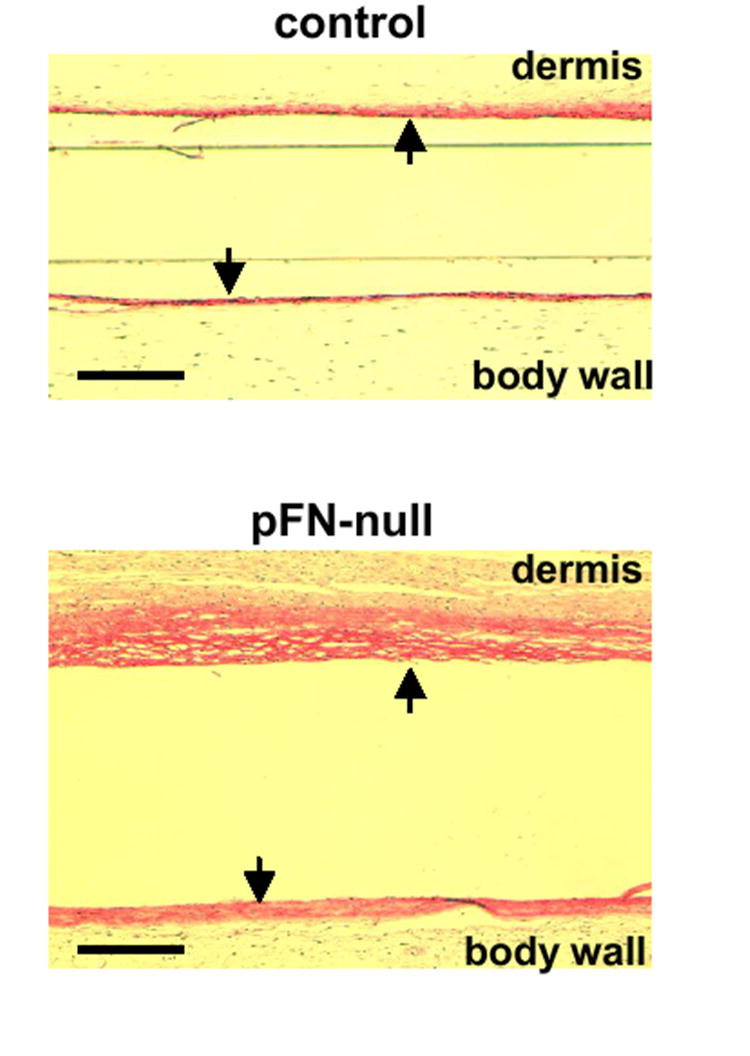
PET discs were also implanted subcutaneously for 14 days to assess the contributions of pFN to the foreign body reaction and fibrous encapsulation of implanted materials. Measurement of fibrous capsule thickness following subcutaneous implantation is a standard measure of chronic inflammation to synthetic materials [2]. PET discs implanted in pFN-null mice were encapsulated by thick, dense fibrous membranes (Fig. 3a), while discs implanted in control animals exhibited considerably thinner capsules (Fig. 3b). Fibrous capsules were thicker on the implant side facing the dermis compared to the implant side facing the body wall as is often seen in this model, but the relative difference in capsule thickness between pFN-null and control animals was equivalent. Measurements of capsule thickness indicated a 2-fold increase in fibrous capsule thickness for pFN-null mice compared to controls (dermis, p < 0.04; body wall, p < 0.02) (Fig. 3c,d). These results indicate that pFN modulates the foreign body reaction and fibrotic response to implanted materials.
Fig. 3.
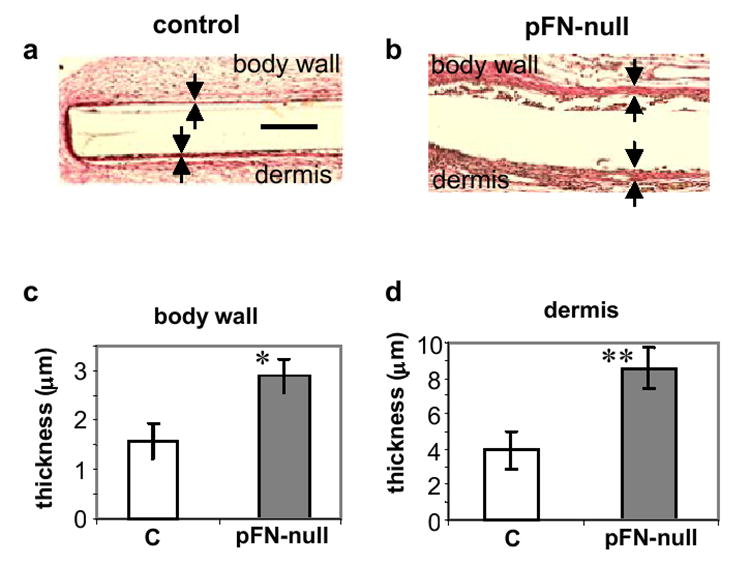
pFN modulates the foreign body reaction and fibrotic response to PET discs implanted subcutaneously for 14 days. Verhoeff-van Geisson stained sections (collagen: pink; cell nuclei: dark blue) of tissue response to implanted biomaterials showing fibrous capsules (arrows) in (a) control (C) and (b) mice with deleted pFN (pFN-null). Measurement of capsule thickness revealed thicker capsules around PET implants in pFN-null mice compared to controls (C) for both (c) body wall (p < 0.02) and (d) dermis (p < 0.04) faces of the implants (scale bar, 50 μm; n = 5 animals/group).
To evaluate whether pFN is involved in macrophage recruitment and fusion into foreign body giant cells (FBGC), sections were stained for the F4/80 antigen (a marker of resident tissue macrophages) and scored for total nuclei, F4/80+ cells with one nucleus (macrophages), and F4/80+ cells containing multiple nuclei (FBGC). More intense staining for the macrophage marker F4/80 antigen was observed for mice lacking pFN than control animals (Fig. 4a,b). A three-fold increase in the number of FBGC associated with the implant was detected in pFN-null mice compared to controls (p < 0.04) (Fig. 4c). No differences were observed in the number of total cells (p = 0.40) or macrophages (p = 0.45). This result suggests that pFN regulates the formation of biomaterial-associated FBGC.
Fig. 4.
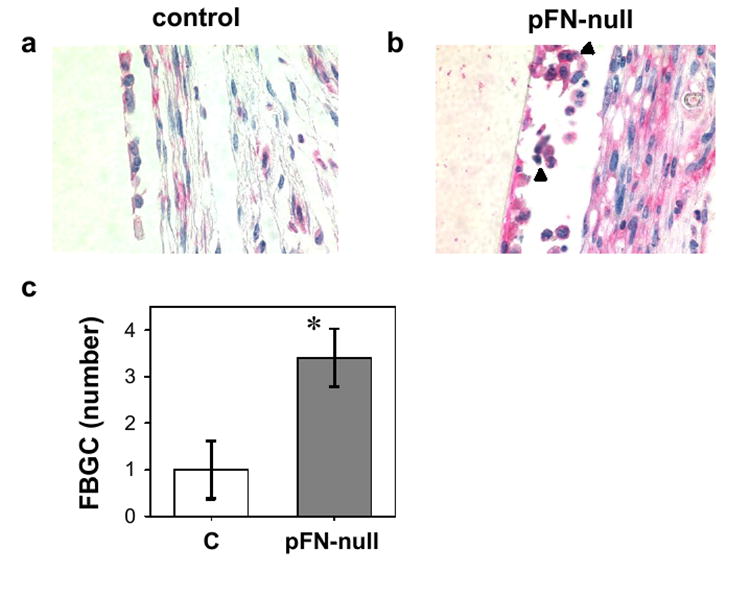
Biomaterial-associated macrophages and FBGC. Immunohistochemical staining for macrophage marker F4/80 (pink) is more intense in pFN-null animals (panel a) compared to control mice (panel b). Multi-nucleated, F4/80+ cells are indicated by black arrowhead. (c) Cell counts showing elevated biomaterial-associated FBGC in pFN-null mice (p < 0.04) (n = 4 animals/group).
Immunohistochemical staining for FN showed intense staining localized to the fibrous capsule for both pFN-null and control animals (Fig. 5). FN was distributed throughout the dense fibrous capsule associated with the biomaterial. Since the polyclonal antibody used recognizes both pFN and cFN, the staining most likely indicates the presence of cFN within the fibrous capsule. As discussed above, for this conditional knock-out model, some tissues retain expression of cFN even though pFN is effectively deleted [15]. Monocytes/macrophages and fibroblasts associated with the fibrous capsule are the likely source of this cFN.
Fig. 5.
Immunohistochemical staining for FN (arrows, dark pink) in fibrous capsules associated with PET discs implanted subcutaneously (scale bar, 50 μm).
DISCUSSION
Our results demonstrate that pFN modulates the foreign body response and fibrous encapsulation of implanted materials. This is the first report directly linking pFN to host inflammatory responses to implanted synthetic materials. Deletion of pFN, however, did not influence leukocyte recruitment and adhesion to the implanted biomaterial, indicating that pFN regulates chronic, but not acute, inflammatory responses to biomaterials. The lack of involvement of pFN in acute leukocyte adhesion to synthetic materials in vivo is consistent with other in vitro and in vivo analyses showing that fibrinogen is the major plasma component mediating leukocyte recruitment to biomaterials [8,19]. Unexpectedly, collagenous capsules surrounding implanted PET discs were twice as thick in pFN-null mice compared to wild-type mice. This finding that collagen capsules are thicker in the absence of pFN is surprising given the requirement of FN on collagen assembly [20,21]. This result indicates that pFN is involved in the deposition and/or remodeling of the fibrous capsule surrounding implanted materials. The mechanism(s) by which pFN regulates fibrous capsule thickness remains unclear, but pFN may modulate host responses by adsorbing directly onto the material or by interacting with other proteins associated with the biomaterial, such as fibrinogen and collagen. pFN associated with the biomaterial may influence the adhesion and/or function of cells involved in the foreign body reaction, including the ability to assemble and remodel the collagenous capsule or secrete cytokines regulating fibrosis. Indeed, we observed a significant increase in the number of FBGC associated with the implant in pFN-null compared to control mice. This result suggests that pFN regulates the formation or maturation of FBGC. This model is consistent with in vitro findings implicating fibronectin-binding β1 integrins and fibronectin-derived binding motifs in FBGC fusion and maturation [10,22]. Moreover, Horbett and colleagues demonstrated that monocyte adhesion to adsorbed fibronectin reduced FBGC formation [34].
Interestingly, considerable levels of FN, corresponding to cFN, were present on the capsules of both pFN-null and control mice. This observation suggests that the differences in fibrous capsule thickness are specific to pFN, and cFN cannot compensate for the loss of pFN. There is evidence that different isoforms of FN support cell adhesion and migration to different extents [23-26]. Finally, host responses to implanted materials are often interpreted in the context of classical wound healing. However, the important role of pFN in the inflammatory response and foreign body reaction observed in our study contrasts with the ability of skin wounds to heal normally in the absence of pFN in the same animal model [15]. These results highlight inherent differences between classical wound healing and host responses to implanted materials.
The use of a conditional knock-out model provides a robust system to evaluate the role of specific proteins in host responses to implanted materials in adult animals without complicating compensatory effects associated with conventional knock-out models. For example, Bornstein and colleagues demonstrated that mice lacking the angiogenesis inhibitor thrombospondin 2 mount an altered foreign body reaction to implanted materials characterized by increased capsule thickness and vascularity [27]. However, interpretation of the results is complicated by the fact that these mice exhibit connective tissue abnormalities, including disordered collagen fibrillogenesis and vascular density [28].
Our results identify a potential target for therapeutic intervention to enhance the biological performance of biomedical devices. Control of pFN activity via drug-, protein-, or gene-based manipulations of plasma concentration, availability, or presentation on the material could be exploited to manipulate host responses. For instance, we have shown that biomaterial surface chemistry-dependent changes in the activity of adsorbed FN modulate integrin receptor binding and cell differentiation responses [29]. Moreover, biomaterial-based strategies focusing on presenting FN or FN-derived adhesion motifs enhance host integration and function in model systems, including bone and cartilage [30-32]. These enhancements are attributed to improved interactions with the host target tissue (e.g., bone) rather than altered inflammatory responses. Our results support pFN-mediated regulation of the foreign body reaction and chronic inflammatory responses as a new mechanism for controlling host responses to synthetic biomedical materials.
CONCLUSION
Using plasma fibronectin conditional knock-out mice, we demonstrate that plasma fibronectin modulates the foreign body response to biomaterial discs implanted subcutaneously. Fibrous collagenous capsules were two-fold thicker and three-fold higher numbers of foreign body giant cells were observed in mice depleted of plasma fibronectin compared to controls. In contrast, deletion of plasma fibronectin did not alter acute leukocyte recruitment to the biomaterial, indicating that plasma fibronectin modulates chronic fibrotic responses. These results implicate plasma fibronectin in the host response to implanted materials and identify a potential target for therapeutic intervention to enhance the biological performance of biomedical devices.
Acknowledgments
Funding provided by the NSF-sponsored Georgia Tech/Emory Center on the Engineering of Living Tissues (ECC-9731643) and NIH (R01 EB-004496). AWB is the recipient of a NSF Graduate Fellowship. Mice containing a floxed (fl; loxP-site containing) fibronectin allele and Mx-Cre were kindly provided by H.P. Erickson and R.O. Hynes.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Ratner BD, Bryant SJ. Biomaterials: where we have been and where we are going. Annu Rev Biomed Eng. 2004;6:41–75. doi: 10.1146/annurev.bioeng.6.040803.140027. [DOI] [PubMed] [Google Scholar]
- 2.Anderson JM. Biological responses to materials. Annu Rev Mater Res. 2001;31:81–110. [Google Scholar]
- 3.Bauer TW, Schils J. The pathology of total joint arthroplasty.II. Mechanisms of implant failure. Skeletal Radiol. 1999;28:483–497. doi: 10.1007/s002560050552. [DOI] [PubMed] [Google Scholar]
- 4.Gorbet MB, Sefton MV. Biomaterial-associated thrombosis: roles of coagulation factors, complement, platelets and leukocytes. Biomaterials. 2004;25:5681–5703. doi: 10.1016/j.biomaterials.2004.01.023. [DOI] [PubMed] [Google Scholar]
- 5.Polikov VS, Tresco PA, Reichert WM. Response of brain tissue to chronically implanted neural electrodes. J Neurosci Methods. 2005;148:1–18. doi: 10.1016/j.jneumeth.2005.08.015. [DOI] [PubMed] [Google Scholar]
- 6.Wu P, Grainger DW. Drug/device combinations for local drug therapies and infection prophylaxis. Biomaterials. 2006;27:2450–2467. doi: 10.1016/j.biomaterials.2005.11.031. [DOI] [PubMed] [Google Scholar]
- 7.Hu WJ, Eaton JW, Ugarova TP, Tang L. Molecular basis of biomaterial-mediated foreign body reactions. Blood. 2001;98:1231–1238. doi: 10.1182/blood.v98.4.1231. [DOI] [PubMed] [Google Scholar]
- 8.Tang L, Eaton JW. Fibrin(ogen) mediates acute inflammatory responses to biomaterials. J Exp Med. 1993;178:2147–2156. doi: 10.1084/jem.178.6.2147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.McNally AK, Anderson JM. Complement C3 participation in monocyte adhesion to different surfaces. Proc Natl Acad Sci U S A. 1994;91:10119–10123. doi: 10.1073/pnas.91.21.10119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McNally AK, Anderson JM. Beta1 and beta2 integrins mediate adhesion during macrophage fusion and multinucleated foreign body giant cell formation. Am J Pathol. 2002;160:621–630. doi: 10.1016/s0002-9440(10)64882-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.McNally AK, Anderson JM. Interleukin-4 induces foreign body giant cells from human monocytes/macrophages. Differential lymphokine regulation of macrophage fusion leads to morphological variants of multinucleated giant cells. Am J Pathol. 1995;147:1487–1499. [PMC free article] [PubMed] [Google Scholar]
- 12.Hynes RO. Fibronectins . New York: Springer-Verlag; 1990. [Google Scholar]
- 13.George EL, Georges-Labouesse EN, Patel-King RS, Rayburn H, Hynes RO. Defects in mesoderm, neural tube and vascular development in mouse embryos lacking fibronectin. Development. 1993;119:1079–1091. doi: 10.1242/dev.119.4.1079. [DOI] [PubMed] [Google Scholar]
- 14.Sakai T, Larsen M, Yamada KM. Fibronectin requirement in branching morphogenesis. Nature. 2003;423:876–881. doi: 10.1038/nature01712. [DOI] [PubMed] [Google Scholar]
- 15.Sakai T, Johnson KJ, Murozono M, Sakai K, Magnuson MA, Wieloch T, Cronberg T, Isshiki A, Erickson HP, Fassler R. Plasma fibronectin supports neuronal survival and reduces brain injury following transient focal cerebral ischemia but is not essential for skin-wound healing and hemostasis. Nat Med. 2001;7:324–330. doi: 10.1038/85471. [DOI] [PubMed] [Google Scholar]
- 16.Ni H, Yuen PS, Papalia JM, Trevithick JE, Sakai T, Fassler R, Hynes RO, Wagner DD. Plasma fibronectin promotes thrombus growth and stability in injured arterioles. Proc Natl Acad Sci U S A. 2003;100:2415–2419. doi: 10.1073/pnas.2628067100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tang L, Jennings TA, Eaton JW. Mast cells mediate acute inflammatory responses to implanted biomaterials. Proc Natl Acad Sci U S A. 1998;95:8841–8846. doi: 10.1073/pnas.95.15.8841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tang L, Lucas AH, Eaton JW. Inflammatory responses to implanted polymeric biomaterials: role of surface-adsorbed immunoglobulin G. J Lab Clin Med. 1993;122:292–300. [PubMed] [Google Scholar]
- 19.Shen M, Horbett TA. The effects of surface chemistry and adsorbed proteins on monocyte/macrophage adhesion to chemically modified polystyrene surfaces. J Biomed Mater Res. 2001;57:336–345. doi: 10.1002/1097-4636(20011205)57:3<336::aid-jbm1176>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- 20.Velling T, Risteli J, Wennerberg K, Mosher DF, Johansson S. Polymerization of type I and III collagens is dependent on fibronectin and enhanced by integrins alpha 11beta 1 and alpha 2beta 1. J Biol Chem. 2002;277:37377–37381. doi: 10.1074/jbc.M206286200. [DOI] [PubMed] [Google Scholar]
- 21.Sottile J, Hocking DC. Fibronectin polymerization regulates the composition and stability of extracellular matrix fibrils and cell-matrix adhesions. Mol Biol Cell. 2002;13:3546–3559. doi: 10.1091/mbc.E02-01-0048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kao WJ, Lee D, Schense JC, Hubbell JA. Fibronectin modulates macrophage adhesion and FBGC formation: the role of RGD, PHSRN, and PRRARV domains. J Biomed Mater Res. 2001;55:79–88. doi: 10.1002/1097-4636(200104)55:1<79::aid-jbm110>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 23.Chauhan AK, Moretti FA, Iaconcig A, Baralle FE, Muro AF. Impaired motor coordination in mice lacking the EDA exon of the fibronectin gene. Behav Brain Res. 2005;161:31–38. doi: 10.1016/j.bbr.2005.02.020. [DOI] [PubMed] [Google Scholar]
- 24.Hashimoto-Uoshima M, Yan YZ, Schneider G, Aukhil I. The alternatively spliced domains EIIIB and EIIIA of human fibronectin affect cell adhesion and spreading. J Cell Sci. 1997;110(Pt 18):2271–2280. doi: 10.1242/jcs.110.18.2271. [DOI] [PubMed] [Google Scholar]
- 25.Liao YF, Gotwals PJ, Koteliansky VE, Sheppard D, Van De WL. The EIIIA segment of fibronectin is a ligand for integrins alpha 9beta 1 and alpha 4beta 1 providing a novel mechanism for regulating cell adhesion by alternative splicing. J Biol Chem. 2002;277:14467–14474. doi: 10.1074/jbc.M201100200. [DOI] [PubMed] [Google Scholar]
- 26.Manabe R, Ohe N, Maeda T, Fukuda T, Sekiguchi K. Modulation of cell-adhesive activity of fibronectin by the alternatively spliced EDA segment. J Cell Biol. 1997;139:295–307. doi: 10.1083/jcb.139.1.295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kyriakides TR, Leach KJ, Hoffman AS, Ratner BD, Bornstein P. Mice that lack the angiogenesis inhibitor, thrombospondin 2, mount an altered foreign body reaction characterized by increased vascularity. Proc Natl Acad Sci U S A. 1999;96:4449–4454. doi: 10.1073/pnas.96.8.4449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kyriakides TR, Zhu YH, Smith LT, Bain SD, Yang Z, Lin MT, Danielson KG, Iozzo RV, LaMarca M, McKinney CE, Ginns EI, Bornstein P. Mice that lack thrombospondin 2 display connective tissue abnormalities that are associated with disordered collagen fibrillogenesis, an increased vascular density, and a bleeding diathesis. J Cell Biol. 1998;140:419–430. doi: 10.1083/jcb.140.2.419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Keselowsky BG, Collard DM, Garcia AJ. Integrin binding specificity regulates biomaterial surface chemistry effects on cell differentiation. Proc Natl Acad Sci U S A. 2005;102:5953–5957. doi: 10.1073/pnas.0407356102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Alsberg E, Anderson KW, Albeiruti A, Rowley JA, Mooney DJ. Engineering growing tissues. Proc Natl Acad Sci U S A. 2002;99:12025–12030. doi: 10.1073/pnas.192291499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lutolf MP, Weber FE, Schmoekel HG, Schense JC, Kohler T, Muller R, Hubbell JA. Repair of bone defects using synthetic mimetics of collagenous extracellular matrices. Nat Biotechnol. 2003;21:513–518. doi: 10.1038/nbt818. [DOI] [PubMed] [Google Scholar]
- 32.Erli HJ, Ruger M, Ragoss C, Jahnen-Dechent W, Hollander DA, Paar O, von Walter M. The effect of surface modification of a porous TiO2/perlite composite on the ingrowth of bone tissue in vivo. Biomaterials. 2006;27:1270–1276. doi: 10.1016/j.biomaterials.2005.08.001. [DOI] [PubMed] [Google Scholar]
- 33.Sokal RR, Rohlf FJ. Biometry: The Principles and Practice of Statistics in Biological Research. W.H. Freeman; New York, NY: 1995. [Google Scholar]
- 34.Shen M, Garcia I, Maier RV, Horbett TA. The effects of adsorbed proteins and surface chemistry on foreign body giant cell formation, TNFα release, and procoagulant activity of monocytes. J Biomed Mater Res. 2004;70A:533–541. doi: 10.1002/jbm.a.30069. [DOI] [PubMed] [Google Scholar]