

Abstract
Much experimental and clinical data suggest that the pharmacological profile of modafinil, a newly discovered waking substance, differs from those of amphetamine and methylphenidate, two classical psychostimulants. The brain targets on which modafinil acts to induce wakefulness, however, remain unknown. A double-blind study using the protooncogene c-fos as experimental marker in the cat was, therefore, carried out to identify the potential target neurons of modafinil and compare them with those for amphetamine and methylphenidate. Cats were sacrificed after a single oral administration of amphetamine, methylphenidate, or modafinil at equivalent doses for wake induction (1, 2.5, or 5 mg/kg, respectively) and brain sections examined for Fos by immunocytochemistry. Administration of either amphetamine or methylphenidate evoked Fos-like immunoreactivity in a large number of neurons in the striatum and whole cortex, especially in the caudate nucleus and mediofrontal cortex, which are known to be dopaminergic targets. In contrast, administration of modafinil resulted in the labeling of few cells in these structures, but did induce marked Fos labeling in neurons of the anterior hypothalamic nucleus and adjacent areas. These results provide evidence for the potential brain targets of modafinil, which differ from those of amphetamine or methylphenidate, and suggest that modafinil induces wakefulness by mechanisms distinct from those of the two stimulants.
Keywords: immediate-early gene, protooncogene, anterior hypothalamic nucleus, striatum, dopaminergic system
Modafinil, a newly discovered wake-improving substance, induces prolonged wakefulness in a number of species, apparently without associated behavioral excitation (1, 2, 3, 4). Unlike amphetamine, a psychomotor stimulant, its waking effect is not followed by an obvious sleep rebound in the cat (4), either under normal conditions or after sleep deprivation. Moreover, pretreatment with α-methylparatyrosine, an inhibitor of catecholamine synthesis prevents the arousal effect of amphetamine but allows that of modafinil to persist (4). Although these results indicate the different pharmacological profiles of modafinil and amphetamine, the underlying mechanisms remain unknown, especially as regards the brain targets by which modafinil induces waking. This is mainly because conventional methods, such as autoradiography, are currently unable to reveal its central sites of action.
Recently, visualization of the expression of immediate-early genes (IEGs) has become a promising method in neuroscience. The proto-oncogene, c-fos, is the most frequently activated IEG in the central nervous system following various internal or external stimuli. c-fos encodes a nuclear phosphoprotein, Fos, which is thought to control target gene expression responsible for cell activation. c-fos has, therefore, been considered as the third messenger, reacting to short-term stimuli and triggering long-term cell reaction cascades (5, 6, 7). Although a basic level of expression of IEGs occurs in the brain, c-fos is strongly expressed in various specific cerebral areas following different stimuli, such as electrical or pharmacological stimulation, stress, or sleep deprivation (8, 9, 10, 11, 12, 13, 14, 15). Thus, visualization of c-fos could serve as a sensitive indicator of gene activation in individual target neurons activated following a given stimulus.
We have, therefore, developed Fos immunocytochemical procedures in both the rat and cat to visualize the potential targets of modafinil and amphetamine in the central nervous system. This comparative study also included methylphenidate, a psychostimulant with a waking effect and certain amphetamine-like characteristics.
MATERIALS AND METHODS
Rats and cats were both used in this study. However, we quickly found in pilot experiments with rats that the stress associated with the route of administration (which has to be intraperitoneal) caused problems, as even a simple injection of physiological saline resulted in strong c-fos expression in a number of cerebral structures (see also ref. 9). In contrast, oral administration is well tolerated by cats, provided that it is very gently and quickly performed, since the animal usually falls asleep soon after (19.8 ± 6.9 min; n = 6) placebo administration, which results in very little Fos immunoreactivity (see Results). Moreover, we have determined in the cat by polygraphic recordings the parameters required for such a study, such as drug dose and duration for amphetamine-, methylphenidate-, and modafinil-induced wakefulness (ref. 4 and data as follows).
The experiments were therefore performed on 16 drug-naive adult male cats. Three days before administration, the animals were put in our usual experimental conditions for sleep–wake recordings. They were placed in a sound-proofed cage (ambient temperature 25°C; 12:12-h light/dark cycle with light on at 7 a.m.) and fed each day at 6 p.m. with a standard normal diet. All external stimuli (e.g., auditory, visual, alimentary, tactile) were strictly avoided. The doses of amphetamine (Sigma), methylphenidate (Sigma), and modafinil (Laboratoire L. Lafon, Maisons Alfozt, France) were set at 1, 2.5, and 5 mg/kg because, at these doses, they induce a continuous waking state of similar duration: 586.3 ± 40.1 min, 579.8 ± 35.1 min, and 596.2 ± 42.1 min, respectively (n = 6; ANOVA, P > 0.1; see also ref. 4). The placebo (lactose) and the drugs used were coded and randomized so that the experiment, cell plotting, and photomicrographs were performed blind. All were administered by the oral route at 11 a.m. and the cats sacrificed 90 or 150 (n = 8) min later, since c-fos expression is generally induced 30 min after cell stimulation, reaches a maximum after 1–2 h, and is maintained for 3–6 h, depending on the nature and intensity of the stimulus (5, 6, 7). Animals were deeply anaesthetized (Nembutal, 50 mg/kg i.v.) and perfused through the ascending aorta with 1 liter of Ringer’s lactate solution containing 0.1% heparin, followed by 2.5 liters of 0.1 M phosphate buffer (PB, pH 7.4, 4°C)/2-4% paraformaldehyde/0.1% glutaraldehyde. After overnight postfixation and rinsing for 48 h with PB/30% sucrose, the brains were coronally sectioned (20 μm) on a cryostat at −25°C. After rinsing, the floating sections were incubated with stirring for 48–72 h at 4°C with a rabbit anti-c-fos antibody (AB-2; Oncogene Science), diluted 1:10,000 to 1:40,000 in PBS/0.3% Triton X-100 (PBST)/0.1% sodium azide. Following rinsing, the sections were incubated with a biotinylated anti-rabbit IgG (Vector Laboratories), diluted 1:1000 to 1:3000, rinsed, then incubated with a horseradish peroxidase conjugated avidin-biotin complex (Vector Laboratories; diluted 1:1000 to 1:2000). Both incubations were in PBST at 4°C overnight with stirring. The sections were then immersed in 0.05 M Tris·HCl buffer (pH 7.6) containing 0.02% 3,3′ diaminobenzidine-4HCl, 0.003% H2O2, and 0.6% nickel ammonium sulfate for 6–12 min at room temperature. Finally, the sections were mounted, dried, dehydrated, and covered with a coverslip, using DePeX (BDH), for light microscopy. Some of them were counterstained with neutral red to identify topographic and cellular structures.
RESULTS
There was no obvious difference in the distribution and intensity of Fos-like immunoreactivity in animals sacrificed at 90 or 150 min, so no distinction will be made between the groups when presenting the results. The placebo led to only a few Fos-immunoreactive (ir) neurons throughout the central nervous system. One or two positive neurons per section were occasionally found in the cortex, basal forebrain, and other regions (see Figs. 1, 2, 3, 4 and Table 1). In addition, with all treatments including the placebo, a group of labeled neurons was detected in the anterior paraventricular nucleus of the thalamus, as were some weakly labeled cells in the inferior colliculus. None of the treatments resulted in much labeling of the main structures known to play important roles in waking and cortical activation: only a few stained cells were seen in the substantia innominata of the basal forebrain, the tuberomammillary area of the posterior hypothalamus, the mesopontine tegmentum, and the locus coeruleus after drug treatment. Few Fos-ir neurons were seen in the intralaminar nuclei and other thalamic areas (Fig. 1 and Table 1).
Figure 1.
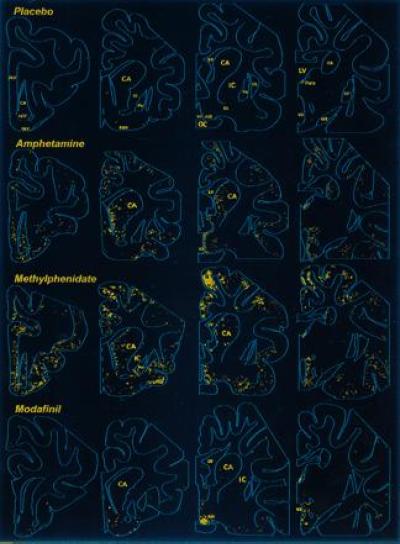
Distribution of Fos-like immunoreactivity in the rostral brain of the cat. Camera lucida drawing of frontal sections showing Fos-ir neurons following oral administration of the indicated substances Note that (i) the placebo induces little labeling, (ii) amphetamine or methylphenidate treatment causes numerous Fos-positive neurons in the cerebral cortex and striatum, and (iii) modafinil treatment induces little labeling of the cortex and striatum, but a large number of aggregated positive neurons is seen in the anterior hypothalamic nucleus (AH). ACC, nucleus accumbens; CA, caudate nucleus; CL, claustrum; DBH, diagonal band of Broca; GP, globus pallidus; IC, internal capsule; LV, lateral ventricle; OC, optic chiasma; OLT, olfactory tubercle; OT, optic tract; Para, anterior paraventricular nucleus of the thalamus; PrF, prefrontal cortex; Pu, putamen; SI, substantia innominata; V3, third ventricle.
Figure 2.
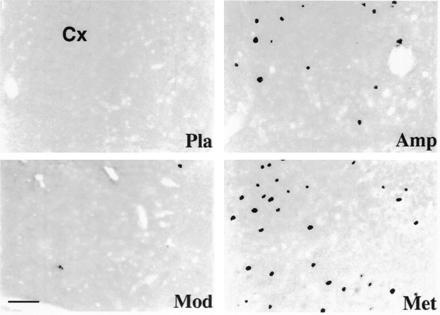
Photomicrographs of frontal sections through the mediofrontal cortex (Cx) of the cat, showing Fos-like immunoreactivity following different administrations. Note that (i) the large number of stained cells seen with methylphenidate (Met) treatment and, to a lesser degree, with amphetamine (Amp) and (ii) few or no labeled neurons seen with modafinil (Mod) or placebo (Pla). (Bar = 50 μm.)
Figure 3.
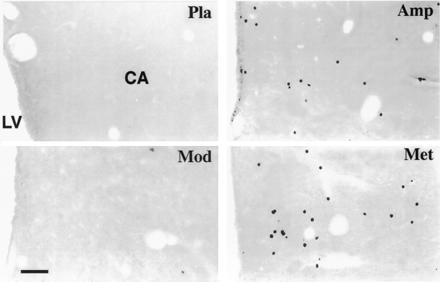
Photomicrographs of frontal sections through the caudate nucleus (CA) of the cat, showing Fos-like immunoreactivity following different administrations. Note the large number of stained cells seen with amphetamine (Amp) or methylphenidate (Met) treatment and that few or no labeled neurons are seen using modafinil (Mod) or placebo (Pla). LV, lateral ventricle. (Bar = 80 μm.)
Figure 4.
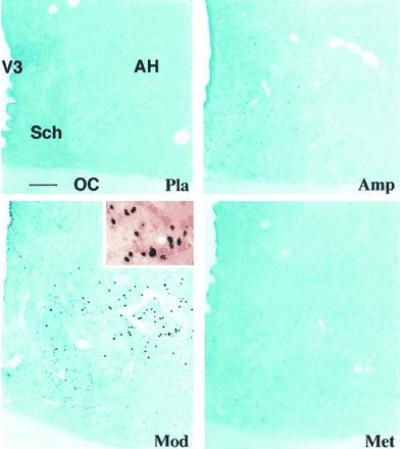
Photomicrographs (blue filter) of frontal sections through the mediocaudal part of the preoptic/anterior hypothalamus of the cat, showing Fos-like immunoreactivity following different administrations. Note that (i) no obvious labeling is seen with placebo (Pla), (ii) some weakly labeled cells are present in the periventricular zone of the third ventricle (V3) following amphetamine (Amp) or methylphenidate (Met) treatment, and (iii) modafinil (Mod) induces some weakly stained neurons in the suprachiasmatic nucleus (Sch) and many strongly labeled cells in the AH. A higher power magnification of the Inset shown in the AH appears in the upper right photomicrograph (without filter), illustrating the morphology of the Fos-ir labeling in the cell nuclei on counterstained background. OC, optic chiasma. (Bar = 50 μm.)
Table 1.
Quantitative evaluation of Fos-like labeling in selected brain areas after the various treatments in the cat
| Site | Placebo (A) | Amphetamine (B) | Methylphenidate (C) | Modafinil (D) | ANOVA | Multiple range analysis |
|---|---|---|---|---|---|---|
| Cortex | ||||||
| Mediofrontal | 0.3 ± 0.1 | 158.8 ± 26.3 | 238.2 ± 28.4 | 5.6 ± 1.5w | ∗∗∗ | AD; B; C |
| Temporal | 0.4 ± 0.1 | 81.8 ± 24.1 | 92.3 ± 7.9 | 15.5 ± 3.6w | ∗∗∗ | AD; BC |
| Amygdaloid | 0.5 ± 0.2 | 91.0 ± 16.9 | 104.7 ± 14.5 | 10.9 ± 2.0w | ∗∗∗ | AD; BC |
| Striatum | ||||||
| Caudate n. | 0.6 ± 0.2 | 105.5 ± 18.6 | 87.6 ± 6.1 | 2.4 ± 0.8 | ∗∗∗ | AD; BC |
| Accumbens | 0.3 ± 0.1 | 31.3 ± 6.5 | 27.5 ± 3.8 | 3.3 ± 0.9 | ∗∗∗ | AD; BC |
| Stria terminalis | 0.3 ± 0.1 | 14.5 ± 3.0 | 8.6 ± 1.6 | 3.0 ± 0.9 | ∗∗∗ | AD; B; C |
| Putamen | 0.3 ± 0.1 | 7.4 ± 2.5 | 5.7 ± 1.6 | 1.6 ± 0.8 | ∗∗ | AD; BC; CD |
| Globus pallidus | <2 | <2 | <2 | <2 | — | — |
| Basal forebrain | ||||||
| Septum | 0.8 ± 0.3 | 26.5 ± 3.4 | 21.5 ± 3.5 | 7.7 ± 1.3 | ∗∗∗ | AD; BC |
| Diagonal band of Broca | 0.1 ± 0.1 | 7.0 ± 1.2 | 15.6 ± 2.2 | 6.6 ± 1.4 | ∗∗∗ | A; BD; C |
| Substantia innominata | <5 | <5 | <5 | <5 | — | — |
| Thalamus | ||||||
| Anterior paraventricular n. | 15.6 ± 1.3 | 22.9 ± 1.4 | 19.2 ± 2.1 | 19.7 ± 2.1 | ∗ | ACD; BCD |
| Midline and intralaminar n. | <2 | <2 | <2 | <2 | — | — |
| Other thalamic areas | <2 | <2 | <2 | <2 | — | — |
| Hypothalamus | ||||||
| Anterior preoptic | 0.3 ± 0.1 | 31.3 ± 4.4 | 30.2 ± 2.4 | 17.9 ± 2.3 | ∗∗∗ | A; BC; D |
| Anterior hypothalamic n. | 0.3 ± 0.2 | 8.3 ± 1.1w | 6.8 ± 1.4w | 77.7 ± 7.1 | ∗∗∗ | ABC; D |
| Suprachiasmatic n. | 1.3 ± 0.5w | 8.5 ± 1.1w | 7.9 ± 1.1w | 33.0 ± 2.1w | ∗∗∗ | A; BC; D |
| Tuberomammillary area | <7 | <7 | <7 | <7 | — | — |
| Other posterior areas | 1.6 ± 0.5 | 26.3 ± 2.8 | 28.0 ± 2.9 | 18.6 ± 2.3 | ∗∗∗ | A; BC; D |
| Brainstem | ||||||
| Superior colliculus | 0.3 ± 0.1 | 24.3 ± 5.4 | 21.8 ± 2.3 | 9.0 ± 1.8 | ∗∗∗ | AD; BC |
| Rostral periacqueductal gray | 0.5 ± 0.2 | 12.3 ± 1.6 | 9.8 ± 0.9 | 25.2 ± 2.9 | ∗∗∗ | A; BC; D |
| Rostral reticular formation | <7 | <7 | <7 | <7 | — | — |
| Substantia nigra | <5 | <5 | <5 | <5 | — | — |
| Dorsal raphe | <5 | <5 | <5 | <5 | — | — |
| Mesopontine tegmentum | <5 | <5 | <5 | <5 | — | — |
| Locus coeruleus | <5 | <5 | <5 | <5 | — | — |
| Inferior colliculus | 16.2 ± 2.8w | 40.8 ± 6.8 | 52.8 ± 8.3 | 32.0 ± 3.8 | ∗∗ | AD; BC; BD |
The results are expressed as the mean number of positive cells per section ± SEM, obtained from cell counting of three sections of each treated cat (four per treatment, thus n = 12) in the representative levels of analyzed structures. The cortical zones are defined in the text. The effect of the drugs on c-fos expression was examined by ANOVA. The relationship among the experimental groups (A, B, C, D) was further evaluated by multiple range analysis with the level of significance set at P < 0.05: they are separated by a semi-colon when they are statistically different. Note that in the great majority of brain regions, the effect of modafinil on c-fos expression is clearly distinct from that of amphetamine or methylphenidate (∗, P < 0.05; ∗∗, P < 0.01; ∗∗∗, P < 0.0001; n, nucleus; w, weak intensity of labeling).
Oral administration of amphetamine at a dose of 1 mg/kg induced Fos-like immunoreactivity in a large number of cells in several cerebral regions of all treated cats, especially the cortex and striatum. Of the cortical zones, the mediofrontal cortex (lateral, splenial, and suprasplenial gyri and gyrus fornicatus) contained the largest number of labeled cells (Table 1 and Fig. 1). Although the intensity of cortical labeling differed from one neuron to another, most were strongly labeled (Fig. 2). Within the striatum, the caudate nucleus was densely labeled throughout its rostrocaudal extension, with more positive neurons being found in its medial part than its lateral part (Figs. 1 and 3). Many Fos-ir cells were also seen in the medial prefrontal cortex, olfactory tubercle, septum, accumbens, and stria terminalis, while lesser or few were found in the putamen and globus pallidus (Table 1 and Fig. 1). Some weakly stained cells were scattered throughout the hypothalamic periventricular zone (Figs. 1 and 4). In the brainstem, scattered labeled neurons were seen in the colliculi, periaqueductal gray and rostroventral pontine gray.
As shown in Fig. 1 and Table 1, methylphenidate (2.5 mg/kg) evoked widespread expression of Fos-like immunoreactivity in various cerebral structures, the distribution pattern and labeling intensity being very similar to those elicited by amphetamine. The number of Fos-ir cells in the mediofrontal cortex was, however, markedly higher than that seen with amphetamine (Table 1 and Figs. 1 and 2). In the striatum, the number and intensity of labeling were similar to that seen with amphetamine (Table 1 and Fig. 3).
In contrast to methylphenidate and amphetamine and in all treated cats, administration of modafinil (5 mg/kg) induced Fos-ir neurons in localized cerebral areas (Table 1 and Fig. 1). Thus, few labeled cells were present in the cortical sites (Fig. 2), except for the temporal (gyrus ectosylvius anterior) and amygdaloid (periamygdalae and praepyriformis) cortex, in which a scattered population of weakly labeled neurons was seen; the number (Table 1) and intensity of labeling in these two areas were, however, lower than that following amphetamine or methylphenidate treatment. In addition, very few positive neurons were present in striatal regions, such as the caudate nucleus (Fig. 3). Conversely, modafinil treatment induced dense Fos-like immunoreactivity in the AH and adjacent area (Figs. 1 and 4). As defined in the cat by different authors (16, 17, 18), the AH is a round nucleus, extending for ≈1 mm within the mediocaudal part of the preoptic/anterior hypothalamus and made up of small- to medium-sized neurons. The Fos-ir neurons in the AH were, for the most part, strongly labeled (Fig. 4). In the dorsolateral portion of the suprachiasmatic nucleus, close to the AH, a population of weakly labeled cells was also present (Table 1 and Fig. 4). A small number of Fos-ir neurons was also found in the rostral basal forebrain and other hypothalamic areas (Table 1 and Fig. 1), as were some scattered cells in the mesencephalic periaqueductal gray and the rostroventral pontine gray. In view of the small size of the AH (Fig. 1) and the large number (Table 1) and high intensity of the labeling on it (Fig. 4), we consider this nucleus and the adjacent portion of the suprachiasmatic nucleus as the major structure in which modafinil induces c-fos expression.
DISCUSSION
The main finding of the present study is that amphetamine, methylphenidate, and modafinil, which all induce wakefulness, cause two distinct patterns of c-fos distribution in the cat brain. In fact, methylphenidate and amphetamine both induce a diffuse Fos-like immunoreactivity in the cortex and striatum. In contrast, modafinil treatment results in few labeled cells in these regions, but, instead, in marked c-fos labeling of neurons of the AH. These striking differences, together with the relative short time between treatment and sacrifice (90 or 150 min), imply that the labeling seen is a reflection of the pharmacological targets of the treatments rather than the consequences of their waking effect. Furthermore, prolonged waking or sleep deprivation causes mainly c-fos expression in the preoptic area (12, 13, 19, 20), data different from those seen in the present study and thus support our hypothesis.
In spite of their powerful awakening potency at the doses used, none of the three drugs caused significant c-fos labeling of the main structures known to be important in waking (e.g., thalamus, basal forebrain, posterior hypothalamus, and mesopontine tegmentum; see refs. 21 and 22). We must point out that most of the neurons expressing c-fos, whether in this trial or certain others (12, 19), were of small to medium size as revealed by neutral red counterstaining, whereas the known waking systems consist mainly of large neurons, and that c-fos is the principal member of the IEGs, but not the only one (5, 6, 7) and so certain categories of neurons might use other IEGs for the transcriptional regulation of genes during cell activation. Although we cannot eliminate the possibility that the amount of induced Fos protein in these systems was below the threshold of the immunocytochemical technique used, these apparently negative results illustrate once again the complexity of the waking mechanisms and tend to suggest that waking may be associated with activation of structures other than those currently defined.
Amphetamine treatment induced dense c-fos expression in the striatum, especially the caudate nucleus, which receives projections from mesencephalic dopaminergic neurons. This fits with its well-known role as dopamine transmission-enhancing agent (23). In fact, numerous studies indicate that amphetamine activates the mesencephalostriatal dopaminergic systems originating from the A8–A10 groups. Moreover, recent studies using c-fos immunohistochemistry in the rat have also found that amphetamine evokes strong expression of c-fos in neurons of the caudate nucleus (24, 25, 26), an effect mediated by dopamine D1 and D2 receptors (24). In agreement with the dopamine transmission enhancing property of amphetamine, we have also observed Fos-like immunoreactivity in various cortical zones, especially the mediofrontal cortex, an essential cortical target of the mesoneocortical system originating from the dopaminergic ventral tegmental area of Tsaï (27, 28). Methylphenidate, which does not seem to have been the subject of previous studies using c-fos, evokes a c-fos distribution highly similar to that caused by amphetamine, both in the cortex and striatum, indicating, its dopamine transmission-enhancing nature as well. The fact that, at equivalent waking potency, methylphenidate induces a greater number of cortical cells expressing c-fos than does amphetamine suggests that the dopaminergic mesoneocortical system might be more sensitive to methylphenidate than to amphetamine. The notable presence of activated cells in the cortex underlines the fact that amphetamine- or methylphenidate-induced awakening would be due not only to activation of structures involved in the control of movement and behavior, such as the striatum, but also to their activating effect on the cortex, probably in a direct manner. Finally, expression of IEGs in the striatal and cortical dopaminergic targets is suggested to mediate the drug tolerance and dependency of psychostimulants (23, 29). The few Fos-ir cells seen here with modafinil might explain these addictive phenomena not being associated with its use (1).
The most distinctive and unusual effect of modafinil seen in this study is the strong c-fos expression in the AH and adjacent suprachiasmatic border. Though this nucleus has been anatomically well defined, its role in sleep-wake control or other brain functions has not yet been attentively investigated (16, 17, 18, 30). It may serve as a key target for the waking effect of modafinil due to its special anatomical disposition. Indeed, it is situated between the preoptic area, which plays an essential role in sleep generation (31, 32, 33, 34, 35), and the posterior hypothalamus, in which histaminergic neurons constitute an important component for arousal (22, 32, 34), and makes numerous reciprocal connections with both (17, 30, 36). Furthermore, it sends efferent inputs to cholinergic and gabaergic basal forebrain (17, 30, 36) involved in the control of cortical activity. The AH may therefore play a key role in integrating and coordinating the functional activity of these basalohypothalamic structures which are particularly important for sleep-wake alternation. Before considering the hypothesis of how modafinil induces wakefulness via the AH, it is necessary to take into account the paucity of cortical Fos labeling seen following modafinil treatment. This singular phenomena implies that no major cortical excitation occurs, at least not in the same manner than that following amphetamine or methylphenidate treatment, and that modafinil’s awakening property cannot be explained in terms of direct pharmacological cortical activation, as occurs with the two agents. A disinhibitory procedure seems, therefore, more plausible.
On the one hand, neurons in the preoptic area have been proposed to induce sleep by inhibiting through their descending efferents the posterior hypothalamus histaminergic arousal system (20, 31, 32, 33, 34, 35, 37). To explain the waking effect of modafinil, it might be suggested that labeled neurons in the AH seen with its use are inhibitory for preoptic sleep-generating neurons. Data supporting a such possibility are the presence in the AH and adjacent area of a large number of neurons containing inhibitory transmitters such as γ-aminobutyric acid (GABA) (37, 38) and met-enkephalin (30, 39) and the massive local efferent projections of this nucleus within the preoptic/anterior hypothalamus. This hypothesis remains to be evaluated by double labeling studies for induced c-fos and inhibitory transmitters. On the other hand, modafinil also causes a marked decrease in cortical GABA outflow in freely moving rodents (40, 41). This effect does not seem to be its direct action on intrinsic cortical gabaergic neurons, since it is not seen in vitro on cortex slices (41) and thus the integrity of the afferent pathways to the cortex appears to be necessary. Extrinsic cortical GABA originates from the gabaergic neurons of the posterior hypothalamus (42) and basal forebrain (43, 44). The forebrain ascending gabaergic neurons form basket-like contacts preferentially with cortical gabaergic interneurons and thus play a decisive role in cortical disinhibition (43, 44, 45). Direct corticopetal projections of gabaergic cells in the AH have not yet been demonstrated; thus, it remains to be determined if the effect of modafinil on cortical GABA is due, via its AH target cells and their large forebrain outputs, to an activation of the gabaergic ascending system, which, in turn, inhibits the cortical gabaergic interneurons, leading to a reduction in GABA (40) and, consequently, to cortical disinhibition and, therefore, to awakening.
The waking state resulting from either the descending or ascending disinhibitory process does not depend on direct pharmacological excitation and is, therefore, more physiological, since the cerebral cortex is maintained in activation purely by natural influxes originating from various ascending systems. In summary, our results suggest that wakefulness induced by different pharmacological agents may be associated with c-fos expression in different brain regions and that modafinil induces wakefulness by mechanisms distinct from those of amphetamine and methylphenidate. The main characteristics of modafinil revealed up to the present seem possibly to be explained by this hypothesis of disinhibition.
Acknowledgments
We thank F. C. Lafon, P. E. Sacksick, F. A. Rambert, and A. M. Frydman for their help and G. Vanni-Mercier for advice. This work was supported by Laboratoire L. Lafon (J.S.L. and Y.H.), Institut National de la Santé et de la Recherche Médicale Unité 52, Centre National de la Recherche Scientifique ERS 5645, Université Claude Bernard, Lyon, France.
Footnotes
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Abbreviations: AH, anterior hypothalamic nucleus; IEG, immediate-early gene; ir, immunoreactive.
References
- 1.Bastuji H, Jouvet M. Prog Neuropsychopharmacol Biol Psychiatry. 1988;12:695–700. doi: 10.1016/0278-5846(88)90014-0. [DOI] [PubMed] [Google Scholar]
- 2.Duteil J, Rambert F A, Pessonnier J, Hermant J F, Gombert R, Assous E. Eur J Pharmacol. 1990;180:49–58. doi: 10.1016/0014-2999(90)90591-s. [DOI] [PubMed] [Google Scholar]
- 3.Hermant J F, Rambert F A, Duteil J. Psychopharmacology. 1991;103:28–32. doi: 10.1007/BF02244069. [DOI] [PubMed] [Google Scholar]
- 4.Lin J S, Roussel B, Akaoka H, Fort P, Debilly G, Jouvet M. Brain Res. 1992;591:319–326. doi: 10.1016/0006-8993(92)91713-o. [DOI] [PubMed] [Google Scholar]
- 5.Morgan J I, Curran T. Trends Neurosci. 1989;12:459–462. doi: 10.1016/0166-2236(89)90096-9. [DOI] [PubMed] [Google Scholar]
- 6.Morgan J I, Curran T. Annu Rev Neurosci. 1991;14:421–451. doi: 10.1146/annurev.ne.14.030191.002225. [DOI] [PubMed] [Google Scholar]
- 7.Sheng M, Greenberg M E. Neuron. 1990;4:477–485. doi: 10.1016/0896-6273(90)90106-p. [DOI] [PubMed] [Google Scholar]
- 8.Hunt S P, Pini A, Evan G. Nature (London) 1989;328:632–634. doi: 10.1038/328632a0. [DOI] [PubMed] [Google Scholar]
- 9.Ceccatelli S, Villar M J, Goldstein M, Hökfelt T. Proc Natl Acad Sci USA. 1989;86:9569–9573. doi: 10.1073/pnas.86.23.9569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fritschy J M, Frondoza C G, Grzanna R. Brain Res. 1991;562:48–56. doi: 10.1016/0006-8993(91)91185-4. [DOI] [PubMed] [Google Scholar]
- 11.Young S T, Porrino L J, Iadarola M J. Proc Natl Acad Sci USA. 1991;88:1291–1295. doi: 10.1073/pnas.88.4.1291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pompeiano M, Cirelli C, Tononi G. Arch Ital Biol. 1992;130:325–335. [PubMed] [Google Scholar]
- 13.Pompeiano M, Cirelli C, Tononi G. J Sleep Res. 1994;3:80–96. doi: 10.1111/j.1365-2869.1994.tb00111.x. [DOI] [PubMed] [Google Scholar]
- 14.Shiromani P J, Kilduff T S, Bloom F E, McCarley R W. Brain Res. 1992;580:351–357. doi: 10.1016/0006-8993(92)90968-f. [DOI] [PubMed] [Google Scholar]
- 15.Yamuy J, Mancillas J R, Morales F R, Chase M H. J Neurosci. 1993;13:2703–2718. doi: 10.1523/JNEUROSCI.13-06-02703.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bleier R. The Hypothalamus of the Cat: A Cytoarchitectonic Atlas in the Horsley-Clarke Co-ordinate System. Baltimore: Johns Hopkins Univ. Press; 1961. [Google Scholar]
- 17.Saper C B, Swanson L W, Cowan W M. J Comp Neurol. 1978;182:575–600. doi: 10.1002/cne.901820402. [DOI] [PubMed] [Google Scholar]
- 18.Berman A L, Jones E G. The Thalamus and Basal Telencephalon of the Cat: A Cytoarchitectonic Atlas With Stereotaxic Coordinates. Madison: Univ. of Wisconsin Press; 1982. [Google Scholar]
- 19.Ledoux L, Sastre J P, Buda C, Luppi P H, Jouvet M. Brain Res. 1996;735:108–118. doi: 10.1016/0006-8993(96)00599-9. [DOI] [PubMed] [Google Scholar]
- 20.Sherin J E, Shiromani P J, McCarley R W, Saper C B. Science. 1996;271:216–219. doi: 10.1126/science.271.5246.216. [DOI] [PubMed] [Google Scholar]
- 21.Jones B E. In: Principles and Practice of Sleep Medicine. Kryger M H, Roth T, Dement W C, editors. Philadelphia: Saunders; 1989. pp. 121–138. [Google Scholar]
- 22.Lin J S, Hou Y, Sakai K, Jouvet M. J Neurosci. 1996;16:1523–1537. doi: 10.1523/JNEUROSCI.16-04-01523.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hyman S E. Neuron. 1996;16:901–904. doi: 10.1016/s0896-6273(00)80111-7. [DOI] [PubMed] [Google Scholar]
- 24.Graybiel A M, Moratalla R, Robertson H A. Proc Natl Acad Sci USA. 1990;87:6912–6916. doi: 10.1073/pnas.87.17.6912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jonansson B, Lindström K, Fredholm B B. Neuroscience. 1994;59:837–849. doi: 10.1016/0306-4522(94)90288-7. [DOI] [PubMed] [Google Scholar]
- 26.Wang J Q, Smith A J W, McGinty J F. Neuroscience. 1995;68:83–95. doi: 10.1016/0306-4522(95)00100-w. [DOI] [PubMed] [Google Scholar]
- 27.Swanson L W. Brain Res Bull. 1982;9:321–353. doi: 10.1016/0361-9230(82)90145-9. [DOI] [PubMed] [Google Scholar]
- 28.Björklund A, Lindvall O. In: Handbook of Chemical Neuroanatomy. Björklund A, Hökfelt T, editors. Amsterdam: Elsevier; 1984. pp. 55–121. [Google Scholar]
- 29.Hughes P, Dragunow M. Pharmacol Rev. 1995;47:133–178. [PubMed] [Google Scholar]
- 30.Risold P Y, Canteras N S, Swanson L W. J Comp Neurol. 1994;348:1–40. doi: 10.1002/cne.903480102. [DOI] [PubMed] [Google Scholar]
- 31.Nauta W J H. J Neurophysiol. 1946;9:285–361. doi: 10.1152/jn.1946.9.4.285. [DOI] [PubMed] [Google Scholar]
- 32.Lin J S, Sakai K, Vanni-Mercier G, Jouvet M. Brain Res. 1989;479:225–240. doi: 10.1016/0006-8993(89)91623-5. [DOI] [PubMed] [Google Scholar]
- 33.Sallanon M, Denoyer M, Kitahama K, Aubert C, Gay N, Jouvet M. Neuroscience. 1989;32:669–683. doi: 10.1016/0306-4522(89)90289-3. [DOI] [PubMed] [Google Scholar]
- 34.Lin J S, Sakai K, Jouvet M. Eur J Neurosci. 1994;6:618–625. doi: 10.1111/j.1460-9568.1994.tb00306.x. [DOI] [PubMed] [Google Scholar]
- 35.Krilowiicz B L, Szymusiak R, McGinty D. Brain Res. 1995;668:30–38. doi: 10.1016/0006-8993(94)90507-x. [DOI] [PubMed] [Google Scholar]
- 36.Morgane J P, Ranksepp J. Handbook of Hypothalamus. Vol. 1. New York: Dekker; 1979. [Google Scholar]
- 37.Gritti I, Mainville L, Jones B E. J Comp Neurol. 1994;339:251–268. doi: 10.1002/cne.903390206. [DOI] [PubMed] [Google Scholar]
- 38.Kitahama K, Sallanon M, Okamura H, Jeffard M, Jouvet M. CR Acad Sci Ser 3. 1989;308:507–511. [PubMed] [Google Scholar]
- 39.Harlan R E, Shivers B D, Romano G J, Howlls R D, Pfaff D W. J Comp Neurol. 1987;258:159–184. doi: 10.1002/cne.902580202. [DOI] [PubMed] [Google Scholar]
- 40.Tanganelli S, Fuxe K, Ferraro L, Janson A M, Bianchi C. Naunyn-Schmiedeberg’s Arch Pharmacol. 1992;345:461–465. doi: 10.1007/BF00176625. [DOI] [PubMed] [Google Scholar]
- 41.Tanganelli S, Mora M P, Ferraro L, Méndez-Franco J, Beani L, Rambert F, Fuxe K. Eur J Pharmacol. 1995;273:63–71. doi: 10.1016/0014-2999(94)00675-w. [DOI] [PubMed] [Google Scholar]
- 42.Vincent S R, Hökfelt T, Skirboll L R, Wu J Y. Science. 1983;220:1309–1311. doi: 10.1126/science.6857253. [DOI] [PubMed] [Google Scholar]
- 43.Freund T F, Antal M. Nature (London) 1988;361:170–173. doi: 10.1038/336170a0. [DOI] [PubMed] [Google Scholar]
- 44.Freund T F, Meskenaite V. Proc Natl Acad Sci USA. 1992;89:738–742. doi: 10.1073/pnas.89.2.738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Smythe J W, Colom L V, Bland B H. Neurosci Biobehav Rev. 1992;16:289–308. doi: 10.1016/s0149-7634(05)80203-9. [DOI] [PubMed] [Google Scholar]