

Abstract
A central theoretical goal of epidemiology is the construction of spatial models of disease prevalence and risk, including maps for the potential spread of infectious disease. We provide three continent-wide maps representing the relative risk of malaria in Africa based on ecological niche models of vector species and risk analysis at a spatial resolution of 1 arc-minute (9 185 275 cells of approximately 4 sq km). Using a maximum entropy method we construct niche models for 10 malaria vector species based on species occurrence records since 1980, 19 climatic variables, altitude, and land cover data (in 14 classes). For seven vectors (Anopheles coustani, A. funestus, A. melas, A. merus, A. moucheti, A. nili, and A. paludis) these are the first published niche models. We predict that Central Africa has poor habitat for both A. arabiensis and A. gambiae, and that A. quadriannulatus and A. arabiensis have restricted habitats in Southern Africa as claimed by field experts in criticism of previous models. The results of the niche models are incorporated into three relative risk models which assume different ecological interactions between vector species. The “additive” model assumes no interaction; the “minimax” model assumes maximum relative risk due to any vector in a cell; and the “competitive exclusion” model assumes the relative risk that arises from the most suitable vector for a cell. All models include variable anthrophilicity of vectors and spatial variation in human population density. Relative risk maps are produced from these models. All models predict that human population density is the critical factor determining malaria risk. Our method of constructing relative risk maps is equally general. We discuss the limits of the relative risk maps reported here, and the additional data that are required for their improvement. The protocol developed here can be used for any other vector-borne disease.
Introduction
A central theoretical goal of epidemiology is the construction of spatial models of disease prevalence and risk, including maps for the potential spread of infectious disease [1], [2]. In particular, Snow et al. [3] have emphasized the need for risk maps for malaria in Africa which accounts for an estimated 85% of the 1 million annual deaths due to this disease [4]. Risk maps can be used to identify appropriate strategies of response to disease outbreaks including targeted vaccination [5] and vector, reservoir, or agent control [6]. Risk maps have been constructed using a variety of techniques including reports of disease cases [7] and distributions of disease agents, reservoirs, or vectors, based on surveys and expert opinion [8]. In recent years, these methods have been extended to use ecological models to predict the potential spatial spread of disease [9]. The use of ecological niche models for quantitative prediction of geographical distributions of agent, reservoir, and vector species has been advocated to augment traditional mapping methods such as splining and kriging [9]. Here we report a systematic attempt to construct niche models for all vectors of malaria in Africa for which data are available.
Niche models predict the “fundamental niche” of a species which identifies the region in ecological space that the species would occupy were its movement unrestricted [9], [10]. Within biodiversity studies, in which the concern is typically with the conservation of the extant habitat of a species, this fundamental niche (representing a potential distribution) is typically restricted to a realized geographical niche by using additional information such as confirmed occurrence records for a species within each accepted contiguous piece of modelled suitable habitat [10]. Such a choice is conservative in the sense that a species may occur outside its predicted realized niche. In typical contexts of biodiversity conservation planning such conservatism is appropriate; it ensures that areas selected for conservation management are maximally likely to contain the species predicted for them. However, in epidemiological contexts, especially if the interest is in identifying risk, the geographical extent of the fundamental niche is more relevant as this range defines the areas to which agent, reservoir, and vector species may potentially spread because of ecological suitability. (This means that, in epidemiological contexts, the fundamental niche or potential distribution should not be clipped to a smaller realized niche in most cases.) In this analysis, fundamental niches as predicted by niche models are used as the basis for the construction of relative risk maps.
We construct niche models for 10 Anopheles species recognized as vectors of malaria in Africa using a maximum entropy method based on known species' occurrences and environmental layers. These niche models predict geographic distributions of the species. For seven of these species the results presented here appear to be the first niche models reported in the literature. We used the Maxent software package [11] for the maximum entropy modelling. Within biodiversity studies, in which niche modelling is a standard technique [12]–[16] this maximum entropy method has emerged as one of the three most reliable techniques for predicting species' distributions [11], [17]. The other two most reliable methods are genetic algorithms (GARP [18]) and regression trees [19]. The advantage of Maxent over GARP is that it is much faster and allows for the simultaneous modelling of an indefinite number of species. The advantage of maximum entropy methods over both genetic algorithms and regression methods is that Maxent predicts relative probabilities of occurrence rather than the simple presence or absence of a species. This permits a finer (more nuanced) risk assessment than what can be achieved from presence-absence predictions alone. Our analysis appears to be the first use of Maxent in an epidemiological context.
Using the niche models, spatial information on human population densities, and the human blood index (HBI) values of Anopheles species for which these values were available, we construct preliminary relative risk maps for malaria in Africa. These maps report the relative risk of malaria occurrence at different geographical locations. We discuss in detail what data are necessary to make such maps more accurate and how our methods can be generalized to other vector-borne diseases.
There have been five recent vector-based attempts to construct risk maps for malaria. Kiszewski et al. [8] produced a global risk map based on the stability of malaria parasite transmission. They divided the world into 260 malarious regions and then identified the dominant vector in each region. The dominant vector was defined as the most abundant Anopheles species in the region that was a malaria vector, contained sporozoites frequently, and fed predominantly on humans. The stability index of a region was defined using the human feeding rate, daily survival rate, and length of the extrinsic incubation period of the dominant vector. The effects of temperature and precipitation on these parameters were quantified and environmental data were used to produce a worldwide projection of malaria transmission.
Lindsay et al. [20] used nonlinear regression to relate known occurrences of A. arabiensis and A. gambiae to environmental parameters. The regression was used to predict the relative proportion of the two species throughout Africa. Kuhn et al. [21] based their map on a database of occurrence records of six Anopheles species in Europe. Statistical analyses correlated these occurrence points with environmental parameters. The correlations were used to predict the distribution of the Anopheles species across Europe. Rogers et al. [22] used a maximum likelihood analysis to identify the optimal environmental parameters to predict the occurrence of five Anopheles species in Africa. Levine et al. [23], [24] used niche models to map malaria risk in Africa, Central America, and the United States. Using species' occurrence and environmental data in a genetic algorithm (in the GARP software package), they predicted the distribution of three Anopheles species in Africa and five species in the United States.
In addition to these vector-based approaches, risk maps have also been constructed by mapping the distribution of the malaria parasite. Kleinschmidt et al. [25], [26] used regression analysis to determine the relationship between the malaria parasite prevalence of an area and environmental variables. The regression model was refined using kriging for spatial interpolation thus producing a map of malaria prevalence. Diggle et al. [27] used a generalized linear mixed model to determine the relationship between malaria presence in children and their age and bed net use along with available medical services and land cover. Gemperli et al. [7] developed a Bayesian model to calculate parasite transmission intensity on the basis of malaria survey information and land cover, temperature, and rainfall data.
We map malaria risk using distributions of malaria vectors rather than malaria parasites. The underlying assumption is that malaria vector abundance is an adequate surrogate for malaria risk without explicit inclusion of parasite abundance. However, even if this assumption is invalid, risk maps based on vector distributions are still useful to augment risk analyses based only on parasite distributions. (In future work, we plan to construct risk maps using more sophisticated models of transmission that include interactions between parasites, vectors, and variable human susceptibility [28]–[30].)
This analysis differs from and extends previous efforts in five ways. First, our niche models are constructed using a maximum entropy method. Second, we model a larger number of species than previously attempted. Third, we include human population distribution data in the risk analysis and associated relative risk models. Fourth, we perform a sensitivity analysis to evaluate the robustness of our results. Fifth, we show how additional data can lead to a more sophisticated risk analysis for malaria and other vector-borne diseases.
Methods
Data
A 1 arc-minute (0.01666°×0.01666° longitude×latitude, approximately 4 km2 at the equator) grid was used to divide Africa into 9 185 275 cells. Twenty environmental layers were obtained from the WorldClim database [31]. Each layer was available at a resolution of 30 arc-seconds (0.008333°×0.008333°) and was resampled at a resolution of 1 arc-minute. These layers are listed in Table 1. (All spatial data manipulation used ArcMap GIS [32].)
Table 1. Environmental Parameters Used in Niche Modelling.
| Parameter |
| Annual Mean Temperature |
| Mean Diurnal Range |
| Isothermality |
| Temperature Seasonality |
| Maximum Temperature of Warmest Month |
| Minimum Temperature of Coldest Month |
| Temperature Annual Range |
| Mean Temperature of Wettest Quarter |
| Mean Temperature of Driest Quarter |
| Mean Temperature of Warmest Quarter |
| Mean Temperature of Coldest Quarter |
| Annual Precipitation |
| Precipitation of Wettest Month |
| Precipitation of Driest Month |
| Precipitation Seasonality |
| Precipitation of Wettest Quarter |
| Precipitation of Driest Quarter |
| Precipitation of Warmest Quarter |
| Precipitation of Coldest Quarter |
| Altitude |
| Land Cover |
Land cover data were obtained from the Global Land Cover Facility [33]. AVHRR satellite data acquired between 1981 and 1994 were used to derive 14 classes of land cover: water; evergreen needleleaf forest; evergreen broadleaf forest; deciduous needleleaf forest; deciduous broadleaf forest; mixed forest; woodland; wooded grassland; closed shrubland; open shrubland; grassland; cropland; bare ground; urban and built. The data were initially available at a resolution of 1 km2 and resampled at a resolution of 1 arc-minute.
A review of the global distribution of malaria vectors [34] was used to identify those African Anopheles species that are capable of transmitting malaria. The 29 species used for this analysis are listed in Table 2. An extensive literature search was performed to obtain records of vector occurrences. Besides using the Mapping Malaria Risk in Africa (MARA) database [3] both PubMed and Google Scholar were searched using “Africa” and “distribution” in conjunction with the names of each of the Anopheles species listed in Table 2. References from those papers so identified were also searched. This resulted in a data set of 3 342 records of 22 malaria vectors, with 2634 of the records drawn from the MARA database.
Table 2. Occurrence Data used in Niche Modelling.
Included in column (i) are each of the 29 Anopheles species responsible for the spread of malaria in Africa. Column (ii) contains the number of records drawn from the MARA database for each species. Column (iii) contains the references from which the MARA data were obtained. Column (iv) contains the number of records drawn from sources not included in the MARA database. Column (v) contains the references from which these additional records were obtained.
Each record was georeferenced to the nearest arc minute and assigned to a corresponding cell. For those cells containing more than one record of a given species only the most recent record was kept. So as to increase the likelihood that current values of the selected environmental parameters represent the environment as it existed when the records were obtained, only those records reporting observations made after 1980 were included. The resulting data set consisted of 977 records of 22 Anopheles species with 367 of these records drawn from the MARA database. Table 2 provides a summary of these records.
Human population density data for the year 2000 were obtained from the Gridded Population of the World database [35]. These data were provided at a resolution of 2.5 arc-minutes (0.041666°×0.041666°) and resampled at a resolution of 1 arc-minute. Figure 1 shows a map of normalized population densities.
Figure 1. Population density in Africa.

The population densities have been normalized so as to range over the unit interval.
HBI values were obtained from the literature. Both PubMed and Google Scholar were searched using the terms “Anopheles” and “human blood index” with each of the species names. References from those papers so identified were also searched. Of the 10 species for which we were able to construct reliable niche models (see below), HBI values were available for nine species, with no value available for A. paludis.
Niche Models
Previous niche models of Anopheles species have used genetic algorithms, as implemented in GARP [23], [24]. In this analysis we use instead a maximum entropy technique implemented in the Maxent software package [11]. Maxent was used instead of GARP because Maxent provides relative probabilities of presence rather than only presence-absence output and because it has, in some recent studies, been shown to outperform GARP with respect to its predictive success [11], [17]. However, there are some preliminary data suggesting that GARP may perform better than Maxent at extrapolating from occurrence data (A. Townsend Peterson, personal communication). This means that models constructed using Maxent may be excessively conservative thus predicting false absences more often than GARP. If this is true then while the areas of high relative risk identified in our maps are probably reliable, those with low relative risk should not be entirely discounted. Regression tree methods (for instance those implemented in the RandomForest software package [19]) have so far not been used in an epidemiological context.
Given a set of records of species occurrences and values of selected environmental variables defined over a chosen geographical region, Maxent predicts the distribution of species in that space by finding the distribution of maximum entropy subject to the constraint that the expectation of the distribution of each species matches its observed average over the sample locations [11]. The distribution thus produced is a relative probability distribution for each species over all cells. If each relative probability in a cell is divided by the maximum such probability across the landscape, then the result is a normalized relative probability distribution which assumes that the species is certainly present in the cell in which it had its highest predicted relative probability. This analysis used this normalization.
Computer memory limitations prevented Maxent runs using as input all 9 185 175 cells simultaneously. Instead, Maxent was run using 100 sets of 10 000 cells drawn randomly from the complete set. Sets of 10 000 cells were used because Maxent performance does not significantly improve when more than 10 000 cells are used (S. Phillips, personal communication), while 100 sets were selected to sample widely from the complete set of cells. The 100 different niche models produced for each species were averaged and subsequently normalized as described above. As we were concerned with the geographical range of the fundamental niche of the vector species, these output maps were not further refined.
The accuracy of the niche models thus produced was evaluated by constructing the models using only 75% of the records with the remaining 25% set aside for testing. The accuracy of each model was then determined by performing both a threshold-dependent binomial test of omission and a threshold-independent receiver operating characteristic analysis with those cells set aside during model development [11].
In the threshold-dependent binomial test of omission a threshold of 0.10 was used to classify each vector as either present or absent in each cell, with a vector present in a given cell if the niche model assigned it a value greater than 0.10 in that cell and absent otherwise. This threshold transformed the continuous data produced by the niche models into binary data to allow a binomial test to be performed. For each vector the number of cells in which the vector was predicted to be present was compared to the number of cells known to contain the vector. A one tailed binomial test was used to determine whether the niche model outperformed a random model predicting the vector to be present in the same number of cells [11]. Maxent provides test statistics for binomial tests for 10 different threshold values. The value of 0.10 used in this analysis was arbitrarily selected from this set of possible threshold values.
In the threshold-independent receiver operating characteristic analysis, the sensitivity and specificity of the niche models were calculated at all possible thresholds. The sensitivity of a model at a threshold was defined as the percentage of species occurrences that were correctly predicted by the model at the threshold, while the specificity of the model at a threshold was defined as the percentage of correctly predicted species absences at the threshold [11]. By calculating the sensitivity and specificity of a model at all possible thresholds a receiver operating characteristic curve was produced with sensitivity plotted on the y-axis and (1–specificity) plotted on the x-axis. The area under the curve (AUC) of the resulting plot provides a measure of model performance independent of the choice of any particular threshold. An optimal model, one that predicted each occurrence of a species and for which each prediction was accurate, would have an AUC of 1.0 while a model that predicted species occurrences at random would have an AUC of 0.5.
These two tests were used to restrict attention to those models that performed significantly better than random. Only those niche models possessing both a p value less than 0.05 for the binomial test of omission and an AUC greater than 0.75 were used in this analysis. The same protocol for model retention has previously been used by Pawar et al. [16].
Risk Models
Three different relative risk models were constructed in which a value between 0 and 1 was assigned to each cell representing the relative risk of malaria posed to the human population residing within it. These models only used the nine species for which an HBI value was available. The models only incorporate the risk from ecological and demographic factors and ignore the modulation of risk through human intervention such as measures to control the spread of parasites or vectors. They also assume that parasites are present at sufficient densities to be capable of spreading whenever vectors are present. These assumptions are generally appropriate for Africa given the continued prevalence of malaria within it. However they would not be appropriate for regions such as northern Australia from which malaria parasites have been eliminated though malaria vectors remain
Let alik be the relative abundance of the l-th vector for the i-th parasite in cell k (relative to other cells in the landscape). Let pk be the human population of cell k. Let hlik be the HBI of the l-th vector for the i-th parasite in cell k. Let elik be the transmission efficiency of the l-th vector for the i-th parasite in cell k, measured by the relative likelihood of parasites being transferred to the human agent with each bite. We use a simple multiplicative model for the relative risk, φlik, due to the l-th vector for the i-th parasite in cell k:
| (1) |
Constraints on available data force further simplification of the model: (i) differences between the two primary parasites for malaria in Africa (Plasmodium vivax and P. falciparum) were ignored; (ii) the HBI was interpreted as an intrinsic property of the vector species that does not vary over geographical space; and (iii) differences in transmission efficiency between vectors were ignored. This results in the simplified relative risk model:
| (2) |
This equation expresses the relative number of individuals in cell k to whom the malaria parasite is expected to be transmitted by the l-th vector. Finally, a linear correlation is assumed between the expected relative abundance of a vector and its relative probability of presence as predicted by its niche model. This assumption must be tested in the field. Because of these four assumptions, the relative risk model presented here must be regarded as very preliminary and treated with caution.
To construct a relative risk map from this model, the relative risk from the different vectors must be compounded for each cell in the landscape. Three different models for compounding relative risk were used for this purpose:
(i) Additive model
The relative risk due to the different vectors were added together and then normalized on a scale of 0 to 1. Let Φu be the relative risk of cell k. Then:
| (3) |
This model thus assumes that there is no interaction between vector species, that each vector is able to inhabit the full extent of its fundamental niche and that there is no competition between vectors for human blood meals. If there is no such interaction then it should be possible to add the relative risks posed by the vectors in arriving at the overall relative risk posed to a given cell. However, the presumed lack of interaction between vectors has been questioned [36] and probably does not hold for all vector species. Moreover, given the methodology of this paper, there is an additional problem with the additive model. The relative probabilities provided by Maxent sum to 1 over the landscape. Thus, even if the relative abundances of a vector in the cells are linearly correlated with these probabilities, there is no way to estimate the absolute abundance. Adding the relative risk values makes the assumption that the highest absolute abundance of a vector among the cells of the landscape is the same for each vector. This assumption is ecologically suspect. Consequently, the next two models for compounding relative risk are more plausible than the additive model.
(ii) Minimax model
The relative risk of each cell was defined as the maximum relative risk from any one of the vectors. Thus:
| (4) |
This is being called a minimax model since, ultimately, in epidemiology, the goal is to minimize the risk of disease while what is being used as a relative risk measure is the maximum risk associated with all the vectors in a given cell.
(iii) Competitive exclusion model
The relative risk associated with the vector of highest relative abundance is identified with the relative risk of the cell. Let 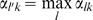 . Then:
. Then:
| (5) |
Since the relative abundances were identified with relative probabilities of occurrence, this model assumes that the vector that has the highest predicted probability of occurrence will displace all others. This assumption is similar to that about a dominant vector incorporated in the risk maps of Kiszewski et al. [8]. However, as studies of malaria transmission have identified the presence of more than one significant malaria vector within some regions [37], [38] there is again reason to doubt the validity of this assumption in many contexts.
Sensitivity Analysis
A sensitivity analysis was performed to determine the robustness of our results to variation in the values of the parameters included in the models. For each of the 100 niche models produced for each of the Anopheles species, 10 sets of HBI values were obtained by drawing randomly from a uniform distribution over the interval defined by the minimum and maximum measured HBI values for each species. For each of the three models, 1 000 relative risk values were produced for each cell using the 100 niche models, the random HBI values, and the actual human population densities. The standard deviation of the relative risk values of each cell was calculated and used to identify areas of high sensitivity.
Results
The need to set aside 25% of the records of each Anopheles species for testing purposes resulted in the production of niche models for only those 17 (out of 29) species for which four or more records were available. Data representing the accuracy of the niche models are provided in Table 3. Of these 17 niche models all but one possessed an AUC greater than 0.75. However, only 10 of the niche models possessed a p value less than 0.05 at a threshold of 0.10. As would be expected, the p values of the niche models were closely correlated with the number of records upon which the models were based. Niche models with an AUC greater than 0.75 and a p value less than 0.05 were produced for A. arabiensis, A. coustani, A. funestus, A. gambiae, A. melas, A. merus, A. moucheti, A. nili, and A. quadriannulatus. Figure 2 presents maps of the predicted geographical distributions of these species. Darker regions are those of greater relative probability of occurrence while lighter areas are those in which the relative probability of occurrence is small.
Table 3. Accuracy of the Niche Models.
| Species | AUC | Omission Rate | p Value |
| A. arabiensis | 0.909 | 0.0722 | <1.0E-6 |
| A. coustani | 0.952 | 0.000 | 6.22E-3 |
| A. funestus | 0.948 | 0.0529 | <1.0E-6 |
| A. gambiae | 0.914 | 0.0782 | <1.0E-6 |
| A. hancocki | 0.987 | 0.000 | 0.145 |
| A. marshallii | 0.856 | 0.000 | 0.226 |
| A. melas | 0.993 | 0.00715 | <1.0E-6 |
| A. merus | 0.988 | 0.0114 | <1.0E-6 |
| A. moucheti | 0.993 | 0.000 | 3.86E-3 |
| A. nili | 0.979 | 0.000 | 2.19E-3 |
| A. paludis | 0.977 | 0.000 | 0.0157 |
| A. pharoensis | 0.869 | 0.245 | 0.145 |
| A. pretoriensis | 0.656 | 0.000 | 0.508 |
| A. quadriannulatus | 0.941 | 0.154 | <1.0E-6 |
| A. rufipes | 0.853 | 0.000 | 0.0924 |
| A. squamosus | 0.920 | 0.000 | 0.641 |
| A. ziemanni | 0.879 | 0.333 | 0.170 |
100 niche models were produced for each of the species listed in column (i). Column (ii) lists the average area under the curve of each model. Column (iii) lists the average omission rate of each model. Column (iv) lists the average p value of each model.
Figure 2. The distributions of 10 malaria vectors in Africa.

Distributions are provided for: (a) A. arabiensis; (b) A. coustani; (c) A. funestus; (d) A. gambiae; (e) A. melas; (f) A. merus; (g) A. moucheti; (h) A. nili; (i) A. paludis; (j) A. quadriannulatus.
Two different tests were performed to determine the contributions of each of the environmental parameters to the niche models. In the first test, the AUC of each niche model was calculated using each of the environmental parameters individually. Those parameters that resulted in the highest AUC were interpreted as those which possessed the most information regarding the niche of a species. In the second test, the AUC of each niche model was calculated after omitting each of the environmental parameters one at a time. The effects of omitting each parameter were determined by comparing the resulting AUC to the actual AUC of the model. Those parameters for which the difference between these values was highest were interpreted as possessing the most information not present in the other environmental parameters.
The results of these two tests are provided in Table 4. As can be seen, there was no clear pattern in the contributions of the environmental parameters. The niches of different species appeared to be determined by different parameters. Over the entire data set of ten species, none of the parameters appeared to be significantly more important than any other parameter.
Table 4. Contributions of the Environmental Parameters.
| Species | Parameters Producing the Largest AUC When Included Separately | Parameters Producing the Smallest AUC when Omitted | ||||
| A. arabiensis | Temperature Seasonality, 0.772 (0.008) | Mean Temperature of Wettest Quarter, 0.760 (0.007) | Annual Precipitation, 0.746 (0.007) | Precipitation of Warmest Quarter, 0.890 (0.006) | Altitude, 0.893 (0.006) | Precipitation of Wettest Month, 0.894 (0.006) |
| A. coustani | Temperature Seasonality, 0.902 (0.007) | Temperature Annual Range, 0.825 (0.008) | Precipitation of Wettest Quarter, 0.796 (0.008) | Precipitation of Coldest Quarter, 0.892 (0.012) | Altitude, 0.905 (0.008) | Temperature Seasonality, 0.927 (0.008) |
| A. funestus | Precipitation of Wettest Month, 0.838 (0.003) | Temperature Seasonality, 0.831 (0.003) | Temperature Annual Range, 0.830 (0.003) | Precipitation of Wettest Month, 0.926 (0.004) | Minimum Temperature of Coldest Month, 0.935 (0.004) | Precipitation of Warmest Quarter, 0.938 (0.003) |
| A. gambiae | Mean Temperature of Coldest Quarter, 0.794 (0.005) | Minimum Temperature of Coldest Month, 0.780 (0.013) | Precipitation of Wettest Month, 0.838 (0.003) | Altitude, 0.891 (0.006) | Precipitation of Warmest Quarter, 0.898 (0.004) | Annual Precipitation, 0.901 (0.005) |
| A. melas | Altitude, 0.961 (0.020) | Mean Temperature of Wettest Quarter, 0.937 (0.020) | Precipitation of Wettest Month, 0.911 (0.007) | Precipitation of Coldest Quarter, 0.987 (0.005) | Precipitation of Warmest Quarter, 0.989 (0.006) | Landscape, 0.990 (0.003) |
| A. merus | Precipitation of Driest Month, 0.922 (0.012) | Precipitation of Coldest Quarter, 0.912 (0.029) | Altitude, 0.884 (0.012) | Precipitation of Warmest Quarter, 0.982 (0.005) | Altitude, 0.982 (0.004) | Mean Temperature of Driest Quarter, 0.984 (0.008) |
| A. moucheti | Temperature Annual Range, 0.980 (0.016) | Mean Diurnal Range, 0.965 (0.036) | Isothermality, 0.965 (0.036) | Landscape, 0.985 (0.003) | Precipitation of Coldest Quarter, 0.990 (0.005) | Mean Temperature of Driest Quarter, 0.991 (0.009) |
| A. nili | Temperature Annual Range, 0.982 (0.016) | Mean Diurnal Range, 0.966 (0.036) | Isothermality, 0.966 (0.036) | Mean Temperature of Wettest Quarter, 0.9623 (0.004) | Precipitation of Coldest Quarter, 0.968 (0.004) | Min Temperature of Coldest Month, 0.973 (0.003) |
| A. paludis | Temperature Annual Range, 0.984 (0.017) | Mean Diurnal Range, 0.968 (0.036) | Isothermality, 0.968 (0.036) | Temperature Annual Range, 0.970 (0.004) | Precipitation of Coldest Quarter, 0.974 (0.004) | Precipitation of Driest Quarter 0.976 (0.004) |
| A. quadriannulatus | Precipitation of Warmest Quarter, 0.874 (0.003) | Precipitation of Wettest Quarter, 0.863 (0.002) | Mean Temperature of Driest Quarter, 0.856 (0.003) | Mean Temperature of Driest Quarter, 0.913 (0.007) | Precipitation of Warmest Quarter, 0.932 (0.006) | Mean Temperature of Coldest Quarter, 0.934 (0.007) |
Column (i) lists the 10 Anopheles species for which niche models were constructed. Columns (ii–iv) list the three parameters that produced the largest AUC when taken individually. These parameters are listed in decreasing order from (ii) to (iv) on the basis of their associated AUC values. Thus column (ii) lists the environmental parameter that possesses the most information regarding the niche of each species. Columns (v–vii) list the three parameters that produced the smallest AUC when omitted. These parameters are listed in increasing order from (ii) to (iv) on the basis of their associated AUC values. Thus column (v) lists the environmental parameter that possesses the most information not possessed by the other parameters regarding the niche of each species. Average AUC values are provided next to each environmental parameter with the standard deviation of the values provided in parenthesis.
Data representing the HBI values of the Anopheles species are provided in Table 5. HBI values were available for 9 of the 10 species for which niche models were produced. An HBI value for A. plaudis could not be found in the literature. The HBI values of the remaining species were observed to vary significantly, with mean values ranging between 0.011 for A. quadriannulatus and 1.00 for A. merus. The values averaged to obtain the mean HBI value were likewise found to vary for different species, with standard deviations as high as 0.269 for A. melas and 0.241 for A. arabiensis.
Table 5. Human Blood Index Values.
| Species | Mean | Standard Deviation | References |
| A. arabiensis | 0.526 | 0.241 | [67], [71], [85], [101], [103], [108], [111], [181]–[194] |
| A. coustani | 0.157 | 0.019 | [52], [101] |
| A. funestus | 0.844 | 0.191 | [67], [111], [129], [146], [184], [185], [191], [195]–[199] |
| A. gambiae | 0.815 | 0.159 | [54], [85], [103], [108], [129], [155], [181], [184], [185], [188]–[193], [200] |
| A. melas | 0.576 | 0.269 | [54], [103], [181] |
| A. merus | 1.00 | - | [191] |
| A. moucheti | 0.931 | 0.080 | [103], [129], [184] |
| A. nili | 0.949 | 0.055 | [52], [129], [184] |
| A. paludis | - | - | - |
| A. quadriannulatus | 0.011 | - | [183] |
A list of the species is included in column (i). Columns (ii) and (iii) list the mean and standard deviation of the HBI values for each species. A list of the references from which the HBI values were drawn is provided in column (iv).
Figure 3 depicts the relative risk maps constructed using the additive, minimax, and competitive exclusion models. Cells in each map were assigned a value between 0 and 1, with 0 representing no risk and 1 representing maximal relative risk. The resulting relative risk values were found to congregate closely to either 0 or 1. To ease the visual discernment of the relative risk faced in different regions, the maps in Figure 3 plot the natural logarithm of the relative risk values.
Figure 3. The distribution of malaria relative risk in Africa.

Three different types of risk were calculated as follows: (a) the probability of occurrence of each vector in each cell was multiplied by both the human population density of the cell and the HBI of the vector. The relative risk of malaria in the cell was calculated as the sum of these values; (b) the vector possessing the maximum probability of occurrence was identified for each cell. Its probability of occurrence was multiplied by its HBI and the human population density of the cell. The relative risk of malaria in the cell was calculated as the product of these three values; and (c) the probability of occurrence of each vector in each cell was multiplied by the human population density of the cell and the HBI of the vector. The relative risk of malaria in the cell was calculated as the maximum of these values. The maps plot the natural logarithm of the relative risk.
In general the relative risk maps produced by the three relative risk models were quite similar. As can be seen in Figure 3, the maps produced from the minimax and competitive exclusion models were nearly identical. These maps differed slightly from the relative risk map produced using the additive model as the relative risk in the additive model was more closely restricted to the areas of high population density. As can be seen in comparing Figure 1 with Figure 3, population density appears to have been the primary determinant of the relative risk of malaria in each of the three models. Areas of high human population density were found to be those in which the relative risk of malaria was greatest. The sole exception to this was in North Africa in which high human population densities were accompanied by relatively low relative risk values.
The sensitivity of the additive model was observed to be less than that of the competitive and minimax models. The standard deviations in the relative risk values ranged from 0 to 0.144 for the additive model to 0 to 0.280 for the competitive model and 0 to 0.279 for minimax model. In each model the majority of cells possessed a standard deviation less than 0.001. In the additive model 98.9% of cells had a standard deviation less than 0.001, while the percentages for the competitive and minimax models were 97.7% and 98.5%, respectively. Most cells were thus robust to variation in the parameter values. There was no clear pattern observed in the distribution of sensitivity across the landscape. While relative risk was concentrated in areas of high human population density, sensitivity was not.
Discussion
As explained below, our relative risk maps are preliminary and must be treated with due caution. However, the methodology developed here can be used to construct relative risk maps for other infectious diseases, with the risk models modified to reflect the presence of multiple agents and reservoirs, more complicated modes of transmission, and other relevant ecological factors. Because, as we and others have shown [9], [11], [16], [17], niche models of acceptable accuracy based on climatic and topographic parameters (as determined by the internal tests of software packages as well as from comparisons of maps produced by experts—see below for more discussion) can be constructed with sparse data, this methodology shows promise for the construction of relative risk maps for infectious diseases even when data are limited and not easily collected. Some suggestive recent work argues that, besides climatic and topographic factors, land use and land cover change influence the spread of malaria [39], [40]. The models presented here used land cover as one of the explanatory variables in the niche models; in future work we plan to include land use and to explore the effects of land cover in more detail, using a finer classification of land cover types.
A variety of different climatic and topographical factors were important for predicting the distributions of different species. Levine et al. [23] report similar variable results for the three Anopheles species that they model. These results have the implication that, to predict malaria vector species distributions, simple ecological heuristic rules (for instance, those incorporating optimal precipitation and/or temperature) will likely be unreliable and should be replaced with predictions of niche models based on as complete a set of environmental parameters as possible. Rogers and Randolph [41] have also noted the lack of relatively simple heuristic rules.
A perhaps not unexpected result is that the most important determinant of relative risk for malaria was human population density, assuming as we do here for Africa that (i) ecological and demographic factors rather than control measures are the determinants of risk and (ii) parasite densities are sufficiently high for disease spread. The extent of the effect of population density in this analysis is likely a result of the multiplicative nature of our risk models. However, this effect cannot be regarded solely as an artifact of the admitted simplicity of these models as any plausible model of infectious disease transmission should include the human population as a multiplicative factor. This dependence on population density has the implication that the expected local increase of population density due to increased urbanization [42], especially in Africa [43], will increasingly exacerbate the risk of malaria unless control measures are implemented [39].
Turning to the details of our results, with a few notable exceptions, the distributions for the niche models in Figure 2 closely follow the previously proposed distributions of most modeled species. This can be seen by comparing by region the vector distributions provided in Figure 2 with those previously presented in the literature.
The niche models of vectors within West Africa appear to coincide quite closely with the expert-based distributions of the modeled vectors within this region. Haworth [34] identified A. arabiensis, A. coustani, A. funestus, A. gambiae, and A. nili as primary and secondary malaria vectors in West Africa. This is consistent with the niche models of Figure 2 in which these three species are shown to possess extremely high relative probabilities of occurrence within this region. Haworth [34] also identified A. melas, A. moucheti, and A. paludis as vectors with limited presence in West Africa. This is consistent with the models in Figure 2 in which these species have limited distributions in parts of that region. In addition, Gillies and Coetzee [44] presented this region as containing A. arabiensis, A. gambiae, and A. melas, while White [45] reported the region as containing A. arabiensis, A. gambiae, A. melas, and A. funestus. The distributions of these species within this region provided by these two sources were thus consistent with the niche models of Figure 2.
In Southeast Africa and Madagascar the niche models again correspond closely to expert-based distributions. Haworth [34] identified A. arabiensis, A. coustani, A. funestus, A. gambiae, A. merus, and A. paludis as the primary and secondary malaria vectors within the region. In Figure 2 these species have high relative probabilities of occurrence in the region. Moreover, the distributions within this region of A. arabiensis, A. gambiae, and A. merus, as presented by Gillies and Coetzee [44], and the distributions of A. arabiensis, A. gambiae, A. funestus, and A. merus, as presented by White [45], were consistent with the niche models in Figure 2.
While the correspondence between the niche models produced in this analysis and the expert-based distributions of the Anopheles species was quite strong in most regions, discrepancies between these distributions are found in Central Africa. Gillies and Coetzee [44] presented the distributions of A. arabiensis and A. gambiae as stretching across Central Africa. However, these distributions conflict with the niche models of Figure 2, in which both vectors have low probabilities of occurrence throughout this region. In slight contrast to Gillies and Coetzee, White [45] depicted both A. gambiae and A. funestus as present in Central Africa, while depicting A. arabiensis as largely absent from the region. While this distribution of A. arabiensis agrees with the niche models of Figure 2, the proposed distributions of A. gambiae and A. funestus do not. Levine et al. [23] found both A. arabiensis and A. gambiae to be distributed throughout Central Africa, unlike what is seen in Figure 2. Similar results were obtained by Lindsay et al. [20] who found the climate throughout Central Africa to be suitable for both of these species. Rogers et al. [22] obtained results similar to those of both Levine et al. and Lindsay et al. with respect to the distribution of A. gambiae, yet found A. arabiensis to be largely absent from Central Africa (which thus agrees with Figure 2).
These discrepancies may simply be a result of the general lack of distributional data for Anopheles species in Central Africa. Fewer than 10 occurrence records within this region were available for use in the present analysis, with the other analyses likewise lacking much data for the region. If the niche models produced by Maxent are excessively conservative, then the low probability of occurrence associated with A. gambiae in Central Africa may be a consequence of the lack of any occurrence data for the species in this region. However, given the current absence of records of Anopheles species within Central Africa, the attribution of a low relative probability of occurrence to both A. arabiensis and A. gambiae within the region may be a correct prediction. The high AUC values associated with the niche models produced in this analysis support this claim. Field data from Central Africa are required to resolve the discrepancies between our predictions and other distributional maps..
Some other discrepancies appear to indicate that our models are an improvement over previous proposed distributions. For example, Coetzee [46] criticized the niche models of Levine et al. [23] for predicting the occurrence of A. quadriannulatus in areas of central Botswana that are supposed to be too arid for the species. She also questioned proposed distributions of Lindsay et al. [20] for including A. arabiensis in arid parts of South Africa. In contrast with these distributions, the niche models in Figure 2 predict small relative probabilities of occurrence for A. quadriannulatus and A. arabiensis in central Botswana and South Africa, respectively. While our results thus avoid these criticisms, since Levine et al.'s criticisms were meant to illustrate the general shortcomings of computer modeling, our results also argue against excessive skepticism about computational approaches in epidemiology.
Finally, the possibly questionable assumptions underlying our preliminary relative risk maps deserve explicit emphasis. The most important of these is that the relative abundances of species can be estimated using the relative probabilities of occurrence predicted by niche models. The relative risk maps based on the minimax model and, especially, the competitive exclusion model are less affected by this assumption than that based on the additive model as, in the former models, the relative probabilities of occurrence of the various vectors species were not aggregated. However, before the predictions of any of these models are fully accepted, the relationship between the predicted relative probabilities of occurrence of these species and their actual distributions must be empirically tested. The importance of such a test extends beyond epidemiology and will be relevant to all disciplines that use niche models (including conservation biology). It is essential to test this assumption for a wide variety of vector species before recommending its adoption. If the assumption holds, then absolute abundances, drawn from a portion of the area under investigation, can be used to calibrate the relative abundance predictions of the niche models, thus allowing for the determination of absolute abundances across the landscape for use in the risk models.
If the assumption does not hold, the situation is more difficult. In almost all epidemiological contexts, at the landscape or at larger scales, it is unlikely that the absolute abundances of vector species can be empirically measured for the entirety of the region of concern. It may be possible to obtain some measurements, and use traditional interpolation techniques such as splining and kriging to acquire estimates for other regions within the geographical boundaries of the measured cells. These can then be used along with the niche models to extrapolate across the landscape using statistical techniques such as regression to other areas that are predicted to be suitable (by the niche models) for the vector species. However, the accuracy of these traditional interpolation techniques in epidemiological contexts must likewise be empirically tested.
In addition to this primary assumption a number of secondary assumptions were made regarding the relationships between the vectors considered in this analysis. One such assumption was that the variable risk posed by P. vivax and P. falciparum could be ignored. As these two parasites differ both in the risk that they pose to humans [47] and in their geographical distribution [48], this assumption should be questioned. In this analysis the assumption was mandated by a lack of data on the varying abilities of most vectors to transmit these two parasites. A more complete analysis will require the explicit incorporation of the differential risk posed by these two parasites.
It was also assumed that the efficiency with which malaria parasites were transmitted, both to and from humans, is the same both for different vectors and in different regions. Since transmission efficiency varies between both species [49], [50] and regions [51], it should be explicitly included for a credible assessment of malaria risk. It was not included in this analysis due to a lack of data.
A similar caveat is needed with respect to the HBI values. Though each HBI value was treated as a species-specific parameter, these values are environmentally influenced. Measured HBI values for a species obtained at one region vary substantially from those measured at other regions, with the variation often depending primarily on the availability of human blood meals [52]. In addition, the data used to derive the HBI values used here were not obtained by random sampling; thus, there is reason to question whether these HBI values adequately reflect the varying anthrophilicity of the vectors. The spatial heterogeneity of HBI values also questions the use of a single HBI value for a vector. However, our sensitivity analysis shows that including such spatial heterogeneity would likely have had little impact upon the results. Relative risk would still have been shown to be concentrated in areas of high population density. Nevertheless, a more precise representation of malaria risk should consider such heterogeneity.
The questionable simplifying assumptions made in the construction of the preliminary relative risk maps presented in this paper show that to construct large-scale risk maps that go beyond the predicted relative probabilities of vector species presence will require much more data, and data of different types (including, for instance, abundance and transmission efficiency data) than what are now available in the literature. Furthermore, the use of more sophisticated transmission models than those on which Equations (1) and (2) are based will only require even more data. For disease risk analysis, research geared towards the acquisition of such data remain a high priority.
Acknowledgments
For discussion and help in developing this work we thank Trevon Fuller, Nelson Guda, Samraat Pawar, Steven Phillips, and Victor Sánchez-Cordero.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: The authors have no support or funding to report.
References
- 1.Waller LA, Carlin BP, Xia H, Gelfand AE. Hierarchical spatio-temporal mapping of disease rates. J Am Stat Assoc. 1997;92:607–617. [Google Scholar]
- 2.MacNab YC, Dean CB. Autoregressive spatial smoothing and temporal spline smoothing for mapping rates. Biometrics. 2001;57:949–956. doi: 10.1111/j.0006-341x.2001.00949.x. [DOI] [PubMed] [Google Scholar]
- 3.Snow RW, Marsh K, Le Sueur D. The need for maps of transmission intensity to guide malaria control in Africa. Parasitol Today. 1996;12:455–457. [Google Scholar]
- 4.World Health Organization. Geneva: World Health Organization; 2003. International travel and health: situations as on 1 January 2003. [Google Scholar]
- 5.Haydon DT, Randall DA, Matthews L, Knobel DL, Tallents LA, et al. Low-coverage vaccination strategies for the conservation of endangered species. Nature. 2006;443:692–695. doi: 10.1038/nature05177. [DOI] [PubMed] [Google Scholar]
- 6.Caldas de Castro M, Yamagat Y, Mtasiwa D, Tanner M, Utzinger J, et al. Integrated urban malaria control: a case study in Dar es Salaam, Tanzania. Am J Trop Med Hyg. 2004;71:103–117. [PubMed] [Google Scholar]
- 7.Gemperli A, Sogoba N, Fondjo E, Mabaso M, Bagayoko M, et al. Mapping malaria transmission in West and Central Africa. Trop Med Int Health. 2006;11:1032–1046. doi: 10.1111/j.1365-3156.2006.01640.x. [DOI] [PubMed] [Google Scholar]
- 8.Kiszewski A, Mellinger A, Spielman A, Malaney P, Sachs S, et al. A global index representing the stability of malaria transmission. Am J Trop Med Hyg. 2004;70:486–498. [PubMed] [Google Scholar]
- 9.Peterson AT. Ecological niche modeling and spatial patterns of disease transmission. Emerg Infect Dis. 2006;12:1822–1826. doi: 10.3201/eid1212.060373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Soberón J, Peterson AT. Interpretation of models of fundamental ecological niches and species' distributional areas. Biodiversity Informatics. 2005;2:1–10. [Google Scholar]
- 11.Phillips SJ, Anderson RP, Schapire RE. Maximum entropy modeling of species geographic distributions. Ecol Model. 2006;190:231–259. [Google Scholar]
- 12.Illoldi-Rangel P, Sanchez-Cordero V, Peterson AT. Predicting distributions of Mexican mammals using ecological niche modeling. J Mammal. 2004;85:658–662. [Google Scholar]
- 13.Ortega-Huerta MA, Peterson AT. Modelling spatial patterns of biodiversity for conservation prioritization in North-Eastern Mexico. Divers Distrib. 2004;10:39–54. [Google Scholar]
- 14.Fuller T, Munguía M, Mayfield M, Sánchez-Cordero V, Sarkar S. Incorporating connectivity into conservation planning: A multi-criteria case study from Central Mexico. Biol Conserv. 2006;133:131–142. [Google Scholar]
- 15.Fuller T, Sánchez-Cordero V, Illoldi-Rangel P, Linaje M, Sarkar S. The cost of postponing biodiversity conservation in Mexico. Biol Conserv. 2007;134:593–600. [Google Scholar]
- 16.Pawar SS, Koo MS, Kelley C, Ahmed MF, Choudhury S, et al. Conservation assessment and prioritization of areas in Northeast India: priorities for amphibians and reptiles. Biol Conserv. 2007 In press. [Google Scholar]
- 17.Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, et al. Novel methods improve predictions of species' distributions from occurrence data. Ecography. 2006;29:129–151. [Google Scholar]
- 18.Stockwell DRB, Peters D. The GARP modeling system: problems and solutions to automated spatial prediction. Int J Geogr Inf Sci. 1999;13:143–158. [Google Scholar]
- 19.Breiman L. Random Forests. Mach Learn. 2001;45:5–32. [Google Scholar]
- 20.Lindsay SW, Parson L, Thomas CJ. Mapping the ranges and relative abundance of the two principle African malaria vectors, Anopheles gambiae sensu stricto and An. arabiensis, using climate data. Proc Biol Sci. 1998;265:847–854. doi: 10.1098/rspb.1998.0369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kuhn KG, Campbell-Lendrum DH, Davies CR. A continental risk map for malaria mosquito (Diptera: Culicidae) vectors in Europe.”. J Med Entomol. 2002;39:621–630. doi: 10.1603/0022-2585-39.4.621. [DOI] [PubMed] [Google Scholar]
- 22.Rogers DJ, Randolph SE, Snow RW, Hay SI. Satellite imagery in the study and forecast of malaria. Nature. 2002;415:710–715. doi: 10.1038/415710a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Levine RS, Peterson AT, Benedict MQ. Geographic and ecologic distributions of the Anopheles gambiae complex predicted using a genetic algorithm. Am J Trop Med Hyg. 2004;70:105–109. [PubMed] [Google Scholar]
- 24.Levine RS, Peterson AT, Benedit MQ. Distribution of members of Anopheles quadrimaculatus Say s.l. (Diptera: Culicidae) and implications for their roles in malaria transmission in the United States. J Med Entomol. 2004;41:607–613. doi: 10.1603/0022-2585-41.4.607. [DOI] [PubMed] [Google Scholar]
- 25.Kleinschmidt I, Omumbo J, Briet O, van de Giesen N, Sogoba N, et al. An empirical malaria distribution map for West Africa. Trop Med Int Health. 2001;6:779–786. doi: 10.1046/j.1365-3156.2001.00790.x. [DOI] [PubMed] [Google Scholar]
- 26.Kleinschmidt I, Bagayoko M, Clarke GPY, Craig M, Le Sueur D. A spatial statistical approach to malaria mapping. Int J Epidemiol. 2000;29:355–361. doi: 10.1093/ije/29.2.355. [DOI] [PubMed] [Google Scholar]
- 27.Diggle P, Moyeed R, Rowlingston B, Thomson M. Childhood malaria in the Gambia: a case study in model-based geostatistics.”. J Roy Stat Soc B. 2002;51:493–506. [Google Scholar]
- 28.Smith DL, Dushoff J, McKenzie FE. The risk of a mosquito-borne infection in a heterogeneous environment. PLoS Bio. 2004;2:e368. doi: 10.1371/journal.pbio.0020368. doi:10.1371/journal.pbio.0020368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Smith DL, McKenzie FE. Statics and dynamics of malaria infection in Anopheles mosquitoes. Malar J. 2004;3:13. doi: 10.1186/1475-2875-3-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Black WC, Moore CG. Population biology as a tool to study vector-borne diseases. In: Marquardt WH, Black WC, Freier JE, Hagedorn HH, Hemingway J, editors. Biology of disease vectors second edition. Burlington: Elsevier Academic Press; 2005. [Google Scholar]
- 31.Robert HJ, Cameron SE, Parra JL, Jones PG, Jarvis A. Very high resolution interpolated climate surfaces for global land areas. Int J Climatol. 2005;25:1965–1978. [Google Scholar]
- 32.ESRI. Redlands: ESRI; 2004. ArcGIS 9.1. [Google Scholar]
- 33.Hansen MC, Defries RS, Townshend JRG, Sohlberg R. Global land cover classification at 1 km spatial resolution using a classification tree approach. Int J Remote Sens. 2000;21:1331–1364. [Google Scholar]
- 34.Haworth J. The global distribution of malaria and the present control effort. In: Wernsdorfer WH, McGregor I, editors. Malaria: principles and practice of malariology. Edinburgh: Churchill Livingstone; 1988. pp. 1379–1419. [Google Scholar]
- 35.Center for International Earth Science Information Network, Columbia University and Centro Internacional de Agricultura Tropical. Palisades: Columbia University; 2004. Gridded Population of the World (GPW), Version 3. Available: http://beta.sedac.ciesin.columbia.edu/gpw. [Google Scholar]
- 36.Schneider P, Takken W, McCall PJ. Interspecific competition between sibling species larvae of Anopheles arabiensis and An. gambiae. Med Vet Entomol. 2000;14:165–170. doi: 10.1046/j.1365-2915.2000.00204.x. [DOI] [PubMed] [Google Scholar]
- 37.Antonio-Nkondjio C, Kerah CH, Simard F, Awono-Ambene P, Chouaibou, et al. Complexity of the malaria vectorial system in Cameroon: contribution of secondary vectors to malaria transmission. J Med Entomol. 2006;43:1215–1221. doi: 10.1603/0022-2585(2006)43[1215:cotmvs]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 38.Cano J, Descalzo MA, Moreno M, Chen Z, Nzambo S, et al. Spatial variability in the density, distribution and vectorial capacity of anopheline species in a high transmission village (Equatorial Guinea). Malar j. 2006;5:21. doi: 10.1186/1475-2875-5-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hay SI, Guerra CA, Tatem AJ, Atkinson PM, Snow RW. Urbanization, malaria transmission, and disease burden in Africa. Nat Rev Microbio. 2005;3:81–90. doi: 10.1038/nrmicro1069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Guerra CA, Snow RW, Hay SI. A global assessment of closed forest, deforestation, and malaria risk. Ann Trop Med Parasitol. 2006;100:189–204. doi: 10.1179/136485906X91512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rogers DJ, Randolph SE. The global spread of malaria in a future, warmer world. Science. 2000;289:1763–1766. doi: 10.1126/science.289.5485.1763. [DOI] [PubMed] [Google Scholar]
- 42.United Nations. New York: United Nations; 2002. World urbanization prospects: the 2001 revision. Data, tables, and highlights. [Google Scholar]
- 43.United Nations. New York: United Nations; 2004. World urbanization prospects: the 2003 revision. Data, tables and highlights. [Google Scholar]
- 44.Gillies MT, Coetzee M. Johannesburg: The South African Institute for Medical Research; 1987. A supplement to the Anophelinae of Africa South of the Sahara. [Google Scholar]
- 45.White GB. Geneva: World Health Organization, WHO/VBC; 1989. Malaria. Geographical Distribution of Arthropod Borne Diseases and their Principal Vectors. pp. 7–22. [Google Scholar]
- 46.Coetzee M. Distribution of the African malaria vectors of the Anopheles. Am J Trop Med Hyg. 2004;70:103–104. [PubMed] [Google Scholar]
- 47.Collins FH, Paskewitz SM. Malaria: current and future prospects for control. Annu Rev Entomol. 1995;40:195–219. doi: 10.1146/annurev.en.40.010195.001211. [DOI] [PubMed] [Google Scholar]
- 48.Mendis K, Sina BJ, Marchesini P, Carter R. The neglected burden of Plasmodium vivax malaria. Am J Trop Med Hyg. 2001;64:97–106. doi: 10.4269/ajtmh.2001.64.97. [DOI] [PubMed] [Google Scholar]
- 49.Gillies MT, Wilkes TJ. A study of the age-composition of populations of Anopheles gambiae Giles and A. funestus Giles in North-Eastern Tanzania. Bull Entomol Res. 1965;56:237–262. doi: 10.1017/s0007485300056339. [DOI] [PubMed] [Google Scholar]
- 50.Haji H, Smith T, Meuwissen JT, Sauerwein R, Charlwood JD. Estimation of the infectious reservoir of Plasmodium falciparum in natural vector populations based on oocyst size. Trans R Soc Trop Med Hyg. 1996;90:494–497. doi: 10.1016/s0035-9203(96)90292-8. [DOI] [PubMed] [Google Scholar]
- 51.Bonnet S, Gouagna LC, Paul RE, Safeukui I, Meunier JY, et al. Estimation of malaria transmission from humans to mosquitoes in two neighbouring villages in South Cameroon: evaluation and comparison of several indices. Trans R Soc Trop Med Hyg. 2003;97:53–59. doi: 10.1016/s0035-9203(03)90022-8. [DOI] [PubMed] [Google Scholar]
- 52.Garrett-Jones C. Geneva: World Health Organization, WHO/Mal/391; 1963. The human blood index of some anopheline mosquitoes, with reference to epidemiological assessment during malaria eradication. [Google Scholar]
- 53.Ameneshewa BB, Service AW. Resting habits of Anopheles arabiensis in the Awash river valley of Ethiopia. Ann Trop Med Parasitol. 1996;90:515–521. doi: 10.1080/00034983.1996.11813077. [DOI] [PubMed] [Google Scholar]
- 54.Bryan JH, Petrarca V, Di Deco MA, Coluzzi M. Adult behavior of members of the Anopheles gambiae complex in the Gambia with special reference to An. melas and its chromsomal variants. Parassitologia. 1987;29:221–249. [PubMed] [Google Scholar]
- 55.Bryan JH, Di Deco MA, Petrarca V, Coluzzi M. Inversion polymorphism and incipient speciation in Anopheles gambiae s.str. in The Gambia West Africa. Genetica. 1982;59:167–176. [Google Scholar]
- 56.Cambournac FJ, Petrarca V, Coluzzi M. Anopheles arabiensis in the Cape Verde archipelago. Parassitologia. 1982;24:265–267. [PubMed] [Google Scholar]
- 57.Charlwood JD, Edoh D. Polymerase chain reaction used to describe larval habitat use by Anopheles gambiae complex (Diptera: Culicidae) in the environs of Ifakara, Tanzania. J Med Entomol. 1996;33:202–204. doi: 10.1093/jmedent/33.2.202. [DOI] [PubMed] [Google Scholar]
- 58.Clarke D, Odialla H, Ouma J, Kenny V, MacCabe R, et al. A malariometric survey in Turkana District Kenya: chemosensitivity in vivo of Plasmodium falciparum infections and identity of the vector. Trans R Soc Trop Med Hyg. 1996;90:302–304. doi: 10.1016/s0035-9203(96)90262-x. [DOI] [PubMed] [Google Scholar]
- 59.Coetzee M. Comparative morphology and multivariate analysis for the discrimination of four members of the Anopheles gambiae group in Southern Africa. Mosq Sys. 1989;21:100–116. [Google Scholar]
- 60.Coetzee M, Hunt RH, Baack L, Davidson G. Distribution of mosquitoes belonging to the Anopheles gambiae complex, including malaria vectors, south of latitude 15°S. S Afr J Sci. 1993;89:227–231. [Google Scholar]
- 61.Constantini C, Li SG, Della-Torre A, Sagnon N, Coluzzi M, et al. Density, survival and dispersal of Anopheles gambiae complex mosquitoes in a west African Savana village. Med Vet Entomol. 1996;10:203–219. doi: 10.1111/j.1365-2915.1996.tb00733.x. [DOI] [PubMed] [Google Scholar]
- 62.Coosemans M, Petrarca V, Barutwanayo M, Coluzzi M. Species of the Anopheles gambiae complex and chromosomal polymorphism in a rice-growing area of the Rusizi Valley (Republic of Burundi). Parassitologia. 1989;31:113–122. [PubMed] [Google Scholar]
- 63.Faye O, Konate L, Mouchet J, Fontenille D, Sy N, et al. Indoor resting by outdoor biting females of Anopheles gambiae complex (Diptera: Culicidae) in the Sahel of northern Senegal. J Med Entomol. 1997;34:285–289. doi: 10.1093/jmedent/34.3.285. [DOI] [PubMed] [Google Scholar]
- 64.Faye O, Gaye O, Faye O, Diallo S. Malaria transmission in villages, near or far of a mangrove area in Senegal. B Soc Pathol Exot. 1994;87:157–163. [PubMed] [Google Scholar]
- 65.Fontenille D, Faye O, Konate L, Sy N, Collins FH. Comparison of PCR and cytogenetic methods for the identification of mosquito species of the Anopheles gambiae complex in Senegal. Ann Parasitol Hum Comp. 1993;68:239–240. doi: 10.1051/parasite/1993685239. [DOI] [PubMed] [Google Scholar]
- 66.Fontenille D, Lepers JP, Coluzzi M, Campbell GH, Rakotoarivony I, et al. Malaria transmission and vector biology on Sainte Marie Island, Madagascar. J Med Entomol. 1992;29:197–202. doi: 10.1093/jmedent/29.2.197. [DOI] [PubMed] [Google Scholar]
- 67.Githeko AK, Adungo NI, Karanja DM, Hawley WA, Vulule JM, et al. Some observations on the biting behavior of Anopheles gambiae s.s Anopheles arabiensis and Anopheles funestus and their implication for malaria control. Exp Parasitol. 1996;82:306–315. doi: 10.1006/expr.1996.0038. [DOI] [PubMed] [Google Scholar]
- 68.Githeko AK, Brandling-Bennett AD, Beier M, Mbogo CM, Atieli FK, et al. Confirmation that Plasmodium falciparum has aperiodic infectivity to Anopheles gambiae. Med Vet Entomol. 1993;7:373–376. doi: 10.1111/j.1365-2915.1993.tb00708.x. [DOI] [PubMed] [Google Scholar]
- 69.Green CA. Cladistic analysis of mosquito chromosome data (Anopheles (Cellia) Myzomyia). J Hered. 1982;73:3–11. [PubMed] [Google Scholar]
- 70.Hogg JC, Thomson MC, Hurd H. Comparative fecundity and associated factors for two sibling species of the Anopheles gambiae complex occurring sympatrically in The Gambia. Med Vet Entomol. 1996;10:385–391. doi: 10.1111/j.1365-2915.1996.tb00761.x. [DOI] [PubMed] [Google Scholar]
- 71.Ijumba JN, Mwangi RW, Beier JC. Malaria transmission potential of Anopheles mosquitoes in the Mwea-Tebere irrigation scheme, Kenya. Med Vet Entomol. 1990;4:425–432. doi: 10.1111/j.1365-2915.1990.tb00461.x. [DOI] [PubMed] [Google Scholar]
- 72.La Grange JJP. Survey of anopheline mosquitoes (Diptera: Culicidae) in a malarious area of Swaziland. Afr Entomol. 1995;3:217–219. [Google Scholar]
- 73.Mbogo CN, Baya NM, Ofulla AV, Githure JI, Snow RW. The impact of permethrin-impregnated bednets on malaria vectors of the Kenyan coast. Med Vet Entomol. 1996;10:251–259. doi: 10.1111/j.1365-2915.1996.tb00739.x. [DOI] [PubMed] [Google Scholar]
- 74.Mekuria Y, Petrarca V, Tesfamariam T. Cytogenetic studies on the malaria vector mosquito Anopheles arabiensis Patton in the Awash Valley, Ethiopia. Parassitologia. 1982;24:237–243. [PubMed] [Google Scholar]
- 75.Mnzava AE, Rwegoshora RT, Wilkes TJ, Curtis CF. Anopheles arabiensis and An. gambiae chromosomal inversion polymorphism feeding and resting behaviour in relation to insecticide house-spraying in Tanzania. Med Vet Entomol. 1995;9:316–324. doi: 10.1111/j.1365-2915.1995.tb00140.x. [DOI] [PubMed] [Google Scholar]
- 76.Mnzava AE, Mutinga MJ, Staak C. Host blood meals and chromosomal inversion polymorphism in Anopheles arabiensis in the Baringo District of Kenya. J Am Mosq Control Assoc. 1994;10:507–510. [PubMed] [Google Scholar]
- 77.Mnzava AE, Kilama WL, Kasigwa PF. Application of a biochemical key to study transmission of malaria and Bancroftian filariasis in sibling species of the Anopheles gambiae complex in north-eastern Tanzania. Acta Trop. 1989;46:323–333. doi: 10.1016/0001-706x(89)90045-4. [DOI] [PubMed] [Google Scholar]
- 78.Mnzava AE, Kilama WL. Observations on the distribution of the Anopheles gambiae complex in Tanzania. Acta Trop. 1986;43:277–282. [PubMed] [Google Scholar]
- 79.Mosha FW, Petrarca V. Ecological studies on Anopheles gambiae complex sibling species on the Kenya coast. Trans R Soc Trop Med Hyg. 1983;77:344–345. doi: 10.1016/0035-9203(83)90161-x. [DOI] [PubMed] [Google Scholar]
- 80.Mosha FW, Subra R. Geneva: World Health Organization; 1982. Ecological studies on Anopheles gambiae complex sibling species in Kenya 1 preliminary observations on their geographical distribution and chromosomal polymorphic inversions. Document WHO/VBC/82867. [Google Scholar]
- 81.Mpofu SM. Seasonal vector density and disease incidence patterns of malaria in an area of Zimbabwe. Trans R Soc Trop Med Hyg. 1985;79:169–175. doi: 10.1016/0035-9203(85)90327-x. [DOI] [PubMed] [Google Scholar]
- 82.Nigatu W, Curtis CF, Lulu M. Test for association of DDT resistance with inversion polymorphism in Anopheles arabiensis from Ethiopia. J Am Mosq Control Assoc. 1995;11:238–240. [PubMed] [Google Scholar]
- 83.Paskewitz SM, Ng K, Coetzee M, Hunt RH. Evaluation of the polymerase chain reaction method for identifying members of the Anopheles gambiae (Diptera: Culicidae) complex in southern Africa. J Med Entomol. 1993;30:953–957. doi: 10.1093/jmedent/30.5.953. [DOI] [PubMed] [Google Scholar]
- 84.Petrarca V, Sabatinelli G, Youre YT, Di Deco MA. Morphometric multivariate analysis of field samples of adult Anopheles arabiensis and An. gambiae s.s. (Diptera: Culicidae). J Med Entomol. 1998;35:16–25. doi: 10.1093/jmedent/35.1.16. [DOI] [PubMed] [Google Scholar]
- 85.Petrarca V, Beier JC, Onyango F, Koros J, Asiago C, et al. Species composition of the Anopheles gambiae complex (Diptera: Culicidae) at two sites in western Kenya. J Med Entomol. 1991;28:307–313. doi: 10.1093/jmedent/28.3.307. [DOI] [PubMed] [Google Scholar]
- 86.Petrarca V, Vercruysse J, Coluzzi M. Observations on the Anopheles gambiae complex in the Senegal River Basin West Africa. Med Vet Entomol. 1987;1:303–312. doi: 10.1111/j.1365-2915.1987.tb00359.x. [DOI] [PubMed] [Google Scholar]
- 87.Petrarca V, Petrangeli G, Rossi P, Sabatinelli G. Chromosomal study of Anopheles gambiae and Anopheles arabiensis in Ouagadougou (Burkina Faso) and various neighboring villages. Parassitologia. 1986;28:41–61. [PubMed] [Google Scholar]
- 88.Petrarca V, Carrara GC, Di Deco MA, Petrangeli G. Observazioni citogenetiche e biometriche sui membri del complesso Anopheles gambiae in Mozambico. Parassitologia. 1984;26:247–259. [PubMed] [Google Scholar]
- 89.Petrarca V, Carrara GC, Di Deco MA, Petrangeli G. Il complesso Anopheles gambiae in Guinea Bissau. Parassitologia. 1983;25:29–39. [PubMed] [Google Scholar]
- 90.Robert V, Carnevale P. Influence of deltamethrin treatment of bed nets on malaria transmission in the Kou valley Burkina Faso. Bull World Health Organ. 1991;69:735–740. [PMC free article] [PubMed] [Google Scholar]
- 91.Robert V, Verhave JP, Carnevale P. Plasmodium falciparum infection does not increase the precocious mortality rate of Anopheles gambiae. Trans R Soc Trop Med Hyg. 1990;84:346–347. doi: 10.1016/0035-9203(90)90309-3. [DOI] [PubMed] [Google Scholar]
- 92.Robert V, Petrarca V, Carnevale P, Ovazza L, Coluzzi M. Analyse cytogenetique du complexe Anopheles gambiae dans la region de Bobo-Dioulasso (Burkina Faso). Ann Parasitol Hum Comp. 1989;64:290–311. [Google Scholar]
- 93.Sharp BL, Le Sueur D, Wilken GB, Bredenkamp BL, Nqxongo S, et al. Assessment of the residual efficiency of lambda-cyhalothrin 2 A Comparison with DDT for the intradomiciliary control of Anopheles arabiensis in South Africa. J Am Mosq Control Assoc. 1993;9:414–420. [PubMed] [Google Scholar]
- 94.Sharp BL, Le Sueur D. Behavioral variation of Anopheles arabiensis (Diptera: Culicidae) populations in Natal, South Africa. Bull Entomol Res. 1991;81:107–110. [Google Scholar]
- 95.Smits A, Roelants P, Van Bortel W, Coosemans M. Enzyme polymorphisms in the Anopheles gambiae (Diptera: Culicidae) complex related to feeding and resting behavior in the Imbo Valley, Burundi. J Med Entomol. 1996;33:545–553. doi: 10.1093/jmedent/33.4.545. [DOI] [PubMed] [Google Scholar]
- 96.Taylor CE, Toure YT, Coluzzi M, Petrarca V. Effective population size and persistence of Anopheles arabiensis during the dry season in West Africa. Med Vet Entomol. 1993;7:351–357. doi: 10.1111/j.1365-2915.1993.tb00704.x. [DOI] [PubMed] [Google Scholar]
- 97.Thomson MC, Connor SJ, Quinones ML, Jawara M, Todd J, et al. Movement of Anopheles gambiae sl malaria vectors between villages in The Gambia. Med Vet Entomol. 1995;9:413–419. doi: 10.1111/j.1365-2915.1995.tb00015.x. [DOI] [PubMed] [Google Scholar]
- 98.Toure YT, Traore SF, Sankare O, Sow MY, Coulibaly A, et al. Perennial transmission of malaria by the Anopheles gambiae complex in a north Sudan savanna area of Mali. Med Vet Entomol. 1996;10:197–199. doi: 10.1111/j.1365-2915.1996.tb00731.x. [DOI] [PubMed] [Google Scholar]
- 99.Van Rensburg AJ, Hunt RH, Koekemoer LL, Coetzee M, Shiff CJ, et al. The polymerase chain reaction method as a tool for identifying members of the Anopheles gambiae complex (Diptera: Culicidae) in northeastern Tanzania. J Am Mosq Control Assoc. 1996;12:271–274. [PubMed] [Google Scholar]
- 100.Walker TW, Robert LL, Copeland RA, Githeko AZ, Wirtz RA, et al. Field evaluation of arthropod repellents deet and a piperidine compound AI3-37220 against Anopheles funestus and Anopheles arabiensis in western Kenya. J Am Mosq Control Assoc. 1996;12:172–176. [PubMed] [Google Scholar]
- 101.Adugna N, Petros B. Determination of the human blood index of some anopheline mosquitos by using ELISA. Ethiop Med J. 1996;34:1–10. [PubMed] [Google Scholar]
- 102.Aranda C, Aponte JJ, Saute F, Casimiro S, Pinto J, et al. Entomological characteristics of malaria transmission in Manhica a rural area in Southern Mozambique. J Med Entomol. 2005;42:180–186. doi: 10.1093/jmedent/42.2.180. [DOI] [PubMed] [Google Scholar]
- 103.Awolola TS, Okwa O, Hunt RH, Ogunrinade AF, Coetzee M. Dynamics of the malaria-vector populations in coastal Lagos, south-western Nigeria. Ann Trop Med Parasitol. 2002;96:75–82. doi: 10.1179/000349802125000538. [DOI] [PubMed] [Google Scholar]
- 104.Carnahan J, Zheng L, Taylor CE, Toure YT, Norris DE, et al. Genetic differentiation of Anopheles gambiae s.s. populations in Mali West Africia using microsatellite loci. J Hered. 2002;93:249–253. doi: 10.1093/jhered/93.4.249. [DOI] [PubMed] [Google Scholar]
- 105.Chen H, Githeko AK, Zhou G, Githure JI, Yan G. New records of Anopheles arabiensis breeding on the Mount Kenya Highlands indicate indigenous malaria transmission. Malar J. 2006;5:17. doi: 10.1186/1475-2875-5-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Constantini C, Sagnon N, Della Torre A, Diallo M, Brady J, et al. Odor-mediated host preferences of West African mosquitoes with particular reference to malaria vectors. Am J Trop Med Hyg. 1998;58:56–63. doi: 10.4269/ajtmh.1998.58.56. [DOI] [PubMed] [Google Scholar]
- 107.Dekker T, Takken W. Differential responses of mosquito sibling species Anopheles arabiensis and An. quadriannulatus to carbon dioxide a man or a calf. Med Vet Entomol. 1998;12:136–140. doi: 10.1046/j.1365-2915.1998.00073.x. [DOI] [PubMed] [Google Scholar]
- 108.Fontenille D, Lochouarn L, Diatta M, Sokhna C, Dia I, et al. Four years' entomological study of the transmission of seasonal malaria in Senegal and the bionomics of Anopheles gambiae and A. arabiensis. Trans R Soc Trop Med Hyg. 1997;91:647–652. doi: 10.1016/s0035-9203(97)90506-x. [DOI] [PubMed] [Google Scholar]
- 109.Habtewold T, Prior A, Torr SJ, Gibson G. Could insecticide-treated cattle reduce Afrotropical malaria transmission? Effects of deltamethrin-treated Zebu on Anopheles arabiensis behavior and survival in Ethiopia. Med Vet Entomol. 2004;18:408–417. doi: 10.1111/j.0269-283X.2004.00525.x. [DOI] [PubMed] [Google Scholar]
- 110.Hunt RH, Coetzee M, Fettene M. The Anopheles gambiae complex: a new species from Ethiopia. Trans R Soc Trop Med Hyg. 1998;92:231–235. doi: 10.1016/s0035-9203(98)90761-1. [DOI] [PubMed] [Google Scholar]
- 111.Ijumba JN, Mosha FW, Lindsay SW. Malaria transmission risk variations derived from different agricultural practices in an irrigated area of northern Tanzania. Med Vet Entomol. 2002;16:28–38. doi: 10.1046/j.0269-283x.2002.00337.x. [DOI] [PubMed] [Google Scholar]
- 112.Klinkenberg E, Takken W, Huibers F, Toure YT. The phenology of malaria mosquitoes in irrigated rice fields in Mali. Acta Trop. 2003;85:71–82. doi: 10.1016/s0001-706x(02)00254-1. [DOI] [PubMed] [Google Scholar]
- 113.Leong Pock Tsy J, Duchemin J, Marrama L, Rabarison P, Le Goff G, et al. Distribution of the species of the Anopheles gambiae complex and first evidence of Anopheles merus as a malaria vector in Madagascar. Malar J. 2003;2:33. doi: 10.1186/1475-2875-2-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Mendis C, Jacobsen JL, Gamage-Mendis A, Bule E, Dgedge M, et al. Anopheles arabiensis and An. funestus are equally important vectors of malaria in Matola coastal suburb of Maputo southern Mozambique. Med Vet Entomol. 2000;14:171–180. doi: 10.1046/j.1365-2915.2000.00228.x. [DOI] [PubMed] [Google Scholar]
- 115.Muturi E, Shililu J, Jacob B, Gu W, Githure J, et al. Mosquito species diversity and abundance in relation to land use in a riceland agroecosystem in Mwea, Kenya. J Vector Ecol. 2006;31:129–137. doi: 10.3376/1081-1710(2006)31[129:msdaai]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 116.Onyabe DJ, Conn JE. The distribution of two major malaria vectors Anopheles gambiae and Anopheles arabiensis in Nigeria. Mem Inst Oswaldo Cruz. 2001;96:1081–1084. doi: 10.1590/s0074-02762001000800009. [DOI] [PubMed] [Google Scholar]
- 117.Petrarca V, Nugud AD, Elkarim Ahmed MA, Haridi AM, Di Deco MA, et al. Cytogenetics of the Anopheles gambiae complex in Sudan with special reference to An. arabiensis: relationships with East and West African populations. Med Vet Entomol. 2000;14:149–164. doi: 10.1046/j.1365-2915.2000.00231.x. [DOI] [PubMed] [Google Scholar]
- 118.Robert V, Awono-Ambene HP, Thioulouse J. Ecology of larval mosquitoes with special reference to Anopheles arabiensis (Diptera: Culicidae) in market-garden wells in urban Dakar, Senegal. J Med Entomol. 1998;35:948–955. doi: 10.1093/jmedent/35.6.948. [DOI] [PubMed] [Google Scholar]
- 119.Simard F, Lehmann T, Lemasson J, Diatta M, Fontenille D. Persistence of Anopheles arabiensis during the severe dry season conditions in Senegal: an indirect approach using microsatellite loci. Insect Mol Biol. 2000;9:467–479. doi: 10.1046/j.1365-2583.2000.00210.x. [DOI] [PubMed] [Google Scholar]
- 120.Taye A, Hadis M, Adugna N, Tilahun D, Wirtz RA. Biting behavior and Plasmodium infection rates of Anopheles arabiensis from Sille, Ethiopia. Acta Trop. 2006;97:50–54. doi: 10.1016/j.actatropica.2005.08.002. [DOI] [PubMed] [Google Scholar]
- 121.Toure YT, Petrarca V, Traore SF, Coulibaly A, Maiga HM, et al. The distribution and inversion polymporphism of chromosomally recognized taxa of the Anopheles gambiae complex in Mali, West Africa. Parasitologia. 1998;40:477–511. [PubMed] [Google Scholar]
- 122.Tripet F, Dolo G, Lanzaro GC. Multilevel analyses of genetic differentiation in Anopheles gambiae s.s. reveal patterns of gene flow important for malaria-fighting mosquito projects. Genetics. 2005;169:313–324. doi: 10.1534/genetics.104.026534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Tripet F, Toure YT, Dolo G, Lanzaro GC. Frequency of multiple inseminations in field-collected Anopheles gambiae females revealed by DNA analysis of transferred sperm. Am J Trop Med Hyg. 2003;68:1–5. [PubMed] [Google Scholar]
- 124.Wondji C, Simard F, Petrarca V, Etang J, Santolamazza F, et al. Species and populations of the Anopheles gambiae complex in Cameroon with special emphasis on chromosomal and molecular forms of Anopheles gambiae s.s. J Med Entomol. 2005;42:998–1005. doi: 10.1093/jmedent/42.6.998. [DOI] [PubMed] [Google Scholar]
- 125.Woyessa A, Gebre-Michael T, Ali A. An indigenous malaria transmission in the outskirts of Addis Ababa, Akaki Town and its environs. Ethiop J Health Dev. 2004;18:2–7. [Google Scholar]
- 126.Yawson AE, McCall PJ, Wilson MD, Donnelly MJ. Species abundance and insecticide resistance of Anopheles gambiae in selected areas of Ghana and Burkina Faso. Med Vet Entomol. 2004;18:372–377. doi: 10.1111/j.0269-283X.2004.00519.x. [DOI] [PubMed] [Google Scholar]
- 127.Coene J. Malaria in urban and rural Kinshasa: the entomological input. Med Vet Entomol. 1993;7:127–137. doi: 10.1111/j.1365-2915.1993.tb00665.x. [DOI] [PubMed] [Google Scholar]
- 128.Diallo M, Ba Y, Sall AA, Diop OM, Ndione JA, et al. Amplification of the sylvatic cycle of dengue virus type 2 Senegal 1999–2000: entomological findings and epidemiological considerations. Emerg Infect Dis. 2003;9:362–367. doi: 10.3201/eid0903.020219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Antonio-Nkondjio C, Awono-Ambene P, Toto JC, Meunier JY, Zebaze-Kemleu S, et al. High malaria transmission intensity in a village close to Yaounde the capital city of Cameroon. J Med Entomol. 2002;39:350–355. doi: 10.1603/0022-2585-39.2.350. [DOI] [PubMed] [Google Scholar]
- 130.Coetzee M. Anopheles crypticus, new species from South Africa is distinguished from Anopheles coustani (Diptera: Culicidae). Mosq Syst. 1994;26:125–131. [Google Scholar]
- 131.Dia I, Diallo D, Duchemin J, Ba Y, Konate L, et al. Comparisons of human-landing catches and odor-baited entry traps for sampling malaria vectors in Senegal. J Med Entomol. 2005;42:104–109. doi: 10.1093/jmedent/42.2.104. [DOI] [PubMed] [Google Scholar]
- 132.Drakeley C, Schellenberg D, Kihonda J, Sousa CA, Arez AP, et al. An estimation of the entomological inoculation rate for Ifakara: a semi-urban area in a region of intense malaria transmission in Tanzania. Trop Med Int Health. 2003;8:767–774. doi: 10.1046/j.1365-3156.2003.01100.x. [DOI] [PubMed] [Google Scholar]
- 133.Elissa N, Karch S, Bureau P, Ollomo B, Lawoko M, et al. Malaria transmission in a region of savanna-forest mosaic Haut-Ogooue Gabon. J Am Mosq Control Assoc. 1999;15:15–23. [PubMed] [Google Scholar]
- 134.Gimnig JE, Ombok M, Kamau L, Hawley WA. Characteristics of larval anopheline (Diptera: Culicidae) habitats in western Kenya. J Med Entomol. 2001;38:282–288. doi: 10.1603/0022-2585-38.2.282. [DOI] [PubMed] [Google Scholar]
- 135.Logan TM, Linthicum KJ, Thande PC, Wagateh JN, Roberts CR. Mosquito species collected from a marsh in western Kenya during the long rains. J Am Mosq Control Assoc. 1991;7:395–399. [PubMed] [Google Scholar]
- 136.Mboera LEG, Takken W. Odour-mediated host preference of Culex quinquefasciatus in Tanzania. Entomol Exp Appl. 1999;92:83–88. [Google Scholar]
- 137.Sylla EHK, Kun JFJ, Kremsner PG. Mosquito distribution and entomological inoculation rates in three malaria-endemic areas in Gabon. Trans R Soc Trop Med Hyg. 2000;94:652–656. doi: 10.1016/s0035-9203(00)90219-0. [DOI] [PubMed] [Google Scholar]
- 138.Antonio-Nkondjio C, Simard F, Awono-Ambene P, Ngassam P, Toto J, et al. Malaria vectors and urbanization in the equatorial forest region of south Cameroon. Trans R Soc Trop Med Hyg. 2005;99:347–354. doi: 10.1016/j.trstmh.2004.07.003. [DOI] [PubMed] [Google Scholar]
- 139.Traore-Lamizana M, Fontenille D, Diallo M, Ba Y, Zeller HG, et al. Arbovirus surveillance from 1990 to 1995 in the Barkedji area (Ferlo) of Senegal, a possible natural focus of rift valley fever virus. J Med Entomol. 2001;38:480–492. doi: 10.1603/0022-2585-38.4.480. [DOI] [PubMed] [Google Scholar]
- 140.Ayala D, Le Goff G, Robert V, de Jong P, Takken W. Population structure of the malaria vector Anopheles funestus (Dipetera: Culicidae) in Madagascar and Comoros. Acta Trop. 2006;97:292–300. doi: 10.1016/j.actatropica.2005.12.002. [DOI] [PubMed] [Google Scholar]
- 141.Cohuet A, Dia I, Simard F, Raymond M, Fontenille D. Population structure of the malaria vector Anopheles funestus in Senegal based on microsatellite and cytogenetic data. Insect Mol Biol. 2004;13:251–258. doi: 10.1111/j.0962-1075.2004.00482.x. [DOI] [PubMed] [Google Scholar]
- 142.Cohuet A, Simard F, Wondji CS, Antonio-Nkondjio C, Awono-Ambene P, et al. High malaria transmission intensity due to Anopheles funestus (Diptera: Culicidae) in a village of savannah-forest transition area in Cameroon. J Med Entomol. 2004;41:901–905. doi: 10.1603/0022-2585-41.5.901. [DOI] [PubMed] [Google Scholar]
- 143.Cohuet A, Simard F, Toto J, Kengne P, Coetzee M, et al. Species identification within the Anopheles funestus group of malaria vectors in Cameroon and evidence for a new species. Am J Trop Med Hyg. 2003;69:200–205. [PubMed] [Google Scholar]
- 144.Cuamba N, Choi KS, Townson H. Malaria vectors in Angola: distribution of species and molecular forms of the Anopheles gambiae complex their pyrethroid insecticide knockdown resistance (kdr) status and Plasmodium falciparum sporozoite rates. Malar J. 2006;5:2. doi: 10.1186/1475-2875-5-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Dossou-yovo J, Doannio JMC, Riviere F, Chauvancy G. Malaria in Côte d'Ivoire wet savannah region: the entomological input. Trop Med Parasitol. 1995;46:263–269. [PubMed] [Google Scholar]
- 146.Duchemin JB, Leong Pock Tsy JM, Rabarison P, Roux J, Coluzzi M, et al. Zoophily of Anopheles arabiensis and An. gambiae in Madagascar demonstrated by odourbaited entry traps. Med Vet Entomol. 2001;15:50–57. doi: 10.1046/j.1365-2915.2001.00276.x. [DOI] [PubMed] [Google Scholar]
- 147.Fillinger U, Sonye G, Killeen GF, Knols BGJ, Becker N. The practical importance of permanent and semipermanent habitats for controlling aquatic stages of Anopheles gambiae sensu lato mosquitoes: operational observations from a rural town in western Kenya. Trop Med Int Health. 2004;9:1274–1289. doi: 10.1111/j.1365-3156.2004.01335.x. [DOI] [PubMed] [Google Scholar]
- 148.Kamau L, Munyekenye GO, Koekemoer LL, Hunt RH, Coetzee M. A survey of Anopheles funestus (Diptera: Culicidae) group of mosquitoes from 10 sites in Kenya with special emphasis on population genetic structure based on chromosomal inversion karyotypes. J Med Entomol. 2003;40:664–671. doi: 10.1603/0022-2585-40.5.664. [DOI] [PubMed] [Google Scholar]
- 149.Kamau L, Hunt R, Coetzee M. Analysis of the population structure of Anopheles funestus (Diptera: Culicidae) from western and coastal Kenya using paracentric chromosomal inversion frequencies. J Med Entomol. 2002;39:78–83. doi: 10.1603/0022-2585-39.1.78. [DOI] [PubMed] [Google Scholar]
- 150.Mukabayire O, Boccolini D, Lochouarn L, Fontenille D, Besansky NJ. Mitochondrial and ribosomal internal transcribed spacer (ITS2) diversity of the African malaria vector Anopheles funestus. Mol Ecol. 1999;8:289–297. doi: 10.1046/j.1365-294x.1999.00567.x. [DOI] [PubMed] [Google Scholar]
- 151.Quakyi IA, Leke RGF, Befidi-Mengue R, Tsafack M, Bomba-Nkolo D, et al. The epidemiology of Plasmodium falciparum malaria in two Cameroon villages: Simbok and Etoa. Am J Trop Med Hyg. 2000;63:222–230. [PubMed] [Google Scholar]
- 152.Zhou G, Minakawa N, Githeko A, Yan G. Spatial distribution patterns of malaria vectors and sample size determination in spatially heterogeneous environments: a case study in the West Kenyan highland. J Med Entomol. 2004;41:1001–1009. doi: 10.1603/0022-2585-41.6.1001. [DOI] [PubMed] [Google Scholar]
- 153.Akogbeto M, Di Deco MA, Romano R, Coluzzi M. Analisi comparativa del ruolo vettore per la malaria de Anopheles gambiae e Anopheles melas nella zona lagunare costiera del Benin. Parassitologia. 1988;30:7–8. [Google Scholar]
- 154.Appawu MA, Baffoe-Wilmot A, Afari EA, Nkrumah FK, Petrarca V. Species composition and inversion polymorphism of the Anopheles gambiae complex in some sites of Ghana west Africa. Acta Trop. 1994;56:15–23. doi: 10.1016/0001-706x(94)90036-1. [DOI] [PubMed] [Google Scholar]
- 155.Bockarie MJ, Service MW, Barnish G, Maude GH, Greenwood BM. Malaria in a rural area of Sierra Leone III Vector ecology and disease transmission. Ann Trop Med Parasitol. 1994;88:251–262. doi: 10.1080/00034983.1994.11812865. [DOI] [PubMed] [Google Scholar]
- 156.Coluzzi M, Petrarca V, Di Deco MA. Chromosomal inversion intergradation and incipient speciation in Anopheles gambiae. . Boll Zool. 1985;52:45–63. [Google Scholar]
- 157.Jaenson TGT, Gomes MJ, Barreto dos Santos RC, Petrarca V, Fortini, et al. Control of endophagic Anopheles mosquitoes and human malaria in Guinea Bissau West Africa by permethrin-treated bednets. Trans R Soc Trop Med Hyg. 1994;88:620–624. doi: 10.1016/0035-9203(94)90197-x. [DOI] [PubMed] [Google Scholar]
- 158.Matola YG, White GB, Magayuka SA. The changed pattern of malaria endemicity and transmission at Amani in the eastern Usambara mountains noth-eastern Tanzania. J Trop Med Hyg. 1987;90:127–134. [PubMed] [Google Scholar]
- 159.Petrarca V, Sabatinelli G, Di Deco MA, Papakay M. The Anopheles gambiae complex in the Federal Islamic Republic of Comoros (Indian Ocean): some cytogenetic and biometric data. Parassitologia. 1990;32:371–380. [PubMed] [Google Scholar]
- 160.Robert V, Le Goff G, Toto JC, Mulder L, Fondjo E, et al. Anthrophilic mosquitoes and malaria transmission at Edea Cameroon. Trop Med Parasitol. 1993;44:14–18. [PubMed] [Google Scholar]
- 161.Smits A, Coosemans M, Van Bortel W, Barutwanayo M, Delacollette C. Readjustment of the malaria vector control strategy in the Rusizi Valley, Burundi. Bull Entomol Res. 1995;85:541–548. [Google Scholar]
- 162.Toure YT, Petrarca V, Coluzzi M. Nuove entita del complesso Anopheles gambiae in Mali. Parassitologia. 1983;25:367–370. [Google Scholar]
- 163.Vulule JM, Beach RF, Atieli FK, Roberts JM, Mount DL, et al. Reduced susceptibility of Anopheles gambiae to permethrin associated with the use of permethrin-impregnated bednets and curtains in Kenya. Med Vet Entomol. 1994;8:71–75. doi: 10.1111/j.1365-2915.1994.tb00389.x. [DOI] [PubMed] [Google Scholar]
- 164.Afrane YA, Lawson BW, Githeko AK, Yan G. Effects of microclimatic changes caused by land use and land cover on duration of gonotrophic cycles of Anopheles gambiae (Diptera: Culicidae) in Western Kenya Highlands. J Med Entomol. 2005;42:974–980. doi: 10.1093/jmedent/42.6.974. [DOI] [PubMed] [Google Scholar]
- 165.Awolola TS, Oyewole IO, Amajoh CN, Idowu ET, Ajayi MB, et al. Distribution of the molecular forms of Anopheles gambiae and pyrethroid knock down resistance gene in Nigeria. Acta Trop. 2005;95:204–209. doi: 10.1016/j.actatropica.2005.06.002. [DOI] [PubMed] [Google Scholar]
- 166.Cano J, Angel Desalzo M, Moreno M, Chen Z, Nzambo S, et al. Spatial variability in the density distribution and vectorial capacity of anopheline species in a high transmission village (Equatorial Guinea). Malar J. 2006;5:21. doi: 10.1186/1475-2875-5-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Charalambous M, Townson H, Harbach RE, Mukwaya LG. Electrophoretic and DNA identification of Anopheles bwambae and A. gambiae (Diptera: Culicidae) in western Uganda. Bull Entomol Res. 1999;89:111–117. [Google Scholar]
- 168.Charlwood JD, Pinto J, Sousa CA, Ferreira C, Petrarca V, et al. ‘A mate or a meal’–Pre-gravid behaviour of female Anopheles gambiae from the islands of Sao Tome and Principe West Africa. Malar J. 2003;2:9. doi: 10.1186/1475-2875-2-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.della Toree A, Fanello C, Akogbeto M, Dossou-yovo J, Favia G, et al. Molecular evidence of incipient speciation within Anopheles gambiae s.s.. in West Africa. Insect Mol Biol. 2001;10:9–18. doi: 10.1046/j.1365-2583.2001.00235.x. [DOI] [PubMed] [Google Scholar]
- 170.Lehman T, Licht M, Elissa N, Maega BTA, Chimumbwa JM, et al. Population structure of Anopheles gambiae in Africa. J Hered. 2003;94:133–147. doi: 10.1093/jhered/esg024. [DOI] [PubMed] [Google Scholar]
- 171.Munga S, Minakawa N, Zhou G, Barrack OJ, Githeko AK, et al. Oviposition site preference and egg hatchability of Anopheles gambiae: effects of land cover types. J Med Entomol. 2005;42:993–997. doi: 10.1093/jmedent/42.6.993. [DOI] [PubMed] [Google Scholar]
- 172.Pinto J, Lynd A, Elissa N, Donnelly MJ, Costa C, et al. Co-occurrence of East and West African kdr mutations suggests high levels of resistance to pyrethroid insecticides in Anopheles gambiae from Libreville, Gabon. Med Vet Entomol. 2006;20:27–32. doi: 10.1111/j.1365-2915.2006.00611.x. [DOI] [PubMed] [Google Scholar]
- 173.Lambert D. A population genetical study of the African mosquito Anopheles marshallii (Theobald). Evolution. 1983;37:484–495. doi: 10.1111/j.1558-5646.1983.tb05565.x. [DOI] [PubMed] [Google Scholar]
- 174.Sharp BL. Anopheles merus (Donitz) its biting cycle in relation to environmental parameters. J Entomol Soc South Afr. 1983;46:367–374. [Google Scholar]
- 175.Sharp BL, Quicke FC, Jansen EJ. Aspects of the behavior of 5 anopheline species in the endemic malaria area of Natal South Africa. J Entomol Soc South Afr. 1984;47:251–258. [Google Scholar]
- 176.Gad AM, el Said S, Hassan AN, Shoukry A. The distribution and ecology of the mosquitoes in the Red Sea governorate, Egypt. J Egypt Soc Parasitol. 1987;17:207–221. [PubMed] [Google Scholar]
- 177.Awone-Ambene HP, Kengne P, Simard F, Antonio-Nkondjio C, Fontenille D. Description and bionomics of Anopheles (Cellia) ovengensis (Diptera: Culicidae) a new malaria vector species of the Anopheles nili group from South Cameroon. J Med Entomology. 2004;41:561–568. doi: 10.1603/0022-2585-41.4.561. [DOI] [PubMed] [Google Scholar]
- 178.Kenge P, Awono-Ambene P, Antonio-Nkondjio C, Simard F, Fontenille D. Molecular identification of the Anopheles nili group of African malaria vectors. Med Vet Entomol. 2003;17:67–74. doi: 10.1046/j.1365-2915.2003.00411.x. [DOI] [PubMed] [Google Scholar]
- 179.el Said S, Beier JC, Kenawy MA, Morsy ZS, Merdan AI. Anopheles population dynamics in two malaria endemic villages in Faiyum Governorate, Egypt. J Am Mosq Control Assoc. 1986;2:158–163. [PubMed] [Google Scholar]
- 180.Fettene M, Koekemoer LL, Hunt RH, Coetzee M. PCR assay for identification of Anopheles quadriannulatus species B from Ethiopia and other sibling species of the Anopheles gambiae complex. Med Vet Entomol. 2002;16:214–217. doi: 10.1046/j.1365-2915.2002.00354.x. [DOI] [PubMed] [Google Scholar]
- 181.Bogh C, Clarke SE, Pinder M, Sanyang F, Lindsay SW. Effect of passive zooprophylaxis on malaria transmission in the Gambia. J Med Entomol. 2001;38:822–828. doi: 10.1603/0022-2585-38.6.822. [DOI] [PubMed] [Google Scholar]
- 182.Charlwood JD, Qassim M, Elnsur EI, Donnelly M, Petrarca V, et al. The impact of indoor spraying with malathion on malaria in refugee camps in eastern Sudan. Acta Trop. 2001;80:1–8. doi: 10.1016/s0001-706x(01)00152-8. [DOI] [PubMed] [Google Scholar]
- 183.Fettene M, Hunt RH, Coetzee M, Tessema F. Behaviour of Anopheles arabiensis and An. quadriannulatus sp. B mosquitoes and malaria transmission in southwestern Ethiopia. Aft Entomol. 2004;12:83–87. [Google Scholar]
- 184.Garrett-Jones C, Boreham PFL, Pant CP. Feeding habits of Anophelines (Diptera: Culicidae) in 1971-1978 with reference to the human blood index: a review. Bull Entomol Res. 1980;70:165–185. [Google Scholar]
- 185.Githeko AK, Service MW, Mbogo CM, Atieli FK, Juma FO. Origin of blood meals in indoor and outdoor resting malaria vectors in western Kenya. Acta Trop. 1994;58:307–316. doi: 10.1016/0001-706x(94)90024-8. [DOI] [PubMed] [Google Scholar]
- 186.Habtewold T, Walker AR, Curtis CF, Osir EO, Thapa N. The feeding behavior and Plasmodium infection of Anopheles mosquitoes in southern Ethiopia in relation to use of insecticide-treated livestock for malaria control. Trans R Soc Trop Med Hyg. 2001;95:584–586. doi: 10.1016/s0035-9203(01)90086-0. [DOI] [PubMed] [Google Scholar]
- 187.Hadis M, Lulu M, Makonnen Y, Asfaw T. Host choice by indoor-resting Anopheles arabiensis in Ethiopia. Trans R Soc Trop Med Hyg. 1997;91:376–378. doi: 10.1016/s0035-9203(97)90245-5. [DOI] [PubMed] [Google Scholar]
- 188.Highton RB, Bryan JH, Boreham PFL, Chandler JA. Studies on the sibling species Anopheles gambiae Giles and Anopheles arabiensis Patton (Diptera: Culicidae) in the Ksumu area. Kenya Bull Entomol Res. 1979;69:43–53. [Google Scholar]
- 189.Joshi GP, Service MW, Pradhan GD. A survey of species A and B of the Anopheles gambiae Giles complex in the Kisumu area of Kenya prior to insecticidal spraying with OMS-43 (fenitrothion). Ann Trop Med Parasitol. 1975;69:91–104. doi: 10.1080/00034983.1975.11686988. [DOI] [PubMed] [Google Scholar]
- 190.Lemasson JJ, Fontenille D, Lochouarn L, Dia I, Simard F, et al. Comparison of behavior and vector efficiency of Anopheles gambiae and An. arabiensis (Diptera: Culicidae) in Barkedji a Sahelian area of Senegal. J Med Entomol. 1997;34:396–403. doi: 10.1093/jmedent/34.4.396. [DOI] [PubMed] [Google Scholar]
- 191.Mwangangi JM, Mbogo CM, Nzovu JG, Githure JI, Yan G, et al. Blood-meal analysis for anopheline mosquitoes sampled along the Kenyan coast. J Am Mosq Control. 2003;19:371–375. [PubMed] [Google Scholar]
- 192.Service MW Joshi GP, Pradhan GD. A Survey of Anopheles gambiae (Species A) and Anopheles arabiensis (species B) of the Anopheles gambiae Giles complex in the Kisumu area of Kenya following insecticidal spraying with OMS-43 (Fenitrothion). Ann Trop Med Parasitol. 1978;72:377–386. doi: 10.1080/00034983.1978.11719332. [DOI] [PubMed] [Google Scholar]
- 193.White GB, Rosen P. Comparative studies on sibling species of the Anopheles gambiae Giles complex (Diptera: Culicidae). II. ecology of species A and B in Savanna around Kaduna, Nigeria, during transition from wet to dry season. Bull Entomol Res. 1973;62:613–625. [Google Scholar]
- 194.White GB, Magayuka SA, Boreham PFL. Comparative studies on sibling species of the Anopheles gambiae Giles complex (Diptera: Culicidae): bionomics and vectorial activity of species A and species B at Segara, Tanzania. Bull Entomol Res. 1972;62:295–317. [Google Scholar]
- 195.Beier JC, Perkins PV, Wirtz RA, Koros J, Diggs D, et al. Bloodmeal identification by direct enzyme linked immunosorbent assay (ELISA) tested on Anopheles (Diptera: Culicidae) in Kenya. J Med Entomol. 1988;25:9–16. doi: 10.1093/jmedent/25.1.9. [DOI] [PubMed] [Google Scholar]
- 196.Bogh C, Pedersen EM, Mukoko DA, Ouma JH. Permethrin-impregnated bednet effects on resting and feeding behavior of lymphatic filariasis vector mosquitoes in Kenya. Med Vet Entomol. 1998;12:52–59. doi: 10.1046/j.1365-2915.1998.00091.x. [DOI] [PubMed] [Google Scholar]
- 197.Chandler JA, Boreham PFL, Highton RB, Hill MN. A study of the host selection patterns of the mosquitoes of the Kisumu area of Kenya. Trans R Soc Trop Med Hyg. 1975;69:415–425. doi: 10.1016/0035-9203(75)90200-x. [DOI] [PubMed] [Google Scholar]
- 198.Mbogo CN, Kabiru EW, Muiruri SK, Nzovu JM, Ouma JH, et al. Bloodfeeding behavior of Anopheles gambiae s.l. and Anopheles funestus in Kilifi District, Kenya. J Am Mosq Control Assoc. 1993;9:225–227. [PubMed] [Google Scholar]
- 199.White GB. Blood feeding habits of mosquitoes in the South Pare District of Tanzania ten years after cessation of a dieldrin residual spraying campaign. East Afr Med J. 1971;48:120–134. [PubMed] [Google Scholar]
- 200.Sousa CA, Pinto J, Almeida AP, do Rosario VE, Charlwood JD. Dogs as a favored host choice of Anopheles gambiae sensu stricto (Diptera: Culicidae) of São Tomé West Africa. J Med Entomol. 2001;38:122–125. doi: 10.1603/0022-2585-38.1.122. [DOI] [PubMed] [Google Scholar]