

Abstract
Generation of a wide variety of nucleoside (and deoxynucleoside) triphosphates (NTPs) from their cognate nucleoside diphosphates (NDPs) is of critical importance in virtually every aspect of cellular life. Their function is fulfilled largely by the ubiquitous and potent nucleoside diphosphate kinase (NDK), most commonly using ATP as the donor. Considerable interest is attached to the consequence to a cell in which the NDK activity becomes deficient or overabundant. We have discovered an additional and possibly auxiliary NDK-like activity in the capacity of polyphosphate kinase (PPK) to use inorganic polyphosphate as the donor in place of ATP, thereby converting GDP and other NDPs to NTPs. This reaction was observed with the PPK activity present in crude membrane fractions from Escherichia coli and Pseudomonas aeruginosa as well as with the purified PPK from E. coli; the activity was absent from the membrane fractions obtained from E. coli mutants lacking the ppk gene. The order of substrate specificity for PPK was: ADP > GDP > UDP, CDP; activity with ADP was 2–60 times greater than with GDP, depending on the reaction condition. Although the transfer of a phosphate from polyphosphate to GDP by PPK to produce GTP was the predominant reaction, the enzyme also transferred a pyrophosphate group to GDP to form the linear guanosine 5′ tetraphosphate.
Supplies of the ribo- and deoxyribonucleoside triphosphates (NTPs, dNTPs) are crucial in virtually all biosynthetic and regulatory activities. Some notable examples in eukaryotic systems are apparent in developmental control (1–5), signal transduction (6, 7), and tumor metastasis (8, 9). The source of the various NTPs is the phosphorylation of the nucleoside diphosphates (NDPs) by the ubiquitous nucleoside diphosphate kinase (NDK), generally with ATP as the donor molecule (10). The cellular consequences of varying the level of NDK have been examined recently in Escherichia coli (11). Mutants lacking NDK grew at a normal rate but showed an increased mutation frequency, presumably due to the imbalance of dNTP pools (11). The residual NDK activity to support growth in the mutants was supplied by adenylate kinase (12), an enzyme previously thought to be specific for adenosine nucleotides.
Among the numerous functions observed or considered for inorganic polyphosphate (polyP) are roles as an ATP substitute and as an energy reservoir (13). A linear polymer of hundreds of phosphate linked by high-energy, phosphoanhydride bonds, polyP is made from the terminal phosphate of ATP in a readily reversible reaction, the phosphorylation of ADP by polyP. polyP has been demonstrated to substitute for ATP in the kinase-catalyzed phosphorylation of sugars (14, 15), protein (16), AMP (17), and ADP [by polyP kinase (PPK); ref. 18].
Although PPK of E. coli is specific for ATP as the nucleoside triphosphate in the formation of polyP, this report will show that in the reverse reaction, polyP can serve as a donor not only for ADP but also for GDP and other NDPs, thus fulfilling the function of a NDK. The PPK activity observed in crude extracts of Pseudomonas aeruginosa has properties similar to those of E. coli PPK.
MATERIALS AND METHODS
Reagents and Proteins.
Sources were as follows: [γ-32P]ATP, Amersham; all nonradiolabeled NDPs and NTPs, guanosine 5′-tetraphosphate (ppppG), and bovine serum albumin, Sigma; and polyethylenimine cellulose (PEI)-TLC plates, Merck.
Bacterial Strains.
The parental E. coli MG1655 (wild type) and the null mutant, Δppk ppx::kan were generously provided by M. Cashel (National Institutes of Health). P. aeruginosa PAO1 was furnished by A. Chakrabarty (University of Illinois).
Preparation of Crude Membrane Fractions.
Cells were grown at 37°C on Luria–Bertani (LB) medium and harvested. The cell pellet was resuspended in 100 μl of buffer A (50 mM Tris·Cl, pH 7.5/10% sucrose), frozen in liquid nitrogen, and stored at −80°C. Lysozyme (250 μg/ml) was added to the thawed cell suspension and incubated at 0°C for 30 min. The cells were lysed by exposure to 37°C for 4 min, followed by immediate chilling in ice water and subjected to centrifugation [65,000 rpm in a model TA100-1 centrifuge (Beckman) for 20 min]. The pellet was washed with buffer A and dispersed by sonication in buffer A containing 5 mM MgCl2, 10 μg/ml RNase, and 10 μg/ml DNase.
polyP-Dependent NDK Assay.
The reaction mixture contained a crude membrane fraction or purified PPK (7 × 107 units per mg of protein) in PPK buffer {50 mM Hepes, pH 7.2/4 mM MgCl2/40 mM (NH4)2SO4 unless otherwise indicated/0.1 mM [32P]polyP/1 mM NDP}. [32P]polyP was made as described previously (18). The mixture was incubated at 37°C for the time indicated, and 1-μl samples were spotted onto a PEI-TLC plate, which was developed with 0.75 M or 1.5 M KH2PO4 (pH 3.5). The plate was dried after developing, exposed to film, and then visualized in a PhosphoImager scanner (Molecular Dynamics). The ratio of the image intensity of the NTP spot to the total (polyP plus NTP) was calculated. One unit of activity generated 1 pmol of [32P]NTP per min.
Two-Dimensional TLC.
Identification of ppppG was performed as described by Bochner and Ames (19). The solvent in the first dimension was 0.75 M Tris/0.45 M HCl/0.5 M LiCl, and the solvent in the second dimension contained 43.5 g of NH42SO4, 0.4 g of NH4HSO4, and 4 g of Na2EDTA per 100 ml.
Digestion of ppppG by Yeast Exopolyphosphatase (PPX).
After development of ppppG on PEI-TLC, the spot containing ppppG was scratched from the plate and then incubated with 0.5 ml of 0.25 M NH4HCO3 for 30 min at room temperature. The PEI cellulose was pelleted by centrifugation in a microfuge, the supernatant was removed, and the elution process was repeated. The two supernatant fractions were pooled, concentrated in a lyophilizer overnight, and suspended in water. Yeast PPX (6.7 × 107 units per mg of protein) was used for digestion of ppppG in the PPX buffer (20).
RESULTS
PPK Catalyzes the Synthesis of NTPs by Transfer of a Phosphate Group from polyP to NDPs.
Purified PPK from E. coli can transfer a phosphate group from polyP to all NDPs, including ADP, GDP, CDP, UDP, dADP, dGDP, dCDP, and TDP (Fig. 1). The efficiency of NDP substrates was as follows: ADP, dADP > dGDP, GDP, TDP > UDP, CDP, dCDP. The kinetic properties of PPK for various NDPs is summarized in Table 1. The specificity of PPK for nucleoside diphosphates was reduced in the absence of (NH4)2SO4 and in the presence of 1 mM CaCl2 in the reaction mixture (Table 2). Zn2+ and Cu2+ reduced the total activity, but Co2+ and Mn2+ had no significant effect (data not shown).
Figure 1.

NDK activity of purified E. coli PPK. The reaction mixture (10 μl) containing 29 ng of purified PPK, PPK buffer, 1 mM (d)NDP, and 0.1 mM [32P]polyP was incubated at 37°C for 30 min. The TLC plate was developed with 1.5 M KH2PO4 (pH 3.5) and autoradiographed.
Table 1.
Km and kcat values of E. coli PPK
| NDP | Km, mM | kcat, min−1 | kcat/Km, mM−1·min−1 |
|---|---|---|---|
| ADP | 0.25 | 630 | 2500 |
| GDP | 0.63 | 5.3 | 8.4 |
| CDP | >3 | 10 | <3 |
| UDP | >3 | 7 | <2 |
Table 2.
NDK specific activity of PPK
| NDP* |
E. coli PPK (purified)
|
Membrane
|
||||
|---|---|---|---|---|---|---|
| −(NH4)2SO4 | +Ca2+ | −(NH4)2SO4, +Ca2+ | E. coli | P. aeruginosa | ||
| ADP | 370 | 26 | 25 | 4.3 | 1.3 | 1.7 |
| GDP | 5.7 | 3.4 | 3.9 | 2.0 | 0.032 | 0.23 |
| CDP | 1.7 | 1.2 | 1.8 | 1.3 | 0.007 | 0.021 |
| UDP | 1.7 | 1.4 | 2.1 | 1.3 | 0.006 | 0.027 |
Reaction buffer is based on 50 mM Hepes (pH 7.2), 4 mM MgCl2, and 40 mM (NH4)2SO4 (18). (NH4)2SO4 was removed from the buffer, or 1 mM CaCl2 was added if indicated. All values are in pmol of NTP per min per mg of protein × 10−4.
[32P]polyP (0.1 mM) was added to each NDP substrate. NDPs were at a concentration of 1 mM.
polyP-Dependent NDK Activity in the Crude Membrane Fraction.
Crude membrane fractions from E. coli contain virtually all the PPK activity catalyzing the synthesis of ATP from ADP and polyP (Fig. 2A). GTP synthesis activity from GDP and polyP was also detected in this fraction (Fig. 2A), as were the less efficient phosphate transfers to CDP and UDP (Fig. 2 and Table 2). Inasmuch as no NTP spot was detected in reactions without NDP (Fig. 2A), it seems likely that the phosphate group transfer from polyP to the NDPs was direct. Most significantly, membrane fractions of the ppk ppx deletion mutant showed no polyP-dependent NDK activity (Fig. 2A).
Figure 2.

polyP-dependent NDK activity in the E. coli membrane fraction. The reaction mixture (10 μl) containing the membrane fraction, PPK buffer, 1 mM NDP, and 0.1 mM [32P]polyP was incubated at 37°C for 40 min. E. coli membrane fractions from MG1655 (wild type; 1.6 μg) and Δppk ppx mutant (2.1 μg) were used. The TLC plates were developed with (A) 1.5 M KH2PO4 (pH 3.5) and (B) 0.75 M KH2PO4 (pH 3.5).
PPK Transfers Pyrophosphate Group from polyP to GDP.
Among the products of the PPK reaction of polyP with GDP was a compound more highly phosphorylated than GTP (Fig. 1). The ratio of GTP to the new spot was ≈10:1. By two-dimensional TLC (Fig. 3), the radioactive spots corresponded to the positions of GTP and ppppG. Inasmuch as PPK failed to make ppppG from GTP and polyP (data not shown), it seems likely that a pyrophosphate group was transferred to GDP from polyP. Yeast PPX (20) cleaved ppppG to Pi and GTP (Fig. 4), as it was known to hydrolyze ppppA to Pi and ATP (S.-J. Liu, personal communication). E. coli PPX (21) also degraded ppppG, yet both the yeast and E. coli enzymes share no amino acid homologies (data not shown).
Figure 3.

Autoradiogram of the products of PPK action on [32P]polyP and GDP. Unlabeled GTP, guanosine 5′-diphosphate 3′-diphosphate (ppGpp), and ppppG were visualized by UV (outlined). The two-dimensional TLC analysis is described in Materials and Methods.
Figure 4.
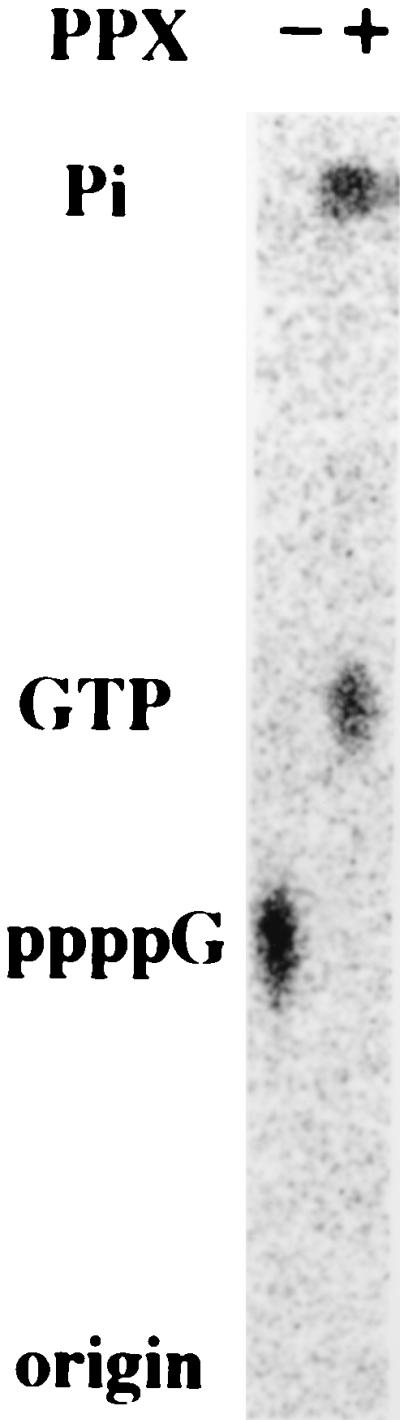
Degradation of ppppG by PPX. α-[32P]ppppG (20 pmol) was digested by yeast PPX (500 units) at 37°C for 60 min. The TLC plate was developed with 1.5 M KH2PO4 (pH 3.5) and autoradiographed.
polyP-Dependent NDK Activity in P. aeruginosa.
Since PPK activity is abundant and membrane attached in P. aeruginosa as in E. coli, it was of immediate interest to determine whether here too the PPK could use NDPs other than ADP as substrates. Membrane fractions of P. aeruginosa PAO1 possessed a polyP-dependent NDP kinase activity (Fig. 5 and Table 2), with a relative specificity for GDP almost 10 times higher than that of E. coli PPK (Table 2). GDP was phosphorylated in the presence of ADP as well as in its absence (Fig. 5). Because of the need for GTP in the major build up of alginate (22) and the concomitant accumulation of polyP at this stage (H.-Y. Kim and A. Kornberg, unpublished data), the capacity of PPK to regenerate GTP from polyP may supply an important function.
Figure 5.

NDK activity of P. aeruginosa membranes. The reaction mixture (10 μl) containing P. aeruginosa membrane fraction (0.73 μg), PPK buffer, 1 mM NDP, and 0.1 mM [32P]polyP was incubated at 37°C for 60 min. The TLC plate was developed with 0.75 M KH2PO4 (pH 3.5) and autoradiographed.
DISCUSSION
In this report, we demonstrate that PPK can transfer a phosphate group from polyP to NDP. Thus PPK may provide an NDK-like activity:
 |
Unlike NDK, the reaction catalyzed by PPK is not reversible, since PPK can synthesize polyP only from ATP. NDP kinase activity of PPK in E. coli lysates is only 0.06–0.3 nmol/min per mg of protein, a value only 0.01% that of NDK (10). Although the contribution of PPK as a NDP kinase may seem minor, the membrane localization of PPK may be significant, since levels of polyP can reach 20 mM in the stationary phase. A mutant of E. coli that lacks NDK grows at a normal rate but shows an elevated mutation rate (11). Tracking the residual NDP kinase activity in the mutant revealed adenylate kinase to have this activity (12). Surprisingly, adenylate kinase can transfer phosphate to an NDP but with 1–5% the specific activity of NDK (12).
Although ammonium sulfate increased the activity of E. coli PPK in polyP synthesis from ATP (18), it alters the preferential order of NDPs as substrates in the NDK reaction. Without ammonium sulfate present, the preference for ADP is markedly reduced (Table 2). Ammonium sulfate increased the kcat value for ADP 20-fold (data not shown), while Ca2+ ion appeared to reduce the preference for ADP. The strong chelation of Ca2+ by polyP and the role of Ca2+-polyP combined with poly-β-hydroxybutyrate in membrane complexes (23) directs more attention to Ca2+ and its influence on PPK activities.
With polyP and GDP as substrates, the predominant reaction catalyzed by PPK was the transfer of a phosphate group to synthesize GTP. About 10% of the time, PPK produced ppppG. Inasmuch as the ppppG was not produced from GTP, we presume that a pyrophosphate was transferred to GDP. A highly phosphorylated compound (likely inosine 5′-tetraphosphate) was also detected in the reaction of IDP and polyP, but nucleoside 5′-tetraphosphate was hardly detected in reactions with ADP, CDP, or UDP. The absence of ppppG among the products of PPK activity in the crude membrane fraction may be due to the presence of PPX, which degrades ppppG. Other known sources of nucleoside 5-tetraphosphates are yeast phosphoglycerate kinase, which forms adenosine 5′-tetraphosphate (24) and ppppG (25) from 1,3-bis-phosphoglycerate, and ATP or GTP. The physiological role of tetraphosphates is unknown. Adenosine, guanosine, and uridine tetraphosphates are strong competitive inhibitors, with nanomolar KI values, of dinucleoside tetraphosphatase (26), which cleaves dinucleoside tetraphosphates to nucleoside tri- and monophosphates. Should diadenosine tetraphosphate prove to have important roles in metabolic regulation, changes in the level of ppppN could modulate its concentration and physiological effects. Another enzyme known to be inhibited by nucleotide 5′-tetraphosphates, with a micromolar KI value, is the soluble guanylate cyclase (27).
The finding of a polyP-dependent NDP kinase activity in P. aeruginosa as well as in E. coli may suggest that this property is a general feature of PPK. The specificity for GDP of the P. aeruginosa PPK was almost 10 times higher than that of E. coli. The need for P. aeruginosa to produce a large amount of alginate dependent on GTP (by way of the alginate precursor, GDP-mannose; ref. 22) and the large accumulations of polyP at this stage of the life cycle suggest that an NDK role for PPK may prove significant.
Acknowledgments
Drs. M. Cashel and A. Chakrabarty generously provided strains. We thank Dr. N. N. Rao for technical help and LeRoy Bertsch for critical reading of this manuscript. A. Kuroda is the recipient of a fellowship from the Human Frontier Science Program. This work was supported by a grant from the National Institutes of Health.
Footnotes
Abbreviations: NDK, nucleoside diphosphate kinase; polyP, inorganic polyphosphate; PPK, polyP kinase; ppppG, guanosine 5′-tetraphosphate; PPX, exopolyphosphatase.
References
- 1.Biggs J, Hersperger E, Steeg P S, Liotta L A, Shearn A. Cell. 1990;63:933–940. doi: 10.1016/0092-8674(90)90496-2. [DOI] [PubMed] [Google Scholar]
- 2.Munoz-Dorado J, Inouye M, Inouye S. J Biol Chem. 1990;265:2702–2706. [PubMed] [Google Scholar]
- 3.Wallet V, Mutzel R, Troll H, Barzu O, Wurster B, Veron M, Lacombe M L. J Natl Cancer Inst. 1990;82:1199–1202. doi: 10.1093/jnci/82.14.1199. [DOI] [PubMed] [Google Scholar]
- 4.Okabe-Kado J, Kasukabe T, Hozumi M, Honma Y, Kimura N, Baba H, Urano T, Shiku H. FEBS Lett. 1995;363:311–315. doi: 10.1016/0014-5793(95)00338-a. [DOI] [PubMed] [Google Scholar]
- 5.Venturelli D, Martinez R, Melotti P, Casella I, Peschle C, Cucco C, Spampinato G, Darzynkiewicz Z, Calabretta B. Proc Natl Acad Sci USA. 1995;92:7435–7439. doi: 10.1073/pnas.92.16.7435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bominaar A A, Molijin A C, Pestel M, Veron M, Van H P J. EMBO J. 1993;12:2275–2279. doi: 10.1002/j.1460-2075.1993.tb05881.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Izumiya H, Yamamoto M. J Biol Chem. 1995;270:27859–27864. doi: 10.1074/jbc.270.46.27859. [DOI] [PubMed] [Google Scholar]
- 8.Steeg P S, Bevilacqua G, Kopper L, Thorgeirsson U P, Talmadge J E, Liotta L A, Sobel M E. J Natl Cancer Inst. 1988;80:200–204. doi: 10.1093/jnci/80.3.200. [DOI] [PubMed] [Google Scholar]
- 9.Rosengard A M, Krutzsch H C, Shearn A, Biggs J R, Barker E, Margulies I M, King C R, Liotta L A, Steeg P S. Nature (London) 1989;342:177–180. doi: 10.1038/342177a0. [DOI] [PubMed] [Google Scholar]
- 10.Parks R E, Agarwal R P. In: The Enzymes. Boyer P D, editor. New York: Academic; 1973. pp. 307–333. [Google Scholar]
- 11.Lu Q, Zhang X, Almaula N, Mathews C K, Inouye M. J Mol Biol. 1995;254:337–341. doi: 10.1006/jmbi.1995.0620. [DOI] [PubMed] [Google Scholar]
- 12.Lu Q, Inouye M. Proc Natl Acad Sci USA. 1996;93:5720–5725. doi: 10.1073/pnas.93.12.5720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kornberg A. J Bacteriol. 1995;177:491–496. doi: 10.1128/jb.177.3.491-496.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kulaev I S. The Biochemistry of Inorganic Polyphosphates. New York: Wiley; 1979. [DOI] [PubMed] [Google Scholar]
- 15.Clark J E, Beegen H, Wood H G. J Bacteriol. 1986;168:1212–1219. doi: 10.1128/jb.168.3.1212-1219.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Skorko R. Biochimie. 1989;71:1089–1093. doi: 10.1016/0300-9084(89)90115-6. [DOI] [PubMed] [Google Scholar]
- 17.Bonting C F, Kortstee G J, Zehnder A J. J Bacteriol. 1991;173:6484–6488. doi: 10.1128/jb.173.20.6484-6488.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ahn K, Kornberg A. J Biol Chem. 1990;265:11734–11739. [PubMed] [Google Scholar]
- 19.Bochner B R, Ames B N. J Biol Chem. 1982;257:9759–9769. [PubMed] [Google Scholar]
- 20.Wurst H, Kornberg A. J Biol Chem. 1994;269:10996–11001. [PubMed] [Google Scholar]
- 21.Akiyama M, Crooke E, Kornberg A. J Biol Chem. 1993;268:633–639. [PubMed] [Google Scholar]
- 22.May T B, Chakrabarty A M. Trends Microbiol. 1994;2:151–157. doi: 10.1016/0966-842x(94)90664-5. [DOI] [PubMed] [Google Scholar]
- 23.Castuma C E, Huang R, Kornberg A, Reusch R N. J Biol Chem. 1995;270:12980–12983. doi: 10.1074/jbc.270.22.12980. [DOI] [PubMed] [Google Scholar]
- 24.Small G D, Cooper C. Biochemistry. 1966;5:26–33. doi: 10.1021/bi00865a004. [DOI] [PubMed] [Google Scholar]
- 25.Garcia-Diaz M, Canales J, Sillero M A, Sillero A, Cameselle J C. Biochem Int. 1989;19:1253–1264. [PubMed] [Google Scholar]
- 26.Moreno A, Lobaton C D, Sillero M A G, Sillero A. Int J Biochem. 1982;14:629–634. doi: 10.1016/0020-711x(82)90047-7. [DOI] [PubMed] [Google Scholar]
- 27.Ignarro L J, Gross R A, Gross D M. J Cyclic Nucleotide Res. 1976;2:337–346. [PubMed] [Google Scholar]