

Abstract
SIRT1 is a mammalian homolog of the Saccharomyces cerevisiae chromatin silencing factor Sir2. Dominant-negative and overexpression studies have implicated a role for SIRT1 in deacetylating the p53 tumor suppressor protein to dampen apoptotic and cellular senescence pathways. To elucidate SIRT1 function in normal cells, we used gene-targeted mutation to generate mice that express either a mutant SIRT1 protein that lacks part of the catalytic domain or has no detectable SIRT1 protein at all. Both types of SIRT1 mutant mice and cells had essentially the same phenotypes. SIRT1 mutant mice were small, and exhibited notable developmental defects of the retina and heart, and only infrequently survived postnatally. Moreover, SIRT1-deficient cells exhibited p53 hyperacetylation after DNA damage and increased ionizing radiation-induced thymocyte apoptosis. In SIRT1-deficient embryonic fibroblasts, however, p53 hyperacetylation after DNA damage was not accompanied by increased p21 protein induction or DNA damage sensitivity. Together, our observations provide direct evidence that endogenous SIRT1 protein regulates p53 acetylation and p53-dependent apoptosis, and show that the function of this enzyme is required for specific developmental processes.
In Saccharomyces cerevisiae, the silent information regulator (Sir) 2 functions as a nicotinamide adenine dinucleotide-dependent histone deacetylase (HDAC) (1), and regulates chromatin silencing (2, 3). Yeast strains with abnormal levels of Sir2 display defects in multiple cellular functions, including transcriptional and recombinational silencing, senescence, and DNA repair (2, 3). Sir2-dependent silencing at the rDNA locus suppresses recombination between rDNA repeats and thereby inhibits recombinational excision of extrachromosomal rDNA circles, the accumulation of which regulates longevity (4). At telomeres, Sir2 is required for establishment and maintenance of telomeric heterochromatin; and, at the mating type loci, Sir2-dependent silencing regulates mating status (2, 3). These functions of Sir2 link it to maintenance of genomic stability by multiple mechanisms. Early studies also suggested that Sir2 functions in DNA repair (5–7). A protein complex containing Sir2 was reported to translocate to DNA double-strand breaks (DSBs) (6, 7). In addition, Sir2-deficient strains showed defects in the nonhomologous end-joining (NHEJ) pathway of DNA DSB repair (5), a process which, in mammals, regulates the lymphocyte-specific variable (diversity) joining [V(D)J] recombination and immunogloblulin heavy-chain class switch recombination (CSR) processes (8, 9). However, more recent evidence indicates that the NHEJ defects in Sir2-deficient yeast are largely secondary to derepression of silent mating type genes, which down-regulate NHEJ (10–12).
In mammalian genomes, seven Sir2 homologs (SIRTs 1–7) have been identified, and could be orthologs of Sir2 with potential roles in regulating chromatin silencing, genomic stability, and cellular or organismal senescence (13, 14). However, relatively little is understood about the substrates and functions of SIRTs. Several SIRTs can deacetylate histones in vitro (1), but none have yet been shown to function as physiologic HDACs. SIRTs 2 and 3 are cytoplasmic proteins. SIRT2 has been shown to have a role in tubulin deacetylation, whereas SIRT3 is a mitochondrial protein with as-yet-unidentified substrates (15–17). Of the seven SIRTs, SIRT1 is the presumed ortholog of Sir2, because it has the most sequence similarity (13). However, in contrast to the critical role Sir2 plays in gene silencing in yeast, studies of a SIRT1-deficient mouse, generated by gene-targeted mutation, failed to find a global defect in gene silencing (18, 19). Instead, the most notable defects in this SIRT1-mutant mouse line were persistent eyelid closure and infertility (18). In addition, depending on the genetic background, SIRT1 deficiency resulted in early postnatal lethality, although the cause of this lethality was unclear (18). In this previous study (18), SIRT1 was found to be widely expressed during early embryogenesis and in adult germ cells, but not in somatic tissues beyond midgestation or the adult mouse.
Several studies have implicated SIRT1 in regulating the tumor suppressor p53 (20–22). In response to DNA damage and other cellular stresses, p53 is stabilized and activated to trigger apoptosis and cell-cycle arrest (23, 24). In addition, p53-dependent apoptosis is also implicated in monitoring spontaneous DNA damage; for example, during neurogenesis (25, 26) and spermatogenesis (27). In the latter context, the infertility of the previously described male SIRT1 mutant mice was attributed to increased p53-dependent spermatocyte apoptosis, although p53 in these mice was not directly analyzed (18). Overexpression and dominant-negative strategies have found that SIRT1 can deacetylate p53 both in vitro and in cell culture overexpression experiments (20–22). Acetylation of p53 occurs on multiple lysine residues, including K320, K373, and K382 in human p53, corresponding to K317, K370, and K379, respectively, in murine p53 (23, 24, 28). Although the specific consequence of p53 acetylation is not known, it correlates with p53 stabilization and activation (23, 24, 28). In this regard, SIRT1 overexpression was shown to inhibit p53 transcriptional activity and p53-dependent apoptosis in response to DNA damage and oxidative stresses, whereas overexpression of a catalytically inactive SIRT1 protein potentiated these cellular stress responses (20, 21).
Although the previous overexpression and dominant-negative studies clearly link SIRT1 function to p53 regulation, they did not completely rule out potential nonphysiologic effects of the overexpressed proteins. Similarly, the phenotypes described in the SIRT1 mutant mouse study (18) could also reflect nonphysiologic or dominant-negative effects, because the SIRT1 mutation consisted of an internal 76-aa deletion predicted to leave behind 90% of the WT SIRT1 protein. To study the function of SIRT1, we have used gene targeting to generate two different SIRT1 mutations, one which expressed a mutant protein lacking part of the catalytic domain, and one which eliminates all protein expression. These different mutations allow us to compare potential dominant-negative effects of mutant SIRT1 protein with complete loss of SIRT1 expression. Our findings provide direct evidence that endogenous SIRT1 protein plays critical and specific roles during development, and regulates acetylation of endogenous p53 protein.
Materials and Methods
Targeting Constructs. The KOII and SKO targeting vectors were constructed in pLNTK (29). The 5′ KOII homology arm is a 2.6-kb SacI/HindIII genomic fragment. The 3′ KOII homology arm is a genomic fragment from the HindIII site upstream of SIRT1 exon 4 extending 3.6 kb (to just upstream of the BglII site), with a loxP site replacing the BglII site immediately downstream of exon 4. The 5′ SKO and 3′SKO homology arms (6-kb HindIII/HindIII and 3.8-kb PacI/PacI genomic fragments, respectively) were amplified from 129/Sv genomic DNA.
Gene Targeting and Generation of Embryonic Stem (ES) Cells. The KOII and SKO targeting vectors were electroporated into TC1 ES cells (30) as described (31). Targeted KOII clones were identified by Southern blotting by using the 5′KOII probe on BamHI-digested genomic DNA (20.6-kb germ line and 16.7-kb targeted), and confirmed with 3′KOII and Neo probes. Targeted KOII clones were infected with AdenoCre to remove the NeoR gene and exon 4. Deleted SIRT1+/Δex4 clones were identified by Southern blotting of BglII-digested genomic DNA with an internal 1.5-kb RI/SalI fragment as probe (3.4-kB germ line, 9.9-kB targeted, and 7.4-kB deleted). Targeted (SIRT1+/Δneo) and Cre-deleted (SIRT1+/Δ) SKO clones were identified by Southern blotting by using the 5′SKO probe on BglI/SwaI-digested genomic DNA (17.4-kB germ line, 12.4-kB targeted, 14.7-kB Cre-deleted), and confirmed with 3′SKO and Neo probes. Independent SIRT1+/Δex4, SIRT1+/Δneo, and SIRT1+/Δ ES cell clones were injected into C57BL/6 blastocysts. Founder chimeras were bred to 129/Sv females and the F1 heterozygotes were intercrossed. SIRT1Δex4/Δex4 ES cells were generated by the high G418-selection method (32).
Analysis of p53 Acetylation. Mouse embryonic fibroblasts (MEFs) isolated from embryonic day (E)12.5 (SIRT1+/Δex4 crosses) or E13.5 (SIRT1+/Δ crosses) embryos were treated with adriamycin (0.2 μg/ml for 8 h) or UV-irradiated (25 J/m2 for 18 h). The HDAC inhibitor trichostatin A (TSA) was added (5 μM) for the last 2 h of culture as indicated. Acetylated p53 was detected by Western analysis of whole-cell extracts or p53 immunoprecipitations (IPs) as described (33, 34). SIRT1Δex4/Δex4 and WT TC1 ES cells were differentiated into fibroblast-like cells with retinoic acid and assayed for p53 acetylation as described (33–35). Antibodies against p53(Ac-317), p53(AcK-379), and p53(Ser-18-P) were described (33, 34). Antibodies against murine p53(Ac-K370) corresponding to human p53(Ac-K373) (Upstate Biotechnology, Lake Placid, NY), SIRT1 (Upstate Biotechnology), tubulin (Sigma), p21 (Ab-4; Oncogene Research Products), and p53 (CM-5; NovoCastra, Newcastle, U.K.) were purchased commercially.
Retroviral Infection. SIRT1 cDNA was cloned into pBabe-puro. Virus was packaged in 293T cells by cotransfection with VSV-G and Gag-Pol expressing plasmids, as described (36). MEFs were infected by incubation with virus and 2 μg/ml polybrene, and 48 h later, were selected in 2.5 μg/ml puromycin.
Thymocyte Apoptosis Assays. Thymocytes (2 × 106 cells) from 2- to 3-week-old mice were irradiated at different doses, cultured for 12 h, and apoptosis assayed as described (37).
Histological Analysis. Mouse tissue was fixed in Bouin's fixative (body) or 4% paraformaldehyde (eye), embedded in paraffin, sectioned at 6 μm, and hematoxylin/eosin staining was performed by standard methods.
Results
Expression of SIRT1 in Mice. To characterize SIRT1 expression patterns, we performed Western analyses with anti-SIRT1 antibody on extracts from a variety of adult murine tissues. In contrast to the results of previous studies (18, 19), these assays revealed readily detectable SIRT1 protein in many adult mouse tissues, including thymus and spleen (Fig. 1A).
Fig. 1.

Generation of SIRT1-deficient mice. (A) Western analysis of SIRT1 and tubulin expression in WT adult mouse tissues. (B) A schematic diagram of the germ-line SIRT1 locus, KOII targeting vector, and SIRT1Δex4 allele. SIRT1 exons comprising the conserved catalytic domain are dark gray. The relative locations of the 5′, 3′, and internal KOII probes are indicated. Restriction sites: BgI, BglI; N, NotI; S, SalI; H, HindIII; E, EcoRI; Bg, BglII; B, BamHI; Pac, PacI; Swa, SwaI. (C) Southern blot of SIRT1+/Δex4, SIRT1+/NeoR, and SIRT1Δex4/Δex4 BglII-digested DNA with the internal KOII probe. (D) Western blot of SIRT1+/+, SIRT1+/Δex4, and SIRT1Δex4/Δex4 MEFs. SIRT1 and SIRT1Δex4 proteins are indicated. (E) A schematic diagram of the germ-line SIRT1 locus, SKO targeting vector, and SIRT1Δ allele, which are depicted as in A. (F) Southern blot of SIRT1+/Δneo and SIRT1+/Δ BglI/SwaI-digested DNA with the 5′SKO probe. (G) Western blot of SIRT1+/+, SIRT1+/Δ, and SIRT1Δ/Δ MEFs with anti-SIRT1 antibodies. Arrows indicate crossreacting bands that control for total protein.
Generation of SIRT1-Deficient Mice. To study the function of SIRT1, we used gene targeting to generate two different SIRT1 mutations, one which expresses a mutant protein, and one which eliminates all protein expression. These different mutations allow us to compare potential dominant negative effects of mutant SIRT1 protein with complete loss of SIRT1 expression.
The targeting vector KOII (Fig. 1B) was designed to conditionally delete SIRT1 exon 4, which encodes 51 amino acids of the conserved SIRT1 catalytic domain. Targeting and Cre-deletion of TC1 ES cells (see Materials and Methods) resulted in cells harboring the SIRT1Δex4 allele; these cells express the expected mutant protein, which migrates slightly faster than WT SIRT1 on Western analysis (Fig. 1D). Because the aberrant SIRT1Δex4 protein might retain partial function or have dominant-negative activity, we also designed a second targeting construct (SKO) to completely abolish SIRT1 protein expression (Fig. 1E). Targeted heterozygous ES cells, either containing a LoxP-pgk-NeoR selection cassette (SIRT1+/Δneo), or with this cassette removed through Cre-deletion (SIRT1+/Δ) (Fig. 1F), were used to generate SIRT1+/Δneo and SIRT1+/Δ mice, respectively. Western analysis with an antibody specific for the SIRT1 N terminus revealed that SIRT1Δ/Δ MEFs expressed no detectable SIRT1 protein (Fig. 1G).
Developmental Defects in SIRT1-Deficient Mice. We intercrossed SIRT1+/Δex4, SIRT1+/Δ, and SIRT1+/Δneo mice, respectively; although we have analyzed larger numbers of offspring from the SIRT1+/Δex4 crosses, we have, where examined, found similar results with all crosses (see below). Consistent with previous reports (18), genotyping of offspring from SIRT1+/Δex4 crosses revealed poor representation of SIRT1Δex4/Δex4 mice, which comprised only 10% of the pups born, 67% of which died within the first week after birth (see Table 1, which is published as supporting information on the PNAS web site, www.pnas.org). In contrast, SIRT1Δex4/Δex4 embryos were present at approximately Mendelian ratios, even at late stages of gestation (E18.5). Thus, the paucity of SIRT1Δex4/Δex4 neonates is likely due to early postnatal lethality.
The SIRT1Δex4/Δex4 mice and embryos were significantly smaller than littermate controls at all stages analyzed, they sometimes exhibited exencephaly, and had dramatically reduced sperm numbers (Fig. 2A and data not shown), which was consistent with findings of a previous study (18). In contrast to the earlier study, we did not observe lung or pancreatic defects, but we frequently observed cardiac defects, in SIRT1Δex4/Δex4 mice. Of five E18.5 SIRT1Δex4/Δex4 embryos analyzed, one had a clear ventricular septal defect (VSD; Fig. 2F), one had an atrial septal defect (ASD; data not shown), and three had abnormally elongated atrioventricular valves and probable VSDs (Fig. 2G). Notably, the few SIRT1Δex4/Δex4 mice that survived to adulthood did not have noticeable cardiac defects (data not shown). Although we have not analyzed hearts of SIRT1Δ/Δ embryos, an E18.5 SIRT1Δneo/Δneo embryo had a clear ASD (data not shown). Thus, our preliminary analyses suggest that SIRT1 likely plays an important role in cardiac septation, and suggests that cardiac defects in SIRT1-deficient mice might contribute to their neonatal lethality.
Fig. 2.

Developmental abnormalities in SIRT1-deficient mice. (A) WT and SIRT1Δex4/Δex4 E16.5 embryos. *, exencephaly in the SIRT1Δex4/Δex4 embryo. (B) Whole-mount eyes from E16.5 embryos. Arrow indicates open optic fissure in the SIRT1Δex4/Δex4 eye. (C) Retinal sections from 5-week-old mice. rs, rosette-like structure. (D) An enlargement of C. gl, ganglion cell layer; ipl, inner plexiform layer; inl, inner nuclear layer; opl, outer plexiform layer; onl, outer nuclear layer; is, inner segment; os, outer segment; pe, pigmented epithelium. (E) Retinal sections from E18.5 WT and SIRT1Δneo/Δneo embryos. (F) Sections through E18.5 hearts. *, the VSD. LV, left ventricle; RV, right ventricle. (G) Section through E18.5 SIRT1Δex4/Δex4 heart showing elongated atrioventricular valve (arrow).
We consistently observed eye abnormalities in SIRT1Δex4/Δex4 mice, which appear to be primary developmental defects, because they were observed in SIRT1Δex4/Δex4 embryos at all stages analyzed, from E12.5 to E18.5 (Fig. 2 B–E). On gross examination, SIRT1Δex4/Δex4 eyes from E16.5 embryos were small, irregularly shaped, and exhibited abnormal closure of the optic fissure (Fig. 2B). By histological analyses of adult eyes, multiple retinal cell layers were significantly thinner in SIRT1Δex4/Δex4 than in SIRT1+/+ eyes, and in some areas, the inner and outer nuclear layers were disorganized, with abnormal rosette-like structures (Fig. 2 C and D). In addition, the inner and outer segments of photoreceptor cells were difficult to discern (Fig. 2D). Similarly, in E13.5 SIRT1Δ/Δ and E18.5 SIRT1Δneo/Δneo embryos, eye morphogenesis was also dramatically perturbed, with multiple retinal involutions (Fig. 2E). These observations indicate that SIRT1 has an important role in eye morphogenesis.
Given the role of Sir2 in recombination in S. cerevisiae, we asked whether V(D)J recombination, which relies on NHEJ (8), or lymphocyte development, is altered by SIRT1-deficiency in the mouse. Analyses of SIRT1Δex4/Δex4 mice revealed, at least at a gross level, normal development of T and B lymphocyte populations as analyzed by expression of stage-specific lymphocyte markers (see Fig. 5, which is published as supporting information on the PNAS web site). In addition, SIRT1Δex4/Δex4 ES cells, generated by the high G418-selection method (32), showed approximately normal ability to generate V(D)J coding and recombination signal joins, as assessed by an extrachromosomal substrate assay (see Table 2, which is published as supporting information on the PNAS web site). Finally, Ig heavy chain CSR, a B lymphocyte-specific recombination process distinct from V(D)J recombination (9), was not grossly affected by SIRT1-deficiency (see Fig. 6, which is published as supporting information on the PNAS web site). Overall, these data suggest that SIRT1 is not required for catalysis of classical NHEJ, V(D)J recombination, or CSR; however, we cannot exclude subtle effects of SIRT1 deficiency on these processes or a role for SIRT1 in the regulation of these processes.
Increased p53 Acetylation in SIRT1-Deficient MEFs. To further elucidate potential roles of SIRT1, we analyzed DNA damage-induced p53 acetylation and function in both SIRT1Δex4/Δex4 and SIRT1Δ/Δ MEFs. SIRT1+/+, SIRT1+/Δex4, and SIRT1Δex4/Δex4 MEFs were mock treated, incubated with adriamycin, or UV irradiated, and levels of acetylated K379 in p53 IPs was determined by using acetylation-specific antibodies (33, 34). Both adriamycin- and UV-irradiation-induced p53 acetylation on K379 were significantly greater in SIRT1Δex4/Δex4 than in SIRT1+/+ and SIRT1+/Δex4 MEFs (Fig. 3A). Total protein was approximately equal in all samples, as assessed by tubulin levels in the IP inputs (Fig. 3A). Notably, acetylation of p53 on K317 and K370 after DNA damage also was greater in SIRT1Δex4/Δex4, than in SIRT1+/+ and SIRT1+/Δex4 MEFs. Thus, SIRT1, directly or indirectly, regulates p53 acetylation, but is not specific to K379, as suggested (20). Similar results were obtained for multiple independent MEF lines and differentiated SIRT1Δex4/Δex4 ES cells (data not shown). SIRT1Δ/Δ MEFs also exhibited increased adriamycin-induced p53 acetylation, compared with SIRT1+/+ and SIRT1+/Δ MEFs (Fig. 3C). This effect was due to SIRT1 loss, because reconstitution with recombinant retroviral SIRT1, but not empty virus control, returned p53 acetylation to levels in WT MEFs (Fig. 3C).
Fig. 3.

p53 hyperacetylation in SIRT1-deficient MEFs after DNA damage. (A) Western analysis of p53 IPs (IP/WB) and direct Western blots (WB) of whole-cell MEF lysates. Cells were incubated with adriamycin, were UV irradiated, and treated with 5 μM TSA as indicated. Blots were probed with antibodies specific for the indicated p53 modifications, total p53, and tubulin. (B) p53 hyperacetylation in the absence of TSA. Western blots of whole-cell extracts were probed as in A. *, a gel artifact in lane 1. Arrow indicates a crossreacting band that controls for total protein levels. (C) p53 acetylation in SIRT1+/+, SIRT1+/Δ, and two different SIRT1Δ/Δ MEF lines, and in SIRT1Δ/Δ MEFs infected with empty virus control (pBabe) or retroviral recombinant SIRT1 (rSIRT1).
We next asked whether the p53 hyperacetylation in SIRT1Δex4/Δex4 MEFs was accompanied by increased levels of p53 protein. Surprisingly, total p53 protein levels in the p53-IP were not increased, and in some cases, were decreased, in SIRT1Δex4/Δex4, as compared with SIRT1+/+ and SIRT1+/Δex4 MEFs (Fig. 3A). Whereas p53 acetylation can interfere with the efficiency of p53 IP (38), we also observed decreased total p53 protein levels in SIRT1Δex4/Δex4 cells when assayed through direct Western blots of whole-cell lysates (Fig. 3A, WB). Notably, phosphorylation of p53 on Serine 18, which accompanies p53 activation, paralleled levels of total p53 in being lower in SIRT1Δex4/Δex4, than in SIRT1+/+ and SIRT1+/Δex4 cells (Fig. 3A).
To maximize detection of acetylated p53, the above experiments were performed in the presence of the HDAC inhibitor TSA. In the absence of TSA, overall levels of detectable acetylated p53 after DNA damage were substantially diminished in MEFs of all genotypes. However, p53 was still clearly more acetylated in SIRT1Δex4/Δex4, than in SIRT1+/+ and SIRT1+/Δex4 MEFs (Fig. 3B), providing evidence that SIRT1 deacetylation of p53 is not fully redundant with HDAC-mediated p53 deacetylation.
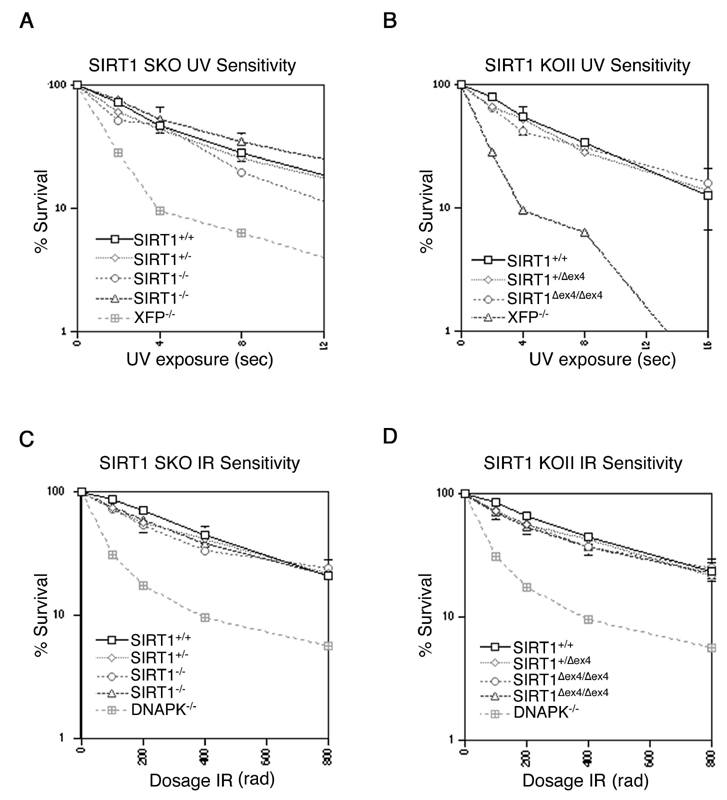
Role of SIRT1 in p53-Dependent Cell-Cycle Arrest. Previous studies reported that p53 hyperacetylation in response to overexpression of a dominant-negative SIRT1 protein was accompanied by increased induction of the p53 response gene p21 (20, 21). In contrast, we found that adriamycin-induced p21 protein levels were not higher in SIRT1Δ/Δ and SIRT1Δex4/Δex4 MEFs than in the corresponding WT and heterozygous MEFs (Fig. 3C and data not shown). In addition, we did not observe substantial differences in UV or ionizing radiation (IR) sensitivity between mutant and WT lines (see Fig. 7, which is published as supporting information on the PNAS web site).
Increased IR-Induced Apoptosis in SIRT1-Deficient Thymocytes. IR-induced apoptosis in CD4+CD8+ mouse thymocytes, which comprise the majority of cells in the thymus, requires p53 (37, 39). Therefore, we asked whether this process was affected by SIRT1 deficiency. We first confirmed that SIRT1 is expressed at significant levels in purified CD4+ and CD4+CD8+ thymocytes, and that the SIRT1Δex4 mutant protein is expressed (at lower levels) in SIRT1Δex4/Δex4 thymocytes (Fig. 4A). Percentages of CD4+, CD8+, and CD4+CD8+ thymocytes were normal in SIRT1-deficient mice (Fig. 5). However, after IR, levels of p53 acetylation were significantly higher in SIRT1Δex4/Δex4, than in SIRT1+/+ thymocytes, whereas total p53 levels were approximately equal (Fig. 4B). Furthermore, SIRT1Δex4/Δex4 thymocytes showed increased IR sensitivity, compared with SIRT1+/+ or SIRT1+/Δex4 thymocytes (Fig. 4C; P < 0.05 at all doses). As a control for p53 dependence, thymocytes from p53–/– mice did not undergo significant IR-induced apoptosis under the same conditions (Fig. 4C). Therefore, deacetylation of p53 by SIRT1 protein appears to contribute to the modulation of p53-dependent thymocyte apoptosis in response to IR.
Fig. 4.

Increased thymocyte apoptosis in SIRT1-deficient cells. (A) Western analysis of SIRT1 and SIRT1Δex4 protein in thymocytes of the indicated genotypes. (B) Western analysis of acetylated p53 in p53 IPs from SIRT1+/+ and SIRT1Δex4/Δex4 thymocytes, at 2.5 h after mock (–) or 500 rads IR (+), in the presence of 1 μM TSA. Western analyses on the IP inputs show total p53 levels, and the arrow indicates a crossreacting band that controls for total protein. (C) The percent nonapoptotic cells in SIRT1+/+, SIRT1+/Δex4, SIRT1Δex4/Δex4, and p53–/– CD4+ thymocytes after IR is shown.
Discussion
We have examined the effects of elimination of SIRT1 expression or expression, in place of SIRT1, of a mutant SIRT1 protein lacking critical catalytic residues on mouse development and p53 function. In all cases analyzed, we observed no significant differences in phenotype between the two different mutations. Whereas many of our observations confirm findings of previous overexpression or gene-targeted mutation studies, certain of our results significantly extend or differ from those of earlier reports. In particular, we find a specific, and not previously described, role for SIRT1 in several different developmental processes. In addition, we provide direct evidence that endogenous SIRT1 protein regulates p53 acetylation, and describe some unexpected effects of p53 hyperacetylation in the absence of SIRT1. Finally, we find that SIRT1 is expressed in adult tissues, where it could well be involved in p53 responses and other processes.
Developmental Defects in SIRT1-Deficient Mice. We have found that SIRT1 deficiency is associated with failure to thrive, abnormal retinal histology with rosette formation, sporadic exencephaly, and cardiac septal and valvular abnormalities. These phenotypes overlap with those of a previous SIRT1 mutational study (18), but they also significantly differ in several respects. Thus, we did not observe the reported (18) lung or pancreatic defects in our lines of SIRT1-mutant or SIRT1-deficient mice. In this context, it is possible that the lung defects observed in the other study were secondary to undetected cardiac abnormalities. We further note that the cardiac defects, detected in our SIRT1-mutant mice, but not in the earlier study, could contribute to neonatal lethality.
Eye defects in postnatal SIRT1 mutant mice were also observed in an earlier study; however, these were concluded to be secondary to persistent eyelid closure, rather than to developmental defects (18). In contrast, our data suggest SIRT1 functions in eye morphogenesis and retinal development, because defects were observed in embryos as early as E12.5. Moreover, the persistence of the optic fissure and disorganized retina morphogenesis are similar to the phenotype observed in mice with targeted deletion of the homeobox gene Vax2 (40); thus, it is conceivable that SIRT1 may deacetylate a factor that functions in this pathway. In addition, the SIRT1-deficient retinal phenotypes (decreased cellularity of multiple retinal layers and abnormal rosette-like structures) have been associated with abnormal proliferation during retinal cell differentiation (41–43), and it is possible that SIRT1 modulates such pathways. Notably, SIRT1 was reported to interact with the transcriptional repressors Hes1 and Hey2 (44). In this regard, mice lacking Hes1 exhibit exencephaly and retinal defects with rosette structures, whereas Hey2-deficient mice develop VSDs and show failure to thrive (41, 45–47). The similarities between these phenotypes and those of our SIRT1-mutant mice raise the possibility that the retinal and cardiac defects observed might result from interference with the Hes1/Hey2 pathways. Alternatively, the developmental abnormalities could reflect dysregulated p53 functions or defects in yet to be determined processes during development.
Functional Consequences of p53 Hyperacetylation in SIRT1-Deficient Cells. We have demonstrated a physiologic role for SIRT1 in regulating p53 acetylation. Previous in vitro studies suggested that SIRT1 specifically deacetylates K382 of human p53 (20), whereas our current studies provide evidence that multiple lysine residues of p53 are hyperacetylated in the absence of functional SIRT1. One interpretation of this observation is that SIRT1 substrate specificity is broader than previously presumed from in vitro deacetylation assays. Alternatively, SIRT1 loss might influence acetylation of some or all of these p53 residues indirectly. Acetylation of p53 is thought to augment its biologic activity (23, 24, 28). We have found that p53 hyperacetylation after DNA damage is accompanied by a significant increase in p53-dependent apoptosis in SIRT1-deficient thymocytes, which was not necessarily expected, because a previous study (18) failed to detect SIRT1 protein in adult somatic tissues. However, we find that SIRT1 protein is detectable in most adult somatic tissues, including thymocytes. The differences in the two studies might be due to sensitivity of different anti-SIRT1 antibodies. Our observation that SIRT1 is expressed in adult somatic tissues is significant, because it leaves open the possibility that SIRT1 could subserve some of the highly anticipated functions for a mammalian Sir2 ortholog in modulating aging, genomic instability, or tumorigenesis.
In MEFs, activation of p53 by DNA damage leads to induction of the cell-cycle inhibitor p21 (48–50). In SIRT1-deficient MEFs, however, p53 hyperacetylation after DNA damage was not accompanied by increased levels of p21 protein induction, or markedly increased sensitivity to DNA damage agents, which was in contrast to the findings of overexpression studies (20, 21). We note, however, that we cannot exclude potential abnormalities in regulation of other p53 response genes in the SIRT1-deficient MEFs. With regard to activation of the p21 pathway, previous studies (51) have shown that acetylation of the p53-family member p73 is selectively coupled to proapoptotic versus cell-cycle arrest pathways, and it is conceivable that the same could be true for p53 acetylation. Alternatively, it is possible that, in addition to deacetylating p53, SIRT1 could regulate other pathways, which could counter p53 effects on p21 induction, either at the protein or RNA level. Thus, we cannot exclude that p53-dependent transactivation of the p21 promoter is abnormal in the absence of SIRT1, and that this abnormality is masked by other as-yet-uncharacterized effects of SIRT1 deficiency. Our observations also indicate that levels of total p53 after DNA damage are not greater in SIRT1-deficient cells than in WT cells. In the latter context, it is possible that net p53 activity in SIRT1-deficient cells correlates better with total p53 levels than acetylation levels. Regardless, our findings suggest that loss of SIRT1 differs from inhibition of HDAC1 activity, with the latter resulting in increased p53 stability (52). We conclude that endogenous SIRT1 protein regulates p53 acetylation, and that p53 hyperacetylation can be uncoupled from p53 stabilization. In the latter context, p53 stability is modulated by diverse factors (23, 24), and such factors could, themselves, be SIRT1 substrates.
Supplementary Material
Acknowledgments
We thank Leonard Guarente, Danesh Moazed, and Yang Xu for critical review of the manuscript; Nicole Stokes, Caitlin Kennedy, JoAnn Sekiguchi, and Wendy Pang for technical assistance; David Lombard, Or Gozani, Connie Cepko, Seohee Cho, and Lenny Guarente for helpful discussions; and Lenny Guarente for the SIRT1 cDNA. K.F.C. is a Pfizer Postdoctoral Fellow in Immunology/Rheumatology. R.M. is a Long-term Fellow of the Human Frontier Science Program. F.W.A. is an Investigator of the Howard Hughes Medical Institute.
Abbreviations: Sir, silent information regulator; SIRT, homolog of Sir2; NHEJ, nonhomologous end joining; ES, embryonic stem; En, embryonic day n; MEF, mouse embryonic fibroblast; HDAC, histone deacetylase; TSA, trichostatin A; IR, ionizing radiation; IP, immunoprecipitation.
References
- 1.Imai, S., Armstrong, C. M., Kaeberlein, M. & Guarente, L. (2000) Nature 403, 795–800. [DOI] [PubMed] [Google Scholar]
- 2.Denu, J. M. (2003) Trends Biochem. Sci. 28, 41–48. [DOI] [PubMed] [Google Scholar]
- 3.Gasser, S. M. & Cockell, M. M. (2001) Gene 279, 1–16. [DOI] [PubMed] [Google Scholar]
- 4.Guarente, L. (2000) Genes Dev. 14, 1021–1026. [PubMed] [Google Scholar]
- 5.Tsukamoto, Y., Kato, J. & Ikeda, H. (1997) Nature 388, 900–903. [DOI] [PubMed] [Google Scholar]
- 6.Mills, K. D., Sinclair, D. A. & Guarente, L. (1999) Cell 97, 609–620. [DOI] [PubMed] [Google Scholar]
- 7.McAinsh, A. D., Scott-Drew, S., Murray, J. A. & Jackson, S. P. (1999) Curr. Biol. 9, 963–966. [DOI] [PubMed] [Google Scholar]
- 8.Bassing, C. H., Swat, W. & Alt, F. W. (2002) Cell 109, Suppl., S45–S55. [DOI] [PubMed] [Google Scholar]
- 9.Manis, J. P., Tian, M. & Alt, F. W. (2002) Trends Immunol. 23, 31–39. [DOI] [PubMed] [Google Scholar]
- 10.Frank-Vaillant, M. & Marcand, S. (2001) Genes Dev. 15, 3005–3012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kegel, A., Sjostrand, J. O. & Astrom, S. U. (2001) Curr. Biol. 11, 1611–1617. [DOI] [PubMed] [Google Scholar]
- 12.Valencia, M., Bentele, M., Vaze, M. B., Herrmann, G., Kraus, E., Lee, S. E., Schar, P. & Haber, J. E. (2001) Nature 414, 666–669. [DOI] [PubMed] [Google Scholar]
- 13.Frye, R. A. (2000) Biochem. Biophys. Res. Commun. 273, 793–798. [DOI] [PubMed] [Google Scholar]
- 14.Frye, R. A. (1999) Biochem. Biophys. Res. Commun. 260, 273–279. [DOI] [PubMed] [Google Scholar]
- 15.North, B. J., Marshall, B. L., Borra, M. T., Denu, J. M. & Verdin, E. (2003) Mol. Cell 11, 437–444. [DOI] [PubMed] [Google Scholar]
- 16.Onyango, P., Celic, I., McCaffery, J. M., Boeke, J. D. & Feinberg, A. P. (2002) Proc. Natl. Acad. Sci. USA 99, 13653–13658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schwer, B., North, B. J., Frye, R. A., Ott, M. & Verdin, E. (2002) J. Cell Biol. 158, 647–657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.McBurney, M. W., Yang, X., Jardine, K., Hixon, M., Boekelheide, K., Webb, J. R., Lansdorp, P. M. & Lemieux, M. (2003) Mol. Cell. Biol. 23, 38–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.McBurney, M. W., Yang, X., Jardine, K., Bieman, M., Th'ng, J. & Lemieux, M. (2003) Mol. Cancer Res. 1, 402–409. [PubMed] [Google Scholar]
- 20.Vaziri, H., Dessain, S. K., Ng Eaton, E., Imai, S. I., Frye, R. A., Pandita, T. K., Guarente, L. & Weinberg, R. A. (2001) Cell 107, 149–159. [DOI] [PubMed] [Google Scholar]
- 21.Luo, J., Nikolaev, A. Y., Imai, S., Chen, D., Su, F., Shiloh, A., Guarente, L. & Gu, W. (2001) Cell 107, 137–148. [DOI] [PubMed] [Google Scholar]
- 22.Langley, E., Pearson, M., Faretta, M., Bauer, U. M., Frye, R. A., Minucci, S., Pelicci, P. G. & Kouzarides, T. (2002) EMBO J. 21, 2383–2396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Appella, E. & Anderson, C. W. (2001) Eur. J. Biochem. 268, 2764–2772. [DOI] [PubMed] [Google Scholar]
- 24.Brooks, C. L. & Gu, W. (2003) Curr. Opin. Cell Biol. 15, 164–171. [DOI] [PubMed] [Google Scholar]
- 25.Frank, K. M., Sharpless, N. E., Gao, Y., Sekiguchi, J. M., Ferguson, D. O., Zhu, C., Manis, J. P., Horner, J., DePinho, R. A. & Alt, F. W. (2000) Mol. Cell 5, 993–1002. [DOI] [PubMed] [Google Scholar]
- 26.Gao, Y., Ferguson, D. O., Xie, W., Manis, J. P., Sekiguchi, J., Frank, K. M., Chaudhuri, J., Horner, J., DePinho, R. A. & Alt, F. W. (2000) Nature 404, 897–900. [DOI] [PubMed] [Google Scholar]
- 27.Beumer, T. L., Roepers-Gajadien, H. L., Gademan, I. S., van Buul, P. P., Gil-Gomez, G., Rutgers, D. H. & de Rooij, D. G. (1998) Cell Death Differ. 5, 669–677. [DOI] [PubMed] [Google Scholar]
- 28.Prives, C. & Manley, J. L. (2001) Cell 107, 815–818. [DOI] [PubMed] [Google Scholar]
- 29.Gorman, J., van der Stoep, N., Monroe, R., Cogne, M., Davidson, L. & Alt, F. (1996) Immunity 5, 241–252. [DOI] [PubMed] [Google Scholar]
- 30.Deng, C., Wynshaw-Boris, A., Zhou, F., Kuo, A. & Leder, P. (1996) Cell 84, 911–921. [DOI] [PubMed] [Google Scholar]
- 31.Sleckman, B. P., Bardon, C. G., Ferrini, R., Davidson, L. & Alt, F. W. (1997) Immunity 7, 505–515. [DOI] [PubMed] [Google Scholar]
- 32.Mortensen, R. M., Conner, D. A., Chao, S., Geisterfer-Lowrance, A. A. & Seidman, J. G. (1992) Mol. Cell. Biol. 12, 2391–2395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chao, C., Saito, S., Anderson, C. W., Appella, E. & Xu, Y. (2000) Proc. Natl. Acad. Sci. USA 97, 11936–11941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chao, C., Saito, S., Kang, J., Anderson, C. W., Appella, E. & Xu, Y. (2000) EMBO J. 19, 4967–4975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sakaguchi, K., Herrera, J. E., Saito, S., Miki, T., Bustin, M., Vassilev, A., Anderson, C. W. & Appella, E. (1998) Genes Dev. 12, 2831–2841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ory, D. S., Neugeboren, B. A. & Mulligan, R. C. (1996) Proc. Natl. Acad. Sci. USA 93, 11400–11406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lowe, S. W., Schmitt, E. M., Smith, S. W., Osborne, B. A. & Jacks, T. (1993) Nature 362, 847–849. [DOI] [PubMed] [Google Scholar]
- 38.Li, M., Luo, J., Brooks, C. L. & Gu, W. (2002) J. Biol. Chem. 277, 50607–50611. [DOI] [PubMed] [Google Scholar]
- 39.Clarke, A. R., Purdie, C. A., Harrison, D. J., Morris, R. G., Bird, C. C., Hooper, M. L. & Wyllie, A. H. (1993) Nature 362, 849–852. [DOI] [PubMed] [Google Scholar]
- 40.Barbieri, A. M., Broccoli, V., Bovolenta, P., Alfano, G., Marchitiello, A., Mocchetti, C., Crippa, L., Bulfone, A., Marigo, V., Ballabio, A. & Banfi, S. (2002) Development (Cambridge, U.K.) 129, 805–813. [DOI] [PubMed] [Google Scholar]
- 41.Tomita, K., Ishibashi, M., Nakahara, K., Ang, S. L., Nakanishi, S., Guillemot, F. & Kageyama, R. (1996) Neuron 16, 723–734. [DOI] [PubMed] [Google Scholar]
- 42.Robanus-Maandag, E., Dekker, M., van der Valk, M., Carrozza, M. L., Jeanny, J. C., Dannenberg, J. H., Berns, A. & te Riele, H. (1998) Genes Dev. 12, 1599–1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lin, S. C., Skapek, S. X., Papermaster, D. S., Hankin, M. & Lee, E. Y. (2001) Oncogene 20, 7073–7084. [DOI] [PubMed] [Google Scholar]
- 44.Takata, T. & Ishikawa, F. (2003) Biochem. Biophys. Res. Commun. 301, 250–257. [DOI] [PubMed] [Google Scholar]
- 45.Donovan, J., Kordylewska, A., Jan, Y. N. & Utset, M. F. (2002) Curr. Biol. 12, 1605–1610. [DOI] [PubMed] [Google Scholar]
- 46.Gessler, M., Knobeloch, K. P., Helisch, A., Amann, K., Schumacher, N., Rohde, E., Fischer, A. & Leimeister, C. (2002) Curr. Biol. 12, 1601–1604. [DOI] [PubMed] [Google Scholar]
- 47.Sakata, Y., Kamei, C. N., Nakagami, H., Bronson, R., Liao, J. K. & Chin, M. T. (2002) Proc. Natl. Acad. Sci. USA 99, 16197–16202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sage, J., Mulligan, G. J., Attardi, L. D., Miller, A., Chen, S., Williams, B., Theodorou, E. & Jacks, T. (2000) Genes Dev. 14, 3037–3050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kastan, M. B., Onyekwere, O., Sidransky, D., Vogelstein, B. & Craig, R. W. (1991) Cancer Res. 51, 6304–6311. [PubMed] [Google Scholar]
- 50.Brugarolas, J., Moberg, K., Boyd, S. D., Taya, Y., Jacks, T. & Lees, J. A. (1999) Proc. Natl. Acad. Sci. USA 96, 1002–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Costanzo, A., Merlo, P., Pediconi, N., Fulco, M., Sartorelli, V., Cole, P. A., Fontemaggi, G., Fanciulli, M., Schiltz, L., Blandino, G., et al. (2002) Mol. Cell 9, 175–186. [DOI] [PubMed] [Google Scholar]
- 52.Ito, A., Kawaguchi, Y., Lai, C. H., Kovacs, J. J., Higashimoto, Y., Appella, E. & Yao, T. P. (2002) EMBO J. 21, 6236–6245. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}