

Abstract
Chondrocytes exposed to nitric oxide (NO) or antibody to Fas undergo cell death by apoptosis. This study examines structural and functional properties of chondrocyte-derived apoptotic bodies. In NO treated cartilage, the dense pericellular matrix that normally surrounds the cells is degraded and apoptotic bodies accumulate within and in the vicinity of the chondrocyte lacunae. Functional analysis shows that apoptotic bodies isolated from NO-treated chondrocytes or cartilage produce pyrophosphate. The levels of pyrophosphate produced by apoptotic bodies are increased by pretreatment of the chondrocytes with transforming growth factor β and decreased by interleukin 1. Apoptotic bodies contain alkaline phosphatase and NTP pyrophosphohydrolase activities and can precipitate calcium. These results suggest that chondrocyte-derived apoptotic bodies express functional properties that may contribute to the pathologic cartilage calcification observed in aging and osteoarthritis.
Cartilage has a high frequency of age-related pathology, and osteoarthritis represents one of the most prevalent diseases in older individuals (1). Among age-related changes in cartilage are a reduction in tissue cellularity and abnormal matrix calcification or crystal deposition (2, 3). Careful studies have shown evidence for a decrease in cartilage cellular density in femoral condyles from autopsy specimens of older donors by approximately 50%. The number of empty lacunae increases with aging, and there is a correlation between the age-related decrease in cell number and the increased frequency of cartilage fibrillation (4, 5). Apparently normal cartilage in osteoarthritic joints is also hypocellular as compared with nonosteoarthritic samples. Collectively, these observations suggest a predisposition to matrix degeneration and possibly osteoarthritis as a consequence of a reduction in cartilage cellularity.
Cell death is part of tissue homeostasis and repair to tissue injury. Apoptosis represents a physiologic form of cell death that results in removal of apoptotic bodies by phagocytic cells and thus prevents the induction of an inflammatory response by the release of cell contents as observed during necrotic cell death (6–8). Articular cartilage has several features that impact on the fate of apoptotic bodies. This tissue is not vascularized and does not contain mononuclear phagocytes. Furthermore, chondrocytes are anchored in the extracellular matrix and are surrounded by a pericellular matrix. Thus there is no apparent mechanism for the clearance of apoptotic bodies.
Chondrocytes undergo apoptosis after exposure to nitric oxide (9) or after treatment with agonistic antibody to Fas (10). Electron microscopy of apoptotic chondrocytes (9) suggested a similarity between apoptotic bodies and matrix vesicles. In the late 1960s, Anderson and Bonucci identified matrix vesicles in epiphyseal growth plates by electron microscopy (11, 12). Matrix vesicles were described as cell-derived, membrane-enclosed units that are associated with hydroxyapatite deposition. It was also suggested that matrix vesicles may derive from budding or disintegrating cells in the upper portion of the epiphyseal plate. Numerous studies have demonstrated the presence of matrix vesicles in the areas of epiphyseal cartilage that eventually will calcify. The role of matrix vesicles in the initial calcification of bone and cartilage has been confirmed in different experimental models, and studies on their biochemical composition have revealed a large number of similarities with cytoplasmic and cell membrane components of cells from which they derive (13–15). Alkaline phosphatase activity is abundant in matrix vesicles and is used as a marker for their identification. Matrix vesicles also contain pyrophosphate-generating nucleoside triphosphate pyrophosphohydrolase (NTPPH) activities. Matrix vesicles can be isolated from collagenase-digested articular cartilage and separated from chondrocytes by differential centrifugation and used for functional, biochemical, and ultrastructural studies (13). Functionally, isolated matrix vesicles incorporate 45Ca, hydrolyze ATP or other nucleotide triphosphates, and form orthophosphate. Matrix vesicles thus are able to raise the Ca × P product above solubility and promote precipitation of crystalline calcium phosphates (16). Matrix vesicles are found in articular cartilage as well as growth plate cartilage (17). Although matrix vesicles isolated from different sources are heterogeneous (18), they are similar with respect to the capacity to mineralize matrix (19, 20).
On the basis of this information the present study examined the hypothesis that expression of matrix vesicle-like activities in apoptotic bodies may contribute to the abnormal calcification of articular cartilage in aging and osteoarthritis.
METHODS
Chondrocyte Isolation and Culture.
Cartilage was obtained at autopsy or from the University of California at San Diego tissue bank from donors (ages 27–64; gender: 37% female, 63% male) without history of joint disease. Cartilage slices were removed from the femoral condyles and washed in DMEM. Tissues were then minced with a scalpel, transferred into a digestion buffer containing DMEM, 5% FBS, l-glutamine, antibiotics, and 2 mg/ml clostridial collagenase type IV (Sigma), and incubated on a gyratory shaker at 37°C until the fragments were digested (8–24 h). Residual multicellular aggregates were removed by sedimentation (1 × g), and the cells were washed three times in DMEM 5% FBS before use. All experiments reported here were performed with primary or first-passage cells. Cells were used in the experiments after culture for at least 1 week after collagenase treatment.
Induction of Apoptosis and Isolation of Apoptotic Bodies.
Chondrocyte monolayer cultures were treated with sodium nitroprusside (SNP) at 1 mM or antibody to the Fas antigen (anti-Fas IgM antibody CH11 from Oncor) at 1 μg/ml for 24 h. To document and quantify induction of apoptosis, aliquots of the cultures were harvested, centrifuged onto glass slides, fixed with 0.25% glutaraldehyde for 1 min, washed with water, and stained with 1 μg/ml 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) for 10 min at 37°C (21). Slides were then washed with water, air dried, and covered with Vectashield (Vector Laboratories) for analysis by fluorescence microscopy using a triple-band filter set (model 61000; Chroma Technology, Brattleboro, VT) on an Olympus microscope.
For isolation of apoptotic bodies the cultures were harvested (media combined with the residual cell layer). Cells were removed by a 10,000 × g centrifugation for 20 min, and the subcellular particles in the supernatant from the first centrifugation were pelleted by ultracentrifugation (150,000 × g for 1 h at 4°C). The protein content in the pellets was determined with the Bio-Rad DC protein assay, based on the Bradford dye-binding procedure. For isolation of apoptotic bodies or matrix vesicles from cartilage, the tissue was digested with collagenase and this digest was subjected to the two-step centrifugation as described for the cell cultures.
Electron Microscopy.
Fixation of the samples was for 1 h at room temperature in 2.5% glutaraldehyde buffered with 0.1 M cacodylate (pH 7.2). The samples were then rinsed in cacodylate buffer, postfixed 1 h in 2% OsO4 buffered with cacodylate, dehydrated in a graded ethanol series, and embedded in Polybed 812 (Polysciences). Thin sections of cell pellets were stained with uranyl acetate and lead citrate. Pellets from ultracentrifugation of cell-free supernatants containing matrix vesicles or apoptotic bodies were fixed and stained according to the same protocol. For examination of cartilage tissue, pieces of untreated or SNP-treated cartilage were fixed and processed for electron microscopy as described above.
PPi Assay.
PPi levels were measured by a radiometric method (22). The method employs a high specific activity UDP-(3H) glucose (Amersham), which is separated from the reaction product 6-phospho-(3H) gluconate by selective adsorption on charcoal activated with phosphoric and sulfuric acids (Sigma). NADP, UDP-glucose, and uridine-5′-diphosphoglucose pyrophosphorylase were products of Sigma, and the phosphoglucomutase and glucose-6-phosphate dehydrogenase were purchased from Boehringer Mannheim. Sodium pyrophosphate used to prepare the standard was purchased from Fisher Scientific. The standard concentrations ranged from 100 to 400 pmol PPi and were included in each assay. To monitor recovery, PPi was added to the samples; recoveries ranged from 95 to 105% with a mean of 96%. Each sample was run in duplicate or triplicate. After adsorption of the reaction mixture on charcoal, and centrifugation at 14,000 rpm for 10 min, 100 μl of the supernatant was carefully removed and counted for radioactivity in 5 ml of Bio-Safe II (Research Products International). Results are expressed as nanomoles PPi in media.
NTPPH and Alkaline Phosphatase Assays.
The 5′-nucleotide phosphodiesterase I assay for NTPPH was performed with 1 mM p-nitrophenyl-thymidine 5′-monophosphate (Sigma) as substrate in 50 mM Hepes-buffered DMEM containing 1.6 mM MgCl2, pH 8.0, in a volume of 0.5 ml to which 0.05 ml of sample was added for 1 h at 37°C (23). The assay was halted by addition of 4 volumes of 100 mM NaOH, and absorbance at OD at 405 nm was determined. Specific enzyme activity was measured against a standard curve of p-nitrophenol and expressed in units (1 unit was equivalent to 1 nanomole of substrate hydrolyzed per hour per μg of protein). Alkaline phosphatase was assayed as described by Oyajobi et al. (24).
Calcium Deposition.
Matrix vesicles or apoptotic bodies (25 μl containing 40 μg protein) were added to 500 μl of calcifying medium [2.2 mM CaCl2 (or 50,000 cpm of 45Ca purchased from Amersham)/1.6 mM KH2PO4/1 mM ATP disodium salt/1 mM MgCl2/85 mM NaCl/15 mM KCl/10 mM NaHCO3/50 mM N-Tris (hydroxymethyl) methyl-2-aminoethanesulfonic acid, pH 7.6/1% penicillin/streptomycin], vortexed and incubated at 37°C for 24 and 48 h. The samples were centrifuged at 10,000 × g for 10 min in a cold microfuge. The supernatants were removed, and the pellets were washed twice in cold calcifying solution without ATP. 45Ca contained in the mineral phase was solubilized in concentrated HCl and counted in 5 ml of scintillation liquid. Results are expressed as percentage of total 45Ca that precipitated. Assay controls included the omission of ATP, incubation at 4°C as opposed to 37°C, and heat inactivation (56°C for 30 min) of the matrix vesicles or apoptotic bodies.
Reagents.
Human recombinant interleukin 1 (IL-1β) and transforming growth factor β1 (TGF-β1) were purchased from R & D Systems. Tissue culture medium was obtained from BioWhittaker, and SNP (sodium nitroprusside) was obtained from Alexis Corporation, San Diego.
Statistical Analysis.
Results are expressed as mean ± SEM. For all results shown, at least two separate experiments with cells from different donors were performed. Within experiments, each individual measurement was either performed in duplicate or triplicate. Statistical analysis was performed with the unpaired two-tailed t test.
RESULTS
Induction of Chondrocyte Apoptosis and Ultrastructural Appearance of Chondrocyte-Derived Apoptotic Bodies.
Treatment of normal articular chondrocytes with SNP (9) or anti-Fas (10) causes apoptosis. Electron microscopy of a chondrocyte in normal articular cartilage shows the intact cell surrounded by a pericellular matrix (Fig. 1A). After treatment of cartilage with SNP, several changes can be observed. The cells shrink and small membrane-enclosed particles are budding off the cytoplasmic membrane (Fig. 1B). The pericellular matrix is degraded and subcellular particles accumulate in the interterritorial matrix (Fig. 1 C and D) or in the chondrocyte lacuna (Fig. 1E).
Figure 1.
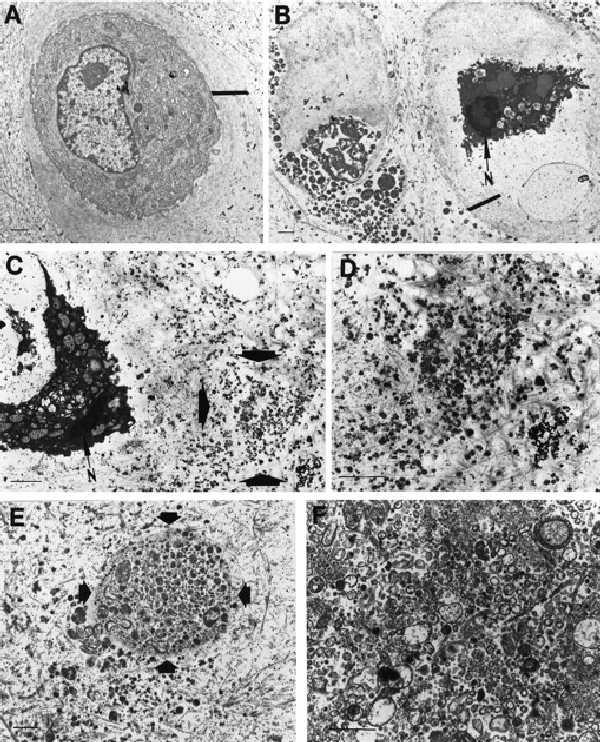
Electron microscopy of apoptotic bodies and matrix vesicles from normal and SNP-treated cartilage. (A) Electron micrograph of a chondrocyte from normal articular cartilage. (×5,400.) Small bar represents 1 μm. Large bar defines pericellular matrix. (B) Two adjacent apoptotic chondrocytes in SNP-treated cartilage. N = nucleus. (×4,300.) Small bar represents 1 μm. Large bar defines pericellular matrix. (C) Apoptotic chondrocyte in SNP-treated cartilage with apoptotic bodies accumulating in the cartilage matrix (area marked by arrows). (×8,500.) (D) High-magnification view of the area indicated by the arrow in C. (×14,800.) (E) Apoptotic bodies in chondrocyte lacuna marked by arrow. (×7,500.) (F) Apoptotic bodies isolated by ultracentrifugation from SNP-treated cartilage. (×15,000.) (Bars = 1 μm.)
To isolate apoptotic bodies, normal articular cartilage was treated with NO and subsequently digested with collagenase. Cells were removed from the digest by a 10,000 × g centrifugation, and the subcellular particles in the supernatant from the first centrifugation were pelleted by ultracentrifugation. Electron microscopy analysis shows that the apoptotic bodies in articular cartilage (Fig. 1E) and after isolation by ultracentrifugation (Fig. 1F) are similar vesicular structures of heterogeneous size.
Pyrophosphate Production by Apoptotic Bodies and Matrix Vesicles.
To assess functional properties of apoptotic bodies, the ability to produce pyrophosphate was analyzed and compared with matrix vesicles. Matrix vesicles (MV) were isolated from normal untreated cartilage. A mixture of MV and apoptotic bodies (MV/AB) was obtained after SNP treatment of an aliquot of the same cartilage, and AB were isolated from SNP-treated chondrocytes in monolayer culture. Subjecting normal monolayer chondrocytes and their conditioned media through the purification protocol for matrix vesicles did not yield an analyzable pellet. Identical amounts of AB and MV were examined, and, in the presence of ATP, the apoptotic bodies produced similar levels of PPi as MV or the mixture of AB and MV (Fig. 2A). We had shown earlier that chondrocytes produce increased levels of PPi after stimulation with TGF-β, whereas IL-1 suppresses PPi generation (25). Chondrocytes thus were pretreated with TGF-β, IL-1, or TGF-β plus IL-1 for 3 days. Apoptosis was induced with SNP, and apoptotic bodies were isolated. Equal amounts of apoptotic bodies, as determined by protein concentration from TGF-β-treated cells, produced higher levels of PPi as compared with apoptotic bodies from untreated cells. In contrast, apoptotic bodies from cultures that were treated with IL-1 produced lower levels of PPi (Fig. 2B).
Figure 2.
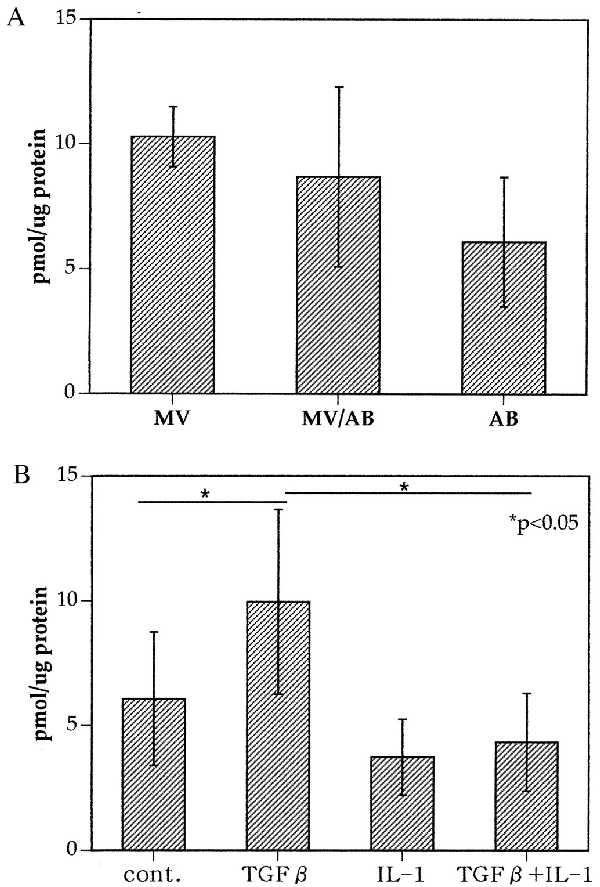
(A) Pyrophosphate production by matrix vesicles and apoptotic bodies. Matrix vesicles (MV) were isolated from normal articular cartilage. Normal articular cartilage was treated with SNP. The tissue was digested with collagenase, cells were pelleted at 20,000 × g, and matrix vesicles and apoptotic bodies in the cell-free supernatants were isolated by ultracentrifugation. Apoptotic bodies were isolated from SNP-treated chondrocytes. Equal amounts (on the basis of protein content) of MV, MV/AB mixtures, and AB were assayed for pyrophosphate production. Values represent pmol PPi/μg protein. (B) Chondrocyte-derived apoptotic bodies produce pyrophosphate; correlation with pretreatment of the cells. Chondrocytes were cultured in medium, TGF-β (10 ng/ml), IL-1β (1 ng/ml), or TGF-β plus IL-1β for 72 h. The conditioned medium from the cell cultures was harvested, and PPi levels were determined. The cells were then treated with SNP for an additional 24 h to induce apoptosis. The cultures were harvested and centrifuged at 20,000 × g for 20 min. The cell-free supernatants were then centrifuged at 150,000 × g. The pellets containing apoptotic bodies were resuspended in buffer containing ATP. PPi levels in the supernatants of the apoptotic bodies were then determined.
These results indicate that chondrocyte-derived apoptotic bodies can produce PPi and the levels of PPi production correlate with the patterns seen in the cells from which the apoptotic bodies are derived.
Enzyme Activities in Apoptotic Bodies: Alkaline Phosphatase, NTPPH, Plasma Cell Membrane Glycoprotein-1 (PC-1).
The presence of enzyme activities that are involved with pyrophosphate formation or degradation was examined in apoptotic bodies. The apoptotic bodies contained alkaline phosphatase activity and NTPPH activity (Fig. 3A). The levels of these enzyme activities were similar in apoptotic bodies or matrix vesicles isolated from chondrocytes, normal cartilage, or NO-treated cartilage. These results suggest that apoptotic bodies derived from NO-treated chondrocytes or cartilage contain alkaline phosphatase activity, a hallmark of matrix vesicles. Furthermore, the relative levels of NTPPH and alkaline phosphatase activity were indistinguishable in apoptotic bodies as compared with matrix vesicles. Comparison of intact chondrocytes with chondrocyte-derived apoptotic bodies did not show enrichment in alkaline phosphatase content but did show a relative increase in NTPPH activity (Table 1).
Figure 3.

(A) NTPPH and alkaline phosphatase activity. Apoptotic bodies (AB), matrix vesicles (MV), and mixtures of apoptotic bodies and matrix vesicles (AB/MV) were prepared as described in the legend to Fig. 2. Identical amounts of protein were assayed for NTPPH and alkaline phosphatase (ALK PHOS) activities. (B) Chondrocyte apoptotic bodies contain the PPi-generating enzyme PC-1. Apoptotic bodies sedimented by ultracentrifugation from SNP-treated chondrocytes were lysed in Triton X-100 and analyzed by SDS/PAGE under reducing conditions and Western blotting, using the polyclonal antibody 1769, as previously described (25).The figure shows a Western blot analysis of 10 μg of protein from the apoptotic bodies. The dark arrow indicates the detection of full-length PC-1 monomer (130 kDa); the smaller arrow indicates the presence of a larger PC-1 species of 260 kDa, attributable to incomplete reduction of PC-1 homodimers as previously observed (23).
Table 1.
Enzymatic activities of apoptotic bodies versus intact chondrocytes
| NTPPH | Alkaline phosphatase | |
|---|---|---|
| Chondrocytes | 2.13 ± 0.06 | 3.65 ± 0.21 |
| Apoptotic bodies | 5.35 ± 0.24 | 3.93 ± 0.41 |
Chondrocytes were cultured in medium or treated with antibody to Fas (1 μg/ml) for 24 h. Cultures were harvested, and apoptotic bodies were isolated by ultracentrifugation. Equal amounts of chondrocytes or apoptotic bodies as assessed by protein content were analyzed for enzyme content. Results are shown as units/5 μg protein.
Because chondrocyte apoptotic bodies generated PPi and possess PPi-forming NTPPH activity, we examined whether the molecular identity of the NTPPH was attributable to PC-1, which accounts for more than half of the NTPPH activity in human chondrocytes (23). The apoptotic bodies contained PC-1 detectable by Western blotting (Fig. 3B). Furthermore, the majority of NTPPH activity in the apoptotic bodies was removed by immunoprecipitation with 3E8, a previously characterized antibody to human PC-1 (25). A mouse control antibody did not reduce NTPPH activity (results not shown). Thus, PC-1 was a functional component of chondrocyte apoptotic bodies.
Calcium Deposition by Apoptotic Bodies.
One of the biologically most important functions of matrix vesicles is the ability to deposit calcium and form crystals. Apoptotic bodies were prepared from cultured chondrocytes and compared with matrix vesicles from normal articular cartilage. In addition, normal cartilage was treated with SNP, and the mixture of apoptotic bodies and matrix vesicles was isolated. Equal amounts of AB, MV, and AB/MV were tested in an in vitro calcium precipitation assay. Calcium deposition was observed only when both ATP and AB, or MV was added to the calcifying solution. All three types of samples formed crystalline deposits, and quantification of the amount of 45Ca in these precipitates showed almost identical values (Fig. 4).
Figure 4.
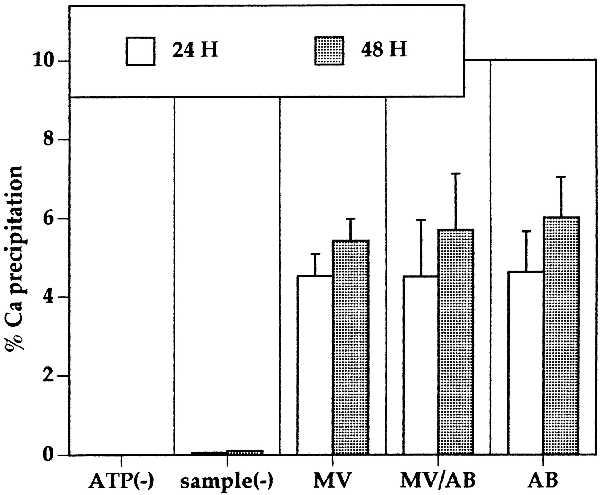
Calcium deposition by matrix vesicles and apoptotic bodies. Matrix vesicles isolated from articular cartilage and apoptotic bodies prepared from SNP-treated chondrocytes were isolated and incubated in calcifying solution containing 45Ca. Crystalline deposits were solubilized in HCl, and radioactivity was quantified by liquid scintillation counting. Assay controls include the omission of ATP [ATP(−)] and negative control without the addition of matrix vesicles or apoptotic bodies [sample(−)].
Apoptotic Bodies Derived from Anti-Fas-Treated Chondrocytes.
The effects of SNP and anti-Fas on chondrocyte apoptosis are quantitatively different and occur through at least partially different mechanisms (9, 10). Thus it was important to compare apoptotic bodies from cells that are either treated with SNP or anti-Fas. Apoptotic bodies were isolated from anti-Fas- or SNP-treated cells, and equal amounts on the basis of protein content were assayed for PPi production and calcium deposition. Table 2 shows that the amount of 45Ca precipitated by anti-Fas- or SNP-induced apoptotic bodies is equal.
Table 2.
Function of apoptotic bodies induced by SNP versus antibody to Fas
| PPi | % 45Ca deposited | |
|---|---|---|
| SNP | 6.1 ± 2.6 (n = 5) | 4.6 ± 1.4 (n = 5) |
| anti-Fas | 7.0 ± 3.2 (n = 3) | 4.0 ± 1.0 (n = 3) |
Chondrocytes were treated with SNP (1 mM) or antibody to Fas (1 μm/ml) for 24 h. Cultures were harvested, and apoptotic bodies were isolated by ultracentrifugation. Equal amounts of material as assessed by protein content were subjected to PPi and calcium deposition assays. PPi values are expressed as nanomoles PPi in medium.
DISCUSSION
Apoptosis is a physiologic clearance mechanism that is important in development and tissue homeostasis (6–8). The apoptotic bodies that are formed in this process are membrane-enclosed structures that are recognized by specific cell surface receptors and phagocytized to prevent exposure of intact tissues to cell constituents that can cause inflammation. The potential of apoptotic bodies to actively participate in a physiologic process or to contribute to pathogenesis has not been addressed experimentally.
The present study focused on the significance of apoptosis in articular cartilage. This tissue has several characteristics that are unique and important in this context. Articular cartilage is not vascularized, and chondrocytes are its only cellular constituent. Furthermore, there is no evidence that under normal conditions other cell types enter cartilage, and chondrocytes are not thought to migrate within the cartilage extracellular matrix. Cartilage is not known to contain phagocytes that can ingest apoptotic bodies. It is thus likely that apoptotic bodies remain within the articular cartilage unless extracellular matrix is degraded and release into the joint space occurs.
Cartilage shows profound, age-related changes in regard to extracellular matrix biochemical and biomechanical properties that appear to be at least in part the consequence of age-associated deficits in chondrocyte function. With advancing age, the cellularity of articular cartilage decreases. This is a possible consequence of chondrocyte senescence (26) or is related to chondrocyte death. In addition, there is a >50% incidence of abnormal calcium crystal deposition in cartilage in individuals above age 60. These age-related changes contribute to the pathogenesis of osteoarthritis, the most prevalent musculoskeletal disorder in humans.
Previously it was shown that chondrocytes produce large amounts of NO (27–29) and that NO can induce chondrocyte apoptosis (9). Antibody to the Fas antigen also induces apoptosis in a subpopulation of chondrocytes (10). Apoptotic bodies that are generated after exposure of cultured chondrocytes to NO donors or anti-Fas can be separated from cells and isolated as subcellular particles by ultracentrifugation. Electron microscopy showed that apoptotic bodies derived from primary cultured chondrocytes are membrane-enclosed structures of varying size. Chondrocyte apoptosis within articular cartilage was associated with several unique changes. The pericellular matrix that normally surrounds the chondrocytes and provides a transition to the interterritorial matrix is degraded, and within the territorial matrix adjacent to the chondrocyte lacunae, accumulation of apoptotic bodies can be observed. This indicates that in association with chondrocyte apoptosis there is degradation of pericellular matrix, and this allows the apoptotic bodies to enter the interterritorial space.
For functional analysis, apoptotic bodies were first isolated from cultured chondrocytes that were treated with NO or anti-Fas. The chondrocyte-derived apoptotic bodies produced pyrophosphate and contained NTPPH activity. We had shown earlier that in chondrocytes, TGF-β increases pyrophosphate production and NTPPH activity whereas IL-1 suppresses these responses (25). This provided an experimental system to study whether the function of apoptotic bodies is a reflection of the activation status of the cells they are derived from. The results showed that pretreatment of chondrocytes with IL-1 or TGF-β alters the levels of PPi and NTPPH activity in the apoptotic bodies that are derived from these cells after SNP or anti-Fas treatment.
Normal articular cartilage contains matrix vesicles (30) that are of similar size and appearance as apoptotic bodies that were obtained from cultures of apoptotic chondrocytes. In our studies we therefore subjected normal, untreated cartilage to collagenase digestion and matrix vesicle purification without in vitro induction of apoptosis. Other cartilage samples were treated with SNP subjected to the same isolation procedure. Matrix vesicles from normal cartilage, subcellular particles from SNP-treated cartilage, and apoptotic bodies from cultured chondrocytes were compared in several functional assays. When samples from the different sources were normalized on the basis of protein content, levels of pyrophosphate production, NTPPH, and alkaline phosphatase activities were all similar. Thus, at least with respect to the variables tested, matrix vesicles and apoptotic bodies are indistinguishable.
One of the most important roles of matrix vesicles in epiphyseal cartilage is the formation of calcium-containing crystals. We examined whether this is also a function of apoptotic bodies. The first assay used was a quantitative test that measures the precipitation of 45Ca. The results showed that apoptotic bodies that are derived from chondrocytes treated with SNP or anti-Fas can indeed precipitate calcium. When identical amounts of matrix vesicles (derived from normal cartilage), apoptotic bodies from cultured chondrocytes, and mixtures of matrix vesicles and apoptotic bodies (obtained from SNP-treated cartilage) were compared, the amount of 45Ca precipitated was similar. These results suggest that apoptotic bodies can precipitate calcium from solution.
These observations have the following implications for concepts on the pathogenesis of osteoarthritis. Apoptosis of chondrocytes in articular cartilage is associated with the destruction of the pericellular matrix and accumulation of apoptotic bodies in chondrocyte lacunae and in the interterritorial space. Apoptotic bodies are likely to remain at these sites because articular cartilage does not contain phagocytic cells. Apoptotic bodies express several functional activities. They can produce pyrophosphate and contain NTPPH and alkaline phosphatase activities. The presence of other enzymatic activities, such as metalloproteinases, is under investigation. Furthermore, apoptotic bodies can precipitate calcium from solution. Important changes occur in chondrocyte production of pyrophosphate with aging. The levels of constitutively produced and TGF-β-induced pyrophosphate increase with aging in articular chondrocytes (31, 32).
Thus, with advancing age there are several changes that represent potential risk factors for the development of osteoarthritis. With aging, cartilage cellularity decreases; in aging and osteoarthritic cartilage there is an increased frequency of apoptotic cells (S.H., R.L.O., and M.L., unpublished data), which is associated with the accumulation of apoptotic bodies; and there is an increase in pyrophosphate production. This constitutes a milieu in which calcium crystals can be formed. This scheme provides a mechanism to explain the age-associated increase in cartilage calcification. Furthermore, it is known that crystals can activate chondrocytes (33), and it is likely that the remaining viable cells in cartilage with abnormal calcium deposits are activated to undergo an unsuccessful remodeling process that is characteristic of osteoarthritis and terminates in the complete loss of articular cartilage.
Acknowledgments
We thank Drs. R. D. Coutts, K. Benirschke, P. Wolf, and C. Colwell for providing cartilage samples and Dr. James Goding for antibody to PC-1. This study was supported by National Institutes of Health Grants AG07996 and P608R40770, the Veterans Affairs Research Service, and the Sam and Rose Stein Institute for Research on Aging.
ABBREVIATIONS
- AB
apoptotic bodies
- SNP
sodium nitroprusside
- PPi
pyrophosphate
- NTPPH
NTP pyrophosphohydrolase
- AP
alkaline phosphatase
- TGF
transforming growth factor
- IL
interleukin
- PC-1
plasma cell membrane glycoprotein-1
Footnotes
To whom reprint requests should be addressed. e-mail: mlotz@scripps.edu.
References
- 1.Hamerman D. Ann Rheum Dis. 1995;54:82–85. doi: 10.1136/ard.54.2.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sokoloff L. In: Cartilage, Biomedical Aspects. Hall B K, editor. New York: Academic; 1983. , 110 pp. [Google Scholar]
- 3.Abyad A, Boyer J T. Curr Opin Rheumatol. 1992;4:153–159. doi: 10.1097/00002281-199204000-00004. [DOI] [PubMed] [Google Scholar]
- 4.Stockwell R A. J Anat. 1971;109:411–421. [PMC free article] [PubMed] [Google Scholar]
- 5.Vignon, E., Arlot, M., Patricot, L. M. & Vignon, G. (1976) Clin. Orthop. 303–308. [PubMed]
- 6.Majno G, Joris I. Am J Pathol. 1995;146:3–15. [PMC free article] [PubMed] [Google Scholar]
- 7.Jacobson M D, Weil M, Raff M C. Cell. 1997;88:347–354. doi: 10.1016/s0092-8674(00)81873-5. [DOI] [PubMed] [Google Scholar]
- 8.White E. Genes Dev. 1996;10:1–15. doi: 10.1101/gad.10.1.1. [DOI] [PubMed] [Google Scholar]
- 9.Blanco F J, Ochs R L, Schwarz H, Lotz M. Am J Pathol. 1995;146:75–85. [PMC free article] [PubMed] [Google Scholar]
- 10.Hashimoto S, Setareh M, Ochs R L, Lotz M. Arthritis Rheum. 1997;40:1749–1755. doi: 10.1002/art.1780401004. [DOI] [PubMed] [Google Scholar]
- 11.Anderson H C. J Cell Biol. 1967;35:81–101. doi: 10.1083/jcb.35.1.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bonucci E. J Ultrastruct Res. 1967;20:33–50. doi: 10.1016/s0022-5320(67)80034-0. [DOI] [PubMed] [Google Scholar]
- 13.Wuthier R E, Chin J E, Hale J E, Register T C, Hale L V, Ishikawa Y. J Biol Chem. 1985;260:15972–15979. [PubMed] [Google Scholar]
- 14.Bonewald L F, Schwartz Z, Swain L D, Boyan B D. Bone Miner. 1992;17:139–144. doi: 10.1016/0169-6009(92)90725-s. [DOI] [PubMed] [Google Scholar]
- 15.Dean D D, Schwartz Z V, Muniz O E, Gomez R, Swain L D, Howell D S, Boyan B D. Bone Miner. 1992;17:172–176. doi: 10.1016/0169-6009(92)90731-r. [DOI] [PubMed] [Google Scholar]
- 16.Ali S Y. Fed Proc. 1976;35:135–142. [PubMed] [Google Scholar]
- 17.Ali S Y, Sajdera S W, Anderson H C. Proc Natl Acad Sci USA. 1970;67:1513–1520. doi: 10.1073/pnas.67.3.1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mitchell N S, Shepard N L. Anat Rec. 1990;227:397–404. doi: 10.1002/ar.1092270403. [DOI] [PubMed] [Google Scholar]
- 19.Derfus B A, Rachow J W, Mandel N S, Boskey A L, Buday M, Kushnaryov V M, Ryan L M. Arthritis Rheum. 1992;35:231–240. doi: 10.1002/art.1780350218. [DOI] [PubMed] [Google Scholar]
- 20.Derfus B, Steinberg M, Mandel N, Buday M, Daft L, Ryan L. J Rheumatol. 1995;22:1514–1519. [PubMed] [Google Scholar]
- 21.Verma R S, Conte R A, Luke S, Sindwani V, Macera M J. Clin Genet. 1992;42:267–270. doi: 10.1111/j.1399-0004.1992.tb03253.x. [DOI] [PubMed] [Google Scholar]
- 22.Cheung C P, Suhadolnik R J. Anal Biochem. 1977;83:52–56. doi: 10.1016/0003-2697(77)90508-5. [DOI] [PubMed] [Google Scholar]
- 23.Huang R, Rosenbach M, Vaughn R, Provvedini D, Rebbe N, Hickman S, Goding J, Terkeltaub R. J Clin Invest. 1994;94:560–567. doi: 10.1172/JCI117370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Oyajobi B O, Caswell A M, Russell R G. J Bone Miner Res. 1994;9:99–109. doi: 10.1002/jbmr.5650090114. [DOI] [PubMed] [Google Scholar]
- 25.Lotz M, Rosen F, McCabe G, Quach J, Blanco F, Dudler J, Solan J, Goding J, Seegmiller J E, Terkeltaub R. Proc Natl Acad Sci USA. 1995;92:10364–10368. doi: 10.1073/pnas.92.22.10364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Guerne P A, Blanco F, Kaelin A, Desgorges A, Lotz M. Arthritis Rheum. 1995;38:960–968. doi: 10.1002/art.1780380712. [DOI] [PubMed] [Google Scholar]
- 27.Stadler J, Stefanovic-Racic M, Billiar T R, Curran R D, McIntyre L A, Georgescu H I, Simmons R L, Evans C H. J Immunol. 1991;147:3915–3920. [PubMed] [Google Scholar]
- 28.Palmer R M, Andrews T, Foxwell N A, Moncada S. Biochem Biophys Res Commun. 1992;188:209–215. doi: 10.1016/0006-291x(92)92371-4. [DOI] [PubMed] [Google Scholar]
- 29.Rediske J, Koehne C, Zhang B, Lotz M. Osteoarthritis Cartilage. 1994;2:199–206. doi: 10.1016/s1063-4584(05)80069-x. [DOI] [PubMed] [Google Scholar]
- 30.Livne E, Oliver C, Leapman R D, Rosenberg L C, Poole A R, Silbermann M. J Anat. 1987;150:61–74. [PMC free article] [PubMed] [Google Scholar]
- 31.Rosenthal A K, Ryan L M. Mech Aging Dev. 1994;75:35–44. doi: 10.1016/0047-6374(94)90026-4. [DOI] [PubMed] [Google Scholar]
- 32.Rosen F, McCabe G, Quach J, Solan J, Terkeltaub R, Seegmiller J E, Lotz M. Arthritis Rheum. 1997;40:1275–1281. doi: 10.1002/1529-0131(199707)40:7<1275::AID-ART12>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- 33.Terkeltaub R. Pathogenesis and Treatment of Crystal Induced Inflammation. Baltimore: Williams & Wilkins; 1997. [Google Scholar]