

Abstract
Marsupials are a diverse group of mammals that occupy a large range of habitats and have evolved a wide array of unique adaptations. Although they are as diverse as placental mammals, our understanding of marsupial brain organization is more limited. Like placental mammals, marsupials have striking similarities in neocortical organization, such as a constellation of cortical fields including S1, S2, V1, V2, and A1, that are functionally, architectonically, and connectionally distinct. In this review, we describe the general lifestyle and morphological characteristics of all marsupials and the organization of somatosensory, motor, visual, and auditory cortex. For each sensory system, we compare the functional organization and the corticocortical and thalamocortical connections of the neocortex across species. Differences between placental and marsupial species are discussed and the theories on neocortical evolution that have been derived from studying marsupials, particularly the idea of a sensorimotor amalgam, are evaluated. Overall, marsupials inhabit a variety of niches and assume many different lifestyles. For example, marsupials occupy terrestrial, arboreal, burrowing, and aquatic environments; some animals are highly social while others are solitary; and different species are carnivorous, herbivorous, or omnivorous. For each of these adaptations, marsupials have evolved an array of morphological, behavioral, and cortical specializations that are strikingly similar to those observed in placental mammals occupying similar habitats, which indicate that there are constraints imposed on evolving nervous systems that result in recurrent solutions to similar environmental challenges.
Keywords: cortical organization, evolution, somatosensory cortex, auditory cortex, visual cortex, motor cortex
1. Introduction
Marsupials are a major order of mammals whose ancestors radiated from stem mammals over 180 million years ago (MYA; Figure 1, based on Murphy et al., 2004). Although most of us are familiar with only a few species of marsupials, such as kangaroos, koalas, and opossums, there are actually hundreds of marsupial species. Like placental mammals, marsupials are a highly diverse group that have adapted to a number of different habitats and lifestyles. For example, they can be terrestrial, arboreal, burrowing, or aquatic. Marsupials can be highly social, like the striped possum (Dactylopsila trivirgata), or solitary, like the koala (Phascolarctos cinereus); they can be carnivorous, herbivorous, or omnivorous. This diversification is tied to the evolution of unique peripheral morphologies and highly specialized behaviors, both of which are related to alterations in overall brain size, neocortical organization, and connectivity.
Figure 1.
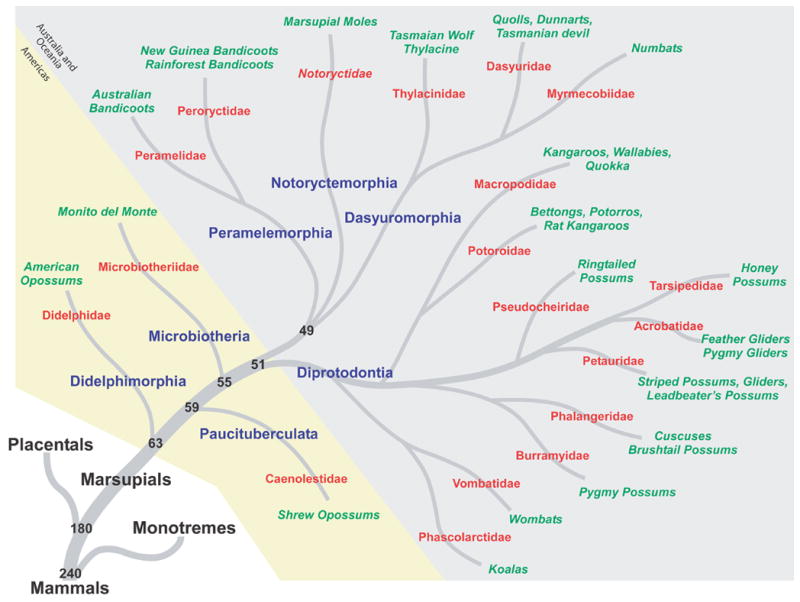
A phylogenetic tree illustrating the relationship between marsupials and other mammals. The seven extant marsupial orders are shown in blue; names of the families within each order are given in red; and examples of well-known species are in green. The three orders that are found in North and South America are shaded in yellow, and the remaining orders, which are found in Australia, Tasmania, New Guinea, and neighboring islands, are shaded in light gray. The timescale is in MYA; phylogenetic relationships are from Murphy et al., 2004.
In terms of gross morphology, marsupial brains vary dramatically in size, shape, lissencephalization, and encephalization (the expected size of the brain relative to body size). For example, some marsupials have small, smooth brains with a small proportion of the brain devoted to the neocortex, such as the Virginia opossum (Didelphis virginiana; Figure 2), while others have relatively larger, gyrencephalic brains with a larger proportion of the brain devoted to the neocortex, such as the Tasmanian wombat (Vombatus ursinus) and the Western gray kangaroo (Macropus fuliginosus; Figure 2). Furthermore, regardless of overall brain size, some animals have proportionately small brains for their body size, and thus small encephalization quotients, such as the long-tailed planigale (Planigale ingrami; Iwaniuk et al., 2001). Conversely, the striped possum has an extremely large brain and a particularly large neocortex, relative to the size of its body, with an encephalization quotient that rivals that of some primates (Figure 2; Iwaniuk et al., 2001; Nelson and Stephan, 1982).
Figure 2.
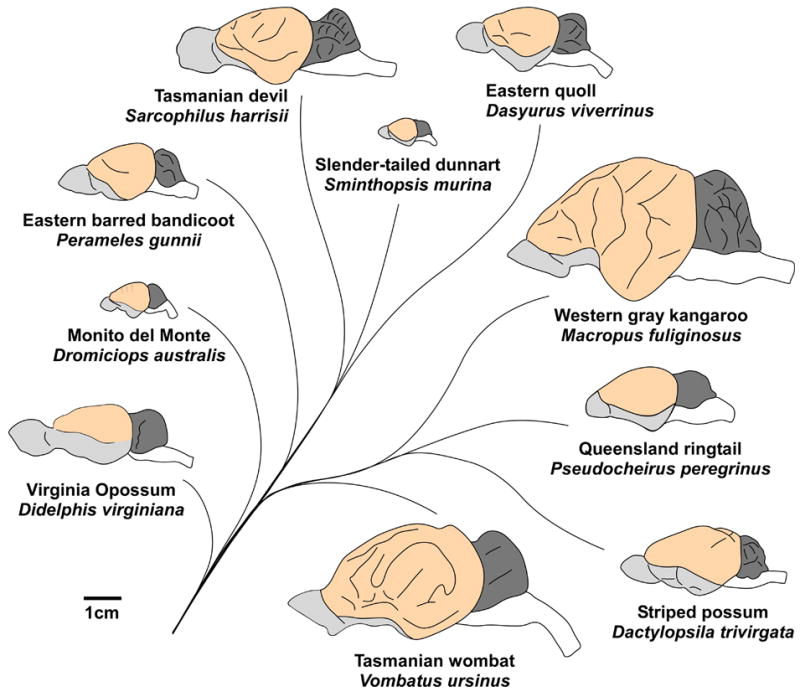
Lateral views of ten representative marsupial brains. Marsupial brains can vary dramatically in their overall size; compare the brain of the Western gray kangaroo with that of the slender-tailed dunnart. The relative size of distinct brain areas also differ between species, such as the size of the olfactory bulb and pyriform cortex (gray), the size of the neocortex (light orange), and the size of the cerebellum (dark gray). Finally, the smoothness of the neocortical surface varies between species, such as the lissencephalic neocortex of the Virginia opossum as compared to the gyrencephalic neocortex of the Tasmanian wombat. Lines denote phylogenetic relationships between species; rostral is left; medial is up; scale bar is 1 cm. Brains were drawn based on photographs from the University of Wisconsin and Michigan State Comparative Mammalian Brain Collections (http://www.brainmuseum.org), which has been funded by the National Science Foundation and the National Institutes of Health, photographs from Johnson, 1977, and photographs provided by John Nelson (personal communication).
Despite the fact that marsupial lifestyle, morphology, and gross brain organization are as diverse as those observed in placental mammals, our understanding of marsupial neocortex organization is more limited than that of placental mammals. Despite this limitation, these mammals are important for understanding larger issues of cortical evolution and development for a number of reasons. First, because marsupials occupy a wide range of habitats and have evolved a large array of unique adaptations, they make excellent models for studying animal ecology and the relationship between brain and behavior. Further, the co-evolution of peripheral morphology, sensory abilities, and cortical organization in marsupials parallels the evolution of these characteristics in placental mammals. Second, since many present day marsupials have retained a number of features of cortical organization from their ancient ancestor, several marsupials (particularly within the order Didelphimorphia) are considered to reflect the ancestral mammalian state more than most other present-day mammals. Because of this, information on marsupial brain organization has spurred a number of theories of cortical field evolution that have a large impact on how we think about brain evolution today. Finally, because of the extremely immature state of marsupials at birth, they serve as important models for studying neural development, since controlled manipulations of the sensory environment and/or the nervous system can be made ex utero at a very early developmental stage.
In this review, we begin by examining the general lifestyle and morphological characteristics of all marsupials. We then describe the organization of somatosensory, motor, visual, and auditory cortex in marsupials. For each sensory system, we will compare the functional organization of the neocortex and the corticocortical and thalamocortical connectivity across species. Differences between placental and marsupial species will be discussed, and we will evaluate the theories on neocortical evolution that have been derived from studying marsupials, particularly the idea of a sensorimotor amalgam. Finally, we will use these comparisons to summarize the aspects of cortical organization that are the same for all mammals and to infer the plan of cortical organization that may have been present in the common ancestor. We will then provide a few examples of how cortical organization has been altered in different lineages and speculate about how alterations in the basic plan of organization are associated with specialized behaviors and individual lifestyles.
2. What characterizes a marsupial?
Marsupials represent one of three subclasses of mammals that include prototherians (monotremes), metatherians (marsupials), and eutherians (placentals). Both the terms ‘marsupial’ and ‘placental’ are misnomers, since not all marsupials have a pouch, or marsupium, and since both placentals and marsupials have a placenta during gestation. The term metatherian indicates that, in terms of evolution, marsupials are midway between prototherian and eutherian mammals, and this term is often used synonymously with the term marsupial. However, recent evidence from gene sequencing studies indicates that marsupials arose concurrently with eutherian mammals, about 180 MYA (Murphy et al., 2004). Conventionally, the first marsupials are thought to have arisen in the New World (either North or South America) and subsequently migrated to Australia and South East Asia, possibly through Antarctica (Marshall Graves and Westerman, 2002; Szalay, 1982; Thenius, 1990; Woodburne and Case, 1996; Woodburne et al., 2003). Alternatively, recent fossil evidence suggests that marsupials may have arisen in Laurasia, somewhere in present-day Asia, before migrating to North America and later to Australia (Luo et al., 2001; Luo et al., 2003).
Extant marsupials have been subdivided into seven different orders, and over 270 species of marsupials have been identified (Figure 1). The exact phylogenetic relationship between marsupial orders, and even between families within the larger orders, is unresolved because different types of data, such as dental and skeletal morphology, DNA sequencing, and mitochondrial genome analysis, support conflicting phylogenetic relationships (Figure 3; Cardillo et al., 2004; Horovitz and Sanchez-Villagra, 2003; Luo et al., 2003; e.g., Marshall Graves and Westerman, 2002; Munemasa et al., 2006; Murphy et al., 2004; Nilsson et al., 2004). In general, the order Didelphimorphia is considered by most to be the oldest group of Marsupials (Figures 1 and 3). These North, Central, and South American marsupials are morphologically homogeneous and, in terms of brain organization, have few cortical fields and relatively small brains for their body size (Iwaniuk et al., 2001). The order Paucituberculata contains one extant family, Caenolestidae, which is composed of five species of small, shrew-like animals found in western South America; very little is known about the organization of their neocortex. Similarly, the order Microbiotheria is a small radiation, having one extant species, Monito del Monte (Dromiciops australis). Although this animal is only found in South America, phylogenic studies demonstrate that it is more closely related to Australian marsupials than to American marsupials (Figure 3). The remaining four orders are found in Australia, Tasmania, New Guinea, and neighboring islands. Dasyuromorphia is composed of three families (Figure 1) and is considered to have retained many primitive features of the Australian common ancestor. The order Peramelemorphia consists of two families of bandicoots, both of which have been found in a wide range of habitats. Notoryctemorphia only contains one species of marsupial mole (Notoryctes typhlops), although there is debate as to whether a second species (Notoryctes caurinus) exists (Nowak, 2005). Finally, the order Diprotodontia is the largest and most diverse order, consisting of 10 families, which include most of the well known marsupials (i.e., kangaroos, koalas, wallabies, and wombats; see Figure 1). The proportion of the brain devoted to the neocortex is largest in this order (Nelson and Stephan, 1982), and with the exception of Didelphimorphia, the most is known about cortical organization within this order.
Figure 3.
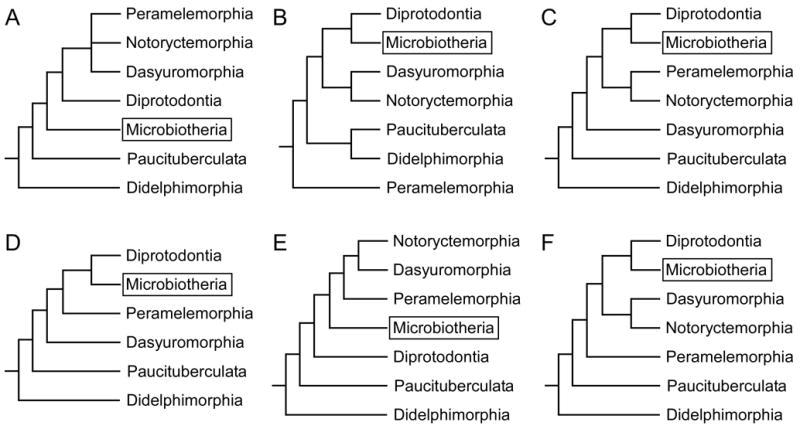
The phylogenetic relationship between marsupial orders is unresolved. For example, the relationship between the order Microbiotheria (boxed) and other marsupial orders changes depending on what type of data is used to construct the phylogenetic tree. The six phylogenies shown are based on DNA sequencing (A, B), dental and skeletal morphology (C,D), mitochondrial genome (B,E), and a meta-analysis that combined numerous phylogenetic relationships described in the literature (F). (A) is the same tree shown in Figure 1 from Murphy et al., 2004; (B) from Marshall Graves and Westerman, 2002; (C) from Horovitz and Sanchez-Villagra, 2003; (D) from Luo et al., 2003; (E) from Nilsson et al., 2004; and (F) from Cardillo et al., 2004.
Regardless of phylogeny, there are a number of morphological features, reproductive functions, and aspects of lifestyle that marsupials share. The most noted morphological feature is the marsupium, or pouch, from which they derive their name. The marsupium varies between species from a closed pouch to a small, primal pouch to no pouch at all (Nowak, 2005). In terms of reproductive functions, female marsupials, like other mammals, possess two ovaries, two oviducts, and two uteri (similar to some rodents, bats, whales, ungulates). However, marsupials are unique in that they possess two vaginas that connect bilaterally to a pseudovagina that functions as a birth canal, and males possess a double gland penis that corresponds to the two lateral vaginas (Klima and Maier, 1990; Moeller, 1990; Nowak, 2005). Marsupials have a short gestation, between 8 and 42 days (Moeller, 1990). The young are born at a very immature state, and they must make their way to the marsupium and attach to the mother’s nipple in order to survive. The immaturity of the young is reflected in the embryonic developmental state of most of their body features and sensory organs. For instance, these animals are hairless, their eyes and ears are closed, the hindlimbs and tail are stumps, and bone development is extremely immature (Figure 4). Only their olfactory system, their forelimbs, and to a lesser extent portions of the somatosensory system, are developed at birth. When marsupials are born, they pass through the pseudovagina and crawl, using their forelimbs, into the pouch using gravitational and/or olfactory cues to navigate (Moeller, 1990; Nowak, 2005). Once in the pouch, the young animals attach to a nipple and reside there for the remainder of early development.
Figure 4.
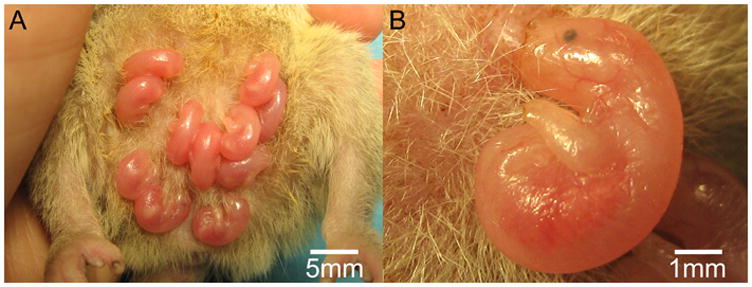
Marsupials are born at a very immature state, and they must make their way to the marsupium, or pouch, and attach to the mother’s nipple in order to survive. Shown in (A) is a litter of short-tailed opossums, a pouchless marsupial, on the day of birth. The immaturity of the young is evident in the embryonic developmental state of most of their body features and sensory organs (B). They are hairless, their eyes and ears are poorly developed, the hindlimbs and tail are stumps, and bone development is extremely immature. Only their olfactory system, the forelimbs, and to a lesser extent portions of the somatosensory system are developed at birth.
In regard to lifestyle, all but three species of marsupials are nocturnal or crepuscular. This is particularly interesting since all monotremes are nocturnal, suggesting that the common mammalian ancestor was nocturnal. Although it is often assumed that nocturnal animals lack color vision, recent studies have demonstrated that the honey possum (Tarsipes rostratus) and the fat-tailed dunnart (Sminthopsis crassicaudata) have three spectrally distinct cone photoreceptors (Arrese et al., 2002), a condition not observed in most placental mammals (see Jacobs and Rowe, 2004 for review). The presence of trichromacy in these marsupials may be a feature retained from ancestral reptiles, or as in primates, it may have been re-acquired and relate to specific requirements of their visual ecology.
Finally, in terms of gross brain organization, perhaps the most notable feature of the neocortex in all marsupials (and monotremes) is the lack of a corpus callosum, which was observed as early as 1837 (Owen, 1837). Instead, the primary form of interhemispheric communication is through an enlarged anterior commissure (Ebner, 1967, 1969; Granger et al., 1985; Heath and Jones, 1971; Loo, 1931; Martin, 1967; Smith, 1902), which has been shown to be functionally equivalent to the corpus callosum in placental mammals (Nelson and Lende, 1965; Putnam et al., 1968). Specifically, responses from one cortical area can be recorded from a homotopic area in the opposite hemisphere, and these responses are abolished by transecting the commissure joining the neocorticies (Nelson and Lende, 1965). Furthermore, in the Virginia opossum, the latencies and waveforms of the response are comparable to placental mammals (Nelson and Lende, 1965; Putnam et al., 1968). In marsupials, the anterior commissure is topographically organized with axons from rostral cortical areas crossing anteriorly, caudal areas crossing posteriorly, dorsal areas crossing dorsally, and ventral areas crossing ventrally, although there is considerable overlap between projections from these different areas (Ashwell et al., 1996a). Finally, the development of the anterior commissure has been extensively studied and shown to follow the same general pattern of formation as the corpus callosum in placental mammals (Ashwell et al., 1996a; Ashwell et al., 1996b; Cabana and Martin 1985; Cummings et al., 1997; Shang et al., 1997; Silver et al., 1982). The anatomy of other gross neuroanatomical structures in marsupials has been comprehensively described elsewhere and will not be covered in this review (Abbie, 1942; Ebner, 1969; e.g., Gray, 1924; Johnson, 1977; Loo, 1930; 1931; Mayner, 1989).
3. Somatosensory cortex
The organization of somatosensory cortex has been examined in several species of American and Australian marsupials. All species examined have a primary somatosensory area (S1) and a second somatosensory area (S2) located caudolateral to S1 (for review see Johnson, 1990; Rowe, 1990). The functional organization of S1 has been described using electrophysiological mapping techniques in several species (Figure 5), including the brush-tailed possum (Trichosurus vulpecula; Adey and Kerr, 1954; Elston and Manger, 1999; Haight and Weller, 1973; Weller, 1993), the Virginia opossum (Beck et al., 1996; Bodemer and Towe, 1963; Lende, 1963c; Pubols et al., 1976), the wallaby (Thylogale eugenii; Lende, 1963a), the white-eared opossum (Didelphis albiventris, previously Didelphis azarae azarae; Magalhaes-Castro and Saraiva, 1971), the Tasmanian wombat (Johnson et al., 1973), the short-tailed opossum (Monodelphis domestica; Catania et al., 2000; Frost et al., 2000; Huffman et al., 1999), the striped possum (Huffman et al., 1999), and the Northern quoll (Dasyurus hallucatus, also called the native cat; Huffman et al., 1999). In all species examined, S1 contains a complete and inverted representation of the contralateral body surface with the tail represented most medially, followed by the representation of the hindlimb, trunk, forelimb, forepaw, face, and oral structures in a medial to lateral progression (Figure 5). Most neurons in S1 respond to cutaneous stimulation, and receptive fields for neurons are generally small, particularly on specialized morphological structures, such as the vibrissae or forepaws.
Figure 5.
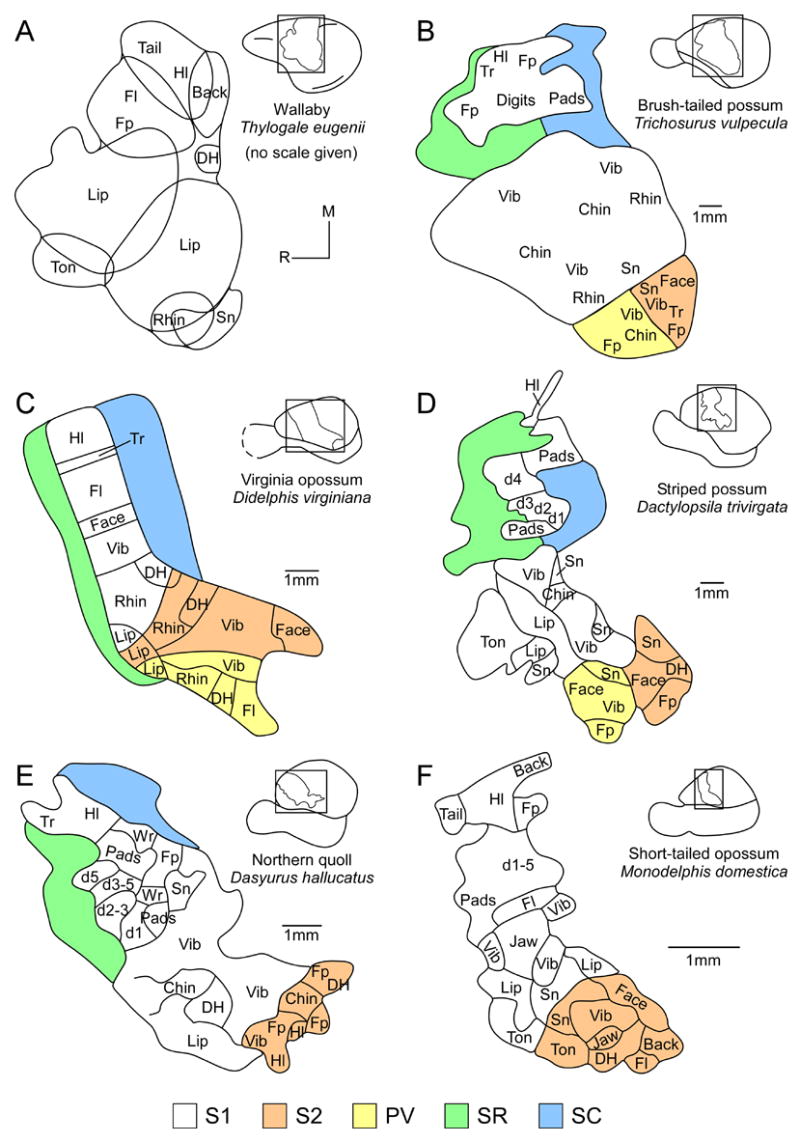
The functional organization of somatosensory cortex has been described using electrophysiological mapping techniques in several marsupial species. Shown here, all species had a complete and inverted representation of the contralateral body surface in S1 (white); individual body areas are given to show receptive field progression (A–F). When available, the receptive field progressions for S2 (orange) and PV (yellow) are also given. Since receptive field progressions have not been studied as extensively in SR (green) and SC (blue), only the outermost borders are shown. d1-5, digits 1-5; DH, dorsal head; Fl, forelimb; Fp, forepaw; Hl, hindlimb; M, medial; R, rostral; Rhin, rhinarium; Sn, snout; Ton, tongue; Tr, trunk; Vib, vibrissae; Wr, wrist. (A) based on Lende, 1963a; (B) based on Elston and Manger, 1999; (C) based on Beck et al., 1996; (D, E) based on Huffman et al., 1999; and (F) based on Frost et al., 2000.
These specialized morphological structures are associated with unique behaviors. For example, the structure of the paw in marsupials can vary dramatically between species, from the spade-shaped forepaw used for digging in the marsupial mole to the specially elongated digit used to extract insects while foraging in the striped possum (Figure 6 compare A and B). Although no information is available on the organization of S1 in the marsupial mole, detailed maps of S1 organization in the striped possum reveal an enlarged representation of the specialized fourth digit, D4. This is particularly apparent when compared to the organization of S1 in marsupials with forepaws that have five digits of similar length (Figure 5 compare D and E). This type of cortical magnification, or enlarged representation of specialized morphological structures, is commonly found not only in marsupials, but also in placental and monotreme mammals (see Krubitzer and Hunt, 2006 for review; Krubitzer and Kahn, 2004).
Figure 6.

Paw morphology can vary dramatically between marsupial species. Shown here are examples of forepaws (A, B) and hindpaws (C–G) from seven different marsupial species. The marsupial mole (A) has a spade-shaped forepaw used for digging. The striped opossum (B) has a specially elongated digit used to extract insects while foraging. The water opossum (C) has webbed feet that it uses to propel itself through water. The brush-tailed possum (E), the Virginia opossum (F), and the tree kangaroo (G) have feet that are adapted for climbing, and the red kangaroo (D) has an elongated foot that is specialized for jumping. (A, C, D, F, G) based on Parker, 1990; (B) based on Huffman et al., 1990; (E) based on Myers et al., 2006.
In addition to being functionally defined, S1 has been shown to be co-extensive with a distinct architectonic appearance (e.g., Adey and Kerr, 1954; Ashwell et al., 2005; Beck et al., 1996; Catania et al., 2000; Christensen and Ebner, 1978; Foster et al., 1981; Weller, 1972; 1993). Using a variety of staining procedures, the cortical architecture of S1 has been described in a number of different marsupials. First, S1 is readily identified by its well-defined granular layer (layer IV), which contains densely packed cells that stain darkly for Nissl (Adey and Kerr, 1954; Ashwell et al., 2005; Elston and Manger, 1999; Foster et al., 1981). In fact, many of the same cell types, as defined by Golgi-Cox and Nissl staining, are found in the parietal cortex (which contains S1) of both marsupials and placental mammals (Walsh and Ebner, 1970). In several marsupials, a distinct barrel field (in the brush-tailed possum) or barrel-like subdivisions (in the Virginia opossum, Tammar wallaby (Macropus eugenii), and striped possum) have been found within S1 in both coronally sectioned and tangentially sectioned tissue processed for Nissl, succinic dehydrogenase (SDH), or myelin (Beck et al., 1996; Elston and Manger, 1999; Waite et al., 1998; Waite et al., 1991; Weller, 1993; Woolsey et al., 1975). Unlike the barrel fields in mice and rats that are characterized by densely packed cells surrounding a loosely packed center, the barrel field in marsupials contains loosely packed cells surrounding a densely packed center (Possibly add figure of barrel cortex; Weller, 1993). Using electrophysiological recording techniques, a direct correspondence between each barrel and an individual whisker has been shown in marsupials, similar to what has been described in rodents (Waite et al., 1991; Weller, 1993; Woolsey et al., 1975). As in placental mammals, barrels are not common to all species, and they have only been described in the species listed above.
Finally, in cortex that has been sectioned parallel (tangential) to the cortical surface, S1 stains darkly for myelin, SDH, cytochrome oxidase (CO), and/or nicotinamide adenine dinucleotide phosphate diaphorase (NADPHd), a marker for GABAergic non-pyramidal neurons (Catania et al., 2000; Frost et al., 2000; Huffman et al., 1999). In addition to the overall appearance of S1, histologically identified modules, or isomorphs, have also been identified in some species. For example, in the brush-tailed possum S1 is densely myelinated and separated by myelin-sparse cortex which contains neurons that are responsive to stimulation of deep receptors (Elston and Manger, 1999). In the striped-possum, distinct CO-rich territories have been observed for different body part representations in S1 (Huffman et al., 1999), and these areas are similar to the isomorphic representation described in some primates (e.g., Jain et al., 2001).
Caudolateral to S1 is a second small representation of the contralateral body surface called S2. This area has been electrophysiologically identified in all marsupials examined (Figure 5), including the brush-tailed possum (Adey and Kerr, 1954; Coleman et al., 1999; Elston and Manger, 1999), the Virginia opossum (Beck et al., 1996; Lende, 1963c; Pubols, 1977), the white-eared opossum (Magalhaes-Castro and Saraiva, 1971), the Northern quoll (Huffman et al., 1999), the striped possum (Huffman et al., 1999), and the short-tailed opossum (Catania et al., 2000; Frost et al., 2000). As in other mammals, S2 in marsupials contains a non-inverted representation of the body with the representation of the face and oral structures located adjacent to similar representations in S1 (Figure 5B–F). The forelimb and hindlimb representations are located progressively caudal in S2, and the trunk and proximal body parts are represented more rostromedially. The receptive fields for neurons in S2 are larger than for neurons in S1, and are often responsive to bilateral somatosensory stimulation and/or to auditory stimulation (Lende, 1963c; Pubols, 1977). In most species examined, the S2 region actually contains two separate fields, a rostral field called the parietal ventral area (PV) and a caudal field termed S2 (e.g., Disbrow et al., 2000; Krubitzer, 1995; Krubitzer et al., 1986). Both fields are somatotopically organized, and neurons in each field have large receptive fields. Separate S2 and PV representations have been identified in several species of marsupials in which this region has been mapped in detail, such as the Virginia opossum (Beck et al., 1996), the brush-tailed possum (Elston and Manger, 1999), and the striped possum (Huffman et al., 1999). A single representation called either S2 or S2/PV has been described in the Northern quoll (Huffman et al., 1999) and the short-tailed opossum (Catania et al., 2000).
As with S1, S2 is architectonically distinct. In Nissl stains, layers III and V are broad, and the granular layer (layer IV) is not well defined (Adey and Kerr, 1954). In tangentially sectioned tissue, S2 is moderately to darkly myelinated, and it stains darkly for CO and NADPHd in most species (Beck et al., 1996; Catania et al., 2000; Huffman et al., 1999); however both SII and PV are sparsely myelinated in the brush-tailed possum (Elston and Manger, 1999).
Two other fields have been identified in the parietal cortex of marsupials; one cortical field is rostral to S1 and the other field is caudal to S1. These fields have been termed the somatosensory rostral (SR) and somatosensory caudal (SC) fields (Beck et al., 1996; Huffman et al., 1999). Neurons in both SR and SC respond to stimulation of deep receptors or high threshold cutaneous receptors (Beck et al., 1996; Huffman et al., 1999). Further, it has been proposed that these areas correspond to fields that have been identified in other mammals, such as rodents and primates (Beck et al., 1996; Huffman et al., 1999; Kaas, 2004b; Krubitzer and Padberg, 2007; Slutsky et al., 2000). Based on location, stimulus preference, and architecture it is possible that the rostral field in marsupials is homologous to area 3a of primates and flying foxes, and the rostral somatosensory area of rodents. The caudal field in marsupials appears to correspond to posterior parietal area (PP; formerly PM) in squirrels and posterior parietal cortex (PPC) in rats, which is hypothesized to be homologous to one or more of the posterior parietal fields identified in primates, such as area 5 (Krubitzer and Padberg, 2007). However, this hypothesis has not been tested.
Studies of corticocortical and thalamocortical connections of somatosensory cortex in marsupials are relatively restricted to connections of S1, and they have been studied using retrograde degeneration techniques (Bodian, 1942; Diamond and Utley, 1963; Goldby, 1943; Pubols, 1968) and anatomical tracers such as horseradish peroxidase (HRP) and/or tritiated amino acids (Beck et al., 1996; Donoghue and Ebner, 1981a; b; Elston and Manger, 1999; Haight and Neylon, 1978; 1981; Marotte et al., 1997). In the brush-tailed possum (Elston and Manger, 1999) and the Virginia opossum (Beck et al., 1996), S1 has dense ipsilateral corticocortical connections with S2, PV, SR, SC, cortex immediately lateral to S1, and in the brush-tailed possum only, with motor cortex. S1 has sparse connections with cortex of the medial wall. Interhemispheric connections are restricted to S1, PV, and cortex lateral to S1 (Elston and Manger, 1999). In one study (Elston and Manger, 1999), one injection was made into cortex caudal to SC. The connections of this region are very broadly distributed. Specifically, cortex caudal to SC receives inputs from S1, S2, PV, SR, SC, motor cortex, visual cortex (areas 17 and 18), limbic cortex of the medial wall, and temporal cortex. Interhemispheric connections of this region are more dense than those of S1, and they are distributed to the same regions as those observed ipsilaterally.
Thalamocortical connections have been examined in the Virginia opossum (Bodian, 1942; Diamond and Utley, 1963; Donoghue and Ebner, 1981a; b; Pubols, 1968), the brush-tailed possum (Haight and Neylon, 1978), the Northern quoll (Haight and Neylon, 1981), and the Tammar wallaby (Marotte et al., 1997). In the Virginia opossum, thalamic projections to S1 arise predominantly from the ventroposterior nucleus (VP; Diamond and Utley, 1963; Pubols, 1968). Moderate projections are also from the ventrolateral nucleus (VL), the central intralaminar nucleus (CIN), the central lateral nucleus (CL), and the ventromedial nucleus (VM; Donoghue and Ebner, 1981b). However, Jones (1985) has proposed that the CIN was misidentified by Donoghue and Ebner (1981b) and that the CIN projections are actually with the posterior nucleus (Po). Similar patterns of thalamocortical connectivity have been observed after injections into parietofrontal cortex (which contains S1) in the brush-tailed possum (Haight and Neylon, 1978) and the Northern quoll (Haight and Neylon, 1981). In the brush-tailed possum (Haight and Neylon, 1978), each division of VP projects to a specific region of cortex.
Taken together, the electrophysiological recording data and architectonic analysis indicate that at least two unimodal somatosensory fields are present in the neocortex of marsupials, S1 and S2. In some species, PV is also clearly present. An additional field, SR, is associated with the somatosensory system, but is likely associated with motor processing as well. Likewise, the field caudal to S1,SC, is clearly associated with somatosensory processing, but may be homologous to one or more fields in the posterior parietal cortex of primates involved in sensorimotor integration.
4. Motor cortex
The organization of motor cortex in marsupials has been of great interest to evolutionary neurobiologists for a number of decades because it is thought to most closely resemble the sensorimotor cortex of the first mammals. The earliest experiments on motor cortex in marsupials used gross electrical stimulation with unipolar and bipolar electrodes, and demonstrated that motor cortex is located at the rostral pole of the neocortex, usually centered around the orbital sulcus (Chapman, 1906; Cunningham, 1898; Flashman, 1906; Herrick, 1898; Herrick and Tight, 1890; Vogt and Vogt, 1907; Weed and Langworthy, 1925; Ziehen, 1897). In these early studies, researchers demonstrated that movements of individual body parts, such as the face, vibrissae, and forepaws, are represented progressively across the cortical surface; however, only half of the studies were able to find a clear representation of body areas posterior to the forepaws (i.e., the hindpaws and tail). Thus, for many years there was a debate as to whether marsupials had a motor representation of the lower body similar to what had been identified in placental mammals (Rogers, 1924; see Gray and Turner, 1924 for review).
In the mid-1900s, as techniques improved, several studies addressed this issue. In 1939, using unipolor AC stimulation techniques in the brush-tailed possum, Golby demonstrated the existence of a motor cortex with a complete representation of the body at the rostral pole of the neocortex (Figure 7A). The field consisted of a topographically organized representation of body movements with the hindlimb represented medially, followed by representations of the forelimb and face more laterally. Architectonic examination of cortex in this stimulated area confirmed that it corresponded to Brodmann’s area 4, as described in marsupial and placental mammals (Brodmann, 1909), and to the primary motor area (M1), as determined using similar techniques in both monotremes (Abbie, 1938; Lende, 1964) and placental mammals (Penfield and Boldrey, 1937).
Figure 7.
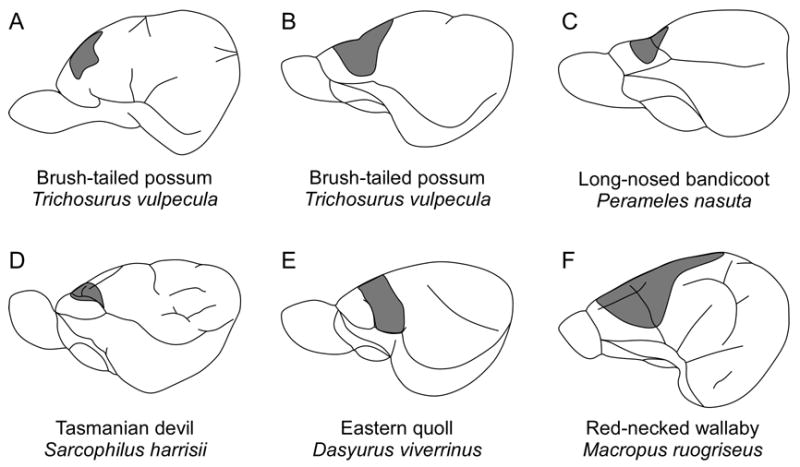
Early studies of motor cortex demonstrate that it is topographically organized and is located at the rostral pole of the neocortex, usually centered around the orbital sulcus. A complete body representation has been found in the brush-tailed possum (A, B) and the red-necked wallaby (F); however there was no representation of the lower body in the long-nosed bandicoot (C), the Tasmanian devil (D), or the Eastern quoll (E). Conventions as in previous figures; (A) based on Goldby, 1939; (B–F) based on Abbie, 1940.
Concurrently, Abbie (1940) demonstrated that body movements could be evoked at the far rostral pole of cortex in five different species (Figure 7B–F): the long-nosed bandicoot (Perameles nasuta), the Tasmanian devil (Sacrophilus harrissi), the Eastern quoll (Dasyurus viverrinus), the brush-tailed possum, the red-necked wallaby (Macropus rufogriseus, previously Macropus ruficollis), and the agile wallaby (Macropus agilis). His results demonstrated that motor cortex is topographically organized in all five species, but that it does not include a representation of the lower body in the long-nosed bandicoot, the Tasmanian devil, or the Eastern quoll (species from the Orders Peramelemorphia and Dasyuromorphia). In the brush-tailed possum, the red-necked wallaby, and the agile wallaby (species from the Order Diprotodontia), there is a motor representation of the hindlimb but not of the tail. He concluded that there is probably no cortical representation of the lower body in Peramelemorphia and Dasyuromorphia, there is a representation of the whole body (except the tail) in Diprotodontia, and that the results are inconclusive in Didelphimorphia (based on early studies in the Virginia opossum). Further, he observed that the excitable cortex in all five species is coextensive with a well developed layer IV, indicative of sensory cortex, which suggested that motor cortex overlapped somatosensory cortex.
In the early sixties, Lende (1963a; 1963b) used evoked potential recordings and somewhat more refined methods of cortical stimulation to explore sensory and motor cortex in the Virginia opossum and the wallaby. His results confirmed the presence of motor representation for the hindlimb and tail on the medial wall of the neocortex in both species (Orders Didelphimorphia and Diprotodontia, respectively). Unlike the earlier studies in marsupials, Lende demonstrated that the neocortex did not contain a separate or partially overlapping representation of motor movements rostral to somatosensory cortex. Rather, he demonstrated that these animals have a complete overlap of the primary somatosensory area and the primary motor area, which he called a sensorimotor amalgam (Lende, 1963a). At the time, the results from his study were surprising, but seemed to agree with data from Vogt and Vogt (1906), as well as with histological data suggesting that motor cortex had a large granular cell layer (Abbie, 1940; Gray, 1924; Loo, 1931). These findings spurred a number of important ideas regarding cortical field evolution in mammals. Most notably, Lende (1969) proposed that marsupials in general represent a primitive state of cortical organization in which the homologous areas of S1 and M1, as described in placental mammals, are completely overlapping in marsupials, and that the evolution of this region in placental mammals is marked by a progressive separation of these fields, ultimately into two complete and separate sensory (S1) and motor (M1) representations (Figure 8A).
Figure 8.
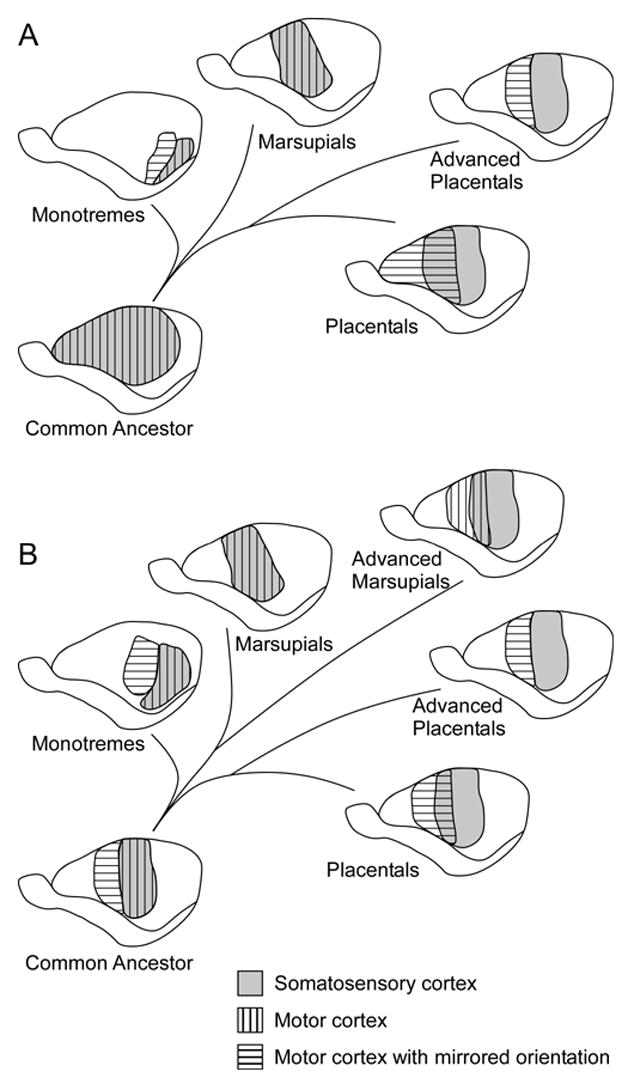
A diagram of the two prominent hypotheses on the evolution of sensorimotor cortex in mammals. (A) Lende (1969) proposed that marsupials represent a primitive state of cortical organization in which S1 (gray) and M1 (striped) are completely overlapping and that the evolution of this region is marked by a progressive separation of these fields in placental mammals, ultimately into two complete and separate sensory (S1) and motor (M1) representations. (B) Haight and Neylon (1979) proposed that there are three types of sensory motor organization. (1) In monotremes (and potentially the common ancestor), there is a dual motor representation with one being co-extensive with S1 and possessing the same orientation (vertical stripes) and the other occupying an anterior, mirrored position (horizontal stripes). (2) In marsupials and placental edentates, S1 and M1 have the same orientation (vertical stripes). In less specialized marsupials and placental edentates, there is a complete overlap of S1 and M1; in advanced marsupials, there is only a partial overlap of S1 and M1. (3) In placental mammals, the body representations of S1 and M1 are mirror images of one another (horizontal stripes). In less specialized placentals, there is a partial overlap of S1 and M1; in advanced placental mammals, S1 and M1 are completely separate. Conventions as in previous figures.
Early observations by Lende have been supported by several subsequent cortical stimulation studies in marsupials, including those in brush-tailed possums (Rees and Hore, 1970), white-eared opossums (Magalhaes-Castro and Saraiva, 1971), and short-tailed opossums (Frost et al., 2000). However, in the short-tailed opossum, only movements of the jaw and vibrissae could be evoked in the sensorimotor amalgam. The finding that no other body part movements (particularly forepaw movements) could be evoked anywhere in the motor cortex of short-tailed opossums is surprising in light of earlier studies which demonstrate that dense corticospinal projections originate in the S1 region of this species (Nudo and Masterton, 1990).
Like cortical stimulation studies, neuroanatomical studies indicate that S1 and M1 in marsupials are partially overlapping, but the data are equivocal with respect to the completely overlapping sensorimotor amalgam. For example, in the Virginia opossum, projections from VL (associated with the motor system) and VP (associated with the somatosensory system) converge on S1 (Killackey and Ebner, 1973). Furthermore, the projections of VL are restricted to S1, supporting the idea of a completely overlapping sensorimotor amalgam (Donoghue and Ebner, 1981a). Similarly, in the quokka wallaby (Setonix brachyurus), the Western gray kangaroo, and the red kangaroo (Megaleia rufa), retrograde degeneration studies indicate that motor cortex lies in the postorbital area, presumably in the sensorimotor amalgam (Watson, 1971a; b),
On the other hand, in the Tasmanian wombat (Johnson et al., 1973), the Tasmanian pademelon (Thylogale billardierii; Weller et al., 1977), and the brush-tailed possum (Haight and Neylon, 1978; 1979), there is a region of cortex located anterior to the proposed sensorimotor amalgam in which neurons do not respond to somatosensory stimulation and do not receive inputs from VP. Further, in the brush-tailed possum, thalamocortical projections terminate in three distinct zones (Haight and Neylon, 1979; Joschko and Sanderson, 1987). Cortex at the far rostral pole receives inputs from the mediodorsal nucleus (MD), generally associated with frontal cortex in placental mammals. Agranular cortex just caudal to this frontal region and rostral to S1, in the location of M1 in placental mammals, receives input predominantly from VL and receives less dense inputs from VP. Projections to S1, as described above, are predominantly from VP and Po, and sparsely from VL. Because the Tasmanian wombat, the Tasmanian pademelon, and the brush-tailed possum, are in the order Diprotodontia, Haight and Neylon (1979) have proposed that, unlike the sensorimotor amalgam in Didelphimorphia, Diprotodontia do not have a complete overlap of the sensory and motor areas. Instead, they hypothesize that there are three types of sensory/motor organization in mammals (Figure 8B; Haight and Neylon, 1979). First, in monotremes, there are two motor representations with one representation being co-extensive with S1 and possessing the same orientation and the other occupying an anterior, mirrored position (Lende, 1964), although alternative theories on monotreme motor cortex have since been proposed (Bohringer and Rowe, 1977; Hassiotis et al., 2004; Krubitzer et al., 1995). Second, in marsupials and placental edentates, S1 and M1 have the same orientation. In less specialized marsupials and placental edentates, there is a complete overlap of S1 and M1, and in advanced marsupials, there is only a partial overlap of S1 and M1. Finally, in placental mammals, the body representations of S1 and M1 are mirror images of one another. In less specialized placentals, there is a partial overlap of S1 and M1, and in advanced placental mammals, S1 and M1 are completely separate. Haight and Neylon (1979) also suggest that the partially separated S1 and M1 in Diprotodontia and the fully separated S1 and M1 in placental mammals may represent evolutionary convergence.
This hypothesis is supported in part by comparative studies of Nudo and Masterton (1990) in which retrograde tracers were injected into the spinal cord of a number of different mammals. Their results indicate that the major source of spinal output from the cortex forms a similar pattern of projections in all mammals investigated, including several marsupial species. Specifically there are two to three corticospinal projection zones (termed A, B, and C) that all mammals have. One of these projection zones (A) appears to correspond to somatosensory and motor cortex. Although no direct correlations have been made, projection zone A in the Virginia opossum and the short-tailed opossum appears to be restricted to somatosensory cortex, while this same projection zone in rodents and primates has a rostral extension into the location of motor cortex.
When considered together, results from cortical stimulation and neuroanatomical studies in marsupials are somewhat conflicting. Early cortical stimulation studies in marsupials indicate that motor cortex is at most partially overlapping with somatosensory cortex, while later stimulation studies indicate that motor cortex is completely overlapping with somatosensory cortex, but only in certain species. Several possible considerations could account for this discrepancy. The first is that earlier studies used gross stimulation techniques in which the spread of current was large, well beyond the site of cells that would actually evoke a movement. Thus, both the rostral and caudal extent of motor cortex was overestimated utilizing these techniques. A second consideration is the effect of anesthetics on marsupials. In our experience and that of other investigators (e.g., see Aitkin et al., 1986), a number of anesthetics commonly used on placental mammals have adverse effects on the responsiveness of neurons in marsupial neocortex. Thus, regions that would normally evoke movements when stimulated are silent and/or thresholds for evoking movement are raised, and therefore the responsive area is reduced in size. Finally, it should be noted that studies in which the data appear to support a complete overlap have been performed in two relatively primitive species, the Virginia opossum and the short-tailed opossum, both from the order Didelphimorphia, and the restriction of thalamic projections from motor nuclei of the thalamus to S1 has been observed only in the Virginia opossum. Thus, it may be that the order Didelphimorphia represents a more primitive state and that more advanced Australian radiations have independently evolved motor cortex organization that closely resembles that of advanced placental mammals. This evolution could be driven by the diversity of morphological specializations, lifestyles, and behaviors that have independently occurred in Australian marsupials which parallel the diversity of lifestyles and behaviors that arose in placental mammals.
In summary, the state of motor cortex is contentious and the data support two types of organization. The first type is exhibited in relatively primitive marsupials that have retained a sensorimotor amalgam. The second type of organization is found in more advanced marsupials which have evolved a partial separation of sensory and motor representations. These two types of organization indicate that motor cortex has evolved differently in separate phylogenetic lineages, and this divergence is likely related to selection pressures experienced by different groups. Neuroanatomical and connectional data support the observation that two types of motor cortex organization exist within marsupials. Despite these differences within the marsupial lineage, there are strong anatomical similarities between marsupials and placental mammals, in that both groups appear to have representatives of the two distinct types of motor cortex organization. This suggests that an independent evolution of distinct motor fields in some groups occurred from an initial amalgamated state.
5. Visual cortex
Although visual cortex had been defined neuroanatomically since the early 1900’s (e.g., Bodian, 1935; Brodmann, 1909; Diamond and Utley, 1963; Goldby, 1943; Gray, 1924; Packer, 1941), Lende (1963c) was the first to functionally define visual cortex using evoked potential techniques in the Virginia opossum. Since these first descriptions of visual cortex, the organization of the primary visual area (V1) has been examined using electrophysiological recording techniques in a several different marsupials, including the Virginia opossum (Christensen and Hill, 1970a; b; Rocha-Miranda et al., 1973), the white-eared opossum (Magalhaes-Castro and Saraiva, 1971), the big-eared opossum (Didelphis aurita; Sousa et al., 1978; Volchan et al., 1988), the brush-tailed possum (Crewther et al., 1984), the Tammar wallaby (Vidyasagar et al., 1992), the Northern quoll (Rosa et al., 1999), and the short-tailed opossum (Kahn et al., 2000). As in other mammals, V1 is located at the caudal pole of the occipital lobe and contains a complete visuotopically organized representation of the contralateral visual field (Figure 9). As in all other mammals, the representation of the upper visual quadrant is located caudolaterally in cortex, the lower visual quadrant representation is located rostromedially in cortex, the horizontal meridian bisects the upper and lower visual quadrants, and the vertical meridian forms the rostrolateral boundary of V1 (Kahn et al., 2000; Rosa et al., 1999; Sousa et al., 1978; Vidyasagar et al., 1992). While some studies demonstrate that cortex immediately rostral to V1 contains neurons responsive to visual stimulation, the second visual area (V2) has only been mapped in detail in the Northern quoll (Figure 9D; Rosa et al., 1999). As in placental mammals, V2 contains a complete representation of the visual hemifield with the vertical meridan represented at the caudomedial border of the field, adjacent to the vertical meridian border of V1. The horizontal meridian bisects the field. The lower visual quadrant is represented rostromedially and the upper visual quadrant is represented caudolaterally.
Figure 9.
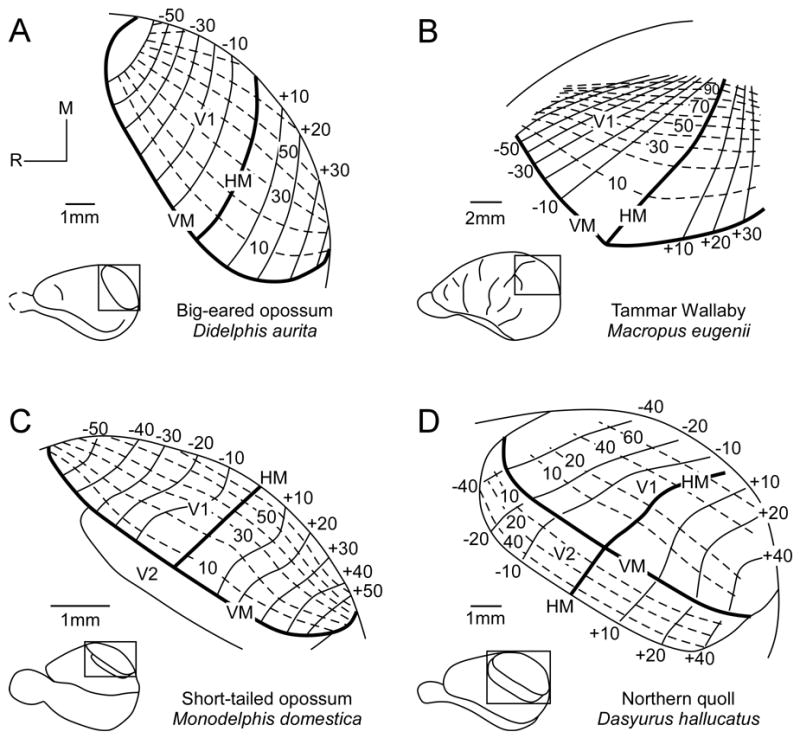
The visuotopic organization of V1 has been described in the big-eared opossum (A), the Tammar wallaby (B), the short-tailed opossum (C), and the Northern quoll (D). As in all other mammals, the representation of the upper visual quadrant is located caudolaterally in cortex, and the lower visual quadrant representation is located rostromedially. The horizontal meridian bisects the upper and lower visual quadrants, and the vertical meridian forms the rostrolateral boundary of V1. V2 has only been described in detail in the Northern quoll (D). The vertical meridan is represented at the caudomedial border of V2, adjacent to the vertical meridian of V1. The horizontal meridian bisects the field. The lower visual quadrant is represented rostromedially and the upper visual quadrant is represented caudolaterally. Numbers denote degrees in either elevation or azimuth; solid lines represent isoelevations; dashed lines represent isoazimuths. HM, horizontal meridian; VM, vertical meridian; Conventions as in previous figures; (A) based on Sousa et al., 1978; (B) based on Vidyasagar et al., 1992; (C) based on Kahn et al., 2000; and (D) based on Rosa et al., 1999.
Response properties of neurons in V1 of marsupials vary between species but generally resemble those of placental mammals. For example, studies of the response properties of neurons in V1 of the brush-tailed possum (Crewther et al., 1984) demonstrate that about one third of the neurons are orientation selective, most of which are directionally selective, and that about 65% of recorded cells are binocularly driven. Receptive fields for neurons range in size from one degree to eleven degrees based on eccentricity. In the Virginia opossum, cells have characteristic “on”, “off” or “on/off” responses, receptive fields are much larger than in the brush-tailed possum with a mean diameter of 19 degrees, and only about 30% of the cells are binocular (Christensen and Hill, 1970a; b). In the big-eared opossum, response properties of neurons are much like those described in placental mammals in that some neurons are orientation selective, some have antagonistic surrounds, and some are involved in processes such as contour integration (Oliveira et al., 2002; Rocha-Miranda et al., 1973). Finally, in the Tammar wallaby, one of the three diurnal marsupial species, response properties of neurons are much like those observed in cats and monkeys in that neurons are highly orientation selective, directional, and tuned to particular spatial frequencies (Ibbotson and Mark, 2003; Ibbotson et al., 2005).
In studies in which electrophysiological recordings have been combined with architectonic analysis (Kahn et al., 2000; Rosa et al., 1999; Vidyasagar et al., 1992), V1 is demonstrated to be architectonically distinct from V2, and its appearance is similar to that described for V1 in placental mammals. Specifically, in Nissl stained tissue V1 has a striated appearance with a densely packed, thick layer IV, and a moderately packed layer VI (Kahn et al., 2000; Vidyasagar et al., 1992). In cortex that is sectioned tangentially and processed for myelin (Kahn et al., 2000; Rosa et al., 1999) or CO (Rosa et al., 1999; also see Martinich et al., 2000; Martinich et al., 1990), V1 stains very darkly. In Northern quolls, the only marsupial in which V2 has been functionally defined, V2 stains lightly for both myelin and CO (Rosa et al., 1999). However architectonic studies indicate that in favorable preparations, V2 is actually composed of heterogeneous light and dark patches in CO stained tissue (Martinich et al., 2000; Martinich et al., 1990).
Architectonic studies that were not done in conjunction with electrophysiological recording have also demonstrated the distinct appearance of V1, similar to observations in placental mammals. For example, in both the Virginia opossum and short-tailed opossum V1 stains very darkly for NADPHd (Franca et al., 2000). In the Parma wallaby (Macropus parma), the staining pattern of zinc-containing neuropil (a possible neuromodulator) in V1 is similar to that observed in placental mammals (Garrett et al., 1994). In the Tammar wallaby, V1 stains darkly in layer IV for CO, lightly in layers III, IV, and V for acetylcholinesterase, and darkly for NADPHd in layers IV and upper VI (Ashwell et al., 2005). Finally, studies of pyramidal cell morphology in V1 indicate that the size and appearance of pyramidal cells in the fat-tailed dunnart and quokka wallaby are similar to those observed in macaque monkeys (Tyler et al., 1998).
The cortical connections of V1 are similar in all species examined. For example, in the Virginia opossum (Benevento and Ebner, 1971a; Ebner, 1967), brush-tailed possum (Crewther et al., 1984), mouse opossum (Marmosa elegans; Bravo et al., 1990), Tammar wallaby (Sheng et al., 1990), big-eared opossum (Martinich et al., 2000), and short-tailed opossum (Kahn et al., 2000), V1 is densely connected with V2 (peristriate cortex), cortex just lateral to V2 in multimodal cortex (parietal cortex and PP), the caudotemporal area (CT), posterolateral peristriate cortex, and perirhinal cortex. Connections of V1 are found throughout ipsilateral V2 and CT, but the connections with V2 are patchy and appear to be related to heterogeneities identified using CO stains (Martinich et al., 2000; see also Martinich et al., 1990). Studies of the overall pattern of commissural connections also demonstrate a patchy distribution with V1 and V2 of the opposite hemisphere (Bravo et al., 1990; Cusick and Kaas, 1986; Martinich et al., 2000), indicating that V2 may be modularly organized as it is in primates (e.g., Collins et al., 2001).
Thalamocortical connections of V1 in marsupials are similar to those described in a number of placental mammals. In the brush-tailed possum (Haight et al., 1980; Packer, 1941; Sanderson et al., 1980), the Virginia opossum (Benevento and Ebner, 1971b; Coleman and Clerici, 1981; Coleman et al., 1977), the big-eared opossum (Linden and Rocha-Miranda, 1983), the Tammar wallaby (Sheng et al., 1990; 1991), and the short-tailed opossum (Karlen et al., 2006), the primary source of input to V1 is from the dorsal division of the lateral geniculate nucleus (LGd). In all of these marsupials, moderate inputs also arise from the lateral posterior nucleus (LP). In the Virginia opossum (Coleman and Clerici, 1981; Coleman et al., 1977) and the brush-tailed possum (Haight et al., 1980), the primary source of input to V2 is from LP, with sparse input from LGd. In the Virginia opossum, the lateral intermediate nucleus (Li) also projects sparsely to V2.
Taken together, the data indicate that visual cortex in marsupials contains at least two cortical areas, V1 and V2. Each field is topographically organized similar to that in placental mammals. Furthermore, features of neural response properties, histochemical appearance, cortical and subcortical connections, and even the presence of a modular organization are common features of visual cortex across all mammals.
6. Auditory cortex
Although subcortical auditory pathways in marsupials have been well studied (see Aitkin, 1995 for review), less is known about the organization and connections of auditory cortex. Early evoked potential studies in the Virginia opossum revealed the presence and location of cortex that is responsive to auditory stimulation (Lende, 1963a; c; Magalhaes-Castro and Saraiva, 1971; Nieder and Randall, 1964). Modern electrophysiological studies have verified the location of auditory cortex in marsupials, but little is known about the number of cortical auditory fields or the detailed organization of auditory cortex across species. Most of what we do know comes from work on two Australian marsupials, the Northern quoll (previously called the native cat) and the brush-tailed possum. In the Northern quoll, detailed maps of the primary auditory area (A1) have been generated by Aitkin and colleagues (1986), and the results from these studies indicate that A1 is tonotopically organized, as in placental mammals. Specifically, isofrequency bands run mediolateral in A1, with high frequencies represented rostrally and low frequencies represented caudally (Figure 10). In these animals, there is a disproportionate representation of high frequencies, and neurons in this high frequency representation have very low thresholds (Aitkin et al., 1986). Studies of adult and juvenile Northern quoll vocalization frequencies indicate that most adult vocalizations occur at the lower end of the hearing range, between 0.4 and 2 kHz, but that juveniles have calls within the peak frequencies of adult hearing, between 6 and 12 kHz (Aitkin et al., 1994). Moreover, vocalizations and other sounds made by predators and prey of this nocturnal hunter have peak energy at frequencies less than 6 kHz and greater than 10 kHz, suggesting that the cortical magnification of high frequencies in the Northern quoll is important for detecting the location of predators and prey (Aitkin et al., 1994). Another auditory field just rostral to A1 has been identified in the Northern quoll, but the data on this field are relatively sparse (Aitkin et al., 1986). However, the lack of electrophysiological support for multiple auditory fields should be interpreted with caution since it has been suggested that auditory neurons in the Northern quoll may be highly susceptible to anesthetic influences (Aitkin et al., 1986).
Figure 10.
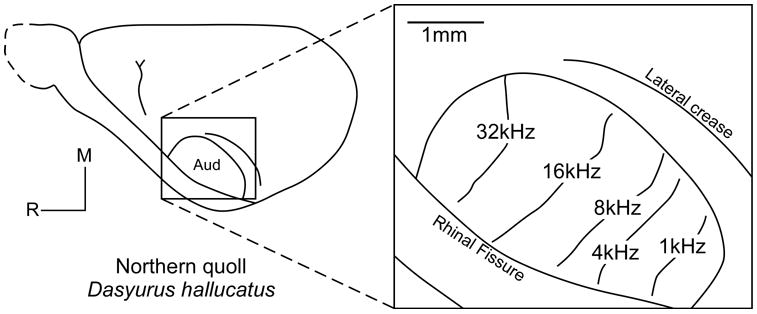
In the Northern quoll, A1 is tonotopically organized. Isofrequency bands run mediolaterally with high frequencies represented rostralmedially and low frequencies represented caudally. Another auditory field just rostral to A1 was also partially identified in this study, but the data were sparse. Conventions as in previous figures; based on Aitkin et al., 1986.
The orientation of A1 in the brush-tailed possum is different than that observed in Northern quolls and other mammals in that high frequencies are represented dorsally and low frequencies are represented ventrally (Gates and Aitkin, 1982). However, as in the Northern quoll, the lowest thresholds for neurons in A1 in the brush-tailed possum falls between 12–21 kHz, similar to the range of frequencies used in vocalizations of the Virginia opossum, the only marsupial that has been examined for a behavioral audiogram (Ravizza et al., 1969).
Several early studies describe the architectonic features of cortex in the location of auditory cortex (Abbie, 1942; Gray, 1924; Loo, 1931), but only in the Northern quoll have architectonic features been directly related to functional boundaries of A1 (Aitkin et al., 1986). In this study, Nissl stains revealed that the physiologically identified A1 is coextensive with a wide, densely packed granule cell layer (layer IV). In other studies where the cortex has been flattened and cut parallel to the cortical surface, cortex in which neurons are highly responsive to auditory stimulation stains very darkly for myelin and moderately for CO. This occurs in a variety of marsupials, including the Virginia opossum, brush-tailed possum, Northern quoll, striped possum, and short-tailed opossum (Beck et al., 1996; Catania et al., 2000; Elston and Manger, 1999; Huffman et al., 1999). This region is similar in appearance and location to A1 described in placental mammals (e.g., Luethke et al., 1988; Merzenich et al., 1976).
Thalamocortical and corticocortical connections of A1 have been examined in the Virginia opossum (Bodian, 1942; Diamond and Utley, 1963; Kudo et al., 1986), the brush-tailed possum (Aitkin and Gates, 1983; Goldby, 1943; Neylon and Haight, 1983), and the Northern quoll (Kudo et al., 1989). In the Virginia opossum, A1 receives projections from the anterior two-thirds of the medial geniculate nucleus (MG), and sparse projections from the posterior MG and the suprageniculate nucleus (SG; Bodian, 1942; Diamond and Utley, 1963; Kudo et al., 1986; Ravizza and Masterton, 1972). In the brush-tailed possum, A1 receives projections from three divisions of the MG: the dorsomedial MG, the ventromedial MG, and the lateral MG, as defined by Aitkin and Gates (1983). Since injections into different best-frequency representations in the cortex produce discrete bands of label in MG, these results indicate that projections from MG to A1 are tonotopically organized (Aitkin and Gates, 1983). In addition to projections from the MG, A1 in the brush-tailed possum also receives input from the SG (Neylon and Haight, 1983). Finally, in the Northern quoll, similar patterns of projections from MG have been observed after injections into A1 (Kudo et al., 1989). However, the densest projections appear to be from the ventral MG.
While ipsilateral cortical connections of A1 have not been described for any marsupial, dense contralateral projections from injections into homotopic locations in A1 have been described for both the brush-tailed possum and the Northern quoll (Aitkin and Gates, 1983; Kudo et al., 1989). Further, in the Northern quoll, A1 has been reported to connect ipsilaterally with the putamen and bilaterally with the lateral amygdala (Kudo et al., 1989). Direct connections between the amygdala and A1 have not been reported for any placental mammal, and may be a specialization of the Northern quoll or may represent a primitive pathway of mammals that has been subsequently lost in placental mammals with the addition of more auditory cortical fields. Support for the latter supposition comes from recent studies in marmoset monkeys which indicate that higher order auditory areas, such as the rostromedial field, have ipsilateral connections with the lateral amygdala (de la Mothe et al., 2006).
While information on the organization and connections of auditory cortex in marsupials is relatively sparse, existing data indicate that there are features of organization that are similar to that described in placental mammals, and that the cortical magnification of specific frequencies appears to be related to behaviorally relevant aspects of their lifestyle.
7. What can marsupials tell us about cortical evolution?
Comparative studies of sensory and motor neocortex in a number of marsupials indicate that despite differences in lifestyle, size, and phylogeny, all marsupials share a common plan of cortical organization (Figure 11). This plan includes a constellation of cortical fields that have distinct architecture, functional organization, and patterns of connectivity. For somatosensory cortex, S1 and S2 are present in all marsupials investigated (Figure 11), as well as in placental and monotreme mammals (Figure 12). This suggests that these fields arose very early in mammalian evolution and have been retained in all lineages. Cortical areas such as PV, SR, and SC are present in most of the marsupials studied, but in some species these fields are more primitive and poorly developed, such as in the Virginia opossum and short-tailed opossum (Figure 11A, B).
Figure 11.
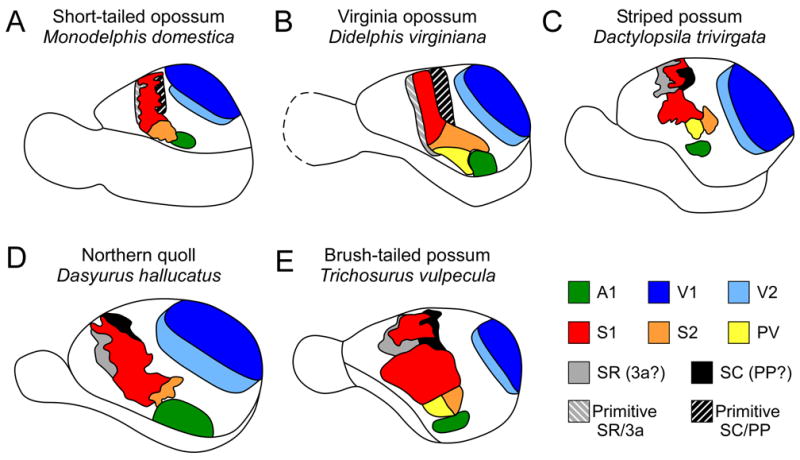
A summary of sensory neocortical organization in five marsupial species. Visual areas V1 (dark blue) and V2 (light blue) have been identified in all examined marsupials (A–E); however V2 has only been fully described in the Northern quoll (D). Similarly, the primary auditory area A1 (green) has been identified in all examined marsupials (A–E) and all examined mammals; however is has only been described in detail in the Northern quoll and brush-tailed possum (D,E). Somatosensory areas S1 (red) and S2 (orange) have been identified in all examined marsupials (A–E), and all examined mammals. In some species S2 has been subdivided in to S2 and PV (yellow; B, C, E). Two additional fields, SR (gray) and SC (black) have also been identified (A–E), although in some species, such as the short-tailed opossum (A) and the Virginia opossum (B), these fields are more primitive (Primitive SR (gray striped) and Primitive SC (black striped)). Based on several factors, we have proposed that the rostral field (SR) may be homologous to area 3a of primates and flying foxes, and the rostral somatosensory area of rodents and that the caudal field (SC) may correspond to posterior parietal area (PP; formerly PM) in squirrels and posterior parietal cortex (PPC) in rats, which is hypothesized to be homologous to one or more of the posterior parietal fields identified in primates, such as area 5 (Krubitzer and Padberg, 2007). However, this hypothesis has not been tested. Conventions as in previous figures; (A) based on Huffman et al., 1999, Frost et al., 2000, Kahn et al., 2000; (B) based on Beck et al., 1996; (C) based on Huffman et al., 1999; (D) based on Rosa et al., 1999, Huffman et al., 1999, Aitkin et al., 1986; and (E) based on Elston and Manger, 1999, Huffman et al., 1999.
Figure 12.
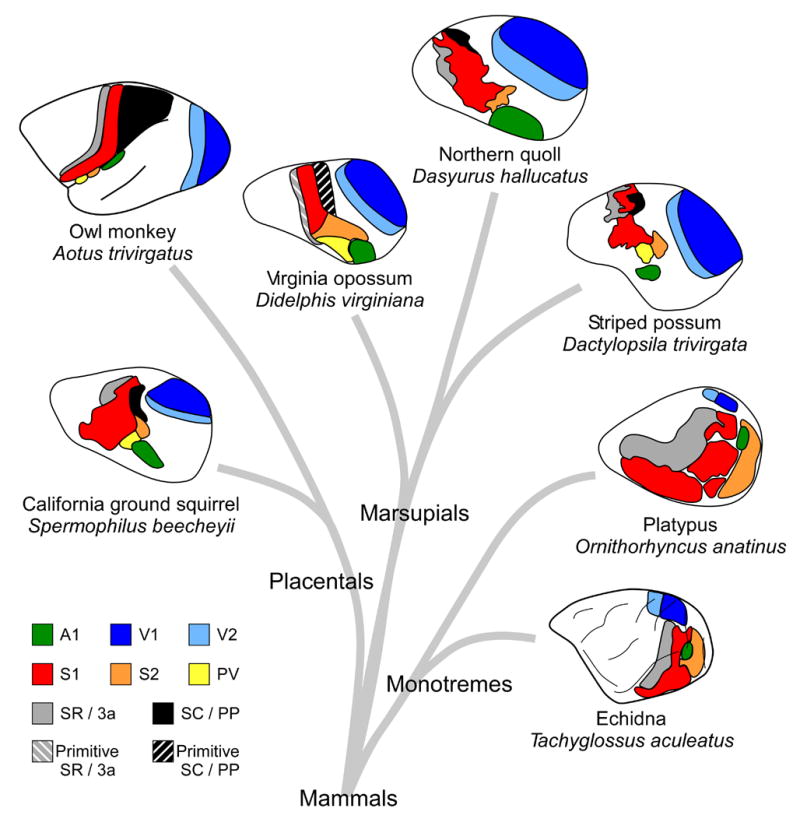
There are striking similarities in the organization of sensory neocortex across mammals. Shown here are examples of placental, marsupial, and monotreme neocortical organization. Somatosensory areas S1 (red) and S2 (orange) have been identified in all examined mammals, and in some species S2 has been subdivided in to S2 and PV (yellow). Two additional fields, SR (3a in placental mammals; gray) and SC (PP in placental mammals; black) have also been identified in placental and marsupial mammals, and SR has been identified in monotremes. In some species, such as the Virginia opossum, these fields are more primitive (Primitive SR (gray striped) and Primitive SC (black striped)). Further, in some animals, such as primates, regions of cortex that likely contain only a single field in marsupials and some placental mammals (e.g. squirrels), have expanded and contain multiple cortical areas. Visual areas V1 and V2, and auditory area A1 have also been identified in all mammals examined. In the owl monkey, A1 is on the lower bank of the lateral sulcus, and S2 and PV are on the upper bank of the lateral sulcus. Conventions as in previous figures; Virginia opossum based on Beck et al., 1996; striped possum and Northern quoll based on Huffman et al., 1999; owl monkey based on Kaas, 2004a; platypus and echidna based on Krubitzer et al., 1995, Krubitzer, 1998; California ground squirrel based on Slutsky et al., 2000.
For visual cortex, V1 and V2 have been identified in all marsupial and placental mammals studied, indicating that both fields were present in the common ancestor (Figures 11 and 12). However, V2 is less well developed in some species than in others. V1 has been identified in monotremes, but V2 does not appear to be present in this group. This suggests that V2 arose after the monotreme radiation.
Although less is known about the organization of auditory cortex, A1 has been identified (either electrophysiologically and/or architectonically) in all marsupials investigated (Figure 11), as well as in placental and monotreme mammals (Figure 12). The ubiquity of this field in all subclasses indicates that it was present in the common ancestor.
Finally, the status of motor cortex in marsupials is contentious. However, when all data are examined it appears that two types of organization exist (Figure 8). The first is represented in primitive marsupials, such as in the order Didelphimorphia. In these animals there appears to be a complete sensorimotor overlap in that S1 is coextensive with M1. In more advanced marsupials, such as in the order Diprotodontia, S1 and M1 only partially overlap. The state of organization in the common ancestor of all mammals is difficult to deduce from existing data. However, when comparing data across species we believe that the common ancestor had a complete sensorimotor overlap. In the three species of extant monotremes, all of which are highly derived, these fields became distinct. In early marsupials, this overlap was inherited from the common ancestor, and in some advanced marsupial lineages, these fields have become more distinguished and now only partially overlap. We propose that in early placental mammals a sensory motor overlap existed, and still exists for some primitive groups, such as insectivores. In other groups, such as rodents, these areas are partially separated, and in some groups, such as primates, these areas have completely separated.
Alternatively, it has been argued that one of the hallmarks of primate evolution has been the expansion of motor cortex (Kaas, 2004b). Kaas and colleagues have argued that the sensorimotor amalgam, defined as the combination of S1 and M1, may instead only be an enhanced S1 with more advanced motor features (Beck et al., 1996), since S1 has traditionally been defined as having a small motor component (i.e., Woolsey, 1958). If this is the case, it would suggest that monotremes and marsupials do not possess a separate motor region. Instead, motor cortex is a specialized field that evolved in placental mammals and has become particularly derived in primates. Although this theory does not address the issue of a partially separated motor area in advanced marsupials (i.e., Johnson et al., 1973; Weller et al., 1977; Haight and Neylon, 1978; 1979), it does agree with data from more primitive marsupials (see Beck et al., 1996 for more detail on this theory).
Although similarities exist in the organization of the neocortex across all mammalian species (Figure 12), there are several differences in cortical field organization that relate to species-specific lifestyle. The first is a difference in the amount of cortex devoted to a particular sensory system. For example, in the Northern quoll (Figure 11D), visual cortex and auditory cortex are relatively large compared to the size of somatosensory cortex. This may be related to the Northern quoll’s carnivorous lifestyle; it is an agile hunter and likely coordinates both auditory and visual information to capture prey. Conversely, the brush-tailed possum, which is a phalanger with a well developed hand, typically eats leaves and flowers and has a larger somatosensory cortex relative to visual and auditory cortex (Figure 11E). The second difference between species is the cortical magnification of behaviorally relevant sensory surfaces. For example, in the striped possum the representation of D4 and tongue are relatively large compared to other body part representations within S1, and compared to similar representations in S1 of other species (Figure 5D). In the striped possum, D4 and the tongue are both used to extract insects from holes in the bark of trees. In the Tammar wallaby, one of only three diurnal marsupials, an enlarged representation of central vision is present within V1, as compared to nocturnal marsupials (Figure 9B). Finally, frequency representations between 8 and 32 kHz are relatively large in the Northern quoll (Figure 10), and these frequencies match the vocalization frequencies of predators and prey of this species.
The types of adaptations that have been made to cortical fields in marsupials are remarkably similar to those described in placental mammals. This is not surprising given that marsupials, like placental mammals, have undergone significant radiations, and have diversified greatly, particularly in Australia. This diversification consists of a wide array of morphological, behavioral, and cortical specializations. Many of the morphological and behavioral specializations are associated with patterns of activity (diurnal versus nocturnal), habitat selection (arboreal, terrestrial, aquatic, or subterranean), and diet (carnivorous, herbivorous, omnivorous, or frugivorous). In fact, striking similarities in lifestyle and behavior have been described for marsupials from the order Diprotondontia and primates (Rassmussen and Sussman, 2007). These include similarities in body size, eye placement, activity patterns, encephalization quotients, jaw structure, and hand structure, to name a few; all of which have evolved independently in marsupials but parallel similar changes in placental mammals. The similarities in cortical field organization that have been described for both groups indicate that there are large constraints imposed on evolving nervous systems that result in recurrent solutions to similar environmental challenges.
Acknowledgments
This work was supported by a McDonnell Foundation grant and an NINDS award (R01-NS35103) to Leah Krubitzer, and by an NSF fellowship (DG-0202740) to Sarah Karlen.
Abbreviations
- A1
primary auditory area
- CIN
central intralaminar nucleus
- CL
central lateral nucleus
- CO
cytochrome oxidase
- CT
caudotemporal area
- D4
fourth digit
- DH
dorsal head
- Fl
forelimb
- Fp
forepaw
- Hl
hindlimb
- HRP
horseradish peroxidase
- LGd
dorsal lateral geniculate nucleus
- Li
lateral intermediate nucleus
- LP
lateral posterior nucleus
- M
medial
- M1
primary motor area
- MD
mediodorsal nucleus
- MG
medial geniculate nucleus
- MYA
million years ago
- NADPHd
nicotinamide adenine dinucleotide phosphate diaphorase
- PM
parietal medial area
- Po
posterior nucleus
- PP
posterior parietal area
- PPC
posterior parietal cortex
- PV
parietal ventral area
- R
rostral
- Rhin
rhinarium
- S1
primary somatosensory area
- S2
second somatosensory area
- SC
somatosensory caudal field
- SDH
succinic dehydrogenase
- SG
suprageniculate nucleus
- Sn
snout
- SR
somatosensory rostral field
- Ton
tongue
- Tr
trunk
- V1
primary visual area
- V2
second visual area
- Vib
vibrissae
- VL
ventrolateral nucleus
- VM
ventromedial nucleus
- VP
ventroposterior nucleus
- Wr
wrist
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Abbie AA. The excitable cortex in the monotremata. Aust J Exp Biol Med Sci. 1938;16:143–152. [Google Scholar]
- Abbie AA. The excitable cortex in Perameles, Sarcophilus, Dasyurus, Trichosurus and Wallabia (Macropus) J Comp Neurol. 1940;72:469–487. [Google Scholar]
- Abbie AA. Cortical lamination in a polyprotodont marsupial, Perameles nasuta. J Comp Neurol. 1942;76:509–536. [Google Scholar]
- Adey WR, Kerr DI. The cerebral representation of deep somatic sensibility in the marsupial phalanger and the rabbit; an evoked potential and histological study. J Comp Neurol. 1954;100:597–624. doi: 10.1002/cne.901000307. [DOI] [PubMed] [Google Scholar]
- Aitkin L. The auditory neurobiology of marsupials: a review. Hear Res. 1995;82:257–66. doi: 10.1016/0378-5955(94)00182-p. [DOI] [PubMed] [Google Scholar]
- Aitkin LM, Gates GR. Connections of the auditory cortex of the brush-tailed possum, Trichosurus vulpecula. Brain Behav Evol. 1983;22:75–88. doi: 10.1159/000121509. [DOI] [PubMed] [Google Scholar]
- Aitkin LM, Irvine DR, Nelson JE, Merzenich MM, Clarey JC. Frequency representation in the auditory midbrain and forebrain of a marsupial, the northern native cat (Dasyurus hallucatus) Brain Behav Evol. 1986;29:17–28. doi: 10.1159/000118669. [DOI] [PubMed] [Google Scholar]
- Aitkin LM, Nelson JE, Shepherd RK. Hearing, vocalization and the external ear of a marsupial, the northern Quoll, Dasyurus hallucatus. J Comp Neurol. 1994;349:377–88. doi: 10.1002/cne.903490305. [DOI] [PubMed] [Google Scholar]
- Arrese CA, Hart NS, Thomas N, Beazley LD, Shand J. Trichromacy in Australian marsupials. Curr Biol. 2002;12:657–60. doi: 10.1016/s0960-9822(02)00772-8. [DOI] [PubMed] [Google Scholar]
- Ashwell KW, Marotte LR, Li L, Waite PM. Anterior commissure of the wallaby (Macropus eugenii): adult morphology and development. J Comp Neurol. 1996a;366:478–94. doi: 10.1002/(SICI)1096-9861(19960311)366:3<478::AID-CNE8>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- Ashwell KW, Waite PM, Marotte L. Ontogeny of the projection tracts and commissural fibres in the forebrain of the tammar wallaby (Macropus eugenii): timing in comparison with other mammals. Brain Behav Evol. 1996b;47:8–22. doi: 10.1159/000113225. [DOI] [PubMed] [Google Scholar]
- Ashwell KW, Zhang LL, Marotte LR. Cyto- and chemoarchitecture of the cortex of the tammar wallaby (Macropus eugenii): areal organization. Brain Behav Evol. 2005;66:114–36. doi: 10.1159/000086230. [DOI] [PubMed] [Google Scholar]
- Beck PD, Pospichal MW, Kaas JH. Topography, architecture, and connections of somatosensory cortex in opossums: evidence for five somatosensory areas. J Comp Neurol. 1996;366:109–33. doi: 10.1002/(SICI)1096-9861(19960226)366:1<109::AID-CNE8>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- Benevento LA, Ebner FF. The areas and layers of corticocortical terminations in the visual cortex of the Virginia opossum. J Comp Neurol. 1971a;141:157–89. doi: 10.1002/cne.901410203. [DOI] [PubMed] [Google Scholar]
- Benevento LA, Ebner FF. The contribution of the dorsal lateral geniculate nucleus to the total pattern of thalamic terminations in striate cortex of the Virginia opossum. J Comp Neurol. 1971b;143:243–60. doi: 10.1002/cne.901430206. [DOI] [PubMed] [Google Scholar]
- Bodemer CW, Towe AL. Cortical localization patterns in the somatic sensory cortex of the opossum. Experimental Neurology. 1963;8:380–394. [Google Scholar]
- Bodian D. The projection of the lateral geniculate body on the cerebral cortex of the opossum, Didelphis virginiana. J Comp Neurol. 1935;62:469–494. [Google Scholar]
- Bodian D. Studies on the diencephalon of the Virginia opossum. III. The thalamo-cortical projection. J Comp Neurol. 1942;77:525–576. [Google Scholar]
- Bohringer RC, Rowe MJ. The organization of the sensory and motor areas of cerebral cortex in the platypus (Ornithorhynchus anatinus) J Comp Neurol. 1977;174:1–14. doi: 10.1002/cne.901740102. [DOI] [PubMed] [Google Scholar]
- Bravo H, Olavarria J, Martinich S. Patterns of interhemispheric and striate-peristriate connections in visual cortex of the South American marsupial Marmosa elegans (mouse opossum) Anat Embryol (Berl) 1990;182:583–9. doi: 10.1007/BF00186465. [DOI] [PubMed] [Google Scholar]
- Brodmann K. Vergleichende Lokalisationsiehre der Grosshirnrinde in ihren Prinzipien Dargestellt auf Grund des Zellenbaues. Leipzig: Barth; 1909. [Google Scholar]
- Cabana T, Martin GF. The development of commissural connections of somatic motor-sensory areas of neocortex in the North American opossum. Anat Embryol (Berl) 1985;171:121–8. doi: 10.1007/BF00319061. [DOI] [PubMed] [Google Scholar]
- Cardillo M, Bininda-Emonds ORP, Boakes E, Purvis A. A species-level phylogenetic supertree of marsupials. Journal of Zoology (London) 2004;264:11–31. [Google Scholar]
- Catania KC, Jain N, Franca JG, Volchan E, Kaas JH. The organization of somatosensory cortex in the short-tailed opossum (Monodelphis domestica) Somatosens Mot Res. 2000;17:39–51. doi: 10.1080/08990220070283. [DOI] [PubMed] [Google Scholar]
- Chapman HG. Note on cerebral localization in the bandicoot (Perameles) Proc Linn Soc NSW. 1906;31:493–494. [Google Scholar]
- Christensen BN, Ebner FF. The synaptic architecture of neurons in opossum somatic sensory-motor cortex: a combined anatomical and physiological study. J Neurocytol. 1978;7:39–60. doi: 10.1007/BF01213459. [DOI] [PubMed] [Google Scholar]
- Christensen JL, Hill RM. Receptive fields of single cells of a marsupial visual cortex of Didelphis virginiana. Experientia. 1970a;26:43–4. doi: 10.1007/BF01900381. [DOI] [PubMed] [Google Scholar]
- Christensen JL, Hill RM. Response properties of single cells of a marsupial visual cortex. Am J Optom Arch Am Acad Optom. 1970b;47:547–56. doi: 10.1097/00006324-197007000-00005. [DOI] [PubMed] [Google Scholar]
- Coleman GT, Zhang HQ, Murray GM, Zachariah MK, Rowe MJ. Organization of somatosensory areas I and II in marsupial cerebral cortex: parallel processing in the possum sensory cortex. J Neurophysiol. 1999;81:2316–24. doi: 10.1152/jn.1999.81.5.2316. [DOI] [PubMed] [Google Scholar]
- Coleman J, Clerici WJ. Organization of thalamic projections to visual cortex in opossum. Brain Behav Evol. 1981;18:41–59. doi: 10.1159/000121775. [DOI] [PubMed] [Google Scholar]
- Coleman J, Diamond IT, Winer JA. The visual cortex of the opossum: the retrograde transport of horseradish peroxidase to the lateral geniculate and lateral posterior nuclei. Brain Res. 1977;137:233–52. doi: 10.1016/0006-8993(77)90336-5. [DOI] [PubMed] [Google Scholar]
- Collins CE, Stepniewska I, Kaas JH. Topographic patterns of v2 cortical connections in a prosimian primate (Galago garnetti) J Comp Neurol. 2001;431:155–67. [PubMed] [Google Scholar]
- Crewther DP, Crewther SG, Sanderson KJ. Primary visual cortex in the brushtailed possum: receptive field properties and corticocortical connections. Brain Behav Evol. 1984;24:184–97. doi: 10.1159/000121316. [DOI] [PubMed] [Google Scholar]
- Cummings DM, Malun D, Brunjes PC. Development of the anterior commissure in the opossum: midline extracellular space and glia coincide with early axon decussation. J Neurobiol. 1997;32:403–14. [PubMed] [Google Scholar]
- Cunningham RH. Cortical motor centers of the opossum. J Physiol. 1898;9:355. doi: 10.1113/jphysiol.1898.sp000691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cusick CG, Kaas JH. Interhemispheric connections of cortical sensory and motor representations in primates. In: Lepore F, Ptito M, Jasper HH, editors. Two hemispheres - one brain: Functions of the corpus callosum. Vol. 17. Alan R. Liss, Inc; New York: 1986. pp. 83–102. [Google Scholar]
- de la Mothe LA, Blumell S, Kajikawa Y, Hackett TA. Cortical connections of the auditory cortex in marmoset monkeys: core and medial belt regions. J Comp Neurol. 2006;496:27–71. doi: 10.1002/cne.20923. [DOI] [PubMed] [Google Scholar]
- Diamond IT, Utley JD. Thalamic retrograde degeneration study of sensory cortex in opossum. J Comp Neurol. 1963;120:129–60. doi: 10.1002/cne.901200202. [DOI] [PubMed] [Google Scholar]
- Disbrow E, Roberts T, Krubitzer L. Somatotopic organization of cortical fields in the lateral sulcus of Homo sapiens: evidence for SII and PV. J Comp Neurol. 2000;418:1–21. doi: 10.1002/(sici)1096-9861(20000228)418:1<1::aid-cne1>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Donoghue JP, Ebner FF. The laminar distribution and ultrastructure of fibers projecting from three thalamic nuclei to the somatic sensory-motor cortex of the opossum. J Comp Neurol. 1981a;198:389–420. doi: 10.1002/cne.901980303. [DOI] [PubMed] [Google Scholar]
- Donoghue JP, Ebner FF. The organization of thalamic projections to the parietal cortex of the Virginia opossum. J Comp Neurol. 1981b;198:365–88. doi: 10.1002/cne.901980302. [DOI] [PubMed] [Google Scholar]
- Ebner FF. Afferent connections to neocortex in the opossum (Didelphis virginiana) J Comp Neurol. 1967;129:241–68. doi: 10.1002/cne.901290303. [DOI] [PubMed] [Google Scholar]
- Ebner FF. A comparison of primitive forebrain organization in metatherian and eutherian mammals. Ann N Y Acad Sci. 1969;167:241–257. [Google Scholar]
- Elston GN, Manger PR. The organization and connections of somatosensory cortex in the brush-tailed possum (Trichosurus vulpecula): evidence for multiple, topographically organized and interconnected representations in an Australian marsupial. Somatosens Mot Res. 1999;16:312–37. doi: 10.1080/08990229970384. [DOI] [PubMed] [Google Scholar]
- Flashman JF. A preliminary note on the motor areas in the cerebral cortex of marsupials. Rep Path Lab Dept Lunacy NSW. 1906;1:24–26. [Google Scholar]
- Foster RE, Donoghue JP, Ebner FF. Laminar organization of efferent cells in the parietal cortex of the Virginia opossum. Exp Brain Res. 1981;43:330–6. doi: 10.1007/BF00238375. [DOI] [PubMed] [Google Scholar]
- Franca JG, Volchan E, Jain N, Catania KC, Oliveira RL, Hess FF, Jablonka M, Rocha-Miranda CE, Kaas JH. Distribution of NADPH-diaphorase cells in visual and somatosensory cortex in four mammalian species. Brain Res. 2000;864:163–75. doi: 10.1016/s0006-8993(00)02058-8. [DOI] [PubMed] [Google Scholar]
- Frost SB, Milliken GW, Plautz EJ, Masterton RB, Nudo RJ. Somatosensory and motor representations in cerebral cortex of a primitive mammal (Monodelphis domestica): a window into the early evolution of sensorimotor cortex. J Comp Neurol. 2000;421:29–51. doi: 10.1002/(sici)1096-9861(20000522)421:1<29::aid-cne3>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- Garrett B, Osterballe R, Slomianka L, Geneser FA. Cytoarchitecture and staining for acetylcholinesterase and zinc in the visual cortex of the Parma wallaby (Macropus parma) Brain Behav Evol. 1994;43:162–72. doi: 10.1159/000113632. [DOI] [PubMed] [Google Scholar]
- Gates GR, Aitkin LM. Auditory cortex in the marsupial possum Trichosurus vulpecula. Hear Res. 1982;7:1–11. doi: 10.1016/0378-5955(82)90078-8. [DOI] [PubMed] [Google Scholar]
- Goldby F. An experimental investigation of the motor cortex and its connexions in the phalanger, Trichosurus vulpecula. J Anat. 1939;74:12–33. [PMC free article] [PubMed] [Google Scholar]
- Goldby F. An experimental study of the thalamus in the phalanger, Trichosurus vulpecula. J Anat. 1943;77:195–224. [PMC free article] [PubMed] [Google Scholar]
- Granger EM, Masterton RB, Glendenning KK. Origin of interhemispheric fibers in acallosal opossum (with a comparison to callosal origins in rat) J Comp Neurol. 1985;241:82–98. doi: 10.1002/cne.902410107. [DOI] [PubMed] [Google Scholar]
- Gray PA. The cortical lamination pattern of the opossum, Didelphys virginiana. J Comp Neurol. 1924;37:221–262. [Google Scholar]
- Gray PA, Turner EL. The motor cortex of the opossum. J Comp Neurol. 1924;36:375–385. [Google Scholar]
- Haight JR, Neylon L. The organization of neocortical projections from the ventroposterior thalamic complex in the marsupial brush-tailed possum, Trichosurus vulpecula: a horseradish peroxidase study. J Anat. 1978;126:459–85. [PMC free article] [PubMed] [Google Scholar]
- Haight JR, Neylon L. The organization of neocortical projections from the ventrolateral thalamic nucleus in the brush-tailed possum, Trichosurus vulpecula, and the problem of motor and somatic sensory convergence within the mammalian brain. J Anat. 1979;129:673–94. [PMC free article] [PubMed] [Google Scholar]
- Haight JR, Neylon L. An analysis of some thalamic projections to parietofrontal neocortex in the marsupial native cat, Dasyurus viverrinus (Dasyuridae) Brain Behav Evol. 1981;19:193–204. doi: 10.1159/000121642. [DOI] [PubMed] [Google Scholar]
- Haight JR, Sanderson KJ, Neylon L, Patten GS. Relationships of the visual cortex in the marsupial brush-tailed possum, Trichosurus vulpecula, a horseradish peroxidase and autoradiographic study. J Anat. 1980;131:387–412. [PMC free article] [PubMed] [Google Scholar]
- Haight JR, Weller WL. Neocortical topography in the brush-tailed possum: variability and functional significance of sulci. J Anat. 1973;116:473–474. [PubMed] [Google Scholar]
- Hassiotis M, Paxinos G, Ashwell KW. Cyto- and chemoarchitecture of the cerebral cortex of the Australian echidna (Tachyglossus aculeatus). I. Areal organization. J Comp Neurol. 2004;475:493–517. doi: 10.1002/cne.20193. [DOI] [PubMed] [Google Scholar]
- Heath CJ, Jones EG. Interhemispheric pathways in the absence of a corpus callosum. An experimental study of commissural connexions in the marsupial phalanger. J Anat. 1971;109:253–70. [PMC free article] [PubMed] [Google Scholar]
- Herrick CL. The cortical motor centres in lower mammals. J Comp Neurol. 1898;8:92–98. [Google Scholar]
- Herrick CL, Tight WG. The central nervous system of rodents. Bull Lab Denison Univ. 1890;5:35–95. [Google Scholar]
- Horovitz I, Sanchez-Villagra MR. A morphological analysis of marsupial mammal higher-level phylogenetic relationships. Cladistics. 2003;19:181–212. [Google Scholar]
- Huffman KJ, Nelson J, Clarey J, Krubitzer L. Organization of somatosensory cortex in three species of marsupials, Dasyurus hallucatus, Dactylopsila trivirgata, and Monodelphis domestica: neural correlates of morphological specializations. J Comp Neurol. 1999;403:5–32. doi: 10.1002/(sici)1096-9861(19990105)403:1<5::aid-cne2>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Ibbotson MR, Mark RF. Orientation and spatiotemporal tuning of cells in the primary visual cortex of an Australian marsupial, the wallaby Macropus eugenii. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2003;189:115–23. doi: 10.1007/s00359-002-0379-6. [DOI] [PubMed] [Google Scholar]
- Ibbotson MR, Price NS, Crowder NA. On the division of cortical cells into simple and complex types: a comparative viewpoint. J Neurophysiol. 2005;93:3699–702. doi: 10.1152/jn.01159.2004. [DOI] [PubMed] [Google Scholar]
- Iwaniuk AN, Nelson JE, Pellis SM. Do big-brained animals play more? Comparative analyses of play and relative brain size in mammals. J Comp Psychol. 2001;115:29–41. doi: 10.1037/0735-7036.115.1.29. [DOI] [PubMed] [Google Scholar]
- Jacobs GH, Rowe MP. Evolution of vertebrate colour vision. Clin Exp Optom. 2004;87:206–16. doi: 10.1111/j.1444-0938.2004.tb05050.x. [DOI] [PubMed] [Google Scholar]
- Jain N, Qi HX, Catania KC, Kaas JH. Anatomic correlates of the face and oral cavity representations in the somatosensory cortical area 3b of monkeys. J Comp Neurol. 2001;429:455–68. doi: 10.1002/1096-9861(20010115)429:3<455::aid-cne7>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Johnson JI. Central nervous system of marsupials. In: Hunsaker D, editor. The biology of marsupials. Academic Press, Inc; New York: 1977. pp. 157–278. [Google Scholar]
- Johnson JI. Comparative development of somatic sensory cortex. In: Jones EG, Peters A, editors. Cerebral Cortex. Plenum; New York: 1990. pp. 335–449. [Google Scholar]
- Johnson JI, Haight JR, Megirian D. Convolutions related to sensory projections in cerebral neocortex of marsupial wombats. J Anat. 1973;114:153. [Google Scholar]
- Jones EG. The Thalamus. Plenum Press; New York: 1985. [Google Scholar]
- Joschko MA, Sanderson KJ. Cortico-cortical connections of the motor cortex in the brushtailed possum (Trichosurus vulpecula) J Anat. 1987;150:31–42. [PMC free article] [PubMed] [Google Scholar]
- Kaas JH. Early visual areas: V1, V2, V3, DM, DL, and MT. In: Kaas JH, Collins CE, editors. The Primate Visual System. CRC Press; Boca Raton: 2004a. pp. 139–159. [Google Scholar]
- Kaas JH. Evolution of somatosensory and motor cortex in primates. Anat Rec A Discov Mol Cell Evol Biol. 2004b;281:1148–56. doi: 10.1002/ar.a.20120. [DOI] [PubMed] [Google Scholar]
- Kahn DM, Huffman KJ, Krubitzer L. Organization and connections of V1 in Monodelphis domestica. J Comp Neurol. 2000;428:337–54. [PubMed] [Google Scholar]
- Karlen SJ, Kahn DM, Krubitzer L. Early blindness results in abnormal corticocortical and thalamocortical connections. Neuroscience. 2006;142:843–58. doi: 10.1016/j.neuroscience.2006.06.055. [DOI] [PubMed] [Google Scholar]
- Killackey H, Ebner F. Convergent projection of three separate thalamic nuclei on to a single cortical area. Science. 1973;179:283–5. doi: 10.1126/science.179.4070.283. [DOI] [PubMed] [Google Scholar]
- Klima M, Maier W. Body Structure. In: Grzimek B, editor. Grzimek’s Encyclopedia of Mammals. McGraw-Hill; New York: 1990. pp. 58–85. [Google Scholar]
- Krubitzer L. The organization of neocortex in mammals: are species differences really so different? Trends Neurosci. 1995;18:408–17. doi: 10.1016/0166-2236(95)93938-t. [DOI] [PubMed] [Google Scholar]
- Krubitzer L. What can monotremes tell us about brain evolution? Philos Trans R Soc Lond B Biol Sci. 1998;353:1127–46. doi: 10.1098/rstb.1998.0271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krubitzer L, Hunt DL. Captured in the net of space and time: Understanding cortical field evolution. In: Kaas JH, Krubitzer L, editors. The Evolution of Nervous Systems in Mammals. IV. Academic Press; Oxford: 2006. pp. 49–72. [Google Scholar]
- Krubitzer L, Manger P, Pettigrew J, Calford M. Organization of somatosensory cortex in monotremes: in search of the prototypical plan. J Comp Neurol. 1995;351:261–306. doi: 10.1002/cne.903510206. [DOI] [PubMed] [Google Scholar]
- Krubitzer L, Padberg J. Evolution of parietal association areas of the neocortex in mammals. In: Butler A, editor. Encyclopedic Reference of Neuroscience. Springer; 2007. In press. [Google Scholar]
- Krubitzer LA, Kahn DM. The evolution of the human neocortex: Is the human brain fundamentally different than that of other mammals? In: Kanwisher JDN, Umilta C, editors. Attention and Performance XX, Functional Brain Imaging of Visual Cognition. 2004. [Google Scholar]
- Krubitzer LA, Sesma MA, Kaas JH. Microelectrode maps, myeloarchitecture, and cortical connections of three somatotopically organized representations of the body surface in the parietal cortex of squirrels. J Comp Neurol. 1986;250:403–30. doi: 10.1002/cne.902500402. [DOI] [PubMed] [Google Scholar]
- Kudo M, Aitkin LM, Nelson JE. Auditory forebrain organization of an Australian marsupial, the northern native cat (Dasyurus hallucatus) J Comp Neurol. 1989;279:28–42. doi: 10.1002/cne.902790104. [DOI] [PubMed] [Google Scholar]
- Kudo M, Glendenning KK, Frost SB, Masterton RB. Origin of mammalian thalamocortical projections. I. Telencephalic projections of the medial geniculate body in the opossum (Didelphis virginiana) J Comp Neurol. 1986;245:176–97. doi: 10.1002/cne.902450205. [DOI] [PubMed] [Google Scholar]
- Lende RA. Cerebral cortex: a sensorimotor amalgam in the marsupiala. Science. 1963a;141:730–2. doi: 10.1126/science.141.3582.730. [DOI] [PubMed] [Google Scholar]
- Lende RA. Motor Representation in the Cerebral Cortex of the Opossum (Didelphis Virginiana) J Comp Neurol. 1963b;121:405–15. doi: 10.1002/cne.901210308. [DOI] [PubMed] [Google Scholar]
- Lende RA. Sensory Representation in the Cerebral Cortex of the Opossum (Didelphis Virginiana) J Comp Neurol. 1963c;121:395–403. doi: 10.1002/cne.901210307. [DOI] [PubMed] [Google Scholar]
- Lende RA. Representation in the Cerebral Cortex of a Primitive Mammal. Sensorimotor, Visual, and Auditory Fields in the Echidna (Tachyglossus Aculeatus) J Neurophysiol. 1964;27:37–48. doi: 10.1152/jn.1964.27.1.37. [DOI] [PubMed] [Google Scholar]
- Lende RA. A comparative approach to the neocortex: localization in monotremes, marsupials, and insectivores. Ann N Y Acad Sci. 1969;167:262–275. [Google Scholar]
- Linden R, Rocha-Miranda CE. Organization of the visual thalamus: corticothalamic projections from the primary visual area in the opossum. Braz J Med Biol Res. 1983;16:247–60. [PubMed] [Google Scholar]
- Loo YT. The forebrain of the opossum, Didelphis virginiana. I. Gross anatomy. J Comp Neurol. 1930;51:13–64. [Google Scholar]
- Loo YT. The forebrain of the opossum, Didelphis virginiana. Part II. Histology. J Comp Neurol. 1931;52:1–148. [Google Scholar]
- Luethke LE, Krubitzer LA, Kaas JH. Cortical connections of electrophysiologically and architectonically defined subdivisions of auditory cortex in squirrels. J Comp Neurol. 1988;268:181–203. doi: 10.1002/cne.902680205. [DOI] [PubMed] [Google Scholar]
- Luo ZX, Cifelli RL, Kielan-Jaworowska Z. Dual origin of tribosphenic mammals. Nature. 2001;409:53–7. doi: 10.1038/35051023. [DOI] [PubMed] [Google Scholar]
- Luo ZX, Ji Q, Wible JR, Yuan CX. An Early Cretaceous tribosphenic mammal and metatherian evolution. Science. 2003;302:1934–40. doi: 10.1126/science.1090718. [DOI] [PubMed] [Google Scholar]
- Magalhaes-Castro B, Saraiva PE. Sensory and motor representation in the cerebral cortex of the marsupial Didelphis azarae azarae. Brain Res. 1971;34:291–9. doi: 10.1016/0006-8993(71)90282-4. [DOI] [PubMed] [Google Scholar]
- Marotte LR, Leamey CA, Waite PM. Timecourse of development of the wallaby trigeminal pathway: III. Thalamocortical and corticothalamic projections. J Comp Neurol. 1997;387:194–214. doi: 10.1002/(sici)1096-9861(19971020)387:2<194::aid-cne3>3.0.co;2-w. [DOI] [PubMed] [Google Scholar]
- Marshall Graves JA, Westerman M. Marsupial genetics and genomics. Trends Genet. 2002;18:517–21. doi: 10.1016/s0168-9525(02)02772-5. [DOI] [PubMed] [Google Scholar]
- Martin GF., Jr Interneocortical connections in the opossum, Didelphis virginiana. Anat Rec. 1967;157:607–15. doi: 10.1002/ar.1091570406. [DOI] [PubMed] [Google Scholar]
- Martinich S, Pontes MN, Rocha-Miranda CE. Patterns of corticocortical, corticotectal, and commissural connections in the opossum visual cortex. J Comp Neurol. 2000;416:224–44. [PubMed] [Google Scholar]
- Martinich S, Rosa MG, Rocha-Miranda CE. Patterns of cytochrome oxidase activity in the visual cortex of a South American opossum (Didelphis marsupialis aurita) Braz J Med Biol Res. 1990;23:883–7. [PubMed] [Google Scholar]
- Mayner L. A cyto-architectonic study of the cortex of the tammar wallaby, Macropus eugenii. Brain Behav Evol. 1989;33:303–16. doi: 10.1159/000115938. [DOI] [PubMed] [Google Scholar]
- Merzenich MM, Kaas JH, Roth GL. Auditory cortex in the grey squirrel: tonotopic organization and architectonic fields. J Comp Neurol. 1976;166:387–401. doi: 10.1002/cne.901660402. [DOI] [PubMed] [Google Scholar]
- Moeller HF. Introduction. In: Grzimek B, editor. Grzimek’s Encyclopedia of Mammals. McGraw-Hill; New York: 1990. pp. 212–219. [Google Scholar]
- Munemasa M, Nikaido M, Donnellan S, Austin CC, Okada N, Hasegawa M. Phylogenetic analysis of diprotodontian marsupials based on complete mitochondrial genomes. Genes Genet Syst. 2006;81:181–91. doi: 10.1266/ggs.81.181. [DOI] [PubMed] [Google Scholar]
- Murphy WJ, Pevzner PA, O’Brien SJ. Mammalian phylogenomics comes of age. Trends Genet. 2004;20:631–9. doi: 10.1016/j.tig.2004.09.005. [DOI] [PubMed] [Google Scholar]
- Myers P, Espinosa R, Parr CS, Jones T, Hammond GS, Dewey TA. 2006 The Animal Diversity Web (online). http://animaldiversity.org.
- Nelson JE, Stephan H. Encephalization in Australian marsupials. In: Archer M, editor. Carnivorous Marsupials. Royal Zoological Society of New South Wales; Mosman, Australia: 1982. pp. 699–706. [Google Scholar]
- Nelson LR, Lende RA. Interhemispheric Responses in the Opossum. J Neurophysiol. 1965;28:189–99. doi: 10.1152/jn.1965.28.2.189. [DOI] [PubMed] [Google Scholar]
- Neylon L, Haight JR. Neocortical projections of the suprageniculate and posterior thalamic nuclei in the marsupial brush-tailed possum, Trichosurus vulpecula (Phalangeridae), with a comparative commentary on the organization of the posterior thalamus in marsupial and placental mammals. J Comp Neurol. 1983;217:357–75. doi: 10.1002/cne.902170402. [DOI] [PubMed] [Google Scholar]
- Nieder PC, Randall W. Sound-Evoked Potentials in Neocortex of Unanesthetized Opossum. Science. 1964;144:429–30. doi: 10.1126/science.144.3617.429. [DOI] [PubMed] [Google Scholar]
- Nilsson MA, Arnason U, Spencer PBS, Janke A. Marsupial relationships and a timeline for marsupial radiation in South Gondwana. Gene. 2004;340:189–196. doi: 10.1016/j.gene.2004.07.040. [DOI] [PubMed] [Google Scholar]
- Nowak RM. Walker’s marsupials of the world. Johns Hopkins University Press; Baltimore: 2005. [Google Scholar]
- Nudo RJ, Masterton RB. Descending pathways to the spinal cord, III: Sites of origin of the corticospinal tract. J Comp Neurol. 1990;296:559–83. doi: 10.1002/cne.902960405. [DOI] [PubMed] [Google Scholar]
- Oliveira L, Volchan E, Pessoa L, Pantoja JH, Joffily M, Souza-Neto D, Marques RF, Rocha-Miranda CE. Contour integration in the primary visual cortex of the opossum. Neuroreport. 2002;13:2001–4. doi: 10.1097/00001756-200211150-00002. [DOI] [PubMed] [Google Scholar]
- Owen R. On the structure of the brain in marsupial animals. Philos Trans R Soc Lond B Biol Sci. 1837;127:87–96. [Google Scholar]
- Packer AD. An experimental investigation of the visual system in the phalanger, Trichosurus vulpecula. J Anat. 1941;75:309–329. [PMC free article] [PubMed] [Google Scholar]
- Parker SP. Marsupials. In: Grzimek B, editor. Grzimek’s Encyclopedia of Mammals. McGraw-Hill; New York: 1990. pp. 210–211. [Google Scholar]
- Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain. 1937;60:389–443. [Google Scholar]
- Pubols BH., Jr Retrograde degeneration study of somatic sensory thalamocortical connections in brain of Virginia opossum. Brain Res. 1968;7:232–51. doi: 10.1016/0006-8993(68)90101-7. [DOI] [PubMed] [Google Scholar]
- Pubols BH., Jr The second somatic sensory area (SmII) of opossum neocortex. J Comp Neurol. 1977;174:71–8. doi: 10.1002/cne.901740105. [DOI] [PubMed] [Google Scholar]
- Pubols BH, Jr, Pubols LM, DiPette DJ, Sheely JC. Opossum somatic sensory cortex: a microelectrode mapping study. J Comp Neurol. 1976;165:229–45. doi: 10.1002/cne.901650208. [DOI] [PubMed] [Google Scholar]
- Putnam SJ, Megirian D, Manning JW. Marsupial interhemispheric relation. J Comp Neurol. 1968;132:227–34. doi: 10.1002/cne.901320202. [DOI] [PubMed] [Google Scholar]
- Rassmussen DT, Sussman RW. Parallelisms among primates and possums. In: Ravosa MJ, Dagosto M, editors. Primate Origins, Adaptations and Evolution. Springer; New York: 2007. pp. 775–803. [Google Scholar]
- Ravizza RJ, Heffner HE, Masterton B. Hearing in primitive mammals, I: Opossum (Didelphis virginianus) Journal of Auditory Research. 1969;9:1–7. [Google Scholar]
- Ravizza RJ, Masterton B. Contribution of neocortex to sound localization in opossum (Didelphis virginiana) J Neurophysiol. 1972;35:344–56. doi: 10.1152/jn.1972.35.3.344. [DOI] [PubMed] [Google Scholar]
- Rees S, Hore J. The motor cortex of the brush-tailed possum (Trichosurus vulpecula): motor representation, motor function and the pyramidal tract. Brain Res. 1970;20:439–51. doi: 10.1016/0006-8993(70)90173-3. [DOI] [PubMed] [Google Scholar]
- Rocha-Miranda CE, Bombardieri RA, Jr, de Monasterio FM, Linden R. Receptive fields in the visual cortex of the opossum. Brain Res. 1973;63:362–7. doi: 10.1016/0006-8993(73)90105-4. [DOI] [PubMed] [Google Scholar]
- Rogers FT. An experimental study of the cerebral physiology of the virginian opossum. J Comp Neurol. 1924;37:265–315. [Google Scholar]
- Rosa MG, Krubitzer LA, Molnar Z, Nelson JE. Organization of visual cortex in the northern quoll, Dasyurus hallucatus: evidence for a homologue of the second visual area in marsupials. Eur J Neurosci. 1999;11:907–15. doi: 10.1046/j.1460-9568.1999.00497.x. [DOI] [PubMed] [Google Scholar]
- Rowe M. Organization of the cerebral cortex in monotremes and marsupials. In: Jones EG, Peters A, editors. Cerebral Cortex. Plenum; New York: 1990. pp. 263–334. [Google Scholar]
- Sanderson KJ, Haight JR, Pearson LJ. Transneuronal transport of tritiated fucose and proline in the visual pathways of the brushtailed possum, Trichosurus vulpecula. Neurosci Lett. 1980;20:243–8. doi: 10.1016/0304-3940(80)90154-8. [DOI] [PubMed] [Google Scholar]
- Shang F, Ashwell KW, Marotte LR, Waite PM. Development of commissural neurons in the wallaby (Macropus eugenii) J Comp Neurol. 1997;387:507–23. doi: 10.1002/(sici)1096-9861(19971103)387:4<507::aid-cne3>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- Sheng XM, Marotte LR, Mark RF. Development of connections to and from the visual cortex in the wallaby (Macropus eugenii). J Comp Neurol. 1990;300:196–210. doi: 10.1002/cne.903000205. [DOI] [PubMed] [Google Scholar]
- Sheng XM, Marotte LR, Mark RF. Development of the laminar distribution of thalamocortical axons and corticothalamic cell bodies in the visual cortex of the wallaby. J Comp Neurol. 1991;307:17–38. doi: 10.1002/cne.903070103. [DOI] [PubMed] [Google Scholar]
- Silver J, Lorenz SE, Wahlsten D, Coughlin J. Axonal guidance during development of the great cerebral commissures: descriptive and experimental studies, in vivo, on the role of preformed glial pathways. J Comp Neurol. 1982;210:10–29. doi: 10.1002/cne.902100103. [DOI] [PubMed] [Google Scholar]
- Slutsky DA, Manger PR, Krubitzer L. Multiple somatosensory areas in the anterior parietal cortex of the California ground squirrel (Spermophilus beecheyii) J Comp Neurol. 2000;416:521–39. doi: 10.1002/(sici)1096-9861(20000124)416:4<521::aid-cne8>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- Smith GE. On a peculiarity of the cerebral commissures in certain marsupialia, not hitherto recognised as a distinctive feature of the diprotodontia. Proc Roy Soc, Lond. 1902;70:226–231. [Google Scholar]
- Sousa AB, Gattass R, Oswaldo-Cruz E. The projection of the opossum’s visual field on the cerebral cortex. J Comp Neurol. 1978;177:569–87. doi: 10.1002/cne.901770404. [DOI] [PubMed] [Google Scholar]
- Szalay X. A new appraisal of marsupial phylogeny and classification. In: Archer M, editor. Carnivorous marsupials. Royal Zoological Society of New South Wales; Mosman, Australia: 1982. pp. 621–640. [Google Scholar]
- Thenius E. Phylogeny. In: Grzimek B, editor. Grzimek’s Encyclopedia of Mammals. McGraw-Hill; New York: 1990. pp. 219–231. [Google Scholar]
- Tyler CJ, Dunlop SA, Lund RD, Harman AM, Dann JF, Beazley LD, Lund JS. Anatomical comparison of the macaque and marsupial visual cortex: common features that may reflect retention of essential cortical elements. J Comp Neurol. 1998;400:449–68. doi: 10.1002/(sici)1096-9861(19981102)400:4<449::aid-cne2>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- Vidyasagar TR, Wye-Dvorak J, Henry GH, Mark RF. Cytoarchitecture and visual field representation in area 17 of the tammar wallaby (Macropus eugenii) J Comp Neurol. 1992;325:291–300. doi: 10.1002/cne.903250211. [DOI] [PubMed] [Google Scholar]
- Vogt C, Vogt O. Zur kenntnis der elektrisch erregbaren hirnrindengebiete bei den saugetieren. J f Psych u Neur. 1906;8:277–456. [Google Scholar]
- Vogt C, Vogt O. Zur Kenntnis der electrisch erregbaren Hirnrindengebiete bei den Saugetieren. J f Psych u Neur. 1907;8:277–456. [Google Scholar]
- Volchan E, Bernardes RF, Rocha-Miranda CE, Gleiser L, Gawryszewski LG. The ipsilateral field representation in the striate cortex of the opossum. Exp Brain Res. 1988;73:297–304. doi: 10.1007/BF00248221. [DOI] [PubMed] [Google Scholar]
- Waite PM, Marotte LR, Leamey CA, Mark RF. Development of whisker-related patterns in marsupials: factors controlling timing. Trends Neurosci. 1998;21:265–9. doi: 10.1016/s0166-2236(97)01217-4. [DOI] [PubMed] [Google Scholar]
- Waite PM, Marotte LR, Mark RF. Development of whisker representation in the cortex of the tammar wallaby Macropus eugenii. Brain Res Dev Brain Res. 1991;58:35–41. doi: 10.1016/0165-3806(91)90234-a. [DOI] [PubMed] [Google Scholar]
- Walsh TM, Ebner FF. The cytoarchitecture of somatic sensory-motor cortex in the opossum (Didelphis marsupialis virginiana): a Golgi study. J Anat. 1970;107:1–18. [PMC free article] [PubMed] [Google Scholar]
- Watson CR. The corticospinal tract of the quokka wallaby (Setonix brachyurus) J Anat. 1971a;109:127–33. [PMC free article] [PubMed] [Google Scholar]
- Watson CR. An experimental study of the corticospinal tract of the kangaroo. J Anat. 1971b;110:501. [PubMed] [Google Scholar]
- Weed LH, Langworthy OR. Developmental study of excitatory areas in the cerebral cortex of the opossum. Am J Physiol. 1925;72:8–24. [Google Scholar]
- Weller WL. Barrels in somatic sensory neocortex of the marsupial Trichosurus vulpecula (brush-tailed possum) Brain Res. 1972;43:11–24. doi: 10.1016/0006-8993(72)90271-5. [DOI] [PubMed] [Google Scholar]
- Weller WL. SmI cortical barrels in an Australian marsupial, Trichosurus vulpecula (brush-tailed possum): structural organization, patterned distribution, and somatotopic relationships. J Comp Neurol. 1993;337:471–92. doi: 10.1002/cne.903370310. [DOI] [PubMed] [Google Scholar]
- Weller WL, Haight JR, Neylon L, Johnson JI. A re-assessment of the mechanoreceptor projections to cerebral neocortex in marsupial wallabies (Thylogale) J Anat. 1977;124:531–532. [Google Scholar]
- Woodburne MO, Case JA. Dispersal, vicariance, and the late Cretaceous to early Tertiary land mammal biogeography from South America to Australia. J Mammal Evol. 1996;3:121–161. [Google Scholar]
- Woodburne MO, Rich TH, Springer MS. The evolution of tribospheny and the antiquity of mammalian clades. Mol Phylogenet Evol. 2003;28:360–85. doi: 10.1016/s1055-7903(03)00113-1. [DOI] [PubMed] [Google Scholar]
- Woolsey CN. Organization of somatic sensory and motor areas of the cerebral cortex. In: Harlow H, Woolsey C, editors. Biological and biochemical bases of behavior. University of Wisconsin Press; Madison: 1958. pp. 63–81. [Google Scholar]
- Woolsey TA, Welker C, Schwartz RH. Comparative anatomical studies of the SmL face cortex with special reference to the occurrence of “barrels” in layer IV. J Comp Neurol. 1975;164:79–94. doi: 10.1002/cne.901640107. [DOI] [PubMed] [Google Scholar]
- Ziehen T. Ueber die motorische Rindenregion von Didelphys. Centrlbt f Physiol. 1897;11:457–461. [Google Scholar]