

Abstract
Mouse cells are non-permissive to human immunodeficiency virus type 1 (HIV) in that there is a pronounced post-integration block to viral replication. We have recently demonstrated that mouse-human somatic cell hybrids that contain human chromosome 2 increase both HIV Capsid (CA) production and infectious virus release. Here we report on the isolation of three mouse-human microcell hybrids (MCHs) that behave similarly, starting from a pool of 500 MCH clones. Release of virus was specific to HIV and cell revertants that no longer contained any human chromosome fragments did not release CA or infectious virus. Two of the three cell clones were identical as judged by PCR STS content and fluorescence in situ hybridization (FISH) and contained a single 2-12 human chromosome chimera. The third cell clone only contained human chromosome 12, as determined by PCR, FISH, and microarray analyses. There were no consistent differences in Gag protein and spliced/unspliced viral RNA levels between mouse cell lines. CMV promoter-driven, codon-optimized gag-pol had no effect on infectious HIV release from these mouse cells, despite allowing Gag targeting and increasing CA production. These permissive mouse-human MCHs and their corresponding non-permissive revertants may prove useful for mechanistic studies and also for identifying the responsible gene(s) or factor(s) involved in the production of HIV.
Keywords: Human immunodeficiency virus, Murine model, Capsid production, Microcell hybrids
Introduction
There is no small animal model permissive for human immunodeficiency virus type 1 (HIV) replication. For example, after provision of the viral entry determinants (i.e., human CD4 and the appropriate chemokine co-receptor) to murine cells, the level of transcription from the HIV long terminal repeat (LTR) was relatively low. Identification of human cyclin T1 allowed for greater transcriptional processivity from the LTR (Garber et al., 1998; Wei et al., 1998) and resulted in higher amounts of viral gene products in murine cells. Still, HIV-infected mouse cells that express human CD4, co-receptor, and cyclin T1 released only trace amounts of capsid p24 (CA) and little infectious virus (Bieniasz and Cullen, 2000; Garber et al., 1998; Mariani et al., 2000), suggesting the presence of a post-integration block to HIV replication.
Because mouse-human cell fusions are more permissive to viral replication, others have proposed that one or more essential factors that permit completion of the viral life cycle are absent from mouse cells (Bieniasz and Cullen, 2000). In the presence of cyclin T1, mouse cells express both unspliced and spliced viral RNAs, although the ratio of unspliced to spliced is considerably lower compared to human cells (Mariani et al., 2000). By electron microscopy amorphous collections of Gag accrue in the cytoplasm near or in unknown vesicular structures, suggesting that the observed replicative block is at a post-translational stage (Mariani et al., 2000). Neither tethered nor extracellular viral particles are seen in mouse cells infected with HIV (Mariani et al., 2000), suggesting that the replication defect is earlier than the final assembly/budding steps from the plasma membrane.
The block to virus production in mouse cells has been circumvented to some extent by the use of a chimeric murine leukemia virus/HIV matrix p17 (MA) gene, although the level of viral replication was much lower in comparison to wild-type HIV in human cells (Chen et al., 2001; Reed et al., 2002). The globular head of MA may actually be inhibitory to plasma membrane binding (Perez-Caballero et al., 2004; Spearman et al., 1997), which is likely related to the myristate switch (Ono and Freed, 1999; Paillart and Gottlinger, 1999) . Gag expression and production of infectious virus in both human and murine cells was increased by deletions in portions of MA (Hatziioannou et al., 2005), although those results have not been consistently observed by all investigators (Hubner and Chen, 2006). These data imply that as in human cells HIV MA may function inefficiently or actually be antagonistic in mouse cells, but there are likely to be membrane-independent steps required for virus assembly. Multimerization of Gag and association at the plasma membrane was found to be highly concentration-dependent (Perez-Caballero et al., 2004), suggesting CA production and infectious virus release may be particularly dependent upon level of Gag protein, perhaps in a non-linear manner. Reduced CA production/release cannot be explained by the action of mouse APOBEC3, since APOBEC proteins are not known to interfere with Gag processing to any significant extent.
Nuclear export and cytoplasmic translation of fully spliced, partially spliced, and unspliced viral mRNAs is absolutely required for proper and efficient assembly of HIV. HIV RNAs that contain introns are exported through the action of the Rev/Rev-response-element (RRE) pathway which involves Crm1 and Ran-GTP (Cullen, 2003). Mason-Pfizer monkey virus (MPMV), a retrovirus, relies on a separate pathway for unspliced mRNA export (Bray et al., 1994). Here, NFX1 (also known as Tap) binds to the constitutive transport element (CTE) of MPMV and RNA export occurs via normal cellular mechanisms, but separate from the Rev/RRE pathway. When the RRE was replaced with a multimerized CTE from MPMV, in mouse cells both Gag trafficking to cellular membranes and CA production were restored (Swanson et al., 2004). Thus, in mouse cells Rev/RRE-containing transcripts may be incorrectly trafficked after nuclear export, perhaps because the ‘marked’ RNAs are functionally deficient in some manner.
Previously we had used a panel mouse-human somatic cell hybrids (SCHs) to demonstrate that the presence of chromosome 2 allowed efficient CA and infectious virus production from murine cells, which was specific to HIV (Coskun et al., 2006). This effect did not appear to be due to alterations in cyclinT1 function and there was no clear increase in the amounts of unspliced mRNA and Gag protein precursor. Presence of Vpu along with chromosome 2 had only a modest impact on infectious virus release (Coskun et al., 2006).
In parallel with the SCH genetic approach, we began with a panel of microcell hybrids (MCHs) (Speevak et al., 1995; Speevak and Chevrette, 1996). At the time we believed that identification and characterization of permissive MCHs would lead to improved genomic resolution of the phenotype of infectious HIV release. Here we report the isolation of three MCH cell clones highly permissive for CA and infectious virus release, which again is specific to HIV. Cells that had lost the human chromosome also reverted their phenotype. Two of the cell clones appear to be identical and contain a single 2-12 chromosome chimera. The third appears to contain only intact chromosome 12. Use of codon-optimized gag-pol did not increase infectious virus production from any of the mouse cell lines. These cell clones and their associated revertants may prove useful for the further study of the block to HIV replication in murine cells.
Results
Isolation of MCH cell clones permissive to HIV release
Mouse cells release little infectious HIV. As a potential genetic approach to this problem, a pool of 500 mouse-human MCHs was obtained and transduced in bulk with HIV vector pHIV-CIB pseudotyped with VSV G at low MOI. The outline of this screen is shown in Figure 1A. The MCHs are also maintained under hygromycin selection since the human chromosome portion has an integrated, CMV-driven hygroTK gene. Parental mouse melanoma B78 cells were also transduced. HIV-CIB encodes all requisite cis-acting sequences along with gag, pol, tat, rev, and a 1.9 kb bicistronic cassette encoding truncated human cyclinT1 linked by an internal ribosome entry site to blasticidin deaminase (Figure 1B). In many human cell types the vector may be recovered or rescued simply by provision of VSV G or other suitable viral glycoprotein and titered by infecting naïve target cells and subsequently enumerating blasticidin-resistant colonies. After transduction, MCHs were stably passaged in the presence of blasticidin and then subdivided into 80 pools of ∼25 cells each. Each of these was transfected with VSV G and the supernatant used to infect 293T cells. Compared to the parental B78 cells, 3 of the pools reproducibly released infectious virus. Each of these three pools was broken down to individual cell clones, and over 150 cell clones were tested to identify 3 (one from each pool) that repeatedly gave efficient HIV release. These clones, designated p11, p19, and p28, were further investigated.
Figure 1. MCH Genetic strategy.
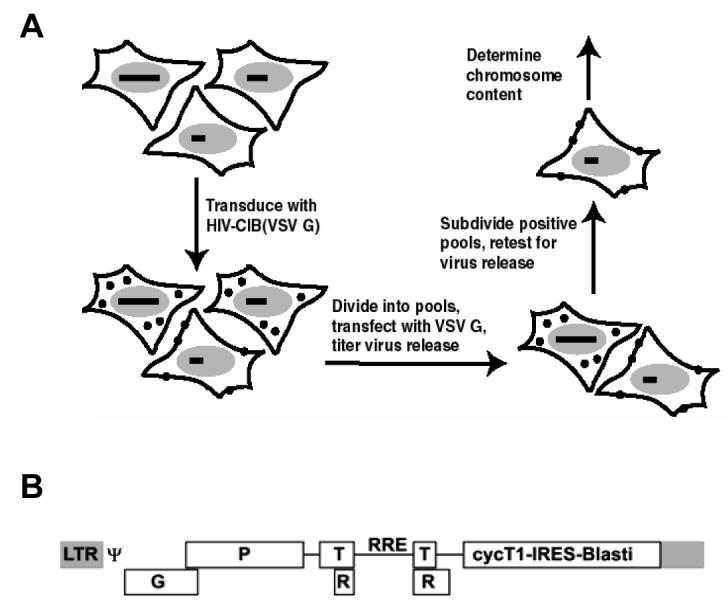
(A) MCH screen. Entire pool was first transduced at low MOI with HIV-CIB(VSV G), selected using 10 μg/ml blasticidin, and then broken down into smaller pools of ∼25 clones. Each smaller pool was transfected with VSV G expression plasmid and rescued HIV titered on naive human cells. Positive pools were broken down to single cells and the process iterated to identify permissive clones (B) Schematic of pHIV-CIB. Truncated cyclin T1 is expressed as part of a bicistronic cassette along with Blasti, blasticidin deaminase. LTR denotes long terminal repeat, G gag, P pol, T tat, R rev, RRE Rev response element, ψ packaging signal. Gene products not to scale.
MCH clones release infectious HIV and CA
The MCH clones of interest were transfected with VSV G and infectious virus present in supernatant was titered on 293T cells. As can be seen in Figure 2A, the MCHs were quite permissive for infectious HIV release, especially when compared to the parental B78 mouse melanoma cells (negative control) and 293Ts (positive control). In order to show that this phenotype was due to the human chromosome(s) present, cells were passaged at low confluency into ganciclovir along with blasticidin S. Surviving cell colonies (arising at a frequency of ∼1 in 10,000) were pooled, tested to verify that they were now sensitive to hygromycin, and then transfected with VSV G. The revertant cells now released little HIV (Figure 2A). PCR across the genome was also performed to confirm loss of associated human DNA (see Table 2 below). Of note, the MCH clones were not as permissive as the 293T cells, perhaps related to transfection efficiency, an increase in unspliced viral RNA (see Figure 4 below), or other unknown factors.
Figure 2. Recovery of HIV and MLV from MCHs.

(A) MCHs that had been transduced with HIV-CIB(VSV G) were transfected with VSV G at least in triplicate and resulting supernatant titered on 293T targets. Titer was normalized to transfection efficiency, as judged by an eYFP reporter. * P<0.05 compared to parental B78 cells; **P>0.05 compared to parental B78 cells; R denotes revertant. (B) MCHs that had been transduced with HIV-CIB(VSV G) were transfected with ADA envelope and resulting supernatant titered on GHOST HI5 targets. Titer was normalized to transfection efficiency, as judged by an eYFP reporter. Experiment was repeated twice with similar results. Typically recovery of infectious HIV(ADA) from similarly transfected 293T cells was >1000. Down arrow indicates no infectious particles were recovered from 2 ml of supernatant in this particular experiment. (C) MCHs and 293Ts were transfected with MLV vector encoding eGFP, VSV G, and MLV gag-pol. Supernatant was titered on adherent human cells by epifluorescence microscopy 72 hours later, normalized toB78 parental cells. Experiment was repeated with similar results (open and closed bars). We occasionally noted variability in recovery of infectious MLV from 293T cells, perhaps due to differences in transfection efficiency of the three plasmids.
Table 2.
PCR Results from Linkage Mapping Set
| Locus1 | Chromosome | Position (bp)2 | p193 | p284 | p11 | p11-R5 | 293T |
|---|---|---|---|---|---|---|---|
| D2S319 | 2 | 3480348-3480477 | + | − | + | − | + |
| D2S2211 | 2 | 7383683-7383944 | + | +/− | + | − | + |
| D2S162 | 2 | 8777748-8777871 | + | − | + | − | + |
| D2S168 | 2 | 8777748-8777871 | + | − | + | − | + |
| D2S165 | 2 | 28456872-28456976 | + | − | + | − | + |
| D2S367 | 2 | 34294707-34294847 | + | − | + | − | + |
| D2S2259 | 2 | 42835897-42836125 | + | − | + | − | + |
| D2S391 | 2 | 46265007-46265152 | + | − | + | − | + |
| D2S337 | 2 | 33790596-33790704 | + | +/− | + | − | + |
| D2S2368 | 2 | 67062334-67062425 | + | − | + | − | + |
| D2S286 | 2 | 75195641-75195784 | + | − | + | − | + |
| D2S2333 | 2 | 85316486-85316739 | + | − | + | − | + |
| D2S2216 | 2 | 88190785-88190917 | + | +/− | + | − | + |
| D2S347 | 2 | 117574432-117574717 | − | − | − | − | + |
| D2S142 | 2 | 149896624-149896883 | − | − | − | − | + |
| D2S2330 | 2 | 160307545-160307709 | − | − | − | − | + |
| D2S335 | 2 | 172274762-172274856 | − | − | − | − | + |
| D2S364 | 2 | 182742779-182743014 | + | − | + | − | + |
| D2S117 | 2 | 195327044-195327241 | + | − | + | − | + |
| D2S325 | 2 | 207979115-207979227 | + | − | + | − | + |
| D2S2382 | 2 | 210818276-210818514 | + | − | + | − | + |
| D2S126 | 2 | 221725283-221725500 | + | − | + | − | + |
| D2S396 | 2 | 230391872-230392111 | + | − | + | − | + |
| D2S206 | 2 | 233416066-233416307 | + | − | + | − | + |
| D2S338 | 2 | 230948422-230948710 | + | − | + | − | + |
| D2S125 | 2 | 240816799-240816894 | + | − | + | − | + |
| D12S352 | 12 | 531659-531820 | + | + | + | − | + |
| D12S99 | 12 | 5435032-5435160 | + | + | + | − | + |
| D12S310 | 12 | 18864808-18865059 | + | + | + | − | + |
| D12S1617 | 12 | 24991299-24991555 | + | + | + | − | + |
| D12S85 | 12 | 45622990-45623114 | + | + | + | − | + |
| D12S83 | 12 | 59175723-59175935 | + | + | + | − | + |
| D12S79 | 12 | 114544839-114545005 | + | + | + | − | + |
| D12S86 | 12 | 117654705-117654838 | + | + | + | − | + |
| D12S1659 | 12 | 127982361-127982460 | + | + | + | − | + |
| D12S1723 | 12 | 130528689-130528851 | + | + | + | − | + |
Footnotes:
Non-informative loci (in which no PCR product was present or was also observed in the water control) have been omitted.
Chromosome bp position as per UCSC genome browser, release date of March 2006.
+ denotes presence, − denotes absence of expected PCR product.
+/− result denotes faint band present and other, immediately surrounding loci tested negative.
R denotes pool of revertants, which was ganciclovir resistant and hygromycin sensitive.
Figure 4. Nucleic acid analysis of MCHs.
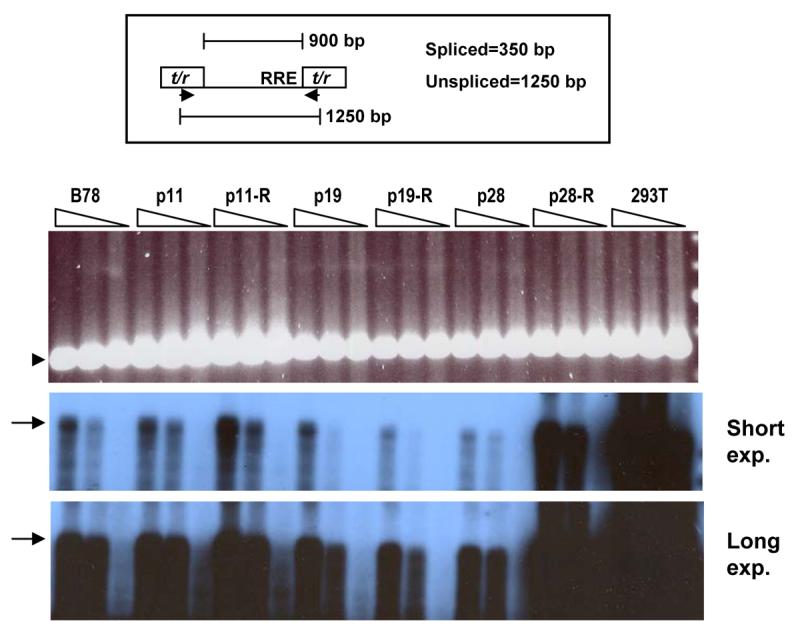
Total RNA was prepared from MCHs and 293Ts stably transduced with HIV-CIB or HIV-LIB, respectively, and subjected to RT-PCR, using oligo(dT) as first strand primer. At top is shown a schematic of the region of the HIV vector amplified, with the DNA primers (arrows) located within the tat/rev (t/r) exons (rectangles). Truncated intron is ∼900 bp. RRE denotes Rev-response element. Bottom two panels: results using HIV primers using 3 different amounts of first strand reaction equivalent to RNA amounts of 15, 45, and 135 ng (wedges), Southern blotted using a 0.7 kb intronic probe (two different film exposure times; expected product of ∼1250 bp indicated by arrow). Top panel: Results using the same primers and amounts of first strand cDNA, with expected spliced product of 350 bp indicated by arrowhead (EtBr stain). Unspliced products have been cropped. No products were observed with water (H) control or in the absence of RT. These RT-PCR experiments were repeated twice with similar results. R denotes revertant cell line.
The MCH clones of interest were also transfected with the M-tropic envelope ADA and released infectious virus titered on GHOST HI5 cells. Again, the MCH cell clones were quite permissive for HIV(ADA) release, whereas the revertants produced little infectious virus (Figure 2B). To demonstrate specificity to HIV, cell clones were tri-transfected with VSV G, pHIT60, and pBABE-IY, and released infectious MLV titered on HOS cell targets. Although there was some variability in MLV production, none of the MCHs had a marked increase in amount of MLV present in the culture supernatant (Figure 2C). The variability of MLV release from 293T cells may best be explained by differences in transfection efficiency from experiment to experiment.
The negligible release of infectious HIV from mouse cells is correlated with low amounts of CA release. Previously we had demonstrated that SCHs that contain human chromosome 2 release significantly greater amounts of infectious HIV and CA. Thus, we also measured the amount of CA produced from the MCH clones, as quantified by ELISA. The absolute amounts of CA in the tissue culture supernatants paralleled the results from the infectious virus assays (Figure 3A). To demonstrate that this was not simply due to more intracellular CA in the individual cell clones, the percentage yield of supernatant CA was also determined, and that paralleled the other results (Figure 3B). This suggests that the increase in CA production observed is not simply due to more Gag protein in those MCH clones (and see next section below).
Figure 3. CA analysis of MCHs.
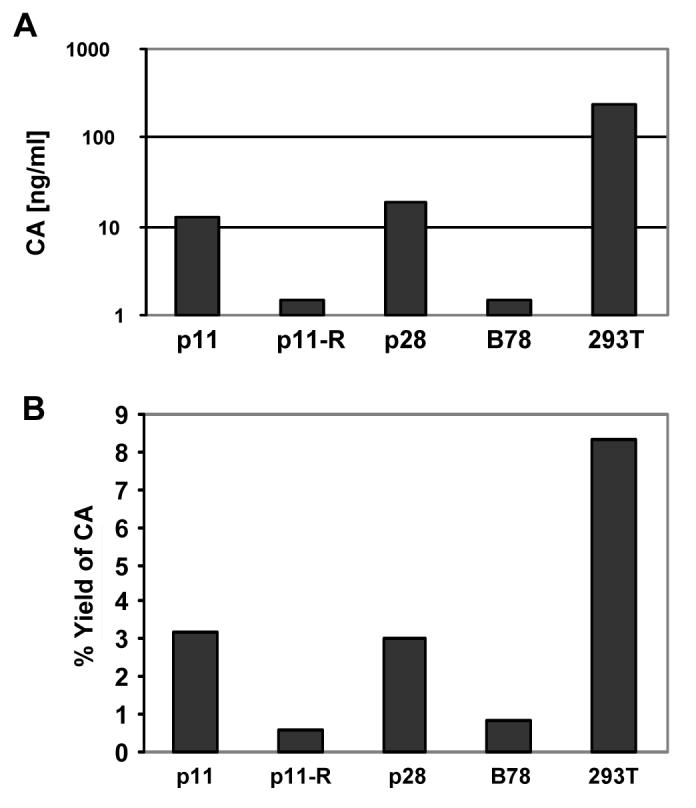
MCHs or 293Ts that had been stably transduced with HIV-CIB or HIV-LIB, respectively, were plated at low density and allowed to reach confluence. At that point equivalent amounts of supernatant and cells were harvested for CA quantitation using commercial ELISA kit. Absolute amounts of supernatant CA are shown in (A). (B) Gag yield (amount of CA in the supernatant divided by amount in cell lysate), using equivalent number of lysed cells. R denotes revertant. This experiment was repeated with similar results.
Many of the MCH clones in the pool have all or a portion of chromosome 12 present ((Speevak et al., 1995; Speevak and Chevrette, 1996) and see below). SCHs that have chromosome 12 have improved cyclin T1 function but release little infectious virus (Coskun et al., 2006). Although the results of the genetic screen implied that only rare MCH clones had the phenotype of virus release, we wished to test randomly picked clones for both presence of chromosome 12 and the ability to release HIV. All of the randomly picked clones had most of chromosome 12 as determined by PCR using 8 different primer pairs spanning 12, whereas none efficiently released HIV (Table 1). This suggests that the mere presence of chromosome 12 is insufficient to allow infectious virus release. Note, however, that the randomly picked MCH clones did not amplify SHGC-81002 at 12q24.23 whereas p28 did, suggesting there may be chromosome 12-specific differences between the non-permissive and permissive clones.
Table 1.
Random MCH clones have most of chromosome 2 but do not support HIV release
| Cell line or clone1 |
Titer2 | SHGC- 123694 (12p13.32)3 |
SHGC- 140086 (12p12.3) |
SHGC- 147202 (12q21.33) |
RH45303 (12q13.12) |
SHGC- 81002 (12q24.23) |
SHGC- 83339 (12q24.31) |
SHGC- 84322 (12q24.32) |
RH39825 (12q24.33)4 |
|---|---|---|---|---|---|---|---|---|---|
| 293T | ∼10000 | + | + | + | + | + | + | + | + |
| M2 | 50 | +/− | + | + | + | − | + | +/− | + |
| M3 | 40 | + | + | + | + | − | + | +/− | + |
| M4 | 24 | + | + | + | + | − | + | +/− | + |
| M8 | 93 | +/− | + | + | − | − | + | +/− | − |
| M9 | 93 | +/− | + | + | − | − | + | +/− | − |
| p28 | 1700 | + | + | + | + | + | + | + | − |
| p28-Revertant | ND | − | − | − | − | − | − | − | − |
Footnotes:
Cells were transduced with HIV-CIB (VSV G) and selected using blasticidin S. p28-Revertant was selected using ganciclovir.
Recovery of HIV (IU/ml) after transfection of cells with VSV G, with transfection efficiency normalized using an eYFP reporter. ND denotes not determined.
+ denotes presence of PCR product, − denotes absence, +/− denotes weak product.
This marker is at position 131081841, near the tip of chromosome 12q.
Quantitation of HIV nucleic acid and p55 Gag protein
It is possible that the observed results are due to increased amounts of proviral DNA and viral RNA in the MCH clones of interest, leading to increased Gag and CA production. In order to determine proviral DNA copy number, genomic DNA was prepared, and integrated viral DNA was amplified along with β-actin (as a normalization control) and compared against a 293T cell clone with a single vector integrant. All of the MCH clones examined had a single vector integrant, within the error of the measurement (not shown).
We next examined levels of spliced and unspliced viral RNAs. Total RNA was prepared, reverse transcribed using oligo(dT) as first strand primer, and PCR performed across the foreshortened tat/rev intron as previously described (Coskun et al., 2006), with β-actin again serving as loading control. With these primers, the PCR product from spliced RNA is 350 bp and from unspliced 1200 bp (see schematic at top of Figure 4). As shown in Figure 4, there were no consistent differences in levels of either spliced or unspliced viral RNAs observed between the B78 parental cells, the MCH clones, and the revertants. We reproducibly observed a lower ratio of unspliced to spliced viral RNAs in the mouse cells compared to the human cells, as we and others have reported before (Coskun et al., 2006). The ratio of unspliced to spliced viral RNA was essentially unchanged across the various MCH cell clones, the corresponding revertants, and the B78 parents, with the notable exception of p28-R, which consistently gave higher levels of unspliced viral RNA (despite little infectious virus release).
Because there could be post-transcriptional effects, we also examined levels of intracellular Gag by immunoblotting. RIPA cell lysates of the transduced murine cells were size-fractionated by SDS-PAGE and HIV proteins detected using anti-HIV antisera. There were no consistent differences in p55 Gag levels observed between the various transduced mouse cell lines, whereas there was more Gag and CA present in the 293T cells (Figure 5). There was also no clear increase in intracellular CA in the MCH cell clones of interest compared to the revertants and B78 parents. These results, taken together, suggest that the phenotype observed in the MCH clones is not due to increased levels of unspliced/spliced viral RNA or precursor Gag protein.
Figure 5. Cell-associated Gag in MCHs.
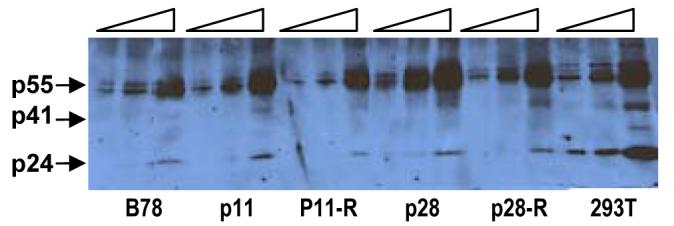
Two-fold increasing amounts (wedges) of cell lysates from indicated MCHs stably transduced with HIV-CIB and 293Ts transduced with HIV-LIB (smaller wedge) were subjected to SDS-PAGE, transferred to nitrocellulose, and immunoblotted using anti-HIV sera with detection using ECL. P55, p41, and p24 are indicated (arrows). Lysates from mock-transduced cells did not give rise to any HIV-specific bands.
DNA and chromosomal content of MCH clones
We also determined the human chromosomal content of the isolated MCH clones. Using a linkage mapping set consisting of 400 pairs of DNA primers spanning the human genome at 10 cM resolution, PCR was performed using genomic DNA prepared from the MCH clones, p11 revertant, and 293Ts, the latter serving as a positive control. Only regions of chromosome 2 and 12 consistently scored positive for the MCHs, and those results are summarized in Table 2. MCH clones p11 and p19 appeared to be identical by this PCR-based analysis. Both had present ∼90 Mb from chromosome 2p and 60 Mb from chromosome 2q, with most of chromosome 12. MCH clone p28 only had what appeared to be intact chromosome 12, without any other clearly identifiable region of the human genome present. Although some PCR products were weakly positive for some of the markers on chromosome 2, a more extensive analysis using flanking markers suggested that those regions were not present in p28 (not shown). As expected, the p11 revertant line had no portion of any human chromosome present.
We next performed FISH on metaphase spreads of the MCH clones. Using chromosome 2 paint, both p11 and p19 MCH clones had an identical complex, but single human chromosome present (Figure 6A, B). Chromosome 12 paint also identified a single chromosome in both p11 and p19, with the fluorescent signal encompassing the centromeric region (not shown). Hybridizing separate 2p and 2q labeled BACs demonstrated that both 2p and 2q were present on opposing arms of the single chromosome in p11 and p19 (Figure 6C). Hybridizing a 2p BAC and a BAC from 12 showed that they were on the same chromosome (Figure 6D). These results suggest that a chimeric 2-12 chromosome is present in MCH clones p11 and p19. Similar analyses were performed on MCH clone p28, but only chromosome 12 was detected, and it appeared to be intact, consistent with the PCR results (Tables 1 and 2).
Figure 6. FISH analysis of MCH clones.
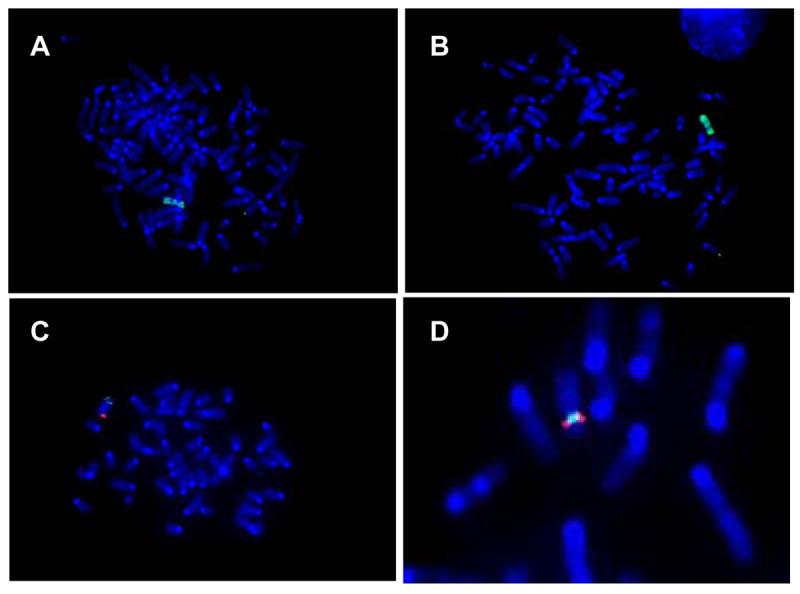
Metaphase spreads were prepared from MCH clones, hybridized with either chromosome-specific paint or BACs, and examined by high power fluorescence microscopy. (A) p11 probed with chromosome 2 paint; (B) p19 probed similarly; (C) p11 probed with a 2q BAC (green) and a 2p BAC (red); (D) p11 probed with a chromosome 2 BAC (red) and a chromosome 12 BAC (green).
Because both the genomic PCR and FISH are of relatively low resolution, we wished to use a higher resolution method to confirm that clone p28 only had chromosome 12 present. RNA was prepared from p28, reverse-transcribed, labeled, and used to probe a U133 Affymetrix microarray. cDNAs were sorted on the basis of numerical order on each chromosome, and the results are shown for all of the chromosomes in Figure 7. As expected, strong hybridizing signals were visible for the entire length of chromosome 12, whereas there was no large, contiguous region present for any other chromosome. Thus, all three methods are consistent and suggest that only whole chromosome 12 is present in MCH clone p28, although we cannot exclude very small pieces of other human chromosomes or mosaicism.
Figure 7. Microarray analysis of MCH clone p28.
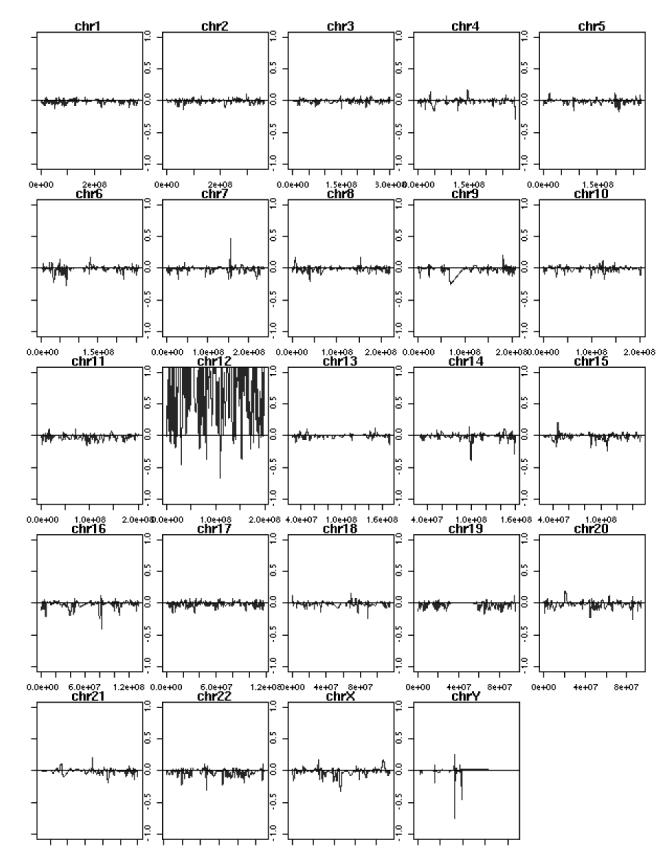
Total RNA was prepared, converted to cDNA, fluorescently labeled, and used to probe a U133 Affymetrix oligonucleotide array. Results are represented graphically as hybridization signal along each human chromosome, in numerical order per UCSC genome browser.
Use of codon-optimized gag constructs
It had been reported that expression of codon-optimized gag driven by the CMV immediate early enhancer-promoter led to Gag accumulation at the plasma membrane in both human and mouse cells (Hatziioannou et al., 2005; Swanson et al., 2004). We confirmed that result in the B78 parental mouse cells (Figure 8A). We next decided to test whether expression of CMV-driven, codon-optimized gag-pol would allow infectious virus release from mouse cells. B78 parental cells and MCH clones were transfected with CMV-gag-pol along with VSV G. These cells have an integrated HIV-CIB vector and already express Tat and Rev. As previously shown by others (Hatziioannou et al., 2005; Swanson et al., 2004), expression of the synthetic gag-pol boosted the amount of CA released from all of the mouse cell lines tested (Figure 8B), but it did not increase infectious HIV release from the B78 cells nor from any of the MCHs (Figure 8C). This suggests that high level expression of Gag-Pol and correct localization of Gag at the murine plasma membrane is not sufficient to rescue integrated HIV as an infectious particle. Similar results had been observed in the SCHs (Coskun et al., 2006).
Figure 8. Effects of synthetic gag-pol on HIV release.

(A) Parental B78 cells were transiently transfected with codon-optimized Gag-eGFP and then subjected to deconvolution microscopy 3 days later. (B, C) MCHs stably transduced with HIV-CIB and 293T cells transduced with HIV-LIB were transiently transfected with HIV-eYFP and pME-VSV G, with or without the Oxford Biomedica synthetic Gag-Pol (SGP) plasmid. (B) Supernatant (upper panel) and cell lysate (lower) were subjected to SDS-PAGE and immunoblotting as per figure 5. Lane 1: parental B78s; 2: B78s+SGP; 3: p11; 4: p11+SGP; 5: p11-R; 6: p11-R+SGP; 7: p28; 8: p28+SGP; 9: 293Ts. Short arrows indicate CA, * MA. (C) Supernatant harvested at 48 hours was titered on naïve human targets. Titer was normalized to transfection efficiency, as judged by epifluorescence microscopy of the producers. Open bars: eYFP titer; closed bars BlastiR titer. R denotes revertant. This experiment was repeated, with similar results.
Discussion
Mouse cells are not permissive for HIV replication, with a prominent block in the latter half of the viral life cycle. Here we used a genetic strategy to isolate three MCH cell clones that allowed efficient infectious HIV release. Production was independent of glycoprotein envelope used and specific to HIV. Amount of CA present in culture supernatants paralleled infectious virus. Importantly, revertant cell clones that no longer had any human chromosome component produced little HIV and CA. Differences in both intracellular Gag and unspliced/spliced viral RNA levels between the mouse cell clones were minor or inconsistent, whereas as previously reported human cells clearly had more unspliced relative to spliced viral RNA. A variety of genomic and chromosomal methods indicated that two of the three MCH clones were identical and had a single 2-12 chimera, whereas the third had solely intact 12. Although codon-optimized Gag-Pol was expressed at the plasma membrane and boosted CA production in all mouse cells tested, it could not rescue the replicative defect in that recoverable infectious titers were unaffected.
Because production of infectious virus may not correlate linearly with the amount of Gag due to cooperative or concentration-dependent assembly effects (Perez-Caballero et al., 2004), we went through some effort to show that compared to B78 parental cells and the MCH revertants the MCH clones i) do not have increased proviral copy number, ii) have roughly equivalent quantities of intracellular precursor Gag protein, and iii) have approximately the same amount of both unspliced and spliced viral RNAs (with the notable exception of p28-R which had more unspliced viral RNA). Importantly, transfection of codon-optimized, CMV promoter-driven HIV gag-pol did not fully rescue the mouse cell defect, as we had observed previously with other murine lines (Coskun et al., 2006).
At this point, however, we cannot exclude the possibility that slight differences in amount of unspliced RNA and p55 Gag are in fact responsible for much higher levels of infectious virus release, perhaps related to highly cooperative effects of Gag at the plasma membrane (Perez-Caballero et al., 2004). Thus, it is possible that one or more factors present on the human chromosome component in the MCH clones influences the amount of unspliced mRNA and Gag expression, either through transcriptional, anti-splicing, translational, or other poorly defined effects. It is clear that the ratio of unspliced to spliced viral RNAs remains low, even in the permissive MCH clones compared to the human cells, suggesting that correction of the ‘over-splicing’ phenomenon is not required to achieve reasonable infectious virus and CA production.
Previously it had been reported that CA production could be increased substantially in mouse cells using a viral RNA construct with a multimerized CTE (Swanson et al., 2004). Additionally, when a CMV-driven, codon-optimized gag plasmid was transfected into mouse cells, CA production was boosted and Gag associated with the plasma membrane (Hatziioannou et al., 2005; Swanson et al., 2004). Here, we confirmed the latter results. The fact that codon-optimized Gag-Pol did not increase infectious virus release in mouse cells suggests that at least one additional fundamental assembly defect may be present in mouse cells, perhaps unrelated to membrane targeting of Gag.
Localization of unspliced, cytosolic HIV mRNA has not been fully evaluated in human or mouse cells. In the case of MLV, use of the ms2 phage protein binding system has suggested unspliced RNA trafficks along an endosomal pathway and co-localizes with Gag, dependent upon MLV envelope protein expression (Basyuk et al., 2003). Determinants and required co-factors for mRNA entry into this pathway await elucidation. Compared to MLV, HIV has additional cis-acting nucleic acid sequences and trans-acting regulatory proteins that function in RNA nuclear export. These may play a role in directing unspliced RNA to a specific, post-nuclear pathway, perhaps in addition to other cellular factors that ‘mark’ the RNA (Swanson et al., 2004). Absence of one or more of these would impede or reduce infectious virus release from mouse cells. It is noteworthy that addition of a multimerized CTE to viral RNA allowed Gag to become localized to the plasma membrane and processed, but infectious virus was not quantified (Swanson et al., 2004).
Previously we had reported that SCHs containing chromosome 2 were permissive for HIV release, whereas the SCH with 12 had enhanced cyclin T1 function but did not produce more infectious virus. We were hoping that the MCHs would lead to improved genomic resolution of this phenotype, but unfortunately this was not the case (likely due to the nature of the cellular clones present in the original MCH pool). Here, two of the three MCHs isolated by the genetic screen appeared to be identical and had a single 2-12 chimeric chromosome, whereas the third only had an intact 12. For unknown reasons, the majority of the MCH clones present in this pool of 500 have a fragment or most of 12 (Speevak et al., 1995; Speevak and Chevrette, 1996). The fact that nearly all of the MCHs and a small panel of randomly chosen MCHs that have most of 12 do not support infectious virus release suggests that 12 is not causative.
We wish to emphasize that we cannot yet rule out the possibility that a gene on 12 is responsible for the observed phenotype, especially since the very permissive clone p28 only has 12 as determined by a variety of low resolution methods. It is also conceivable that there is a small, unresolvable piece of another chromosome in p28 or that p28 is a mosaic such that a fraction of the cells has another chromosome fragment. For example, the PCR primers used had an average spacing of 10 cM and the whole chromosome 2 FISH hybridization has a resolution of ∼20 Mb, depending upon the signal amplification methods used. The results from the U133 microarray suggest that if p28 does have a segment of 2 it is likely no greater than a few Mb in length, which could have been easily missed by the PCR and FISH methods. Mosaicism at the level of 10% or less would also have gone undetected. p28 revertants that have lost 12 and the HygroTK marker are non-permissive, suggesting that the responsible factor(s) is genetically linked to 12 and the selectable gene.
We now have several mouse-human hybrid cell lines, each with a single human chromosome, that are permissive for HIV and CA production. The question remains as to the identity of the gene(s) involved. Both Crm1, which binds to Rev to allow export of RRE-containing mRNAs (Fornerod et al., 1997; Fukuda et al., 1997), and hRIP, involved in the movement of Rev/RRE-dependent HIV RNAs from the nuclear periphery to the cytoplasm (Sanchez-Velar et al., 2004), are located on opposite ends of 2. Introduction of these genes by transient transfection into mouse cells, alone and in combination, failed to increase CA or infectious virus release (not shown). SAM68 may cooperate with Rev/RRE to allow more efficient nuclear export of unspliced mRNAs (Modem et al., 2005; Reddy et al., 1999). That gene is not present in any of the MCHs or SCHs of interest, and transient transfection of SAM68 into mouse cells failed to increase CA or infectious HIV release (not shown). Other genes implicated in RNA export encoded on chromosome 2 (Fang et al., 2004) have not been tested.
We and others (Ned Landau and Paul Bieniasz, personal communications) have attempted without success direct and exhaustive cDNA and genetic complementation approaches to identify possible missing or altered factors in mouse cells. Radiation-hybrid mapping using the Stanford G3 hamster-human panel failed to achieve highly significant LOD scores (not shown). We are now faced with the prospect of engineering the single human chromosome present in the permissive MCHs and SCHs to further localize the gene(s) responsible, which could be a daunting task. These cell lines, however, may also serve as useful reagents to mechanistically dissect the post-integration block to HIV production in murine cells.
Materials and Methods
Plasmids and viral vectors
HIV vectors pHIV-cycT1-IRES-bsd (pHIV-CIB), pHIV-luciferase-IRES-bsd (pHIV-LIB), and pHIV-IRES-eYFP have been described (Coskun and Sutton, 2005), as has murine leukemia virus (MLV) vector pBABE-IY (Segall, Yoo, and Sutton, 2003.). M-tropic HIV envelope ADA expression plasmid was a gift from Dan Littman. pHIT60 is an expression construct of MLV Gag-Pol (Soneoka et al., 1995), and pME VSV G is a VSV glycoprotein expression plasmid. CMV-driven codon-optimized (synthetic) gag-pol and gag-eGFP were gifts of Oxford Biomedica (Kotsopoulou et al., 2000) and Paul Bieniasz, respectively.
Cells and production of pseudotyped particles
Cells were maintained in Dulbecco's modified Eagle's high-glucose medium supplemented with 10% fetal calf serum, penicillin, streptomycin, and ciprofloxacin (termed DMEM complete), with other antibiotic supplements as necessary. Cells were grown in 5% CO2, 37°C water-jacketed incubators and passaged twice weekly using trypsin-EDTA. Pool of 500 MCHs were originally obtained from Mario Chevrette (Speevak et al., 1995) and maintained in complete DMEM supplemented with 400 μg/ml hygromycin B (Calbiochem). In order to revert the MCHs and lose the human chromosome component, cells were passaged at low confluency into complete DMEM plus 35 μM ganciclovir. Ganciclovir-resistant colonies typically arose after 10-12 days at a frequency of ∼0.01% and were shown to be sensitive to hygromycin B, consistent with loss of the human chromosome.
Retroviral and lentiviral pseudotyped particles were produced by calcium phosphate co-transfection of 293T cells with the appropriate viral envelope (here typically VSV G) and vector (Sutton and Littman, 1996). For MLV, pHIT60 was also included in the transfection. After 72 hours, vector supernatant was harvested as previously described (Sutton and Littman, 1996). Target cells were transduced in 18 or 35 mm dish format and 48 hours later passaged into selective medium (typically DMEM complete supplemented with 10 μg/ml of blasticidin S (Invitrogen or ICN).
In order to recover and titer integrated HIV vector encoding bsd from cell lines, cells were plated one day prior in 35 mm or 10 cm format, transfected the next day at high confluence with 1-2 μg each of the viral glycoprotein envelope plasmid (e.g., pME VSV G) along with an autofluorescent control (e.g., HIV-IRES-eYFP) using Lipofectamine 2000 reagent (Invitrogen) and following the manufacturer's instructions. Forty-eight hours later culture supernatant was harvested, centrifuged at 2000 × g for 10 minutes, and inoculated onto adherent human targets, typically HOS TK- or 293T cells. Transfected cells were examined after supernatant harvest by epifluorescence microscopy or subjected to flow cytometry using a FACScan (Becton-Dickinson) to determine overall transfection efficiency. Target human cells were passaged into DMEM complete containing 10 μg/ml of blasticidin S two days later. Cells were fixed in methanol and acetic acid, stained with crystal violet, and colonies enumerated after 7-9 days.
Analysis of CA
Amount of CA produced by vector-transduced or infected cells was quantified by commercial ELISA (Beckman-Coulter) or by immunoblotting. For the latter, RIPA cell lysates were size-separated by SDS-PAGE and transferred to nitrocellulose. Membrane was blocked using Tris-buffered saline in the presence of 0.02% Tween-20 and 2.5% nonfat dried milk (TBS-T+NFDM), probed using primary human anti-HIV antibody from the AIDS Research and Reference Reagent Program (Cat. No. 3957) at 1:1000 in the same buffer, washed extensively in TBS-T, probed using secondary rabbit anti-human IgG antibody conjugated to HRP (Sigma) at 1:10000, washed, and developed using enhanced chemiluminescence.
Nucleic acid analyses
Genomic DNA was prepared from cell lines as described (Coskun et al., 2006). PCR for mouse and human ß-actin was performed as described (Coskun et al., 2006). HIV DNA primers for PCR amplification were NL4-3 831L (Sutton et al., 1998) and NL4-3 569U (Coskun et al., 2006), using conditions 94°C for 30 sec, 62°C for 30 sec, and 68°C for 30 sec for 31 cycles and Expand Long DNA polymerase (Roche) to give rise to a ∼260 bp product, which was analyzed by agarose gel electrophoresis and ethidium bromide staining. Positive control cell line had a single HIV vector integrant. Gels were Southern blotted at high stringency using the corresponding PCR product as 32P-labeled probe, amplified from HIV vector plasmid template.
Total RNA was prepared from cell lines as described previously (Coskun et al., 2006). First strand reaction was performed using Superscript II RT (Invitrogen) using 1-3 μg of total RNA, and oligo(dT) as primer for 60 min at 42 °C. DNA PCR for both mouse and human β-actin was performed as described above for 21 cycles using three different amounts of first strand reaction. For HIV, primers spanning the tat-rev intron were 5'-CTGTGGCATTGAGCAAGCTAACAGCAC-3' and 5'-ACAGCGACGAAGAGCTCATCAGAACAG-3'. Amplification conditions were 94°C for 30 sec, 62°C for 30 sec, and 68°C for 60 sec for 35 cycles using Expand Long DNA polymerase and the equivalent of 15, 45, and 145 ng of total RNA from the first strand reaction. This resulted in DNA products of 1250 and 350 bp, corresponding to unspliced and spliced viral mRNAs, respectively. The spliced 350 bp product was directly visualized by agarose gel electrophoresis and EtBr staining. For the unspliced product, Southern blotting was performed using a 0.7 kb Hind III fragment specific to the intron as a probe (Coskun et al., 2006). As a control for contaminating DNA, Superscript II was omitted from the first strand reaction. Total RNA from 293T cells that had been transduced with an HIV vector encoding both luciferase and bsd was used as a positive control for both spliced and unspliced viral RNAs.
Genome-wide nucleic acid and chromosome analyses
PCR was performed on genomic DNA using the ABI PRISM Linkage Mapping Set (Applied Biosystems) following the manufacturer's recommendations for amplification conditions. DNA products (typically less than 300 bp in length) were size-separated by agarose gel electrophoresis and stained with ethidium bromide (EtBr). Water was included as a negative control and genomic DNA from 293T cells served as a positive control.
For metaphase spread preparations, cells were incubated in 10 μg/ml colcemid solution (Invitrogen) for 20 min at 37°C. Cells were trypsinized, treated with 75 mM KCl hypotonic solution for 20 minutes, and fixed with 3:1 methanol:glacial acetic acid. FISH analyses were performed using whole chromosome painting (WCP) probes for human chromosome 2 (Vysis) or to labeled BACs (purchased from Children's Hospital Oakland Research Institute, Oakland, California), according to manufacturer' protocol and examined using fluorescence microscopy. For each cell line 25-50 representative metaphase spreads were examined.
Microarray analyses were performed using total RNA converted to fluorescently labeled cDNA and hybridized to the U133 Plus 2.0 Affymetrix array chip. Raw hybridization scores for individual genes were plotted in numerical order (numbering per UCSC genome browser) for the entire set of chromosomes, with each chromosome represented graphically by a single line.
Acknowledgements
We thank Mario Chevrette (McGill University), Dan Littman (NYU School of Medicine), Paul Bieniasz (Aaron Diamond AIDS Research Center), and Oxford Biomedica for generous reagent gifts. We acknowledge the assistance of Chad Shaw (Baylor) for the chromosome/gene ordering of the microarray data. The following reagents were obtained from the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: HIV-IG from NABI and NHLBI, GHOST HI5 cells from Vineet KewalRamani (NCI-Frederick) and Dan Littman. This work was initially supported by the American Foundation for AIDS Research (AmfAR) and then the National Institutes of Health and the Edward Mallinckrodt, Jr. Foundation. R.E.S. was an Edward Mallinckrodt, Jr. Foundation Scholar.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Basyuk E, Galli T, Mougel M, Blanchard JM, Sitbon M, Bertrand E. Retroviral genomic RNAs are transported to the plasma membrane by endosomal vesicles. Dev Cell. 2003;5(1):161–74. doi: 10.1016/s1534-5807(03)00188-6. [DOI] [PubMed] [Google Scholar]
- Bieniasz PD, Cullen BR. Multiple blocks to human immunodeficiency virus type 1 replication in rodent cells. J Virol. 2000;74(21):9868–77. doi: 10.1128/jvi.74.21.9868-9877.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bray M, Prasad S, Dubay JW, Hunter E, Jeang KT, Rekosh D, Hammarskjold ML. A small element from the Mason-Pfizer monkey virus genome makes human immunodeficiency virus type 1 expression and replication Rev-independent. Proc. Natl. Acad. Sci. USA. 1994;91:1256–1260. doi: 10.1073/pnas.91.4.1256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen BK, Rousso I, Shim S, Kim PS. Efficient assembly of an HIV-1/MLV Gag-chimeric virus in murine cells. Proc Natl Acad Sci U S A. 2001 doi: 10.1073/pnas.261563198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coskun AK, Sutton RE. Expression of glucose transporter 1 confers susceptibility to human T-cell leukemia virus envelope-mediated fusion. J Virol. 2005;79(7):4150–8. doi: 10.1128/JVI.79.7.4150-4158.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coskun AK, van Maanen M, Nguyen V, Sutton RE. Human chromosome 2 carries a gene required for production of infectious human immunodeficiency virus type 1. J Virol. 2006;80(7):3406–15. doi: 10.1128/JVI.80.7.3406-3415.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullen BR. Nuclear mRNA export: insights from virology. Trends Biochem Sci. 2003;28(8):419–24. doi: 10.1016/S0968-0004(03)00142-7. [DOI] [PubMed] [Google Scholar]
- Fang J, Kubota S, Yang B, Zhou N, Zhang H, Godbout R, Pomerantz RJ. A DEAD box protein facilitates HIV-1 replication as a cellular co-factor of Rev. Virology. 2004;330(2):471–80. doi: 10.1016/j.virol.2004.09.039. [DOI] [PubMed] [Google Scholar]
- Fornerod M, Ohno M, Yoshida M, Mattaj IW. CRM1 is an export receptor for leucine-rich nuclear export signals [see comments] Cell. 1997;90(6):1051–60. doi: 10.1016/s0092-8674(00)80371-2. [DOI] [PubMed] [Google Scholar]
- Fukuda M, Asano S, Nakamura T, Adachi M, Yoshida M, Yanagida M, Nishida E. CRM1 is responsible for intracellular transport mediated by the nuclear export signal. Nature. 1997;390(6657):308–11. doi: 10.1038/36894. [DOI] [PubMed] [Google Scholar]
- Garber ME, Wei P, KewalRamani VN, Mayall TP, Herrmann CH, Rice AP, Littman DR, Jones KA. The interaction between HIV-1 tat and human cyclin T1 requires zinc and a critical cysteine residue that is not conserved in the murine CycT1 protein [In Process Citation] Genes Dev. 1998;12(22):3512–27. doi: 10.1101/gad.12.22.3512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatziioannou T, Martin-Serrano J, Zang T, Bieniasz PD. Matrix-induced inhibition of membrane binding contributes to human immunodeficiency virus type 1 particle assembly defects in murine cells. J Virol. 2005;79(24):15586–9. doi: 10.1128/JVI.79.24.15586-15589.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubner W, Chen BK. Inhibition of viral assembly in murine cells by HIV-1 matrix. Virology. 2006;352(1):27–38. doi: 10.1016/j.virol.2006.04.024. [DOI] [PubMed] [Google Scholar]
- Kotsopoulou E, Kim VN, Kingsman AJ, Kingsman SM, Mitrophanous KA. A Rev-independent human immunodeficiency virus type 1 (HIV-1)-based vector that exploits a codon-optimized HIV-1 gag-pol gene. J Virol. 2000;74(10):4839–52. doi: 10.1128/jvi.74.10.4839-4852.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mariani R, Rutter G, Harris ME, Hope TJ, Krausslich HG, Landau NR. A block to human immunodeficiency virus type 1 assembly in murine cells. J Virol. 2000;74(8):3859–70. doi: 10.1128/jvi.74.8.3859-3870.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modem S, Badri KR, Holland TC, Reddy TR. Sam68 is absolutely required for Rev function and HIV-1 production. Nucleic Acids Res. 2005;33(3):873–9. doi: 10.1093/nar/gki231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ono A, Freed EO. Binding of human immunodeficiency virus type 1 Gag to membrane: role of the matrix amino terminus. J Virol. 1999;73(5):4136–44. doi: 10.1128/jvi.73.5.4136-4144.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paillart JC, Gottlinger HG. Opposing effects of human immunodeficiency virus type 1 matrix mutations support a myristyl switch model of gag membrane targeting. J Virol. 1999;73(4):2604–12. doi: 10.1128/jvi.73.4.2604-2612.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez-Caballero D, Hatziioannou T, Martin-Serrano J, Bieniasz PD. Human immunodeficiency virus type 1 matrix inhibits and confers cooperativity on gag precursor-membrane interactions. J Virol. 2004;78(17):9560–3. doi: 10.1128/JVI.78.17.9560-9563.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy TR, Xu W, Mau JK, Goodwin CD, Suhasini M, Tang H, Frimpong K, Rose DW, Wong-Staal F. Inhibition of HIV replication by dominant negative mutants of Sam68, a functional homolog of HIV-1 Rev. Nat Med. 1999;5(6):635–42. doi: 10.1038/9479. [DOI] [PubMed] [Google Scholar]
- Reed M, Mariani R, Sheppard L, Pekrun K, Landau NR, Soong NW. Chimeric Human Immunodeficiency Virus Type 1 Containing Murine Leukemia Virus Matrix Assembles in Murine Cells. J Virol. 2002;76(1):436–443. doi: 10.1128/JVI.76.1.436-443.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez-Velar N, Udofia EB, Yu Z, Zapp ML. hRIP, a cellular cofactor for Rev function, promotes release of HIV RNAs from the perinuclear region. Genes Dev. 2004;18(1):23–34. doi: 10.1101/gad.1149704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segall H, Yoo E, Sutton RE. Characterization and detection of artificial replication-competent lentivirus of altered host range. Molecular Therapy. 2003;8:118–129. doi: 10.1016/s1525-0016(03)00134-5. [DOI] [PubMed] [Google Scholar]
- Soneoka Y, Cannon PM, Ramsdale EE, Griffiths JC, Romano G, Kingsman SM, Kingsman AJ. A transient three-plasmid expression system for the production of high titer retroviral vectors. Nucleic Acids Res. 1995;23(4):628–33. doi: 10.1093/nar/23.4.628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spearman P, Horton R, Ratner L, Kuli-Zade I. Membrane binding of human immunodeficiency virus type 1 matrix protein in vivo supports a conformational myristyl switch mechanism. J Virol. 1997;71(9):6582–92. doi: 10.1128/jvi.71.9.6582-6592.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Speevak MD, Berube NG, McGowan-Jordan IJ, Bisson C, Lupton SD, Chevrette M. Construction and analysis of microcell hybrids containing dual selectable tagged human chromosomes. Cytogenetics and Cell Genetics. 1995;69:63–65. doi: 10.1159/000133939. [DOI] [PubMed] [Google Scholar]
- Speevak MD, Chevrette M. Human chromosome 3 mediates growth arrest and suppression of apoptosis in microcell hybrids. Mol Cell Biol. 1996;16(5):2214–25. doi: 10.1128/mcb.16.5.2214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutton RE, Littman DR. Broad host range of human T-cell leukemia virus type 1 demonstrated with an improved pseudotyping system. J. Virol. 1996;70:7322–7326. doi: 10.1128/jvi.70.10.7322-7326.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutton RE, Wu HT, Rigg R, Bohnlein E, Brown PO. Human immunodeficiency virus type 1 vectors efficiently transduce human hematopoietic stem cells. J Virol. 1998;72(7):5781–5788. doi: 10.1128/jvi.72.7.5781-5788.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swanson CM, Puffer BA, Ahmad KM, Doms RW, Malim MH. Retroviral mRNA nuclear export elements regulate protein function and virion assembly. Embo J. 2004;23(13):2632–40. doi: 10.1038/sj.emboj.7600270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei P, Garber ME, Fang SM, Fischer WH, Jones KA. A novel CDK9-associated C-type cyclin interacts directly with HIV-1 Tat and mediates its high-affinity, loop-specific binding to TAR RNA. Cell. 1998;92(4):451–62. doi: 10.1016/s0092-8674(00)80939-3. [DOI] [PubMed] [Google Scholar]