

Abstract
The catalytic subunit, gp91phox (a.k.a., Nox2) of the NADPH-oxidase of mammalian phagocytes is activated by microbes and immune mediators to produce large amounts of reactive oxygen species (ROS) which participate in microbial killing. Homologs of gp91phox, the Nox and Duox enzymes, were recently described in a range of organisms, including plants, vertebrates, and invertebrates such as Drosophila melanogaster. While their enzymology and cell biology is being extensively studied in many laboratories, little is known about in vivo functions of Noxes. Here, we establish and use an inducible system for RNAi to discover functions of dNox, an ortholog of human Nox5 in Drosophila. We report here that depletion of dNox in musculature causes retention of mature eggs within ovaries, leading to female sterility. In dNox-depleted ovaries and ovaries treated with a Nox inhibitor, muscular contractions induced by the neuropeptide proctolin are markedly inhibited. This functional defect results from a requirement for dNox for the proctolin-induced calcium flux in Drosophila ovaries. Thus, these studies demonstrate a novel biological role for Nox-generated ROS in mediating agonist-induced calcium flux and smooth muscle contraction.
INTRODUCTION
Reactive oxygen species (ROS) are generally thought of as being broadly reactive, mutagenic and cytotoxic entities that are produced largely as an “accidental” byproduct of aerobic metabolism. While such “unintentional” production of ROS clearly occurs, it is also known that ROS can be produced in a regulated manner and serve useful biological purposes. The classical example of this is the NADPH oxidase of professional phagocytes, of which gp91phox is the catalytic subunit. This enzyme produces large amounts of ROS that form an important part of the phagocyte’s bactericidal machinery (2, 42). Observations in many other cell types in addition to phagocytes of the regulated production of ROS by NADPH oxidase-like enzymatic sources led to the discovery that gp91phox is in fact a member of a family of NADPH oxidases termed the Nox/Duox family, which have now been identified in a wide range of organisms including plants, vertebrate and invertebrate animals (21).
The human genome encodes 7 homologs in the Nox/Duox family termed Nox 1–5, Duox1 and Duox2 (21). Nox1–4 consist of 6 transmembrane α-helices that bind 2 hemes plus an FAD domain containing an NADPH binding site (21). Nox1–3 are activated by regulatory subunits, while Nox4 is constitutively active (2, 7, 15). The N-terminus of Nox5 contains an additional EF-hand-containing calcium-binding domain that mediates its calcium-dependent activation (3, 4). Duoxes build further upon the Nox5 structure with an additional N-terminal peroxidase domain (8).
While the enzymology and cell biology of Noxes is being extensively studied, little is known about their in vivo functions. In recent years, studies using cultured mammalian cells have supported a role for reactive oxygen species as as potential signaling molecules. ROS are produced by cells in a regulated manner in response to a wide range of growth factors and hormones. Oxidation by ROS of key amino acid residues such as low pKa cysteines may mediate the signaling by of ROS by altering the activity of downstream enzymes and transcription factors. NADPH oxidases are implicated as the source of agonist-induced ROS in a variety of cells, including insulin stimulation of adipocytes (26) and Angiotensin II stimulation of vascular smooth muscle cells (28). While cell culture experiments support a role for Noxes in cell signaling, so far there is very little in vivo data regarding signaling or other non-immune functions for these enzymes.
The Drosophila genome encodes only 2 Noxes rather than the 7 seen in humans: dNox, an ortholog of h-Nox5, and dDuox. The relative simplicity of the Drosophila genome and the availability of genetic tools in fly provided an opportunity to explore the biological functions of Noxes in an intact animal. We used RNAi coupled to the GAL-UAS binary system to conditionally knock down the expression of dNox in fly. RNAi of dNox resulted in a marked deficit in female egg-lying caused by a defect in ovulation, which was due to the failure of ovarian muscles to contract in an agonist-dependent manner. We also show herein that dNox regulates an agonist-stimulated calcium flux in ovarian tissue. This study provides some of the first definitive in vivo evidence of a role for Nox enzymes in cell signaling.
MATERIALS AND METHODS
Fly stocks and crosses
Drosophila were cultured on standard cornmeal food at 25°C. Transgenic UAS-dNox-IR flies were generated by subcloning a 1,000 bp portion of the dNox coding sequence in an inverted repeat orientation, separated by a 330 bp spacer sequence downstream of the UAS element in the pUAST vector (34). The first 1,000 bp of the dNox coding sequence was amplified by polymerase chain reaction (PCR) using primer sets 5′-gcatgaattcatggactttgccgagcaaatt-3′ 5′-gcataagcttggacaacccacgacccctg-3′ and 5′-gcatggtaccatggactttgccgagcaaatt-3′ 5′gcatctcgagggacaacccacgacccctg-3′. 330 bp of green fluorescent protein sequence was amplified by PCR for use as spacer sequence using primers 5′-gcataagctttgcttcagccgctaccccga-3′ and 5′-gcatctcgagggcgagctgcacgc-3′. The resulting PCR fragments were digested using appropriate restriction enzymes and cloned sequentially into equivalent sites in Bluescript SK. The completed dNox sense – spacer – dNox antisense fragment was restricted from Bluescript using EcoRI and KpnI and inserted into the equivalent sites in pUAST. Transformation of Drosophila embryos was carried out in the w1118 stock using standard techniques (39). All insertions in transgenic stocks were mapped to a chromosome and either made homozygous or balanced with appropriate balancer chromosomes. GAL4 drivers TubP, c179 and c855a, and 49A UAS-aequorin flies were obtained from the Bloomington Stock Center. The Nrv1-GAL4 driver was a gift from Paul Salvaterra. Adult flies carrying one copy of a UAS-dNox-IR construct and one copy of a GAL4 driver were used for the assays. For aequorin assays, male flies carrying one copy of UAS-dNox-IR and one copy of Nrv1-GAL4 were generated and subsequently crossed with females homozygous for the UAS-aequorin insertion. From this cross, all progeny carried a copy of UAS-aequorin, and flies also carrying one copy of both Nrv1-GAL4 and UAS-dNox-IR were selected based on an easily recognized head morphology defect that occurs when dNox RNAi is driven by Nrv1-GAL4.
Fertility and ovulation assays
To determine fertility, virgin females of the indicated genotype were collected and crossed 5 per vial with 5 w1118 males. After 24 hours, flies were cleared from vials and the total number of progeny resulting from crosses was determined 14 days later. To determine egg-laying ability, 50 virgin females of the appropriate genotype were crossed to w1118 males on grape juice medium with fresh yeast, and the total number of eggs laid was determined 24 hours later. For ovulation assays, 4–5 day old virgin females were crossed individually with w1118 males for 1 hour and then placed on standard medium. At the indicated times, the females were dissected and scored positive for ovulation if an egg was present in the common oviduct or uterus.
Ovary contraction assays
Virgin females of the appropriate genotype were collected and placed on standard medium with freshly added yeast for 3–5 days. Ovaries were dissected in Schneider’s Drosophila medium (Invitrogen). Where indicated, ovaries were pre-incubated with 20 μM diphenylene iodinium (DPI) (Sigma) for 15 minutes prior to the addition of 1 μM proctolin (Phoenix Pharmaceuticals). Movements of ovaries were recorded as QuickTime movies using a Nikon Coolpix 4500 camera. Movies were analyzed for ovarian movement using Videopoint software. The tip of one ovary at t0 was defined as the origin and the location of that same ovarian tip relative to the origin was measured every 0.1 seconds. Data were recorded as distance from origin in pixels and were graphed as a function of time using GraphPad Prism 4 software.
Superoxide generation assays
Virgin w1118 females were collected and placed on standard medium with freshly added yeast for 3–5 days. Ovaries were dissected in Schneider’s Drosophila medium and placed 3 to a well in a white 96 well plate and 50 μM L-012 (31) (Wako Chemicals) was added. Where indicated M40403 (gift from Altana Pharma), BAPTA-AM (Calbiochem) and DPI were pre-incubated with tissue for 15 minutes prior to the start of the assay. L-012 luminescence was read using a FLUOstar OPTIMA (BMG Labtech) plate-based luminometer. Proctolin and ionomycin (Sigma) were injected automatically at the times indicated.
Aequorin assays
Virgin females were collected and placed on standard medium with freshly added yeast for 3–5 days. Ovaries were dissected in Schneider’s Drosophila medium and placed 3 to a well in a white 96 well plate. Ovaries were incubated in 2.5 μM native coelenterazine (Sigma) for 5 hours while gently shaking in the dark at RT. Aequorin luminescence was read using a FLUOstar OPTIMA (BMG Labtech) plate-based luminometer using the well mode. Where indicated, ovaries were pre-incubated with DPI at the indicated final concentration for 15 minutes. Proctolin and ionomycin were added where indicated using pumps built into the luminometer.
RESULTS
Establishment of a System for RNAi of dNox
When coupled to the GAL4/UAS binary system in Drosophila, RNA interference is specific, efficient, stable, heritable, and conditional based on the spatial and temporal patterns of GAL4 expression. This method has been successfully applied to loss of function studies for many genes in Drosophila, including EcR, β-FTZ-F1 (20), yellow (35), and dFADD (24).
We generated ~50 transgenic stocks harboring a dNox-specific inverted repeat (IR) sequence under the control of UAS. When crossed with flies expressing GAL4, dNox RNAi is produced in progeny in a pattern reflecting GAL4 expression. These lines were first crossed with flies expressing GAL4 ubiquitously under the control of the tubulin promoter (tubP-GAL4). Resulting dNox RNAi progeny were viable and produced apparently normal adults except for enlarged heads and subtle defects in clypeus morphology (data not shown). The efficacy of RNAi in reducing dNox mRNA in adults was evaluated using quantitative RT-PCR. The rp49 ribosomal RNA was used as an internal control for the concentration of RNA in all samples. This analysis revealed reduction of dNox expression by 75% to 90% in individual stocks (Fig. 1A).
Fig. 1.
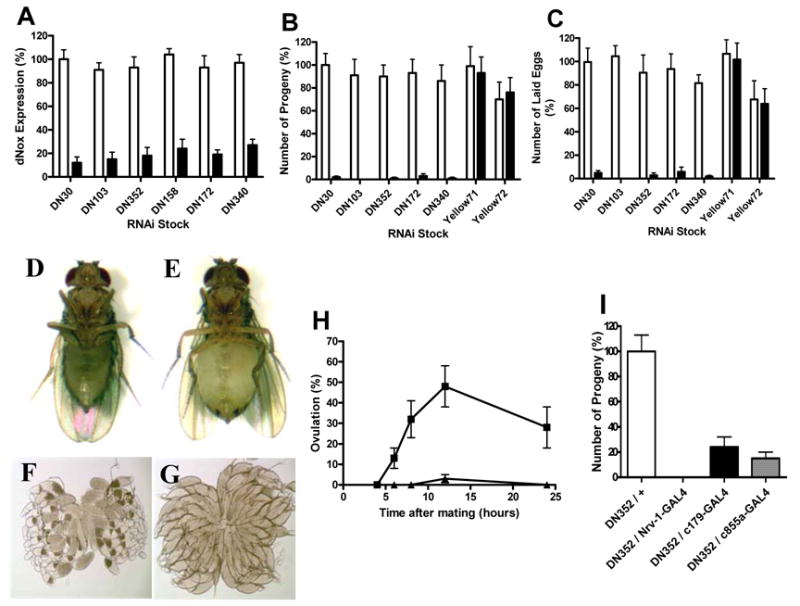
Muscle dNox regulates ovulation. (A–C) White bars, UAS-dNox-IR; black bars, UAS-dNox-IR + TubP-GAL4. (A) Quantitative RT-PCR of dNox in adult flies. (B) Progeny produced from crosses of control and dNox RNAi females with w1118 males. (C) Eggs laid by control and dNox RNAi females mated with w1118 males. (D–H) Control, UAS-DN352-IR; dNox RNAi, UAS-DN352 + TubP-GAL4. (D–E) Photographs of 10 day old control (D) and dNox RNAi (E) females. (F–G) Light micrographs of ovaries dissected from mated control (F) and dNox RNAi (G) females. (H) Ovulation rate of control and dNox RNAi females at various time points after crossing with w1118 males. Squares, control; triangles, dNox RNAi. (I) Progeny produced from crosses of females of the indicated genotype with w1118 males.
RNAi of dNox Causes Female Sterility
Control (UAS-dNox-IR) and dNox-RNAi (UAS-dNox-IR + TubP-GAL4) flies were generated as above through crosses of UAS-dNox-IR flies with TubP-Gal4 flies. Control and dNox-RNAi males and females were crossed separately with wild-type w1118 flies to assess fertility. While dNox-RNAi males were fully fertile compared to control males (data not shown), dNox-RNAi females produced very few or no progeny (Fig. 1B). This was true for all five dNox-RNAi stocks tested, showing that the phenotype is independent of the position of the transgene insertion. As a control for all fertility experiments, stocks that induce a similarly structured double stranded RNA targeting the yellow pigment gene were used (35), and showed no effect on female fertility. Further analysis revealed that dNox-RNAi-mediated female sterility is fully accounted for by a defect in egg-laying (Fig. 1C).
RNAi of dNox Results in a Specific Defect in Ovulation
A deficiency in egg-laying can result either from a defect in oogenesis (formation or maturation of eggs) or from a defect in deposition of mature oocytes. Drosophila ovaries consist of bundles of individual egg “assembly lines” termed ovarioles. Developing oocytes progress through 14 characteristic stages of development as they pass down ovarioles from the germarium in the ovary tip where the germ stem cells reside, toward the calyx area, where fully developed oocytes enter the lateral oviducts (6).
We observed that dNox-RNAi females had markedly expanded abdomens several days after eclosion (Fig. 1E) compared to controls (Fig. 1D). Dissected ovaries revealed that ovaries from control females contained oocytes at all stages of development with no more than one stage 14 oocyte per ovariole (Fig. 1F). In contrast, ovaries from dNox-RNAi ovaries lacked intermediate stage oocytes but contained 3–4 stage 14 oocytes per ovariole (Fig. 1G). The few eggs that are laid by dNox-RNAi females have normal hatching rates compared with eggs laid by control females, indicating that eggs produced by dNox-RNAi females are fully viable. Thus, while oocyte maturation is apparently normal in dNox RNAi females and these eggs are fully viable, most mature eggs fail to be deposited.
Egg-laying in Drosophila consists of ovulation (movement of egg from ovary to the uterus through the lateral and common oviducts) and oviposition (deposition of egg from uterus to the substrate). Ovulation occurs at a very low rate in unmated Drosophila females and is triggered upon mating (6).
Distended abdomens and “backed-up” ovarioles are characteristic phenotypes of mutant flies that have previously been shown to have ovulation defects (22, 29). Therefore, we investigated whether the dNox-RNAi egg-laying defect derived from a defect in ovulation. To determine the disruption in RNAi animals, control and RNAi females were mated to w1118 males and ovulation rates were compared following mating by scoring dissected ovaries for the presence of an egg(s) in the oviducts or uterus. dNox RNAi females mated normally, but ovulated at markedly lower rates (Fig. 1H), indicating that it is the movement of the egg from ovary into oviducts that is defective.
RNAi of dNox Blunts Proctolin-Induced Ovarian Muscle Contractions
Ovulation has not been studied extensively in Drosophila, but studies in other insects indicate that ovulation involves rhythmic contraction of visceral musculature surrounding the ovarioles and oviducts (6). These contractions regulate passage of developing oocytes along the ovariole as well as into the lateral oviducts. Insect ovarian muscle contraction is stimulated by neuropeptides that are released by local neurons and/or by neurohormones distributed through the hemolymph (16, 32, 33)
We screened tissue-specific GAL4 drivers to explore further how dNox regulated ovulation. About 250 tissue specific GAL4 drivers were used, including many that are either neuron- or muscle-specific. This screen uncovered three drivers with muscle expression (Nrv1 (48), c179 (27) and c855a (13)) that recapitulated the dNox sterility phenotype seen with the ubiquitous tubP-GAL4 driver (Fig. 1I). No other drivers, including 23 neuron-specific drivers tested (including the widely used elav-GAL4 and Nrv2-GAL4) caused the sterility phenotype (not shown). The Nrv1-GAL4 driver fully recapitulated the ovulation defect, evidenced by severely “backed-up” ovarioles, as seen with the TubP-GAL4 driver (Supp. Fig. 1).
The effect of dNox RNAi on proctolin-induced ovarian muscle contraction was then evaluated. Ovary pairs were dissected from females and placed in Drosophila S2 culture medium and their movement in response to ex vivo treatments was recorded. Movement in excised ovaries from control (UAS-dNox-IR) and RNAi (UAS-dNox-IR + Nrv1-GAL4) females was recorded without and with proctolin (movies in Supplemental Data). In the absence of agonists, control ovaries showed little or no movement (Fig. 2A), while proctolin induced rhythmic, high amplitude contractions (Fig. 2B). The effect of proctolin on muscle contractions was markedly attenuated in ovaries expressing dNox RNAi (Fig. 2C) and in those pre-treated with the general Nox inhibitor diphenylene iodinium (DPI) (Fig. 2D). Staining of ovaries with FITC-labeled phalloidin did not reveal any differences in muscle structure of ovaries between control (Supp. Fig. 2A) and dNox RNAi (Supp. Fig. 2B) animals indicating dNox RNAi does not affect ovarian muscle development. Immunofluorescence using anti-proctolin receptor antibody (14) also showed normal receptor expression in dNox RNAi ovaries (Supp. Fig. 3B) compared with control (Supp. Fig. 3A). Thus, dNox regulates ovarian muscle contractions in response to the neuropeptide, proctolin.
Fig. 2.
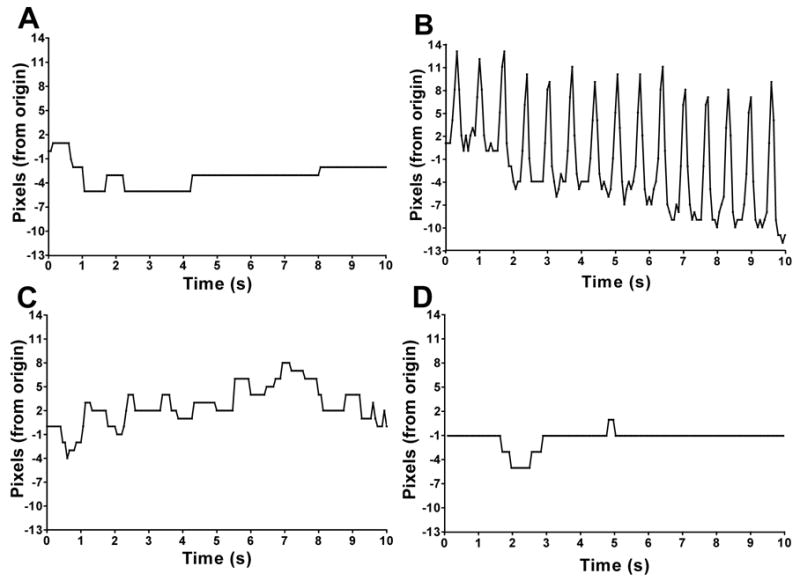
dNox regulates proctolin-induced muscle contractions in ovaries. (A–D) Graphs of ovarian movement caused by muscle contractions. (A) UAS-DN352-IR ovary with no stimulus. (B) UAS-DN352-IR ovary + 1 μM proctolin. (C) UAS-DN352-IR + Nrv1-GAL4 ovary + 1 μM proctolin. (D) UAS-DN352-IR ovary + 20 μM DPI + 1 μM proctolin.
Fig. 3.
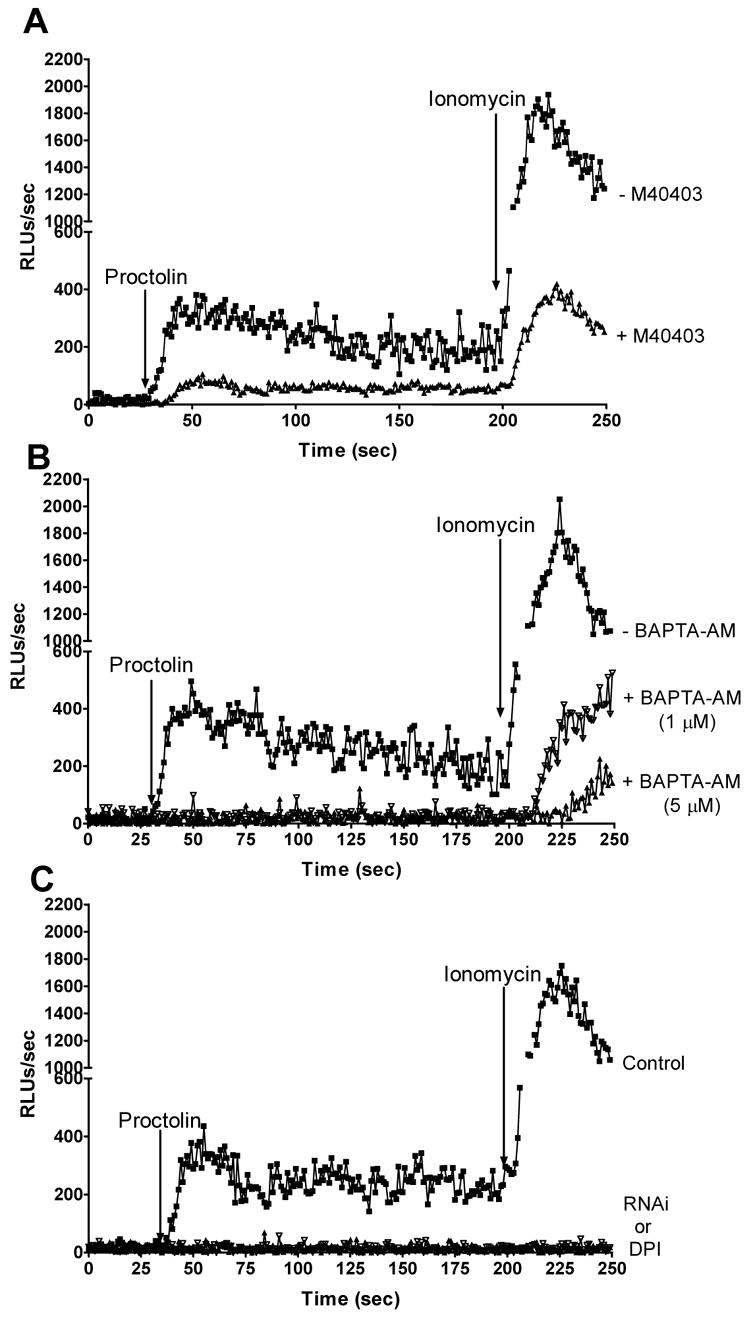
Proctolin activates ROS production through dNox. (A–C) L-012 luminescence after addition of 1 μM proctolin and 1 μM ionomycin. Control, UAS-DN352-IR; dNox RNAi, UAS-DN352 + TubP-GAL4. (A) Squares, control ovaries; triangles, control ovaries + 20 μM M40403. (B) Squares, control ovaries; open triangles, control ovaries + 1 μM BAPTA-AM; closed triangles, control ovaries + 5 μM BAPTA-AM. (C) Squares, control ovaries; filled triangles, control ovaries + 20 μM DPI; open triangles, dNox RNAi ovaries.
dNox generates ROS in response to proctolin in a calcium-dependent manner
We investigated whether proctolin directly activates dNox in ovaries. Superoxide production was monitored using L-012, a luminescent, ROS detecting reagent (31). Proctolin stimulated ROS generation in isolated ovaries, and this was further increased by the calcium ionophore ionomycin, consistent with the expected regulation of dNox by calcium (Fig. 3A). Pre-treatment with the cell-permeant superoxide dismutase mimetic M40403 (41) (Fig. 3A) markedly inhibited the ROS signal, consistent with the reported specificity of L-012 for superoxide (31). Pre-treatment with the calcium chelator BAPTA-AM abrogated proctolin-induced ROS generation (Fig. 3B), demonstrating the requirement for calcium for ROS production. In dNox-RNAi ovaries and ovaries pre-treated with the NADPH oxidase inhibitor diphenylene iodonium (DPI), proctolin- and ionomycin-stimulated ROS production was undetectable (Fig. 3C), demonstrating that ROS generation requires dNox. Thus, proctolin stimulates dNox directly in Drosophila ovaries through a calcium-dependent mechanism.
dNox regulates proctolin-induced calcium fluxes in ovaries
It was previously known that proctolin-induced increases in cytosolic calcium concentration are required for increases in insect visceral muscle contractions (46, 47). To test if dNox regulates proctolin-induced changes in calcium levels in ovarian muscle, we used a transgenic fly strain that expresses the luminescent calcium sensor protein apo-aequorin under GAL4 control (38). When apo-aequorin expression was driven using Nrv1-GAL4 and holo-aequorin was reconstituted by incubating ovaries with the co-factor coelenterazine, proctolin stimulated a marked increase in calcium-dependent luminescence (Fig. 4A) in flies expressing normal levels of dNox. As a control, ionomycin added after the proctolin response markedly increased aequorin luminescence (Fig. 4A). When dNox dsRNA and aequorin were co-expressed, the proctolin-induced calcium flux was markedly inhibited by about 90%, while the total ionomycin-induced signals were similar (Fig. 4A). Similarly, the general Nox inhibitor diphenylene iodonium dose-dependently inhibited the proctolin-induced increase in calcium (Fig. 4B). Thus, dNox participates in proctolin-induced calcium fluxes.
Fig. 4.
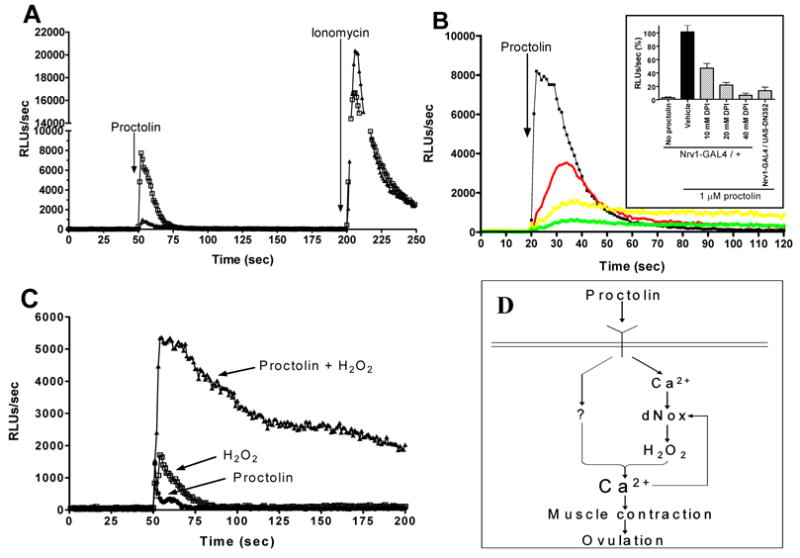
dNox regulates proctolin-induced calcium influx in ovarian muscle. (A–B) Aequorin luminescence after addition of 1 μM proctolin and 1 μM ionomycin. Control, UAS-DN352-IR; dNox RNAi, UAS-DN352 + TubP-GAL4. (A) Squares, control ovaries; triangles, dNox RNAi ovaries. (B) Black, control ovaries; red, control ovaries + 10 μM DPI; yellow, control ovaries + 20 μM DPI; green, control ovaries + 40 μM DPI. (B inset) Quantification of data from (A) and (B); n=5 for all samples and error bars represent standard deviation, n=5. (C) Circles, control ovaries + 1 μM proctolin; squares, control ovaries + 100 μM hydrogen peroxide; triangles, control ovaries + 1 μM proctolin + 100 μM hydrogen peroxide. (D) Schematic of mechanism for dNox regulation of Drosophila ovarian function.
H2O2 functions synergistically with dNox to stimulate a calcium flux in ovaries
There has been a growing appreciation in recent years of the potential for ROS, and hydrogen peroxide in particular, to function as intracellular second messengers (1, 36, 37). We hypothesized that dNox regulates intracellular calcium flux via hydrogen peroxide, which is formed by dismutation of the primary NADPH-oxidase product superoxide.
When H2O2 alone was added to aequorin-expressing ovaries from dNox-RNAi females, a small calcium flux was seen that was slightly greater than that seen with proctolin alone (Fig. 4C). However, when H2O2 and proctolin were added together, a strong and sustained increase in calcium was seen (Fig. 4C). The synergistic interaction of proctolin and H2O2 in the dNox-RNAi ovaries indicates that H2O2 is a co-stimulatory signal downstream of dNox that functions cooperatively with another proctolin receptor-initiated pathway (e.g., inositol triphosphate) to trigger an increase in intracellular calcium.
DISCUSSION
This study describes for the first time a role for dNox in egg-laying in Drosophila. This study demonstrates that dNox-derived ROS regulates proctolin-stimulated ovarian muscle contraction, and demonstrates that Nox-derived ROS participates as a co-signal for regulating a calcium flux in this tissue. Thus, both dNox-derived H2O2 and an additional signal from the proctolin receptor converge to activate an as-yet unidentified calcium channel, elevating cytosolic calcium and triggering muscle contraction (Fig. 4D). The calcium requirement for dNox activation itself, and the calcium regulating function of Nox-derived ROS suggests that in fly muscle dNox and calcium could comprise a positive feedback loop that regulates the level of cytosolic calcium achieved in proctolin-stimulated muscle. Thus, the activation and/or expression of Noxes may determine the extent to which a system is able to elevate calcium in response to an agonist, in effect acting as a rheostat to regulate the extent of the response.
While ROS have typically been thought to be widely and promiscuously reactive, there is now growing evidence that ROS transiently produced by NADPH oxidases actually participate in a fairly narrow range of reactions with biomolecules. One such reaction is the oxidation of low pKa thiols of cysteine residues. Interestingly, such low pKa thiols exist in key residues of many signaling proteins, such as the active site cysteines of protein tyrosine phosphatases (PTPs) (23) and the lipid phosphatase PTEN (19), as well as in transcription factors such as AP-1 and NF-κB (25). Oxidation of active site cysteines of PTPs and PTEN causes their inactivation, thus favoring the activation of signaling pathways by tyrosine kinases (40). In addition to their role in regulating tyrosine phosphorylation, ROS have also been implicated in plants in cellular calcium homeostasis. ROS production by plant NADPH oxidases occurs in response to a variety of physiologic stimuli including pathogens, hormones, polar growth and gravitropism (30). Recent genetic evidence indicates that plant Noxes and their derived ROS regulate cytosolic calcium as part of a mechanism to regulate root hair tip growth (9) and stomatal closure (18).
Increasingly, data from cell culture experiments suggest that ROS-regulated calcium signaling may also be relevant to the animal kingdom. For example, exogenously added hydrogen peroxide increases cellular calcium in pancreatic islet cells (45), cardiac muscle (44), smooth muscle (5, 44), and other mammalian cells (17). While studies in mammalian cell culture systems are intriguing, the use of supraphysiologic levels of ROS in many of these studies as well as the lack of genetic studies have prevented extrapolation to the physiological context, and these data have been interpreted mostly in the context of oxidant toxicity. Some recent data however in murine B-cells using RNAi demonstrated a role for Duox in calcium signaling in response to B-cell receptor activation, suggesting that Nox-ROS-calcium may be a physiologically relevant signaling cassette in some mammalian cells (43).
The biochemical mechanism by which dNox regulates proctolin-induced calcium flux remains an open question. Exogenously added ROS affect the function of variety of calcium channels and other proteins involved in calcium homeostasis, but due to the use of supraphysiologic levels of ROS, the physiologic relevance of these effects is unclear. Nevertheless, these experiments provide candidates for physiologically relevant, Nox-regulated systems. Among these are voltage-gated calcium channels, the IP3-receptor, ryanodine receptor and the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase (SERCA) (17). As with PTPs, ROS regulation of calcium channels seems to be mediated through oxidation of specific sulfhydryl residues. For instance, RyR1 channels have a few cysteine residues that are highly sensitive to oxidative modification at physiological pH, and oxidation of these residues in vitro enhances channel activity (11). The IP3R1 directly interacts with ERp44, a protein of the thioredoxin family that inhibits IP3R1 activity, and whose interaction with IP3R1 depends on the presence of free cysteine residues in IP3R1. IP3R1 and ERp44 fail to interact in their oxidized forms, suggesting that oxidation of key cysteine residues in IP3R1 leads to dissociation of ERp44 and channel activation (12).
Drosophila visceral muscles are closest in structure and function to smooth muscles of mammals and the results presented herein may have implications concerning roles for NADPH oxidases and their derived ROS in human vascular smooth muscle cells (VSMCs). NADPH oxidases have been extensively studied in VSMCs in the context of signaling by Ang II. Ang II acutely activates NADPH oxidase activity in VSMCs and Nox-derived ROS has been shown to regulate kinase signaling in these cells (10). Ang II also acutely activates calcium fluxes in VSMCs. While Ang II-stimulated calcium fluxes and Ang II-induced ROS production through NADPH oxidase are well-documented individually, the possible connections between these two pathways in this context have not been explored. Preliminary data from our lab indicates that Nox-derived ROS is indeed important for Ang II-induced calcium fluxes in hVSMCs (Meera Penumetcha and J.D.L.). Thus, Nox-ROS regulation of agonist-induced calcium flux may be a conserved mechanism operating in smooth muscles and may have important implications for vascular physiology and pathophysiology.
In summary, our results demonstrate a role for dNox in regulating agonist-induced calcium flux, ovarian muscle contraction and egg-laying. These data provide clear genetic evidence in an intact animal for a role of NADPH oxidases in cell signaling. Taken together with genetic data from plants and unpublished studies in VSMCs, these results indicate that a Nox-ROS-calcium signaling cassette is broadly conserved in nature, and may play an important general role in agonist-induced smooth muscle contractions.
Supplementary Material
Supplemental Fig. 1. dNox RNAi driven by Nrv1-GAL4 recapitulates the ovulation defect. (A–B) Light micrographs of ovaries from (A) Mated UAS-DN352-IR female. (B) Mated UAS-DN352-IR + Nrv1-GAL4 female.
Supplemental Fig. 2. Muscle structure is intact in dNox RNAi ovaries. (A–B) Epifluorescent photographs of FITC-phalloidin stained ovaries from (A) UAS-DN352-IR female and (B) UAS-DN352-IR + Nrv1-GAL4 female.
Supplemental Fig. 3. Proctolin receptor expression is conserved in dNox RNAi ovaries. (A–B) Epifluorescent photographs of ovaries labeled with proctolin receptor primary antibody and Alexa Fluor 555-conjugated secondary antibody. (A) ovaries from UAS-DN352-IR female and (B) ovaries from UAS-DN352-IR + Nrv1-GAL4 female.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Aslan M, Ozben T. Oxidants in receptor tyrosine kinase signal transduction pathways. Antioxid Redox Signal. 2003;5:781–8. doi: 10.1089/152308603770380089. [DOI] [PubMed] [Google Scholar]
- 2.Babior BM, Lambeth JD, Nauseef W. The neutrophil NADPH oxidase. Arch Biochem Biophys. 2002;397:342–4. doi: 10.1006/abbi.2001.2642. [DOI] [PubMed] [Google Scholar]
- 3.Banfi B, Molnar G, Maturana A, Steger K, Hegedus B, Demaurex N, Krause KH. A Ca(2+)-activated NADPH oxidase in testis, spleen, and lymph nodes. J Biol Chem. 2001;276:37594–601. doi: 10.1074/jbc.M103034200. [DOI] [PubMed] [Google Scholar]
- 4.Banfi B, Tirone F, Durussel I, Knisz J, Moskwa P, Molnar GZ, Krause KH, Cox JA. Mechanism of Ca2+ activation of the NADPH oxidase 5 (NOX5) J Biol Chem. 2004;279:18583–91. doi: 10.1074/jbc.M310268200. [DOI] [PubMed] [Google Scholar]
- 5.Bielefeldt K, Whiteis CA, Sharma RV, Abboud FM, Conklin JL. Reactive oxygen species and calcium homeostasis in cultured human intestinal smooth muscle cells. Am J Physiol. 1997;272:G1439–50. doi: 10.1152/ajpgi.1997.272.6.G1439. [DOI] [PubMed] [Google Scholar]
- 6.Bloch Qazi MC, Heifetz Y, Wolfner MF. The developments between gametogenesis and fertilization: ovulation and female sperm storage in Drosophila melanogaster. Dev Biol. 2003;256:195–211. doi: 10.1016/s0012-1606(02)00125-2. [DOI] [PubMed] [Google Scholar]
- 7.Cheng G, Ritsick D, Lambeth JD. Nox3 regulation by NOXO1, p47phox, and p67phox. J Biol Chem. 2004;279:34250–5. doi: 10.1074/jbc.M400660200. [DOI] [PubMed] [Google Scholar]
- 8.Edens WA, Sharling L, Cheng G, Shapira R, Kinkade JM, Lee T, Edens HA, Tang X, Sullards C, Flaherty DB, Benian GM, Lambeth JD. Tyrosine cross-linking of extracellular matrix is catalyzed by Duox, a multidomain oxidase/peroxidase with homology to the phagocyte oxidase subunit gp91phox. J Cell Biol. 2001;154:879–91. doi: 10.1083/jcb.200103132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Foreman J, Demidchik V, Bothwell JH, Mylona P, Miedema H, Torres MA, Linstead P, Costa S, Brownlee C, Jones JD, Davies JM, Dolan L. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature. 2003;422:442–6. doi: 10.1038/nature01485. [DOI] [PubMed] [Google Scholar]
- 10.Hanna IR, Taniyama Y, Szocs K, Rocic P, Griendling KK. NAD(P)H oxidase-derived reactive oxygen species as mediators of angiotensin II signaling. Antioxid Redox Signal. 2002;4:899–914. doi: 10.1089/152308602762197443. [DOI] [PubMed] [Google Scholar]
- 11.Hidalgo C, Sanchez G, Barrientos G, Aracena-Parks P. A transverse tubule NOX activity stimulates calcium release from isolated triads via RYR1 S-Glutathionylation. J Biol Chem. 2006 doi: 10.1074/jbc.M600451200. [DOI] [PubMed] [Google Scholar]
- 12.Higo T, Hattori M, Nakamura T, Natsume T, Michikawa T, Mikoshiba K. Subtype-specific and ER lumenal environment-dependent regulation of inositol 1,4,5-trisphosphate receptor type 1 by ERp44. Cell. 2005;120:85–98. doi: 10.1016/j.cell.2004.11.048. [DOI] [PubMed] [Google Scholar]
- 13.Hrdlicka L, Gibson M, Kiger A, Micchelli C, Schober M, Schock F, Perrimon N. Analysis of twenty-four Gal4 lines in Drosophila melanogaster. Genesis. 2002;34:51–7. doi: 10.1002/gene.10125. [DOI] [PubMed] [Google Scholar]
- 14.Johnson EC, Garczynski SF, Park D, Crim JW, Nassel DR, Taghert PH. Identification and characterization of a G protein-coupled receptor for the neuropeptide proctolin in Drosophilamelanogaster. Proc Natl Acad Sci U S A. 2003;100:6198–203. doi: 10.1073/pnas.1030108100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kawahara T, Ritsick D, Cheng G, Lambeth JD. Point mutations in the proline-rich region of p22phox are dominant inhibitors of Nox1- and Nox2-dependent reactive oxygen generation. J Biol Chem. 2005;280:31859–69. doi: 10.1074/jbc.M501882200. [DOI] [PubMed] [Google Scholar]
- 16.Kiss T, Varanka I, Benedeczky I. Neuromuscular transmission in the visceral muscle of locust oviduct. Neuroscience. 1984;12:309–22. doi: 10.1016/0306-4522(84)90156-8. [DOI] [PubMed] [Google Scholar]
- 17.Kourie JI. Interaction of reactive oxygen species with ion transport mechanisms. Am J Physiol. 1998;275:C1–24. doi: 10.1152/ajpcell.1998.275.1.C1. [DOI] [PubMed] [Google Scholar]
- 18.Kwak JM, I, Mori C, Pei ZM, Leonhardt N, Torres MA, Dangl JL, Bloom RE, Bodde S, Jones JD, Schroeder JI. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. Embo J. 2003;22:2623–33. doi: 10.1093/emboj/cdg277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kwon J, Lee SR, Yang KS, Ahn Y, Kim YJ, Stadtman ER, Rhee SG. Reversible oxidation and inactivation of the tumor suppressor PTEN in cells stimulated with peptide growth factors. Proc Natl Acad Sci U S A. 2004;101:16419–24. doi: 10.1073/pnas.0407396101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lam G, Thummel CS. Inducible expression of double-stranded RNA directs specific genetic interference in Drosophila. Curr Biol. 2000;10:957–63. doi: 10.1016/s0960-9822(00)00631-x. [DOI] [PubMed] [Google Scholar]
- 21.Lambeth JD. NOX enzymes and the biology of reactive oxygen. Nat Rev Immunol. 2004;4:181–9. doi: 10.1038/nri1312. [DOI] [PubMed] [Google Scholar]
- 22.Lee HG, Seong CS, Kim YC, Davis RL, Han KA. ctopamine receptor OAMB is required for ovulation in Drosophila melanogaster. Dev Biol. 2003;264:179–90. doi: 10.1016/j.ydbio.2003.07.018. [DOI] [PubMed] [Google Scholar]
- 23.Lee SR, Kwon KS, Kim SR, Rhee SG. Reversible inactivation of protein-tyrosine phosphatase 1B in A431 cells stimulated with epidermal growth factor. J Biol Chem. 1998;273:15366–72. doi: 10.1074/jbc.273.25.15366. [DOI] [PubMed] [Google Scholar]
- 24.Leulier F, Vidal S, Saigo K, Ueda R, Lemaitre B. Inducible expression of double-stranded RNA reveals a role for dFADD in the regulation of the antibacterial response in Drosophila adults. Curr Biol. 2002;12:996–1000. doi: 10.1016/s0960-9822(02)00873-4. [DOI] [PubMed] [Google Scholar]
- 25.Liu H, Colavitti Rovira R, II, Finkel T. Redox-dependent transcriptional regulation. Circ Res. 2005;97:967–74. doi: 10.1161/01.RES.0000188210.72062.10. [DOI] [PubMed] [Google Scholar]
- 26.Mahadev K, Motoshima H, Wu X, Ruddy JM, Arnold RS, Cheng G, Lambeth JD, Goldstein BJ. The NAD(P)H oxidase homolog Nox4 modulates insulin-stimulated generation of H2O2 and plays an integral role in insulin signal transduction. Mol Cell Biol. 2004;24:1844–54. doi: 10.1128/MCB.24.5.1844-1854.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Manseau L, Baradaran A, Brower D, Budhu A, Elefant F, Phan H, Philp AV, Yang M, Glover D, Kaiser K, Palter K, Selleck S. GAL4 enhancer traps expressed in the embryo, larval brain, imaginal discs, and ovary of Drosophila. Dev Dyn. 1997;209:310–22. doi: 10.1002/(SICI)1097-0177(199707)209:3<310::AID-AJA6>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 28.Matsuno K, Yamada H, Iwata K, Jin D, Katsuyama M, Matsuki M, Takai S, Yamanishi K, Miyazaki M, Matsubara H, Yabe-Nishimura C. Nox1 is involved in angiotensin II-mediated hypertension: a study in Nox1-deficient mice. Circulation. 2005;112:2677–85. doi: 10.1161/CIRCULATIONAHA.105.573709. [DOI] [PubMed] [Google Scholar]
- 29.Monastirioti M. Distinct octopamine cell population residing in the CNS abdominal ganglion controls ovulation in Drosophila melanogaster. Dev Biol. 2003;264:38–49. doi: 10.1016/j.ydbio.2003.07.019. [DOI] [PubMed] [Google Scholar]
- 30.Mori IC, Schroeder JI. Reactive oxygen species activation of plant Ca2+ channels. A signaling mechanism in polar growth, hormone transduction, stress signaling, and hypothetically mechanotransduction. Plant Physiol. 2004;135:702–8. doi: 10.1104/pp.104.042069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nishinaka Y, Aramaki Y, Yoshida H, Masuya H, Sugawara T, Ichimori Y. A new sensitive chemiluminescence probe, L-012, for measuring the production of superoxide anion by cells. Biochem Biophys Res Commun. 1993;193:554–9. doi: 10.1006/bbrc.1993.1659. [DOI] [PubMed] [Google Scholar]
- 32.Nykamp DA, Lange AB. Interaction between octopamine and proctolin on the oviducts of Locusta migratoria. J Insect Physiol. 2000;46:809–816. doi: 10.1016/s0022-1910(99)00170-5. [DOI] [PubMed] [Google Scholar]
- 33.Osborne RH. Insect neurotransmission: neurotransmitters and their receptors. Pharmacol Ther. 1996;69:117–42. doi: 10.1016/0163-7258(95)02054-3. [DOI] [PubMed] [Google Scholar]
- 34.Phelps CB, Brand AH. Ectopic gene expression in Drosophila using GAL4 system. Methods (Duluth) 1998;14:367–79. doi: 10.1006/meth.1998.0592. [DOI] [PubMed] [Google Scholar]
- 35.Piccin A, Salameh A, Benna C, Sandrelli F, Mazzotta G, Zordan M, Rosato E, Kyriacou CP, Costa R. Efficient and heritable functional knock-out of an adult phenotype in Drosophila using a GAL4-driven hairpin RNA incorporating a heterologous spacer. Nucleic Acids Res. 2001;29:E55–5. doi: 10.1093/nar/29.12.e55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Reth M. Hydrogen peroxide as second messenger in lymphocyte activation. Nat Immunol. 2002;3:1129–34. doi: 10.1038/ni1202-1129. [DOI] [PubMed] [Google Scholar]
- 37.Rhee SG, Kang SW, Jeong W, Chang TS, Yang KS, Woo HA. Intracellular messenger function of hydrogen peroxide and its regulation by peroxiredoxins. Curr Opin Cell Biol. 2005;17:183–9. doi: 10.1016/j.ceb.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 38.Rosay P, Davies SA, Yu Y, Sozen A, Kaiser K, Dow JA. Cell-type specific calcium signalling in a Drosophila epithelium. J Cell Sci. 1997;110(Pt 15):1683–92. doi: 10.1242/jcs.110.15.1683. [DOI] [PubMed] [Google Scholar]
- 39.Rubin GM, Spradling AC. Genetic transformation of Drosophila with transposable element vectors. Science. 1982;218:348–53. doi: 10.1126/science.6289436. [DOI] [PubMed] [Google Scholar]
- 40.Salmeen A, Barford D. Functions and mechanisms of redox regulation of cysteine-based phosphatases. Antioxid Redox Signal. 2005;7:560–77. doi: 10.1089/ars.2005.7.560. [DOI] [PubMed] [Google Scholar]
- 41.Salvemini D, Wang ZQ, Zweier JL, Samouilov A, Macarthur H, Misko TP, Currie MG, Cuzzocrea S, Sikorski JA, Riley DP. A nonpeptidyl mimic of superoxide dismutase with therapeutic activity in rats. Science. 1999;286:304–6. doi: 10.1126/science.286.5438.304. [DOI] [PubMed] [Google Scholar]
- 42.Segal AW, Shatwell KP. The NADPH oxidase of phagocytic leukocytes. Ann N Y Acad Sci. 1997;832:215–22. doi: 10.1111/j.1749-6632.1997.tb46249.x. [DOI] [PubMed] [Google Scholar]
- 43.Singh DK, Kumar D, Siddiqui Z, Basu SK, Kumar V, Rao KV. The strength of receptor signaling is centrally controlled through a cooperative loop between Ca2+ and an oxidant signal. Cell. 2005;121:281–93. doi: 10.1016/j.cell.2005.02.036. [DOI] [PubMed] [Google Scholar]
- 44.Suzuki YJ, Ford GD. Redox regulation of signal transduction in cardiac and smooth muscle. J Mol Cell Cardiol. 1999;31:345–53. doi: 10.1006/jmcc.1998.0872. [DOI] [PubMed] [Google Scholar]
- 45.Tang J, Zhang JH. Mechanisms of [Ca2+]i elevation by H2O2 in islets of rats. Life Sci. 2000;68:475–81. doi: 10.1016/s0024-3205(00)00944-9. [DOI] [PubMed] [Google Scholar]
- 46.Wegener C, Nassel DR. Peptide-induced Ca(2+) movements in a tonic insect muscle: effects of proctolin and periviscerokinin-2. J Neurophysiol. 2000;84:3056–66. doi: 10.1152/jn.2000.84.6.3056. [DOI] [PubMed] [Google Scholar]
- 47.Wilcox CL, Lange AB. Role of extracellular and intracellular calcium on proctolin-induced contractions in an insect visceral muscle. Regul Pept. 1995;56:49–59. doi: 10.1016/0167-0115(95)00006-w. [DOI] [PubMed] [Google Scholar]
- 48.Xu P, Sun B, Salvaterra PM. Organization and transcriptional regulation of Drosophila Na(+), K(+)-ATPase beta subunit genes: Nrv1 and Nrv2. Gene. 1999;236:303–13. doi: 10.1016/s0378-1119(99)00269-3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Fig. 1. dNox RNAi driven by Nrv1-GAL4 recapitulates the ovulation defect. (A–B) Light micrographs of ovaries from (A) Mated UAS-DN352-IR female. (B) Mated UAS-DN352-IR + Nrv1-GAL4 female.
Supplemental Fig. 2. Muscle structure is intact in dNox RNAi ovaries. (A–B) Epifluorescent photographs of FITC-phalloidin stained ovaries from (A) UAS-DN352-IR female and (B) UAS-DN352-IR + Nrv1-GAL4 female.
Supplemental Fig. 3. Proctolin receptor expression is conserved in dNox RNAi ovaries. (A–B) Epifluorescent photographs of ovaries labeled with proctolin receptor primary antibody and Alexa Fluor 555-conjugated secondary antibody. (A) ovaries from UAS-DN352-IR female and (B) ovaries from UAS-DN352-IR + Nrv1-GAL4 female.