

Abstract
Populations that live in environments with different habitats have to distribute their offspring over these habitats. When population densities go to equilibrium, the evolutionary optimum is an ideal free distribution. Under an ideal free distribution, no offspring should be put into sink habitats. However, when the environmental conditions in a habitat are not constant but fluctuate, allocating offspring to sink habitats can increase the long term growth rate of a population. We demonstrate this principle in a simple model for offspring allocation. As a consequence, it is possible that populations persist in environments that only consist of sink habitats.
In ecological systems, some habitats are better than others. It is believed that habitats can differ to the extent that some local populations act as sinks and only exist because of the spill over from other, source, populations. Sink habitats, by definition, are habitats in which populations cannot survive when they are isolated from other populations (1).
When habitat quality differs, the way in which individuals distribute their offspring over the different habitats affects the total number of surviving offspring produced. Individuals who distribute their offspring differently have different fitnesses. A genetically determined distribution of offspring is therefore under evolutionary control. When the environment consists of habitats of different but constant quality, the evolutionarily optimal solution is, simply, to put all offspring in the best habitat. The problem becomes more complicated when the quality of the habitat depends on the local population density (the quality of a habitat is the probability that an individual born in that habitat gives rise to an adult). If every individual allocated its offspring to the habitat that at that moment is the best, this habitat would soon degrade to be the worst.
If population densities settle at an equilibrium value, the solution to this evolutionary problem is to distribute offspring such that the quality of all habitats becomes equal. When the quality of habitats differ, any individual that allocates fewer offspring to the worst habitat and instead allocates them to the best habitat will have a higher than average reproductive success. Eventually, evolution thus leads to a distribution of offspring for which, when the whole population adopts it, all habitats are of equal quality and for which an individual cannot improve its reproductive success by using a different distribution. This evolutionary stable state is known as the ideal free distribution (2, 3).
When the population is at equilibrium, every individual gives rise to exactly one adult individual in the next generation. When an individual has m children, the survival probability of each child, averaged over all habitats, is 1/m. When the distribution of offspring is ideal free, the probability of a child to survive to an adult is the same in all habitats and therefore it is 1/m in each and every habitat. But when a habitat is the only one available and every individual is forced to allocate all its m children to that one habitat, at equilibrium density, the survival probability of offspring is also 1/m. Therefore, under an ideal free distribution, local population densities are equal to what they would be when all habitats were isolated. The fraction of offspring allocated to a particular habitat, under the evolutionary stable distribution, is proportional to the equilibrium density if that habitat were isolated (4, 5). As a consequence, no offspring should be allocated to a sink habitat because (by definition) the equilibrium density of a sink population is zero (6, 5).
The existence of sink populations in nature is well documented (7, 8). This leaves open the question, “Why do sink populations exist at all?” A possible answer is that constraints of various type keep populations from evolving towards the ideal free distribution (1, 9, 10). Recently, it has been shown that when population densities fluctuate, the evolutionary process may lead to distributions in which offspring are allocated to sink populations (10). The reason for this apparently counter-intuitive result is that when the quality of the source population occasionally becomes worse than the sink population, the geometric mean of the reproductive success of an individual that does allocate to the sink can exceed that of an individual that only uses the source habitat (11) (Fig. 1).
Figure 1.

Long term population growth rates in a system in which female butterflies allocate 100 eggs between farmed and wild habitat. The farmed habitat is mostly high in quality, but in 1 out of 10 generations, all larvae are killed by insecticide spraying or early harvesting. The wild habitat is of constant but low quality. The optimal allocation is 80 eggs in the farmed habitat and 20 in the wild habitat. (After ref. 11).
We will take this idea further in a simple model in which offspring have to be allocated to two sink habitats. The population growth rate depends on the way in which offspring are distributed. Moreover, the optimal distribution can result in a growing population despite the fact that both habitats are, on their own, sinks.
A Model for the Allocation of Offspring
Suppose that an individual distributes its offspring over two types of habitat with different qualities. The probability of offspring survival differs between the two habitats. For simplicity, we will assume that this probability is independent of the local population size. One habitat, say habitat 1, is highly productive but suffers from occasional catastrophes in which only very few offspring survive. We assume that a population confined to this habitat cannot persist in the long run. The other habitat is constant in quality but the quality is low so that a population that uses this habitat only is doomed to extinction.
An individual produces in total m offspring, of which a fraction f is deposited in habitat 1 and the remainder in habitat 2. Let N(t) be the size of the population in generation t and Si(t) the survivorship of offspring in habitat i for generation t. The population size in the next generation is:
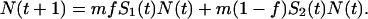 |
1 |
The variable S1(t) is a stochastic variable which takes value Sa < 1/m with probability p, and value Sb > 1/m with probability 1 − p; p is the probability of a catastrophe to occur. We will choose habitat 2 to be constant and low in quality: S2(t) = Sc < 1/m (i.e., it is a sink). The parameters Sa, Sb, and Sc are all constants. Assuming that fraction f is genetically determined, we analyze the optimal genotype f*, which maximizes the expected long term population growth rate. The long term average growth rate of the population is given by the geometric mean of the yearly rates of increase:
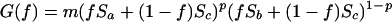 |
2 |
If this quantity exceeds unity the population persists. Maximizing G(f) is identical to maximizing its logarithm, and therefore the optimal fraction f* can be calculated from:
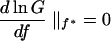 |
which gives:
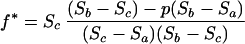 |
3 |
This is a unique maximum because
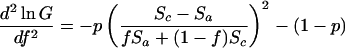 |
4 |
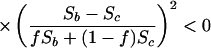 |
The long term growth rate of a population confined to habitat 1 is given by the geometric mean of its growth rate, which is G(1) = mSapSb1−p, whereas for a population confined to habitat 2 the long term growth rate is G(0) = mSc. We will choose the parameters such that G(0), G(1) < 1, thus guaranteeing that the population cannot survive in a single habitat. Hence, both habitats are sinks by definition.
The population will increase in number in an environment of combined sinks when G(f*) > 1. Because the closed form expression for this quantity is not very transparent, we will present our results graphically. In Figure 2a, the geometric growth rate is depicted for a sink–sink population as a function of f. Clearly, it is possible to distribute the offspring among the two sinks in such a way that the population increases. When catastrophes become more likely, the optimal fraction f* and the optimal growth rate G(f*) decrease (Fig. 2b).
Figure 2.

(a) The long term growth rate G(f) of a population described by equation (1) as a function of f, the fraction of offspring put in habitat 1. The growth rate has an optimum for f = f*. Note: when all offspring are put in a single habitat, the long term growth is less than unity, but by distributing offspring over both habitats, the population can grow. Parameters: mSa = 0.005, mSb = 5, mSc = 0.7, p = 1/3. (b) The maximum of growth rate, G(f*), and the fraction of offspring in habitat 1 for which it is attained, f*, as a function of the probability of having a catastrophe, p. An increase in risk causes the maximum growth rate to decrease whereas more offspring are put in the constant low quality patch. Parameters used are as in a.
Strictly speaking, a population that persists should not only grow away from very small numbers but also be bounded away from infinitely large numbers (12). This requires density dependence. Fig. 3 shows a density-dependent analog of our offspring allocation model. The number of individuals that survives in a habitat depends on the number of offspring present in the habitat through the Ricker model. The growth rates of the model can vary; habitat 1 has a variable growth rate, whereas habitat 2 has a constant low growth rate. The figure shows the average population density as a function of f. The population can persist only when offspring are suitably distributed over the two habitats.
Figure 3.

For the density-dependent analog of the model described by Eq. 1, we replace the survival probabilities Si(t) by the Ricker functions: S1(t) = r1(t)me−fmN(t) with r1 = Sa with probability p and r1 = Sb with probability 1 − p, and S2(t) = Scme−(1−f)mN(t). In the figure, the population densities are shown as averaged over time. The population becomes extinct when all offspring are allocated to only one habitat, but when offspring are distributed over the two habitats, population persistence becomes feasible. Parameters used are as in Fig. 2, m = 100.
Discussion
In stable environments, populations can maximize growth rates by not allocating offspring to sink habitats. However, when the quality of habitats fluctuates over time, a population that puts a fraction of its offspring into a sink habitat can have a higher growth rate than one that only uses sources. When the main habitat of a species is struck occasionally by catastrophes so that the population could not survive when confined to this habitat (i.e., it is a sink), populations can persist by spreading their risk and distributing their offspring over different types of habitat. When the quality of the other habitats is higher than that of the source during the catastrophe, population growth rate is increased by spreading offspring over the habitats. Surprisingly, these habitats also may be sinks. In other words, although this population cannot survive in a single habitat, it may persist in an environment consisting only of sinks.
Although data on the temporal variability in reproduction success are scarce, catastrophic extinctions of source populations have been reported (15). A further empirical illustration of our theoretical example is described in (16): Cakile edula is an annual plant found in a range of coastal habitats stretching from open beaches to vegetated dunes. In vegetated areas, the plants occur commonly although the number of fruits produced per fruit sown is less than one; hence, these areas are sinks. On open beaches, the number of fruits per fruit greatly exceeds one. The dunes are a relatively constant environment but reproductive success is low probably because of competition with other plant species. In contrast, the open beaches are a very productive habitat but reproductive success can be catastrophically reduced by storms and high sea events that occur with a high spatial correlation. In the long term, Cakile edula might therefore not be able to survive when confined to either dunes or open beaches; nevertheless, it is a common species on the sandy sea sides of eastern North America.
The mechanism through which such populations persist is the spreading of risk (13). In its original form, the theory of risk spreading was a verbal argument explaining the distribution of offspring over similar but uncorrelated environments (13). This situation is encountered in classic metapopulation models (14) in which the geometric mean growth rate of a population confined to a single patch is zero, due to occasional catastrophes. The geometric mean growth rate of a population that distributes its offspring over many patches depends on the correlation between the patches and the costs of distributing offspring. Whether or not the metapopulation persists depends on the balance between these two. When the patches are sufficiently productive and uncorrelated a population will persist through the spreading of risk. However, when catastrophes are correlated spatially, risk spreading over similar habitats has little effect. Then, spreading of risk between different types of habitat will be more effective. Here, we provide a quantitative description of this idea that enables us to demonstrate that populations can persist through risk spreading even if some of the habitats are sink habitats of constant but low quality. The same result can be achieved by allocating of offspring to life stages in which the reproductive success is low but constant as is the case with, for instance, diapause, dormancy, and seed banks.
The use of source–sink theory is gaining attention in conservation biology (17). The underlying assumption is that environments are stable and densities converge to an equilibrium, so that sinks play no role in the survival of the species. Natural environments can be highly variable. As we have shown, species can depend on sink habitats for their survival. When local environments are variable, predictions from standard source–sink theory may not hold and species can survive in sinks only.
Acknowledgments
We thank John Lawton and Bob Holt for comments on the manuscript.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Pulliam H R. Am Nat. 1988;132:652–661. [Google Scholar]
- 2.Morishita M. Contrib Physiol Ecol Kyoto. 1950;65:1–149. [Google Scholar]
- 3.Fretwell S D, Lucas H L. Acta Biotheor. 1969;19:16–36. [Google Scholar]
- 4.Holt R D. Am Nat. 1984;124:377–406. doi: 10.1086/284280. [DOI] [PubMed] [Google Scholar]
- 5.Holt R D. Theor Popul Biol. 1985;28:181–208. [Google Scholar]
- 6.Fretwell S D. Populations in Seasonal Environments. Princeton: Princeton Univ. Press; 1972. [Google Scholar]
- 7.Dias P C. Trends Ecol Evol. 1996;11:326–330. doi: 10.1016/0169-5347(96)10037-9. [DOI] [PubMed] [Google Scholar]
- 8.Pulliam H R. In: Population Dynamics in Ecological Space and Time. Rhodes O E, Chesser R K, Smith H D, editors. Univ. of Chicago Press; 1996. pp. 45–70. [Google Scholar]
- 9.Van Baalen M, Sabelis M W. Am Nat. 1993;142:646–670. doi: 10.1086/285562. [DOI] [PubMed] [Google Scholar]
- 10.Holt R D. Evol Ecol. 1997;11:723–731. [Google Scholar]
- 11.Yoshimura J, Jansen V A A. Res Pop Ecol. 1996;38:165–182. [Google Scholar]
- 12.Hofbauer J, Sigmund K. The Theory of Evolution and Dynamical Systems. Cambridge: Cambridge Univ. Press; 1988. [Google Scholar]
- 13.Den Boer P J. Acta Biotheor. 1968;18:165–194. doi: 10.1007/BF01556726. [DOI] [PubMed] [Google Scholar]
- 14.Levins R. Bull Entomol Soc Am. 1969;15:237–240. [Google Scholar]
- 15.Thomas C D, Singer M C, Boughton D A. Am Nat. 1996;148:957–975. [Google Scholar]
- 16.Keddy P A. J Ecol. 1981;69:615–630. [Google Scholar]
- 17.Conroy M J, Noon B R. Ecol Appl. 1996;6:763–773. [Google Scholar]