

Abstract
We previously reported a role for the IZH2 gene product in metal ion metabolism. Subsequently, Izh2p was also identified as a member of the PAQR family of receptors and, more specifically, as the receptor for the plant protein osmotin. In this report, we investigate the effect of Izh2p on iron homeostasis. We show that overproduction of Izh2p prevents the iron-dependent induction of the Fet3p component of the high-affinity iron-uptake system and is deleterious for growth in iron-limited medium. We demonstrate that the effect of Izh2p requires cAMP-dependent kinase and AMP-dependent kinase and is not mediated by general inhibition of the Aft1p iron-responsive transcriptional activator. We also show that Izh2p-overproduction negatively regulates Nrg1p/Nrg2p- and Msn2p/Msn4p-dependent reporters. Furthermore, we show that the Nrg1p/Nrg2p and Msn2p/Msn4p pairs are epistatic to each other with respect to their effects on FET3 expression. Finally, we show that the mechanism by which PAQR receptors activate signal transduction pathways is likely to be conserved from yeast to humans.
Keywords: FET3, IZH2, Osmotin, PAQR, adiponectin
Introduction
Iron and zinc are cofactors for hundreds of proteins making them essential for viability. When grown in metal-limiting environments, microorganisms must increase their ability to scavenge these metals to maintain maximal growth rates. As a result, intricate regulatory systems have evolved to up-regulate metal ion acquisition in response to nutritional depletion [1]. In Saccharomyces cerevisiae, iron- and zinc-acquisition are controlled by Aft1p and Zap1p, respectively. These transcriptional activators sense the nutritional status of their respective micronutrients and, in response to deficiency, induce genes involved in uptake. However, some studies indicate that iron- and zinc-uptake systems are inducible under metal replete conditions when Aft1p and Zap1p are believed to be silent. For example, the expression of genes involved in high-affinity iron- and zinc-uptake fluctuate with the phase of the cell cycle [2] and seem to be regulated by carbon starvation [3]. These studies suggest that iron and zinc bioavailability are not the only environmental stimuli that affect metal accumulation and that Aft1p and Zap1p are not the only regulators of iron and zinc uptake. Therefore, identification of novel genes involved in the regulation of iron and zinc homeostasis is of critical importance for a proper understanding of metal metabolism.
The catalyst for the studies reported herein was data we previously published implicating the integral membrane protein, Izh2p, in the metabolism of zinc in S. cerevisiae [4]. Specifically, we showed that IZH2 expression is induced by both zinc-deficiency via the Zap1p zinc-sensor and zinc-toxicity via the Mga2p hypoxia-sensing transcription factor. More importantly, however, we showed that overexpression of Izh2p negatively regulates the expression of a Zap1p-dependent reporter construct. While exploring the effect of Izh2p on gene transcription, we discovered that increased dosage of Izh2p also represses the expression of FET3, a gene whose product collaborates with Ftr1p to form an oxidase-permease complex responsible for high-affinity iron-uptake [5]. While both FET3 and FTR1 are induced during iron-deprivation by Aft1p, the effect of Izh2p on FET3 expression is independent of Aft1p. These findings support a functional link between Izh2p, a zinc-regulated gene, and high-affinity iron-uptake, indicating cross-talk between the iron- and zinc-dependent regulons in yeast.
We present evidence suggesting that Izh2p exerts its effects on FET3 by regulating the activities of four transcription factors with previously unrecognized roles in iron homeostasis - Msn2p, Msn4p, Nrg1p and Nrg2p. Msn2p and Msn4p are stress-responsive transcriptional activators [6] while Nrg1p and Nrg2p are carbon source-dependent transcriptional repressors [7]. We demonstrate that the Msn2p/Msn4p activators and the Nrg1p/Nrg2p repressors are epistatic with respect to the expression of FET3. Furthermore, we demonstrate that Izh2p dependent-repression requires both cAMP-dependent protein kinase (protein kinase A, PKA) and AMP-dependent protein kinase (AMPK). Since PKA inhibits Msn2p/Msn4p [6] and AMPK inhibits Nrg1p/Nrg2p [8], our findings fit a model in which Izh2p regulates FET3 expression via negative regulation of AMPK, positive regulation PKA or both. It remains to be seen how Izh2p affects PKA and AMPK and whether these transcription factors affect FET3 expression through binding to cis-regulatory elements.
The physiological importance of FET3 regulation by Izh2p is still a mystery. However, Izh2p was recently identified as a cell surface receptor for the plant protein osmotin [9]. While the exact function of osmotin remains a matter of debate, the fact that it is induced by plants as part of the innate immune response and possesses potent antifungal activity suggests that it functions as a primary line of defense against fungal pathogens [10]. Therefore, from a pharmacological standpoint, there is significant interest in understanding how osmotin affects fungal physiology. Since yeast with defects in high-affinity iron-uptake show decreased virulence [11, 12], the regulation of high affinity iron-uptake represents a reasonable mechanism by which osmotin, via Izh2p, may exert an antifungal effect.
The importance of this work is not limited to gaining a better understanding of metal metabolism in yeast. Izh2p belongs to a newly discovered family of receptors known as PAQRs (Progesterone and AdipoQ Receptors) that are ubiquitous in eukaryotes [13]. The first members of this family to be characterized as receptors were the membrane progesterone receptors from seatrout [14] and the adiponectin (AdipoQ) receptors from human [15]. Adiponectin is an insulin-sensitizing hormone and the human adiponectin receptors are believed to play an important role in the etiology of type II diabetes. Despite the medical importance of adiponectin receptors, little is known about how they convert extracellular signals into physiological changes inside cells. Indeed, all that seems to be known is that they somehow transmit signals to AMPK [16]. In this report, we show heterologous expression of two human adiponectin receptors in yeast. Not only do these receptors repress FET3 in response to adiponectin, this effect requires the same signaling proteins as Izh2p overexpression, including AMPK. The functional expression of these receptors in yeast demonstrates mechanistic conservation in the PAQR receptor family and establishes a valuable model system for the investigation of a pair of receptors that are critical for human health.
Materials and Methods
Yeast Strains
Genotypes for strains used in this paper are listed in Supplemental Table 1. MCY5326 wild type, MCY5338 (msn2Δmsn4Δ), MCY5378 (nrg1Δnrg2Δ) and MCY5385 (msn2Δmsn4Δnrg1Δnrg2) were generously provided by Dr. Marian Carlson at Columbia University [17]. The pFLO11-lacZ strain, in which the lacZ ORF has replaced the FLO11 ORF in the genome of Σ1278b, was generously provided by Dr. Florian Bauer at the University of Stellenbosch, Matieland, South Africa. [18] All other yeast strains used in this study were purchased from Euroscarf (http://web.uni-frankfurt.de/fb15/mikro/euroscarf/) and are in the BY4742 (MATα his3 leu2 ura3 lys2) background.
lacZ Reporters and Plasmids
pFET3-397 and pFET3-297 are episomal reporter plasmids (Provided by Dr. Andrew Dancis at the University of Pennsylvania) in which lacZ is driven by different truncations of the FET3 promoter (−397 to +3 and −297 to +3, respectively) [19]. Several different reporters were obtained that have the lacZ gene driven by a minimal CYC1 promoter in which the native upstream activating sequence has been replaced with fragments from various other promoters. pCYC1-FeRE (pFL-W, provided by Dr. Andrew Dancis) contains a fragment of the FET3 promoter that includes the Aft1-binding site known as an iron-response element or FeRE [19]. pCYC1-STRE (pCZ-oligo31/32, provided by Dr. Janet Trager, UCLA) contains a fragment of the DDR2 promoter that includes tandem stress response elements (STRE) to which the Msn2p and Msn4p transcription factors bind [20].
IZH1-4 were cloned into pRS316-GAL1 via gap repair as previously described [4]. The triple hemaglutinin epitope (3xHA) tagged construct of the IZH2 gene was generated by replacing the ZRC1 promoter and open reading frame in the YCpZRC1-HA plasmid [21] with those of IZH2. This was accomplished by gap repair of Age I-digested YCpZRC1-HA to generate pIZH2-3xHA. This construct has the IZH2 gene driven by its native promoter and retains the ZRC1 terminator sequence. The native IZH2 promoter was then exchanged with the GAL1 promoter using gap repair of pIZH2-3xHA plasmids cut with EcoN I. AdipoR1 and AdipoR2 were amplified from cDNAs obtained from OpenBiosystems (Accession #: BC010743 and BC051858, respectively). PCR products were cloned into NcoI/BamHI digested pYES260 vector [22] by gap repair allowing for GAL1-driven expression. Primer sequences are reported in Supplemental Table 2. Plasmids containing GAL1-driven the TAP (Tandem Affinity Purification) tagged NRG1 and NRG2 constructs were purchased from OpenBiosystems.
Biochemical Assays
Most experiments were performed in a defined EDTA-containing medium known as low iron medium (LIM) [23]. Iron-deficiency and repletion were generated by adding either 1 μM or 1 mM of FeCl3, respectively, to LIM. 2% galactose was used to fully induce GAL1-driven gene overexpression. Total sugar concentration was maintained at 2% by the addition of raffinose in those experiments in which the % galactose was modulated to decrease expression levels. Cells were allowed to grow to mid-log phase in LIM and β-galactosidase assays on permeabilized yeast were performed as previously described [24]. Ferroxidase activity assays on intact yeast cells followed published procedures using ferrozine as a colorimetric indicator of iron oxidation state [25]. All experiments include triplicate data points and each experiment was performed a minimum of three times. Representative experiments are shown. All error bars represent ± 1 standard deviation within a single experiment. Because of the simplicity of β-galactosidase assays and the fact that Izh2p overexpression similarly affects both lacZ and ferroxidase activity, lacZ assays are shown for most experiments. For some strains with significant growth defects, such as snf1Δ, lacZ assays were not performed because of the low efficiency of co-transformation of lacZ reporter and overexpression plasmids. For these strains, only ferroxidase activity is shown. Iron-limited agar plates were made by adding 20 μM FeCl3 to plates containing synthetic media and 1 mM of the iron-specific chelator, ferrozine. Western blots were performed using standard protocol and commercially available anti-HA antibodies.
Results
Effect of Izh2p dosage on FET3 expression
The pFET3-397 lacZ reporter responds reciprocally to the amount of iron added to LIM. (Figure 1A) The induction during iron-deficiency, which is due to the presence of the FeRE from −250 to −244 to which the Aft1p protein must bind for transcriptional activation [19], is repressible by IZH2 gene overexpression. (Figure 1A) We demonstrate that iron-deficiency and IZH2 gene dosage have the same effect on cell surface ferroxidase activity - the physiological function of Fet3p - as they do on the β-galactosidase activity of pFET3-397. (Figure 1A) pFET3-397 repression is proportional to the amount of galactose used to induce Izh2p overexpression. (Figure 1B) Wild type cells carrying an empty overexpression vector are capable of growing on agar plates made iron-limited by the addition of ferrozine, while those carrying an Izh2p overexpression plasmid are not. (Figure 1C)
Figure 1. Izh2p represses FET3 transcription and Fet3p activity.
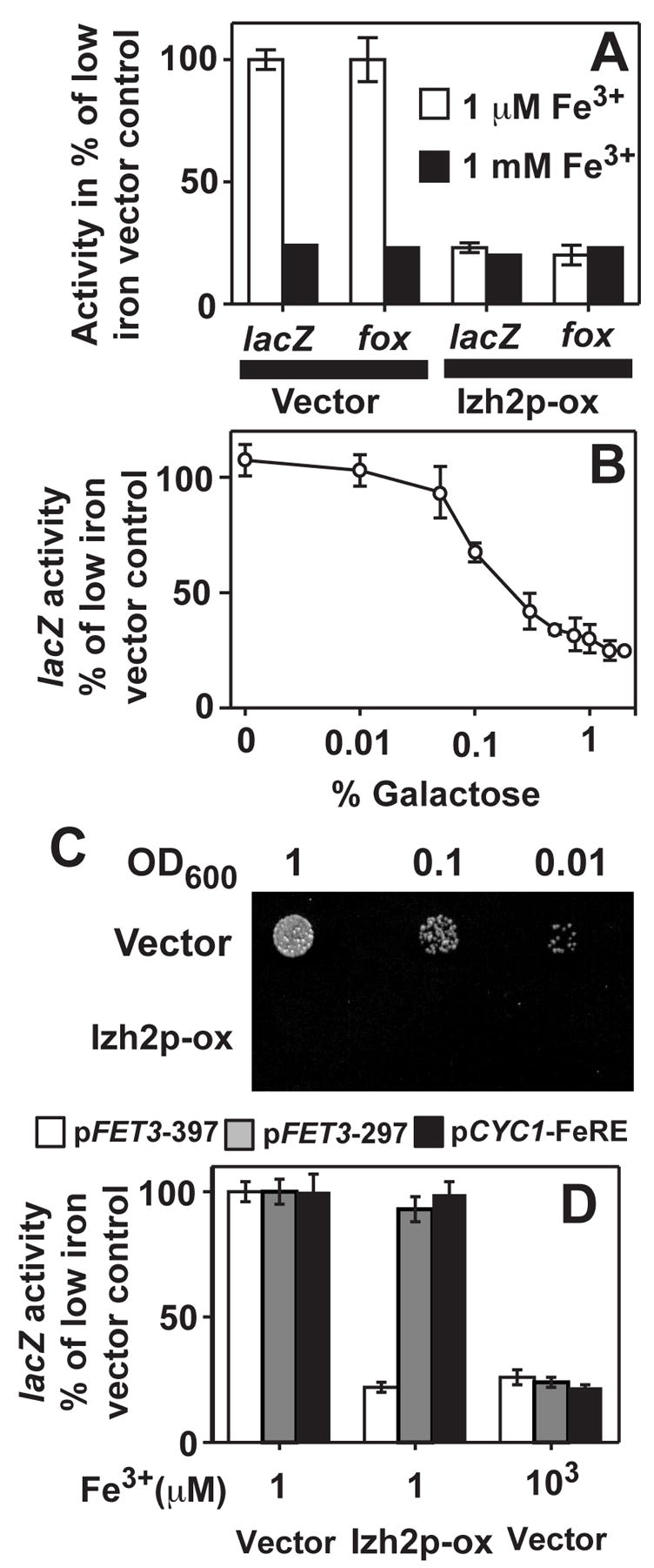
BY4742 wild type strain is used in all panels. Activities in panels A and D are shown as a percentage of fully induced activity in a strain carrying empty overexpression vector grown in iron-deficient LIM. (A) β-galactosidase activity (lacZ) of the pFET3-397 reporter and cell surface ferroxidase activity (fox) are induced by low iron in wild type yeast carrying empty overexpression vector, but not in yeast carrying a plasmid that overexpresses Izh2p. (B) The effect of Izh2p on pFET3-397 decreases as the % galactose in iron-deficient LIM is decreased. Activities for each galactose concentration are normalized to % of activity in a strain carrying empty overexpression vector grown at the same galactose concentration. (C) Overexpression of Izh2p in a wild type strain causes a growth defect in plates containing synthetic media supplemented with 1 mM ferrozine and 20 μM Fe3+. (D) lacZ activity is induced by growth in iron-deficient LIM carrying pFET3-397, pFET3-297 or pCYC1-FeRE. Overexpression of Izh2p in low iron represses only the pFET3-397 construct.
To ensure that Izh2p is functionally expressed under the conditions of our experiment, we tagged the IZH2 gene with a 3x-HA epitope at the C-terminus. This construct functionally complements the phenotypes of an izh2Δ strain [4] and is fully capable of repressing pFET3-397. (Supplemental Figure 1A) Furthermore, we demonstrate by Western blot that the tagged construct is, indeed, galactose inducible and expressed in iron-deficient LIM. (Supplemental Figures 1B and 1C)
Izh2p does not globally affect the activity of Aft1p and specifically affects a 100 bp region of the FET3 promoter
Like pFET3-397, the pFET3-297 and pCYC1-FeRE lacZ reporters are inducible by growth in iron-deficient LIM due to the presence of the FeRE from the FET3 promoter. (Figure 1D) While pFET3-397 responds to Izh2p overexpression, pFET3-297 and pCYC1-FeRE do not. Furthermore, the Aft1p-dependent induction of pFET3-397 or ferroxidase activity in iron-deficient LIM is unaffected in all the mutant strains we tested with the exception of snf1Δ, msn2Δ, msn4Δ, msn2Δmsn4Δ, gal83Δ and sak1Δ. (Supplemental Figure 2)
Msn2p and Msn4p positively regulate FET3 expression
FET3 is significantly induced during the diauxic shift in iron-replete medium [3]. Since Msn2p and Msn4p are known to activate gene transcription during this growth phase, we examined their role in FET3 regulation. In msn2Δ and msn4Δ strains, ferroxidase activity is constitutively repressed in iron-deficient LIM. (Figure 2A) Ferroxidase activity is even more repressed in an msn2Δ msn4Δ double mutant relative to the proper isogenic wild type control. (Figure 2B) The pCYC1-STRE reporter contains tandem STREs and is conditionally activated by Msn2p/Msn4p [20]. In keeping with a role for Msn2p/Msn4p in gene activation during carbon depletion, the activity pCYC1-STRE decreases as cells exit stationary phase and enter log phase (Figure 2C). Izh2p overexpression significantly repressed pCYC1-STRE at low OD600 when Msn2p/Msn4p are active but not at higher OD600.
Figure 2. Msn2p, Msn4p and PKA regulate FET3.

The legend above panel A applies to panels A–D. Activities in panels A–E are shown as a percentage of fully induced activity in the isogenic wild type strain carrying empty overexpression vector grown in iron-deficient LIM. (A and B) Cell surface ferroxidase activity (fox) is constitutively repressed in msn2Δ and msn4Δ strains when compared to the isogenic BY4742 wild type and in the msn2Δmsn4Δ mutant when compared to the isogenic MCY5326 wild type. Overexpression of Izh2p in any of the msn mutant strains had no effect on fox activity. (C) β-galactosidase activity (lacZ) in BY4742 carrying pCYC1-STRE reporter is repressed when Izh2p is overproduced in early log phase (OD600 = 0.04) but not in mid-log phase (OD600 = 1.60). (D) pFET3-397 repression by Izh2p overexpression seen in the BY4742 wild type strain is alleviated in ras2Δ, tpk1Δ, tpk2Δ and tpk3Δ strains. (E) Insensitivity of pFET3-397 to Izh2p overexpression in the ras2Δ strain can be overcome by cAMP addition to the growth medium prior to assay. (1) BY4742 WT strain carrying pFET3-397, empty overexpression vector and treated with 3 mM cAMP (2) ras2Δ carrying pFET3-397, empty overexpression vector and treated with 3 mM cAMP (3) ras2Δ carrying pFET3-397, Izh2p overexpression vector and treated with 3 mM cAMP.
Involvement of Ras-cAMP and PKA in IZH2-dependent FET3 repression
Since PKA negatively regulates the activity of Msn2p/Msn4p and has been implicated in the repression of Aft1-target genes [26], we examined the role of PKA in Izh2p-dependent signal transduction. Deletion of any of the three genes encoding catalytic isoforms of PKA (TPK1, TPK2 and TPK3) results in a complete loss of pFET3-397 repression due to Izh2p overexpression in iron-deficient LIM. (Figure 2D) PKA is positively controlled by input from the Ras2p-cAMP pathway [27]. Deletion of RAS2 results in impaired Izh2p-dependent pFET3-397 repression (Figure 2D), however, Izh2p overexpression still represses pFET3-397 in the ras2Δ mutant if cAMP is added to the culture medium. (Figure 2E)
Nrg1p and Nrg2p negatively regulate FET3 expression
Data from Candida albicans suggests that iron-uptake genes may be under the control of the CaNrg1 transcriptional repressor [28]. Thus, we examined the role of the S. cerevisiae CaNrg1p homologues, Nrg1p and Nrg2p, in FET3 expression. Izh2p-dependent pFET3-397 repression is lost in nrg1Δ or nrg2Δ strains. (Figure 3A) The effect of Izh2p on ferroxidase activity is also lost in an nrg1Δ nrg2Δ double mutant strain. (Figure 3B) Nrg1p/Nrg2p negatively regulate the pFLO11-lacZ reporter [18]. Izh2p represses the activity of pFLO11-lacZ in iron-deficient LIM. (Figure 3C) Furthermore, we obtained GAL1-driven TAP-tagged Nrg1p or Nrg2p constructs and found that Nrg2p-TAP overexpression has the same effect on pFET3-397 as does Izh2p overexpression. (Figure 3D) Nrg1p-TAP overexpression has no effect, however, the Nrg1p-TAP construct does not restore the effect of Izh2p on pFET3-397 in the nrg1Δ strain (data not shown) suggesting that Nrg1p-TAP may be nonfunctional or not expressed. Nrg2p-TAP overexpression is not capable of repressing ferroxidase activity in an nrg1Δ nrg2Δ strain. (Figure 3E)
Figure 3. Nrg1p, Nrg2p, Msn2p and Msn4p in Izh2p-dependent FET3 repression.

For panels A, B, C and F, white bars show strains carrying empty expression vector and black bars show strains carrying Izh2p overexpression vector. For panels D and E, white bars show strains carrying empty expression vector and grey bars show strains carrying Nrg2p-TAP overexpression vector. For all panels except C, activities are shown as a percentage of fully induced activity in the isogenic wild type strain grown in iron-deficient LIM. (A and B) The repression of pFET3-397 lacZ activity by Izh2p overexpression in the BY4742 wild type strain is absent in the nrg1Δ and nrg2Δ mutant strains. Similarly the repression of activity seen in MCY5326 is not seen in the nrg1Δnrg2Δ strain. (C) Overexpression of Izh2p represses a FLO11-lacZ construct in the Σ1278b wild type strain. lacZ activities are shown in Miller Units (D and E) Overexpression of TAP-tagged Nrg2p resulted in repression of pFET3-397 in BY4742 wild type. Similarly Nrg2p-TAP repressed ferroxidase activity in MCY5326 wild type but not in the isogenic nrg1Δnrg2Δ strain. (F) The constitutive repression of ferroxidase activity in low iron medium seen in an msn2Δmsn4Δ strain relative to the MCY5326 isogenic wild type strain can be partially alleviated by concomitant deletion of nrg1Δ and nrg2Δ to generate the quadruple mutant. Izh2p overexpression has no effect on ferroxidase activity in strains lacking Nrg1p/Nrg2p.
The negative effect of Msn2p/Msn4p deletion on ferroxidase activity is alleviated if Nrg1p and Nrg2p are concomitantly knocked-out. (Figure 3F) In addition, overexpression of Izh2p has no effect on ferroxidase activity in the quadruple mutant strain.
AMP-dependent kinase and the repression of FET3
AMP-dependent kinase (AMPK) is a known negative regulator of Nrg1p/Nrg2p [8] and has been implicated in the regulation of iron-regulated genes [3]. AMPK has multiple isoforms depending on subunit composition. There is a lone catalytic α-subunit of AMPK (Snf1p) which forms a heterotrimeric complex with a lone stimulatory γ-subunit (Snf4p) and one of three β-subunits (Gal83p, Sip1p or Sip2p) that regulate AMPK cellular localization [29]. Its activity is regulated by AMP and by phosphorylation by an upstream activating kinase (AMPKK) of which there are three partially redundant isoforms (Elm1p, Sak1p or Tos3p) [30]. Complete loss of AMPK activity caused by SNF1 deletion results in constitutive repression of the FET3 gene as measured by basal ferroxidase activity in iron-deficient LIM. (Figure 4A) Selective inactivation of nuclear AMPK activity by deletion of the β subunit (Gal83p) that targets AMPK to the nucleus or the AMPK activating kinase (AMPKK, Sak1p) responsible for activation of nuclear AMPK also results in constitutive pFET3-397repression. (Figure 4B) Finally, strains lacking Snf4p also show an approximately 40% decrease in pFET3-397 activity. (Figure 4C)
Figure 4. The role of AMPK in FET3 repression.
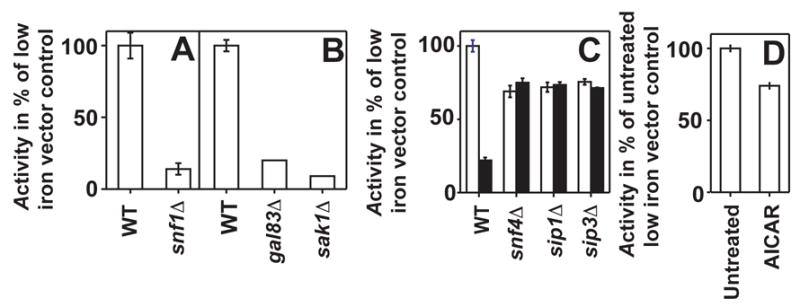
For all panels, β-galactosidase or ferroxidase activities are shown as a percentage of activity produced in the isogenic wild type strain carrying an empty overexpression vector and grown in iron-deficient LIM. (A) In a strain lacking Snf1p, cell surface ferroxidase activity is repressed relative to the isogenic BY4742 wild type. (B) In gal83Δ and sak1Δ strains, pFET3-397 is similarly constitutively repressed. (C) In strains lacking various AMPK interacting proteins, there is a slight, but significant reduction in pFET3-397. In addition, Izh2p overexpression (black bars) in these strains has no effect on lacZ activity when compared to activity in strains carrying empty overexpression vector (white bars). (D) Addition of 500 μM AICAR to the growth medium results in a small but reproducible repression of pFET3-397.
Perturbations of a cytoplasmic isoform of AMPK by deletion of Sip1p results in a decrease in pFET3-397 activity similar to that seen in the snf4Δ mutant. (Figure 4C) Deletion of another cytoplasmic Snf1p-interacting protein, Sip3p, has an identical effect on pFET3-397. (Figure 4C) In the snf4Δ, sip1Δ and sip3Δ strains, Izh2p overexpression no longer represses pFET3-397. The addition of 500 μM AICAR, an activator of mammalian AMPK [31], to the growth medium resulted in a small but reproducible decrease in pFET3-397 activity, suggesting that AMPK activation may play a role in signal transduction. (Figure 4D)
PAQR overexpression mimics receptor activation
In this paper, we show that overexpression of three homologous yeast PAQR receptors (Izh1p, Izh3p and Izh4p) results in pFET3-397 repression and decreased ferroxidase activity in iron-deficient LIM. (Figure 5A) We also demonstrate that overexpression of the human adiponectin receptor, AdipoR1, but not its functional homologue, AdipoR2, represses pFET3-397 and ferroxidase activity. (Figure 5A) By decreasing the amount of galactose, the effect of AdipoR1 on pFET3-397 can be incrementally diminished. (Figure 5B) At first glance it appears as though AdipoR2 does not function in yeast. When adiponectin is added to cells expressing either AdipoR2 (in 2% galactose) or low levels of AdipoR1 (in 0.02% galactose), pFET3-397 is repressed in a dose-dependent manner. (Figure 5C) As in the case of Izh2p overexpression, pFET3-397 repression by either overexpression of AdipoR1 in the absence of adiponectin or AdipoR2 in the presence of adiponectin required RAS2, TPK1, TPK2, TPK3, SIP1, SNF4, SIP3, NRG1 and NRG2. (Figure 5F)
Figure 5. Functional expression of human PAQR receptors in yeast.

For panels A and D, β-galactosidase (lacZ) or ferroxidase (fox) activities are shown as a percentage of activity produced in the BY4742 wild type strain carrying an empty overexpression vector and grown in iron-deficient LIM. (A) Overexpression of three homologous yeast receptors (Izh1p, Izh3p and Izh4p) and one homologous human receptor (AdipoR1) has similar effects on pFET3-397 and ferroxidase activity. Overexpression of a second human homologue (AdipoR2) has no effect. (B) Decreased expression of AdipoR1 by lowering the % galactose diminishes its effect on pFET3-397. Activities for each galactose concentration are normalized to % of activity in a strain carrying empty overexpression vector grown at the same galactose concentration. (C) Expression of AdipoR2 in 2% galactose fully has no effect on pFET3-397 unless adiponectin is added to the medium (black squares). Adiponectin has no effect on pFET3-397 in a strain carrying empty vector control (white squares). Expression of AdipoR1 in 1.98% raffinose/0.02% galactose has no effect on pFET3-397 unless adiponectin is added (black circles) while adiponectin alone has no effect (white circles). For cells grown with 0.02% galactose, activities are normalized to activity in a strain carrying the empty overexpression vector and grown in iron-deficient LIM with the same concentration of galactose. (D) Dose dependent pFET3-397 repression by either AdipoR1 overexpression in the absence of adiponectin or AdipoR2 activation in the presence of adiponectin is alleviated in strains lacking RAS2, TPK1, TPK2, TPK3, SIP1, SNF4, SIP3, NRG1 or NRG2.
Discussion
Repression of FET3 by Izh2p overexpression
In this study, we provide genetic evidence that supports a new mechanism for the regulation of the FET3 gene encoding a ferroxidase involved in high-affinity iron-uptake. The first clues to this regulatory mechanism came from a previous study in which we showed Izh2p-dependent repression of a zinc-dependent reporter [4]. This effect was not due to a generalized growth defect, aberrant expression of any random membrane protein or a global defect in transcription, translation, or β-galactosidase activity. While investigating the specificity of Izh2p-dependent repression, we discovered - and herein report - the finding that Izh2p also represses the iron-dependent induction of the FET3 gene. More importantly, Izh2p causes a decrease in Fet3p enzymatic activity and cell viability in iron-limited medium, thus confirming that the Izh2p effect is neither an artifact of our reporter constructs nor a physiologically irrelevant phenomenon. The level of repression of both FET3 expression and ferroxidase activity by Izh2p are comparable to repression caused by full iron-repletion (approximately 25% of fully induced levels), suggesting that the loss of ferroxidase activity is mainly caused by decreased gene expression. A generic iron-dependent reporter was unaffected by Izh2p overproduction, indicating that Izh2p modulates FET3 activity in an iron-independent manner. Due to the importance of iron acquisition in host environments, the negative effect of Izh2p on iron-uptake represents a viable mechanism by which osmotin, the proposed ligand for Izh2p, might exert its fungicidal effects.
Msn2p/Msn4p, PKA and Ras-cAMP in the negative regulation of FET3
We envision two probable mechanisms by which Izh2p and its homologous receptors may function to repress FET3. In the first scenario, the signaling pathway inactivates an essential co-activator of FET3 expression. We demonstrate that - at least under the conditions of our experiments - the Msn2p and Msn4p stress-responsive transcriptional activators are indeed essential for FET3 induction and that Izh2p may affect FET3 by negatively regulating Msn2p/Msn4p. This possibility is bolstered by several lines of evidence. First, Msn2p/Msn4p are the main transcription factors responsible for inducing genes during the diauxic shift - a growth phase during which FET3 is known to be induced [3]. Second, Izh2p-dependent repression requires PKA, a kinase known to negatively regulate Msn2p/Msn4p [32] and purported to regulate a subset of Aft1p-target genes [26]. Third, PKA activity is positively regulated by cAMP produced by the Ras-cAMP module [32], which is required for Izh2p-dependent repression. Fourth, Izh2p overexpression represses a generic stress responsive reporter that is directly activated by Msn2p/Msn4p. Thus, Izh2p negatively affects the activity of Msn2p/Msn4p and, in turn, these are essential co-activators of FET3.
In this model, Izh2p, through Msn2p/Msn4p, directly or indirectly affect FET3 transcription by altering flux through the Ras-cAMP/PKA pathway. The fact that osmotin signaling in yeast [9, 33] and FET3 repression via Izh2p both require the Ras2p G-protein has led to the supposition that Izh2p is coupled with Ras2p. Since the PAQR family of proteins has been postulated to comprise a novel class of G-protein coupled receptor (GPCR) [34, 35], the involvement of Ras-cAMP/PKA would seem to support the conclusion that Izh2p is a Ras2p-coupled GPCR. However, if Izh2p were a Ras2p-coupled GPCR, then the physical presence of the Ras2p polypeptide would be essential for Izh2p-dependent signaling. We have demonstrated that it is not Ras2p, but cAMP that is essential, thus demonstrating that Izh2p is not directly coupled to Ras2p. Our data suggest that the Ras-cAMP module is only required by virtue of the essentiality of cAMP in PKA activation and that it is likely that signals from Izh2p converge on PKA downstream of Ras-cAMP.
Nrg1p/Nrg2p and AMPK in the negative regulation of FET3
A second, non-mutually-exclusive, possibility for the mechanism of Izh2p-dependent repression is the activation of a repressor of FET3 transcription. An exhaustive review of the literature led us to a paper showing increased expression of genes involved in high-affinity iron-uptake in C. albicans strains lacking the CaNrg1p repressor [28]. CaNrg1p has two partially redundant homologues in S. cerevisiae named Nrg1p and Nrg2p. We demonstrate that without these repressors, Izh2p was incapable of repressing FET3. We also show that overexpression of Izh2p represses a generic reporter known to be directly regulated by Nrg1p/Nrg2p. Furthermore, we show that Nrg2p overexpression was sufficient to recapitulate the effect of Izh2p overproduction. Repression by Nrg1p/Nrg2p is believed to be mediated by their binding to a CCCTC motif in the promoters of target genes [36]. A CCCTC motif can be found from −316 to −312 in the FET3 promoter as well as in the zinc-dependent reporter described in the beginning of the discussion. The fact that pFET3-397 construct is Izh2p-regulated, while pFET3-297 is not, strongly suggests that there is an element - possibly the CCCTC motif at −312 - directly responsible for repression, however, we cannot yet state unequivocally that Nrg1p/Nrg2p bind the FET3 promoter at a cis-regulatory element.
Nrg1p and Nrg2p physically interact with and are negatively regulated by AMPK [37], thus it is possible that Izh2p activates Nrg1p/Nrg2p by inactivation of AMPK. A possible role for AMPK in Izh2p-dependent signaling is very attractive since human adiponectin receptors are known to function through AMPK [16] and yeast AMPK has been shown to positively regulate genes involved in iron homeostasis [3]. We present two pieces of evidence that are consistent with an important role for nuclear AMPK in FET3 regulation by inactivation of nuclear Nrg1p/Nrg2p. First, strains completely lacking AMPK or those lacking the nuclear isoform of AMPK show constitutively repressed ferroxidase activity. Second, strains lacking a variety of AMPK subunits show a 40% reduction in FET3 expression. Evidence for an essential role for cytoplasmic AMPK in Izh2p-dependent repression comes from data showing FET3 repression by the addition of an AMPK activator (AICAR) and loss of Izh2p-dependent repression in strains lacking cytoplasmic isoforms of AMPK. Finally, the finding that the Snf1p-interacting protein, Sip3p [38], is required for Izh2p-dependent repression is particularly exciting considering that the human homologue of Sip3p, APPL1, physically interacts with and is required for signal transduction via the human adiponectin receptor, AdipoR1 [39].
Epistasis of Msn2p/Msn4p and Nrg1p/Nrg2p
Nrg1p/Nrg2p and Msn2p/Msn4p have opposing effects on FET3 expression. Nrg1p and Nrg2p are predicted to bind the CCCTC motif as well as the CCCCT STRE that functions as a binding site for Msn2p and Msn4p [36]. Due to the similarity of the STRE and CCCTC motifs to which Nrg1p/Nrg2p bind, it is possible that both the Nrg1p/Nrg2p repressors and Msn2p/Msn4p activators recognize the same elements. Indeed, a recent study suggests that Nrg1p/Nrg2p and Msn2p/Msn4p compete with each other for binding to the same regulatory elements in a subset of stress-responsive promoters [17]. Our data shows that Nrg1p/Nrg2p are epistatic to Msn2p/Msn4p with respect to FET3 expression, however, we cannot yet conclude if these transcription factors act on FET3 through cis-regulatory elements. Based on our data, we propose that Msn2p/Msn4p function competitively and antagonistically to Nrg1p/Nrg2p in the direct or indirect regulation of FET3 and that Izh2p overexpression may alter this competition by inactivating Msn2p/Msn4p via PKA or activating Nrg1p/Nrg2p via AMPK.
Izh2p overexpression vs. receptor activation
A final point to address is that the Izh2p receptor causes FET3 repression in the absence of its supposed activating ligand osmotin. It is possible that Izh2p is actually a receptor for an, as yet unidentified, endogenous molecule and that this hypothetical ligand is present in high enough concentration to activate Izh2p when it is overexpressed. It is also possible that Izh2p, like many signaling proteins, has an intrinsic basal signaling capability that is amplified by overexpression making the presence of activating ligand unnecessary. In the latter case, overexpression would function equivalently to activation. Due to the fact that osmotin is neither commercially available nor easily produced, we cannot yet directly test the latter model. However, we have functionally expressed two homologous human adiponectin receptors in yeast. In both cases, activation of the receptor with adiponectin represses FET3 expression in a manner that is nearly identical to that of Izh2p overexpression.
Conclusions
Our findings support several important conclusions. First, Izh2p affects iron homeostasis via the Nrg1p/Nrg2p and Msn2p/Msn4p transcription factors. Second, the regulation of Izh2p by zinc suggests cross-talk between the iron- and zinc-dependent regulons, thus adding to the growing body of data indicating a complex relationship between iron and zinc homeostasis in yeast [40–43]. Third, every PAQR protein tested, regardless of activating ligand and physiological function, activate a similar intracellular signaling cascade, suggesting a conserved mechanism of signal transduction. Specifically, these studies shed light on the physiology of Izh2p and its relation to iron metabolism in yeast. More generally, they provide a simple assay to expedite investigations into the structure/function relationship in human PAQR proteins of biomedical interest.
Supplementary Material
Supplemental Figure 1: Functional expression of Izh2p-3HA. (A) Izh2p or Izh2p-3HA overexpression represses the induction of pFET3-397 in low iron. (B) Western blot showing that the Izh2p-3HA tagged protein is expressed in cells grown in galactose but not in cells grown in glucose. (C) Western blot showing that the Izh2p-3HA tagged protein is induced in cells carrying the Izh2p-3HA overexpression construct but not in those carrying the untagged Izh2p construct. Protein samples in B and C were heated to 37º C rather than boiled prior to loading.
Supplemental Figure 2: Aft1p responds normally to iron-deficiency in most mutant strains. (A) Ferroxidase activity is induced by iron-deficiency in the BY4742 wild type strain. Fet3p activity is not induced in isogenic strains lacking SNF1, MSN2 or MSN4. (B) Ferroxidase activity is inducible by iron-deficiency in the MCY5326 wild type strain, the isogenic nrg1Δnrg2Δ double mutant and the isogenic nrg1Δnrg2Δmsn2Δ msn4Δ quadruple mutant but not in the isogenic msn2Δmsn4Δ double mutant. (C) b-galactosidase activity of the pFET3-397 construct is inducible by iron-deficiency in the BY4742 wild type strain and in all isogenic mutants tested except for the gal83Δ, sak1Δ and msn4Δ mutants. For all panels, activities are shown as a % of isogenic wild type control grown in low iron media. The legend above panel A applies to panels A–C.
Acknowledgments
We would like to thank Drs. Marian Carlson, Andrew Dancis, Florian Bauer and Janet Treger for generously providing strains and plasmids. This research was funded by the University of Florida, Department of Chemistry and by NIH-NIDDK grant to TJL 1R21DK074812-01.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Rutherford JC, Bird AJ. Metal-responsive transcription factors that regulate iron, zinc, and copper homeostasis in eukaryotic cells. Eukaryot Cell. 2004;3:1–13. doi: 10.1128/EC.3.1.1-13.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cho RJ, Campbell MJ, Winzeler EA, Steinmetz L, Conway A, Wodicka L, Wolfsberg TG, Gabrielian AE, Landsman D, Lockhart DJ, Davis RW. A genome-wide transcriptional analysis of the mitotic cell cycle. Mol Cell. 1998;2:65–73. doi: 10.1016/s1097-2765(00)80114-8. [DOI] [PubMed] [Google Scholar]
- 3.Haurie V, Boucherie H, Sagliocco F. The Snf1 protein kinase controls the induction of genes of the iron uptake pathway at the diauxic shift in Saccharomyces cerevisiae. J Biol Chem. 2003;278:45391–6. doi: 10.1074/jbc.M307447200. [DOI] [PubMed] [Google Scholar]
- 4.Lyons TJ, Villa NY, Regalla LM, Kupchak BR, Vagstad A, Eide DJ. Metalloregulation of yeast membrane steroid receptor homologs. Proc Natl Acad Sci USA. 2004;101:5506–11. doi: 10.1073/pnas.0306324101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Singh A, Severance S, Kaur N, Wiltsie W, Kosman DJ. Assembly, activation, and trafficking of the Fet3p.Ftr1p high affinity iron permease complex in Saccharomyces cerevisiae. J Biol Chem. 2006;281:13355–64. doi: 10.1074/jbc.M512042200. [DOI] [PubMed] [Google Scholar]
- 6.Smith A, Ward MP, Garrett S. Yeast PKA represses Msn2p/Msn4p-dependent gene expression to regulate growth, stress response and glycogen accumulation. EMBO J. 1998;17:3556–64. doi: 10.1093/emboj/17.13.3556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Berkey CD, Vyas VK, Carlson M. Nrg1 and nrg2 transcriptional repressors are differently regulated in response to carbon source. Eukaryot Cell. 2004;3:311–7. doi: 10.1128/EC.3.2.311-317.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kuchin S, Vyas VK, Carlson M. Snf1 protein kinase and the repressors Nrg1 and Nrg2 regulate FLO11, haploid invasive growth, and diploid pseudohyphal differentiation. Mol Cell Biol. 2002;22:3994–4000. doi: 10.1128/MCB.22.12.3994-4000.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Narasimhan ML, Coca MA, Jin J, Yamauchi T, Ito Y, Kadowaki T, Kim KK, Pardo JM, Damsz B, Hasegawa PM, Yun DJ, Bressan RA. Osmotin is a homolog of mammalian adiponectin and controls apoptosis in yeast through a homolog of mammalian adiponectin receptor. Mol Cell. 2005;17:171–80. doi: 10.1016/j.molcel.2004.11.050. [DOI] [PubMed] [Google Scholar]
- 10.Linthorst JJM. Pathogenesis-related proteins of plants. Crit Rev Plant Sci. 1993;10:123–150. [Google Scholar]
- 11.Eck R, Hundt S, Hartl A, Roemer E, Kunkel W. A multicopper oxidase gene from Candida albicans: cloning, characterization and disruption. Microbiology. 1999;145(Pt 9):2415–22. doi: 10.1099/00221287-145-9-2415. [DOI] [PubMed] [Google Scholar]
- 12.Ramanan N, Wang Y. A high-affinity iron permease essential for Candida albicans virulence. Science. 2000;288:1062–4. doi: 10.1126/science.288.5468.1062. [DOI] [PubMed] [Google Scholar]
- 13.Tang YT, Hu T, Arterburn M, Boyle B, Bright JM, Emtage PC, Funk WD. PAQR proteins: a novel membrane receptor family defined by an ancient 7-transmembrane pass motif. J Mol Evol. 2005;61:372–80. doi: 10.1007/s00239-004-0375-2. [DOI] [PubMed] [Google Scholar]
- 14.Zhu Y, Rice CD, Pang Y, Pace M, Thomas P. Cloning, expression, and characterization of a membrane progestin receptor and evidence it is an intermediary in meiotic maturation of fish oocytes. Proc Natl Acad Sci USA. 2003;100:2231–6. doi: 10.1073/pnas.0336132100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, Sugiyama T, Miyagishi M, Hara K, Tsunoda M, Murakami K, Ohteki T, Uchida S, Takekawa S, Waki H, Tsuno NH, Shibata Y, Terauchi Y, Froguel P, Tobe K, Koyasu S, Taira K, Kitamura T, Shimizu T, Nagai R, Kadowaki T. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature. 2003;423:762–9. doi: 10.1038/nature01705. [DOI] [PubMed] [Google Scholar]
- 16.Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Invest. 2006;116:1784–92. doi: 10.1172/JCI29126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vyas VK, Berkey CD, Miyao T, Carlson M. Repressors Nrg1 and Nrg2 regulate a set of stress-responsive genes in Saccharomyces cerevisiae. Eukaryot Cell. 2005;4:1882–91. doi: 10.1128/EC.4.11.1882-1891.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.van Dyk D, Pretorius IS, Bauer FF. Mss11p is a central element of the regulatory network that controls FLO11 expression and invasive growth in Saccharomyces cerevisiae. Genetics. 2005;169:91–106. doi: 10.1534/genetics.104.033704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yamaguchi-Iwai Y, Stearman R, Dancis A, Klausner RD. Iron-regulated DNA binding by the AFT1 protein controls the iron regulon in yeast. EMBO J. 1996;15:3377–84. [PMC free article] [PubMed] [Google Scholar]
- 20.Treger JM, Magee TR, McEntee K. Functional analysis of the stress response element and its role in the multistress response of Saccharomyces cerevisiae. Biochem Biophys Res Commun. 1998;243:13–9. doi: 10.1006/bbrc.1997.8061. [DOI] [PubMed] [Google Scholar]
- 21.MacDiarmid CW, Milanick MA, Eide DJ. Biochemical properties of vacuolar zinc transport systems of Saccharomyces cerevisiae. J Biol Chem. 2002;277:39187–94. doi: 10.1074/jbc.M205052200. [DOI] [PubMed] [Google Scholar]
- 22.Melcher K. A modular set of prokaryotic and eukaryotic expression vectors. Anal Biochem. 2000;277:109–20. doi: 10.1006/abio.1999.4383. [DOI] [PubMed] [Google Scholar]
- 23.Eide D, Guarente L. Increased dosage of a transcriptional activator gene enhances iron-limited growth of Saccharomyces cerevisiae. J Gen Microbiol. 1992;138:347–54. doi: 10.1099/00221287-138-2-347. [DOI] [PubMed] [Google Scholar]
- 24.Lyons TJ, Gasch AP, Gaither LA, Botstein D, Brown PO, Eide DJ. Genome-wide characterization of the Zap1p zinc-responsive regulon in yeast. Proc Natl Acad Sci USA. 2000;97:7957–62. doi: 10.1073/pnas.97.14.7957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.De Silva DM, Askwith CC, Eide D, Kaplan J. The FET3 gene product required for high affinity iron transport in yeast is a cell surface ferroxidase. J Biol Chem. 1995;270:1098–101. doi: 10.1074/jbc.270.3.1098. [DOI] [PubMed] [Google Scholar]
- 26.Robertson LS, Causton HC, Young RA, Fink GR. The yeast A kinases differentially regulate iron uptake and respiratory function. Proc Natl Acad Sci USA. 2000;97:5984–8. doi: 10.1073/pnas.100113397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jiang Y, Davis C, Broach JR. Efficient transition to growth on fermentable carbon sources in Saccharomyces cerevisiae requires signaling through the Ras pathway. EMBO J. 1998;17:6942–51. doi: 10.1093/emboj/17.23.6942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Murad AM, d’Enfert C, Gaillardin C, Tournu H, Tekaia F, Talibi D, Marechal D, Marchais V, Cottin J, Brown AJ. Transcript profiling in Candida albicans reveals new cellular functions for the transcriptional repressors CaTup1, CaMig1 and CaNrg1. Mol Microbiol. 2001;42:981–93. doi: 10.1046/j.1365-2958.2001.02713.x. [DOI] [PubMed] [Google Scholar]
- 29.Vincent O, Townley R, Kuchin S, Carlson M. Subcellular localization of the Snf1 kinase is regulated by specific beta subunits and a novel glucose signaling mechanism. Genes Dev. 2001;15:1104–14. doi: 10.1101/gad.879301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hedbacker K, Hong SP, Carlson M. Pak1 protein kinase regulates activation and nuclear localization of Snf1-Gal83 protein kinase. Mol Cell Biol. 2004;24:8255–63. doi: 10.1128/MCB.24.18.8255-8263.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Corton JM, Gillespie JG, Hawley SA, Hardie DG. 5-aminoimidazole-4-carboxamide ribonucleoside A specific method for activating AMP-activated protein kinase in intact cells? Eur J Biochem. 1995;229:558–65. doi: 10.1111/j.1432-1033.1995.tb20498.x. [DOI] [PubMed] [Google Scholar]
- 32.Cameroni E, Hulo N, Roosen J, Winderickx J, De Virgilio C. The novel yeast PAS kinase Rim 15 orchestrates G0-associated antioxidant defense mechanisms. Cell Cycle. 2004;3:462–8. [PubMed] [Google Scholar]
- 33.Narasimhan ML, Damsz B, Coca MA, Ibeas JI, Yun DJ, Pardo JM, Hasegawa PM, Bressan RA. A plant defense response effector induces microbial apoptosis. Mol Cell. 2001;8:921–30. doi: 10.1016/s1097-2765(01)00365-3. [DOI] [PubMed] [Google Scholar]
- 34.Thomas P, Dressing G, Pang Y, Berg H, Tubbs C, Benninghoff A, Doughty K. Progestin, estrogen and androgen G-protein coupled receptors in fish gonads. Steroids. 2006;71:310–6. doi: 10.1016/j.steroids.2005.09.015. [DOI] [PubMed] [Google Scholar]
- 35.Thomas P, Pang Y, Dong J, Groenen P, Kelder J, de Vlieg J, Zhu Y, Tubbs C. Steroid and G protein binding characteristics of the seatrout and human progestin membrane receptor alpha subtypes and their evolutionary origins. Endocrinology. 2007;148:705–18. doi: 10.1210/en.2006-0974. [DOI] [PubMed] [Google Scholar]
- 36.Park SH, Koh SS, Chun JH, Hwang HJ, Kang HS. Nrg1 is a transcriptional repressor for glucose repression of STA1 gene expression in Saccharomyces cerevisiae. Mol Cell Biol. 1999;19:2044–50. doi: 10.1128/mcb.19.3.2044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Vyas VK, Kuchin S, Carlson M. Interaction of the repressors Nrg1 and Nrg2 with the Snf1 protein kinase in Saccharomyces cerevisiae. Genetics. 2001;158:563–72. doi: 10.1093/genetics/158.2.563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lesage P, Yang X, Carlson M. Analysis of the SIP3 protein identified in a two-hybrid screen for interaction with the SNF1 protein kinase. Nucleic Acids Res. 1994;22:597–603. doi: 10.1093/nar/22.4.597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mao X, Kikani CK, Riojas RA, Langlais P, Wang L, Ramos FJ, Fang Q, Christ-Roberts CY, Hong JY, Kim RY, Liu F, Dong LQ. APPL1 binds to adiponectin receptors and mediates adiponectin signalling and function. Nat Cell Biol. 2006;8:516–23. doi: 10.1038/ncb1404. [DOI] [PubMed] [Google Scholar]
- 40.Courel M, Lallet S, Camadro JM, Blaiseau PL. Direct activation of genes involved in intracellular iron use by the yeast iron-responsive transcription factor Aft2 without its paralog Aft1. Mol Cell Biol. 2005;25:6760–71. doi: 10.1128/MCB.25.15.6760-6771.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rutherford JC, Ojeda L, Balk J, Muhlenhoff U, Lill R, Winge DR. Activation of the iron regulon by the yeast Aft1/Aft2 transcription factors depends on mitochondrial but not cytosolic iron-sulfur protein biogenesis. J Biol Chem. 2005;280:10135–40. doi: 10.1074/jbc.M413731200. [DOI] [PubMed] [Google Scholar]
- 42.Santos R, Dancis A, Eide D, Camadro JM, Lesuisse E. Zinc suppresses the iron-accumulation phenotype of Saccharomyces cerevisiae lacking the yeast frataxin homologue (Yfh1) Biochem J. 2003;375:247–54. doi: 10.1042/BJ20030835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Waters BM, Eide DJ. Combinatorial control of yeast FET4 gene expression by iron, zinc, and oxygen. J Biol Chem. 2002;277:33749–57. doi: 10.1074/jbc.M206214200. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Figure 1: Functional expression of Izh2p-3HA. (A) Izh2p or Izh2p-3HA overexpression represses the induction of pFET3-397 in low iron. (B) Western blot showing that the Izh2p-3HA tagged protein is expressed in cells grown in galactose but not in cells grown in glucose. (C) Western blot showing that the Izh2p-3HA tagged protein is induced in cells carrying the Izh2p-3HA overexpression construct but not in those carrying the untagged Izh2p construct. Protein samples in B and C were heated to 37º C rather than boiled prior to loading.
Supplemental Figure 2: Aft1p responds normally to iron-deficiency in most mutant strains. (A) Ferroxidase activity is induced by iron-deficiency in the BY4742 wild type strain. Fet3p activity is not induced in isogenic strains lacking SNF1, MSN2 or MSN4. (B) Ferroxidase activity is inducible by iron-deficiency in the MCY5326 wild type strain, the isogenic nrg1Δnrg2Δ double mutant and the isogenic nrg1Δnrg2Δmsn2Δ msn4Δ quadruple mutant but not in the isogenic msn2Δmsn4Δ double mutant. (C) b-galactosidase activity of the pFET3-397 construct is inducible by iron-deficiency in the BY4742 wild type strain and in all isogenic mutants tested except for the gal83Δ, sak1Δ and msn4Δ mutants. For all panels, activities are shown as a % of isogenic wild type control grown in low iron media. The legend above panel A applies to panels A–C.