

Abstract
Patients with cancer frequently report pain that can be difficult to manage. This study examined the antihyperalgesic effects of a cannabinoid receptor agonist, CP 55,940, in a murine model of cancer pain. Implantation of fibrosarcoma cells into and around the calcaneous bone in mice produced mechanical hyperalgesia (decreased paw withdrawal thresholds and increased frequency of paw withdrawals). On day 13 after implantation, mechanical hyperalgesia, nociception, and catalepsy were assessed. Mice were randomly assigned to receive CP 55,940 (0.01–3 mg/kg, i.p.) or vehicle and behavioral measures were redetermined. CP 55,940 dose-dependently attenuated tumor-evoked mechanical hyperalgesia. To examine the effect of catalepsy on the antihyperalgesic effect of CP 55,940, mice with tumor-evoked hyperalgesia were pretreated with the dopamine agonist apomorphine prior to administration of CP 55,940. Apomorphine attenuated the cataleptic effect of CP 55,940 but did not attenuate its antihyperalgesic effect. In a separate group of mice with tumor-evoked hyperalgesia, administration of the dopamine antagonist spiperone produced catalepsy that was ~2.5 fold greater than that produced by CP 55,490. Whereas this dose of CP 55,940 completely reversed tumor-evoked mechanical hyperalgesia, spiperone only attenuated mechanical hyperalgesia by ~35%. Thus, the cataleptic effects of CP 55,940 did not fully account for its antihyperalgesic effect. The antihyperalgesic effect of CP 55,940 was mediated via the cannabinoid CB1 but not CB2 receptor. Finally, repeated administration of CP 55,940 produced a short-term and a longer term attenuation of tumor-evoked hyperalgesia. These results suggest that cannabinoids may be a useful alternative or adjunct therapy for treating cancer pain.
Keywords: Cannabinoids, cancer pain, fibrosarcoma, mechanical hyperalgesia, nociception, catalepsy, dopamine
1. Introduction
An estimated 1.4 million new cases of cancer will be diagnosed in the United States in 2006 (Jemal et al., 2006). Pain will often be the first symptom reported (Daut and Cleeland, 1982; Miser et al., 1987) and becomes more prevalent and more severe with progression of the disease until up to 85% of patients with terminal cancer report severe pain (Foley, 2000; Portenoy, 1989). Bone is a common site of tumor metastases, particularly in patients with lung, breast, and prostate cancer (Rubens, 1998). Patients with bone metastases report cancer-related pain more often than patients with local and regional disease (Ahles et al., 1984; Daut and Cleeland, 1982). Frequently, the first indication of bone metastases is pain, and pain is the most frequent symptom for patients with bone cancer (Pecherstorfer and Vesely, 2000). Thus, pain related to metastasis of cancer to bone is a common and debilitating complication of cancer.
The primary treatment for severe cancer pain is administration of opioids (Cherny, 2004). Effective analgesia can be obtained in 70–90% of patients with cancer pain by using the guidelines of the World Health Organization’s (WHO’s) ‘analgesic ladder’ (Cherny, 2004). Unfortunately, many cancer patients do not obtain sufficient analgesia in routine care (Beck and Falkson, 2001; McMillan et al., 2000). Therefore, alternative or adjunctive therapeutic approaches are needed for the management of cancer pain.
One potential approach for the treatment of cancer pain is the use of cannabinoids. Cannabinoids are compounds that bind to cannabinoid receptors, of which there are two cloned subtypes, CB1 and CB2 (Matsuda et al., 1990; Munro et al., 1993). Cannabinoid CB1 receptors are located in the central and peripheral nervous systems, including areas that contribute to nociceptive signaling and modulation (Hohmann, 2002; Pertwee, 2001). Cannabinoid CB2 receptors are found primarily on immune cells (Hohmann, 2002; Mbvundula et al., 2004). Cannabinoids attenuate behavioral responses to noxious stimuli and attenuate hyperalgesia produced by inflammation and nerve injury in animals (for review see (Hohmann, 2002; Martin and Lichtman, 1998; Walker et al., 1999) and humans (Karst et al., 2003). In preliminary studies, THC produced analgesia equivalent to that of codeine in patients with cancer pain (Noyes et al., 1975). Using a murine model of bone cancer pain, we found that a cannabinoid (WIN 55,212-2) attenuated (~50%) deep-tissue hyperalgesia produced by implantation of fibrosarcoma cells into the humerus (Kehl et al., 2003). The aim of this study was to determine if a more potent cannabinoid, CP 55,940, attenuates tumor-evoked cutaneous mechanical hyperalgesia in a well-characterized murine model of cancer pain. In this model, implantation of osteolytic fibrosarcoma cells into and around the calcaneous bone produces robust mechanical hyperalgesia and sensitization of C nociceptors (Cain et al., 2001; Wacnik et al., 2001). This model will allow us to correlate the antihyperalgesic effects of cannabinoids to its effects on tumor-evoked sensitization of nociceptors. To provide clinically useful pain relief for patients with cancer pain, it is important that cannabinoids be effective when given repeatedly. Thus, we also examined if repeated administration of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia over a 6.5 day period.
2. Materials and Methods
All experiments complied with the guidelines set forth by the International Association for the Study of Pain for the use of experimental animals (Zimmermann, 1983) and were approved by the Institutional Animal Care and Use Committee at the University of Minnesota. For all the experiments, the observer was blinded to the treatment administered.
2.1. Subjects
Adult (>5 weeks old) male C3H/HeNCr mice (National Cancer Institute, Bethesda, MD) weighing 20–30 g were used in all experiments. Mice were housed 4 per box, had free access to mouse chow and water, and were maintained on a 12-h light/dark schedule. Each mouse was used in only one experiment.
2.2. Implantation of Fibrosarcoma Cells
NCTC 2472 fibrosarcoma cells (ATCC, Manassas, VA) were maintained as described previously (Clohisy et al., 1996). Briefly, fibrosarcoma cells were grown to confluency in 75 cm2 flasks in NCTC 135 medium (pH 7.4) containing 10% horse serum and prepared for implantation by creating a cell suspension with trypsin. Fibrosarcoma cells were counted using a hemocytometer, pelleted and resuspended in phosphate buffered saline for implantation.
Fibrosarcoma cells were implanted into the hind paw as described previously (Wacnik et al., 2001). Mice were briefly anesthetized with halothane (2–3%) and fibrosarcoma cells (2 × 105 cells/10 μl) were injected into and around the calcaneous bone of the left hind paw using a 0.3 cc insulin syringe with a 29.5 gauge needle. No mice showed signs of motor dysfunction after implantation of fibrosarcoma cells.
2.3. Mechanical Hyperalgesia
Hyperalgesia, characterized by spontaneous pain, decreased threshold to pain, and increased painfulness to suprathreshold stimuli (Campbell et al., 1979), was examined with von Frey (Semmes-Weinstein) filaments (Stoelting Co, Wood Dale, IL) using two methods. First, paw withdrawal thresholds were determined using the up-down method described by Chaplan and colleagues (Chaplan et al., 1994). Second, the frequency of paw withdrawals evoked by a standard von Frey filament was determined. For both methods, mice were placed on a wire mesh platform, covered with a glass container (10 × 6.5 × 6.5 cm), and allowed to acclimate for at least 30 min. Each von Frey filament was applied to the plantar surface of the hind paw at random locations avoiding the heel, toe, and toe pads. For each trial, the filament was applied for 1–2 s with an interstimuli interval of approximately 5 s. Only vigorous paw withdrawals were counted.
In a preliminary experiment, the frequency of paw withdrawals of nontumor-bearing mice (n=6) to a series of von Frey filaments was examined to determine the range of filaments to be used in the up-down method and to select the standard filament. The forces produced by these von Frey filaments (bending forces of 0.11 to 93 mN; 0.011 to 9.5 g) were determined using a pan balance. Each von Frey filament, beginning with the filament with the weakest bending force, was applied 10 times to the plantar surface of the hind paw and the frequency of paw withdrawals was recorded for each mouse. Then, the frequency of paw withdrawals to the filament with the next weakest bending force was determined. The von Frey filaments with bending forces greater than 58 mN (5.9 g) lifted the hind paw before bending and were not used further. After the frequency of paw withdrawals to the remaining filaments were determined, a stimulus-response curve was plotted.
To determine paw withdrawal thresholds, testing was initiated with the 6.0 mN (0.61 g) von Frey filament. This von Frey filament evoked a paw withdrawal in ~50% of the applications in the preliminary experiment and its bending force was the fourth in intensity of 8 filaments (0.38 to 18.8 mN, 0.039 to 1.91 g) for use in the up-down method. Subsequent stimuli were presented based on the response to the previous stimulus. In the absence of a paw withdrawal to a given stimulus, a stronger stimulus was presented. In the case of a paw withdrawal, a weaker stimulus was presented. Six responses, starting with the negative response immediately before the first paw withdrawal, were recorded. The resulting pattern was tabulated and the 50% paw withdrawal threshold was calculated using the formula (modified from (Chaplan et al., 1994)):
Where Xf = value (in log units) of the final von Frey filament used, K = tabular value for the pattern of positive/negative responses, and δ = mean difference of forces (0.242) in log units produced by the von Frey filaments. Mechanical hyperalgesia was defined as a decrease in paw withdrawal threshold.
To determine responses to a standard stimulus, a von Frey filament with a bending force of 3.7 mN was used to evoke paw withdrawals. This filament evoked a paw withdrawal in approximately 15% of trials in nontumor-bearing mice and therefore was not a suprathreshold stimulus (i.e., it did not evoke a paw withdrawal in >50% of trials). However, this filament was chosen to allow for a greater increase in the frequency of paw withdrawals as mechanical hyperalgesia developed. Tumor-evoked mechanical hyperalgesia was defined as an increase in the frequency of paw withdrawals. Only mice that exhibited sufficient mechanical hyperalgesia, defined as paw withdrawals in at least 50% of the 10 trials, were used in subsequent experiments. Approximately 90% of mice implanted with fibrosarcoma cells exhibited sufficient mechanical hyperalgesia on day 13 after implantation.
2.4. Catalepsy
Cannabinoids produce catalepsy, a state in which an animal remains in an externally imposed position (Sanberg et al., 1988). Catalepsy may decrease a mouse’s ability to withdraw its hind paw, which could be misinterpreted as antihyperalgesia. The bar test was used to determine the level of catalepsy produced by CP 55,940, (Kuschinsky and Hornykiewicz, 1972). Each mouse was placed with its fore limbs on a metal bar (diameter = 1 cm) positioned 5 cm above and parallel to the counter top. Its hind paws rested on the counter top. The time each mouse spent in this position in 2 consecutive trials (60 s each) was recorded then averaged. The bar test was performed immediately after evaluation of mechanical hyperalgesia. Catalepsy was defined as an increase in the time that mice were motionless on the bar.
2.5. Nociception
The antinociceptive effect of CP 55,940 was assessed after the bar test using the tail-flick assay (Harris et al., 1969). Each mouse was gently held in one hand with a 1 cm section of its tail, 1.5 cm from the tip, positioned over a radiant heat source. The intensity of the heat source was adjusted so that, at baseline, mice flicked their tail at ~ 4 s. A cut off time of 12 s was chosen to avoid tissue damage.
2.6. Motor Function
An accelerating (5–40 rpm) treadmill apparatus, the Rota-rod (Stoelting, Wood Dale, IL), was used to examine the effect of CP 55,940 on motor function (Dunham and Miya, 1957). Naïve mice (>5 weeks old) were used in this experiment because mice implanted with fibrosarcoma cells developed enlargement of the hind paw that made gripping the Rota-rod difficult. On the day prior to the experiment, mice were trained on the Rota-rod apparatus for 6 consecutive trials of 5 min, and the length of time (s) until they fell off of the Rota-rod was recorded. Mice that failed to stay on the Rota-rod for at least 180 s during the 6th trial underwent additional trials until they did. The time that each mouse stayed on the Rota-rod apparatus during its last trial was recorded as its predrug score. The next day, mice were weighed and then randomly assigned to receive CP 55,940 (0.03–3 mg/kg, i.p.) or vehicle. Thirty minutes after administration of CP 55,940 or vehicle, the length of time that each mouse remained on the Rota-rod was determined. Motor dysfunction was indicated by a decrease in the time that a mouse stayed on the Rota-rod.
2.7. Drug Preparation and Administration
The nonselective cannabinoid receptor agonist, (-)-cis-3-[2-hydroxy-4(1,1-dimethylheptyl)phenyl]-trans-4-(3-hydroxypropyl)cyclohexanol (CP 55,940; 0.003–1 mg/ml; Tocris-Crookson, Ellisville, MO), the cannabinoid CB1 receptor selective antagonist, N-(piperidin-1-yl)-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide (SR 141716A; 1 mg/ml, National Institute of Drug Abuse, Bethesda, MD) and the cannabinoid CB2 receptor selective antagonist, N-[(1S)-endo-1,3,3-trimethylbicyclo[2.2.1]heptan-2-yl]-5-(4-chloro-3-methylphenyl)-1-(4-methylbenzyl)-pyrazole-3-carboxamide (SR 144528; 0.3 mg/ml, National Institute of Drug Abuse, Bethesda, MD) were prepared in 5% emulphor, 5% ethanol and 90% normal saline. The dopamine receptor agonist, (R)-5,6,6a,7,-tetrahydro-6-methyl-4H-dibenzo[de,g]quinoline-10,11-diol hydrochloride (apomorphine; 3.0 mg/ml; Sigma, St. Louis, MO) was dissolved in 0.9% saline and the dopamine receptor antagonist, 8-[4-(4-fluorophenyl)-4-oxobutyl]-1-phenyl-1,3,8-triazaspiro[4,5]decan-4-one hydrochloride (spiperone; 0.4 mg/ml; Tocris-Crookson, Ellisville, MO) was dissolved in distilled water. All drugs were administered intraperitoneally, except spiperone and its vehicle, which were administered subcutaneously in the back. All drugs were administered in a volume of 0.1 ml/10 gm of body weight.
2.8. Experimental design
2.8.1 Comparison of two measures of tumor-evoked mechanical hyperalgesia
Baseline paw withdrawal thresholds and the frequencies of paw withdrawals to the standard von Frey filament were determined for each mouse for 2–3 days before implantation of fibrosarcoma cells. After fibrosarcoma cells were implanted, these measures of mechanical hyperalgesia were determined on days 3, 5, 7, 9, and 11 with day 0 being the day of implantation. On day 13, predrug levels of mechanical hyperalgesia were determined. Then, mice were randomized into groups and injected intraperitoneally with CP 55,940 (0.1 or 0.3 mg/kg) or vehicle. Postdrug levels of mechanical hyperalgesia were determined 30 min later.
2.8.2 Time- and dose-dependent antihyperalgesia, catalepsy and antinociception induced by CP 55,940 in mice with tumor-evoked mechanical hyperalgesia
Fibrosarcoma cells were implanted as described above and the levels of mechanical hyperalgesia, catalepsy, and nociception were determined on days 3, 5, 7, 9, and 11. On day 13, predrug levels of mechanical hyperalgesia, catalepsy and nociception were determined. Mice were randomly assigned to receive an injection of CP 55,940 (0.03–3 mg/kg) or vehicle. Postdrug levels of mechanical hyperalgesia, catalepsy, and nociception were determined 30 min, 90 min, 3 h and 24 h later.
2.8.3 Contribution of catalepsy to the antihyperalgesic effect of CP 55,940
To examine the contribution of catalepsy to the antihyperalgesic effect of CP 55,940, two experiments were performed. In the first experiment, we attempted to block cannabinoid-induced catalepsy by pretreating mice with apomorphine (30 mg/kg) 30 min prior to administration of CP 55,940 (0.3 mg/kg). Then, we examined whether apomorphine altered the effect of CP 55,940 on tail-flick latencies and frequency of paw withdrawals. Apomorphine attenuates opioid-induced catalepsy in mice (Erzin-Waters et al., 1976; Kuschinsky and Hornykiewicz, 1972). In the second experiment, we induced catalepsy by administering spiperone (4 mg/kg) (Cook et al., 2000; Yurek and Randall, 1985) and, after 30 min, determined whether spiperone attenuated the frequency of paw withdrawals in mice with tumor-evoked mechanical hyperalgesia.
2.8.4 Effect of motor dysfunction on CP 55,940-induced antihyperalgesia
The dose-response relationship of CP 55,940 induced motor dysfunction was determined in nontumor-bearing mice. Mice were trained on the Rota-rod apparatus and the time that mice remained on the apparatus was determined. The next day, mice were randomly assigned to receive CP 55,940 (0.03 to 3 mg/kg) or vehicle. Thirty minutes later, the time that mice remained on the Rota-rod apparatus was redetermined.
2.8.5 Role of cannabinoid CB1 and CB2 receptor subtypes in the effects of CP 55,940
Predrug values of tumor-evoked mechanical hyperalgesia, catalepsy and nociception were obtained on day 13 after implantation of the fibrosarcoma cells. SR 141716A (10 mg/kg), SR144528 (3 mg/kg), or vehicle was administered intraperitoneally. Then 15 min later, CP 55,940 (1 mg/kg) or vehicle was administered and mechanical hyperalgesia, catalepsy, and nociception were determined 30 min later.
2.8.6 Antihyperalgesic effect of repeated administration of CP 55,940
The clinical usefulness of any treatment for a chronic condition will depend upon its effectiveness when given repeatedly. Therefore, we examined whether twice-daily administration of CP 55,940 for 6.5 days would attenuate tumor-evoked mechanical hyperalgesia in this murine model of cancer pain. Mice were implanted with fibrosarcoma cells and tumor-evoked mechanical hyperalgesia was allowed to develop. On day 9 after implantation, the level of mechanical hyperalgesia was determined, and mice were randomly assigned to receive CP 55,940 (0.1 mg/kg) or vehicle. Thirty minutes after administration of CP 55,940 or vehicle, the level of mechanical hyperalgesia was redetermined. Approximately 6 h later, mice received a second daily administration of CP 55,940 or vehicle. This procedure was repeated on days 11, 13, and 15. On days 10, 12, and 14, mice received twice-daily administration of CP 55,940 or vehicle but no behavioral testing was performed.
2.9. Statistical Analyses
Frequency of paw withdrawals and paw withdrawal threshold force (mechanical hyperalgesia), tail-flick withdrawal latency (nociception), time on the bar (catalepsy), and time to fall from the Rota-rod (motor dysfunction) are presented as mean ± S.E.M. Each dependent measure was compared between treatment groups (dose of CP 55,940 or vehicle) and across test times (predrug, 30 min, 90 min, 3 h and 24 h) using a two-way repeated measures ANOVA. In experiments in which mice were pretreated (i.e., apomorphine, SR 141716A, or SR 144528), each dependent measure was compared between treatment groups using a three-way repeated measures ANOVA with two treatment factors (pretreatment with SR 141716A, SR 144528 or vehicle; treatment with CP 55,940 or vehicle) and one repeated measure (predrug or postdrug test time). In the experiments in which mice received repeated administration of CP 55,940 or vehicle, paw withdrawal frequencies were compared between treatment groups, across post-implantation days and across test times (predrug vs postdrug) using a three-way repeated measures ANOVA. Individual pairwise comparisons were made using paired and unpaired t-tests with the Bonferroni correction for multiple comparisons.
To examine the dose-response relationships of CP 55,940’s effect on tumor-evoked mechanical hyperalgesia, nociception, catalepsy, and motor dysfunction, data were converted to % Maximum Possible Effect (%MPE) using the following equations:
Frequency of paw withdrawals:
Tail-flick latency, time on the bar, and time to fall
The cut off scores were 12 s for the tail-flick assay, 60 s for the bar test, and 300 s for the Rota-rod. A one-way ANOVA was performed to determine if the frequency of paw withdrawals, tail-flick latency, bar test time, and Rota-rod time differed among the doses of CP 55,940 administered. Then, the ED50 (50% Effective Dose) and 95% confidence intervals (95% CI) were calculated from the %MPE using the method of Tallarida and Murray (1987).
Associations between the measures of mechanical hyperalgesia (i.e., threshold force and frequency of paw withdrawals) were determined by calculating Pearson’s correlation coefficients. For all statistical analyses, a probability value < 0.05 was considered significant.
3. Results
3.1. Tumor-evoked mechanical hyperalgesia
Implantation of fibrosarcoma cells into and around the calcaneous bone of the left hind paw of mice produced mechanical hyperalgesia as indicated by a decrease in paw withdrawal threshold and an increase in the frequency of paw withdrawals to a standard von Frey filament (Fig. 1). Prior to implantation of fibrosarcoma cells, the threshold force for evoking a paw withdrawal did not differ between left and right hind paws (Fig. 1A). Paw withdrawal thresholds decreased from baseline values in the tumor-bearing hind paw beginning 3 days after implantation of fibrosarcoma cells (P < 0.001). Paw withdrawal thresholds remained lower than baseline values and lower than the contralateral control hind paw at all post-implantation test days (P < 0.001). Hence, implantation of fibrosarcoma cells into and around the calcaneous bone produced a decrease in paw withdrawal threshold.
Fig. 1.
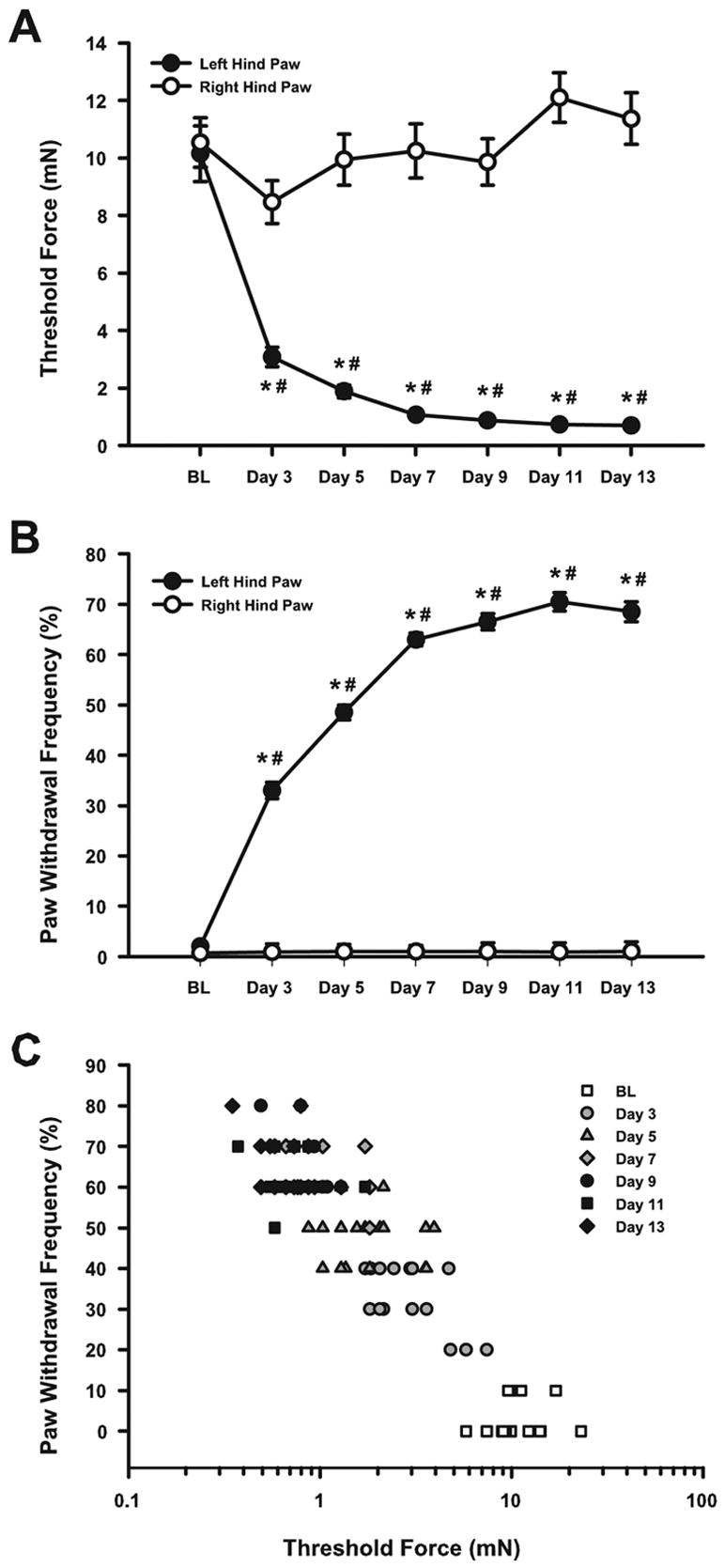
Paw withdrawal threshold force and the frequency of paw withdrawals during development of tumor-evoked mechanical hyperalgesia. (A) Implantation of fibrosacroma cells decreased the threshold force to evoke a paw withdrawal beginning 3 days after implantation. (B) Implantation of fibrosarcoma cells increased the frequency of paw withdrawals to the standard von Frey filament (3.7 mN). (C) During the development of tumor-evoked mechanical hyperalgesia, paw withdrawal thresholds were negatively correlated with frequencies of paw withdrawals. N = 20 mice, *P < 0.001 for baseline (BL) compared to post-implantation day (3–13). #P < 0.001 for left compared to right hind paw at each test day.
The frequency of paw withdrawals in response to stimulation of the plantar surface of the tumor-bearing hind paw with a von Frey filament (3.7 mN) increased from baseline as early as 3 days after implantation and progressively increased until 11 days after implantation (P < 0.001; Fig. 1B). In contrast, the frequency of withdrawals of the right hind paw did not change over time. The frequency of withdrawals of the left hind paw was higher than for the right paw at all post-implantation test days (P < 0.001).
The time course of tumor- evoked decrease in paw withdrawal threshold (Fig. 1A) corresponded to the time course of the increase in paw withdrawal frequency (Fig. 1B). Furthermore, as illustrated in Fig. 1C, the threshold force was negatively correlated (r = −0.82) with the frequency of paw withdrawals from baseline to day 13 after implantation (P < 0.001). The paw withdrawal threshold decreased and the frequency of paw withdrawals increased with increasing time after implantation of fibrosarcoma cells. Thus, these two criteria for hyperalgesia were strongly negatively correlated during the development of tumor-evoked mechanical hyperalgesia.
3.2. CP 55,940 attenuated tumor-evoked mechanical hyperalgesia
Systemic administration of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia. Thirteen days after the implantation of fibrosarcoma cells, there were no differences in paw withdrawal thresholds between groups (predrug) (Fig. 2A). Thirty minutes after administration of CP 55,940 (0.3 mg/kg), threshold increased to 16.2 ± 1.4 mN (P <0.001). This value was similar to the threshold of these mice prior to implantation of fibrosarcoma cells (10.5 ± 0.97 mN, Fig. 1A).
Fig. 2.
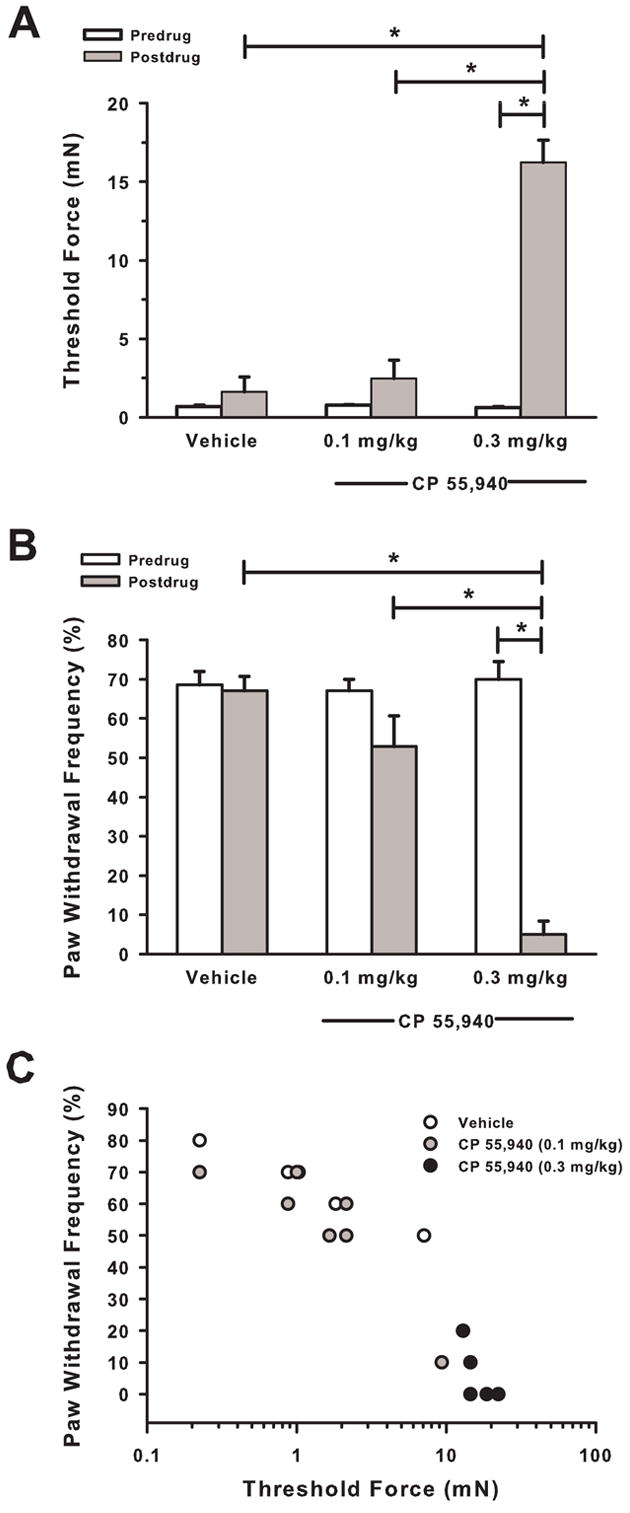
Paw withdrawal threshold force and frequency of paw withdrawals during attenuation of tumor-evoked mechanical hyperalgesia by CP 55,940. In mice with tumor-evoked mechanical hyperalgesia, CP 55,940 (0.3 mg/kg) (A) increased paw withdrawal thresholds and (B) decreased the frequency of paw withdrawals to a standard von Frey filament (3.7 mN). (C) During the attenuation of tumor-evoked mechanical hyperalgesia, paw withdrawal thresholds were negatively correlated with frequencies of paw withdrawals. N = 6–7 mice/group, *P < 0.001 for predrug compared to postdrug (30 min) values.
Similarly, mice withdrew their left hind paw in ~70% of the trials, and there were no differences between groups (Fig. 2B). Thirty minutes after administration of CP 55,940, the frequency of paw withdrawals decreased (P <0.001). In contrast, there was no change in the frequency of paw withdrawals for mice that received vehicle. Thus, systemic administration of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia as indicated by a return in threshold forces and the frequency of paw withdrawals to baseline values.
The two measures of mechanical hyperalgesia, paw withdrawal thresholds and the frequencies of paw withdrawals, were highly negatively correlated (r = −0.94) after administration of CP 55,940 (0.1 or 0.3 mg/kg) or vehicle (P < 0.001, Fig. 2C). Mice that exhibited the greatest increase in threshold force also exhibited the greatest decrease in the frequency of paw withdrawals. Thus, these two measures of mechanical hyperalgesia were strongly negatively correlated during the development of tumor-evoked mechanical hyperalgesia (Fig. 1C) and during attenuation of mechanical hyperalgesia by CP 55,940 (Fig. 2C). Because these measures were highly correlated, we used the frequency of paw withdrawals as our measure of mechanical hyperalgesia in subsequent experiments.
Systemic administration of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia in a time-dependent manner. Thirteen days after implantation of fibrosarcoma cells, mice withdrew their left hind paw in ~68% of the trials (Fig. 3A). Thirty minutes after administration of CP 55,940 (0.1, 0.3, 1 and 3 mg/kg), the frequency of paw withdrawals decreased from predrug values (P < 0.01). The antihyperalgesic effect of CP 55,940 continued at 90 min for the 0.1 to 1 mg/kg doses and at 3 h for the 0.3 to 1 mg/kg doses. The dose of 3 mg/kg was only tested at 30 min and in not shown in the time course (Fig. 3A). The greatest decrease in the frequency of paw withdrawals occurred at 30 and 90 min after administration of CP 55,940 (0.3 and 1 mg/kg) and gradually returned to predrug values by 24 h. Because the antihyperalgesic effect of CP 55,940 was similar at 30 and 90 min, 30 min was chosen as the postdrug time point for determination of the dose-response relationships and in subsequent experiments.
Fig. 3.
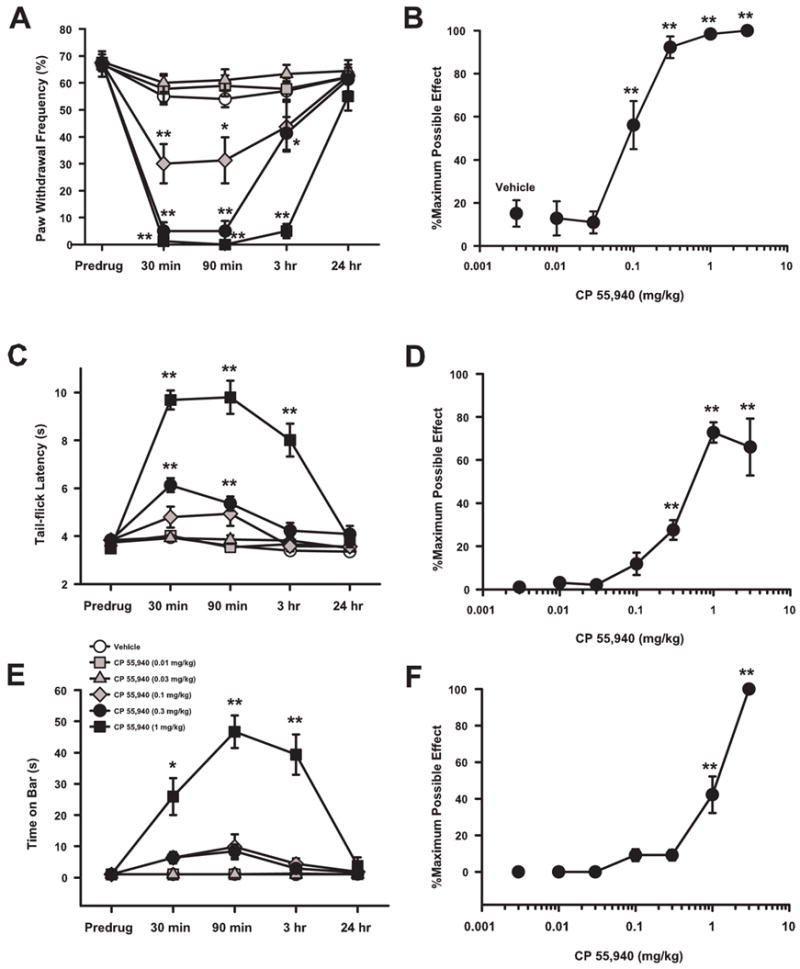
Antihyperalgesic, antinociceptive, and cataleptic effects of CP 55,940 in mice implanted with fibrosarcoma cells. (A) CP 55,940 (0.1–1 mg/kg) attenuated tumor-evoked mechanical hyperalgesia with peak antihyperalgesia occurring at 30 and 90 min. (B) CP 55,940 dose-dependently attenuated tumor-evoked mechanical hyperalgesia with nearly complete attenuation occurring at doses of 0.3 mg/kg and above. (C) CP 55,940 (0.3–1 mg/kg) decreased tail-flick latencies with peak antinociception occurring at 30 and 90 min. (D) CP 55,940 dose-dependently increased tail-flick latencies beginning at doses of 0.3 mg/kg and above. (E) CP 55,940 (1 mg/kg) produced catalepsy with peak time on the bar occurring at 90 min. (F) CP 55,940 dose-dependently increased time on the bar for doses of 1 and 3 mg/kg. The dose of 3 mg/kg was only tested at 30 min and therefore is not shown in the time courses (Panels A, C, & E). N = 8–10 mice/group. For panels A, C, and E; *P < 0.05, **P < 0.01 for predrug compared to postdrug values. For panels B, D, and F; *P < 0.05, **P < 0.01 for dose of CP 55,940 compared to vehicle.
Systemic administration of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia in a dose-dependent manner (Fig. 3B). The frequency of paw withdrawals 30 min after administration of CP 55,940 or vehicle was converted to % maximum possible effect (%MPE). CP 55, 940, at the doses of 0.1, 0.3, 1, and 3 mg/kg, produced higher %MPEs than did vehicle (P < 0.01). The 50% effective dose (ED50) was 0.089 mg/kg (95% CI = 0.064 to 0.124 mg/kg).
3.3. Attenuation of nociception by CP 55,940
The tail-flick assay was performed to examine the antinociceptive effect of CP 55,940 in mice with tumor-evoked mechanical hyperalgesia. CP 55,940 produced an increase in tail-flick latencies compared to the predrug latencies when administered at doses of 0.3 and 1 mg/kg (P < 0.01; Fig. 3C). The time course of the antinociceptive effect of CP 55,940 was similar to its antihyperalgesic effect in that the antinociceptive effect was greatest at 30 and 90 min after administration and returned to predrug levels by 24 h.
CP 55,940 produced an increase in the %MPE for tail-flick latency at doses of 0.3, 1, and 3 mg/kg when compared to vehicle (Fig. 3D; P < 0.01). The ED50 for the antinociceptive effect of CP 55, 940 was 1.31 mg/kg (95% CI = 0.86 to 2.25 mg/kg). Thus, CP 55,940 produced antinociception in a dose-dependent manner.
3.4. Effect of catalepsy on CP 55,940-induced antihyperalgesia
Thirty minutes after administration, CP 55,940 (1 mg/kg) increased the time spent on the bar when compared to predrug times (P < 0.01; Fig. 3E). Unlike the antihyperalgesic and antinociceptive effects of CP 55,940, the cataleptic effect was greatest 90 min after administration. By 24 h, the time spent on the bar returned to predrug values.
The dose-response relationship for the cataleptic effect of CP 55,940 at 30 min after administration showed that CP 55,940 at 1 and 3 mg/kg produced an increase in %MPE when compared to that produced by vehicle (P < 0.01; Fig. 3F). The ED50 for the cataleptic effect of CP 55,940 was 1.38 mg/kg (95% CI = 0.87 to 2.21 mg/kg). Thus, CP 55,940 produced catalepsy that was dose-dependent but peaked later than its antihyperalgesic and antinociceptive effects.
The role of the cataleptic effect of CP 55,940 on paw withdrawal was examined by administering a dopamine receptor agonist, apomorphine (30 mg/kg), 30 min prior to the administration of CP 55,940. When CP 55,940 (0.3 mg/kg) was administered to tumor-bearing mice pretreated with apomorphine vehicle, mice exhibited catalepsy as indicated by an increase in the time spent on the bar compared to predrug levels (P < 0.05; Fig. 4A). Mice pretreated with apomorphine prior to administration of the vehicle for CP 55,940 did not exhibit significant changes in time spent on the bar. Pretreatment with apomorphine decreased the time spent on the bar produced by CP 55,940 compared to that produced by pretreatment with apomorphine vehicle (P < 0.05). However, apomorphine did not completely reverse the catalepsy produced by CP 55,940 when compared to the predrug value (P < 0.01).
Fig. 4.
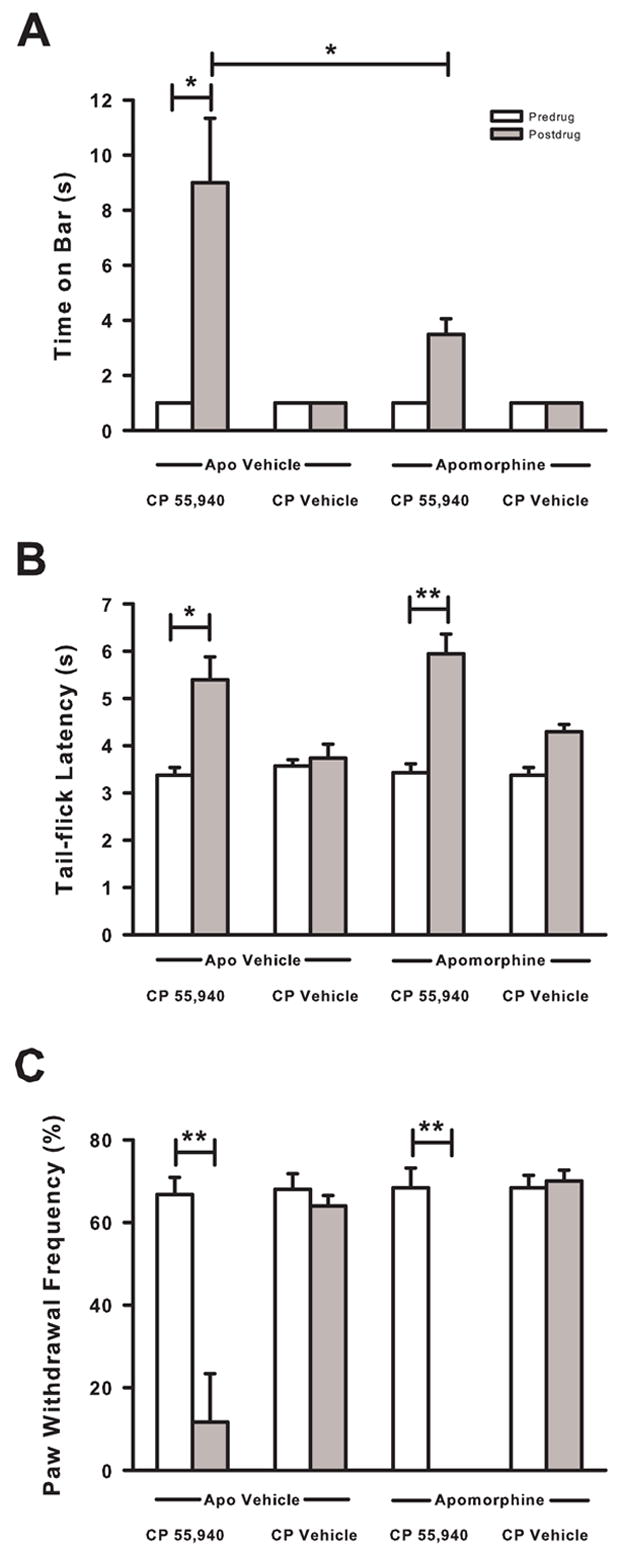
Effect of pretreatment with apomorphine on CP 55,940 induced catalepsy, antinociception, and antihyperalgesia in mice with tumor-evoked mechanical hyperalgesia. (A) Pretreament with apomorphine (30 mg/kg) 30 min prior to administration of CP 55,940 (0.3 mg/kg) decreased the time that mice spent on the bar by ~60%. (B) CP 55,940 induced increase in tail-flick latency was not attenuated by pretreatment with apomorphine. (C) Pretreatment with apomorphine did not attenuate the decrease in the frequency of paw withdrawals following administration of CP 55,940. N=5–6 mice/group. Postdrug = 30 min after administration of CP 55,940 or CP Vehicle (Veh). *P < 0.05 and **P < 0.01.
The effect of catalepsy on CP 55,940-induced antinociception was examined using the tail-flick assay (Fig. 4B). Pretreatment with the vehicle for apomorphine did not attenuate the increase in tail-flick latency produced by CP 55,940 (P <0.05). Mice pretreated with apomorphine prior to administration of the vehicle for CP 55,940 did not exhibit changes in tail-flick latencies. When apomorphine was administered prior to administration of CP 55,940, tail-flick latency increased (P < 0.01). Thus, apomorphine did not attenuate CP 55,940-induced antinociception.
The effect of pretreatment with apomorphine on CP 55,940-induced antihyperalgesia was examined (Fig. 4C). Pretreatment with the vehicle for apomorphine did not block the decrease in the frequency of paw withdrawals produced by CP 55,940 compared to predrug levels (P < 0.01). Pretreatment with apomorphine followed by administration with the vehicle for CP 55,940 did not change the frequency of paw withdrawals. However, CP 55,940 decreased the frequency of paw withdrawals in mice pretreated with apomorphine (P <0.01). Hence, pretreatment with apomorphine attenuated CP 55,940-induced catalepsy, but it did not attenuate the antinociceptive or antihyperalgesic effects of CP 55,940.
CP 55,940-induced catalepsy was not completely blocked by pretreatment with apomorphine. The small amount (~3.5 s) of catalepsy that remained may have been enough to decrease a mouse’s ability to withdraw its hind paw. Therefore, a dopamine antagonist, spiperone, was used to produce catalepsy in mice with tumor-evoked hyperalgesia and its ability to increase tail-flick latency and decrease the frequency of paw withdrawals were examined. The vehicles for CP 55,940 and spiperone did not increase the time mice spent on the bar (Fig. 5A). In contrast, CP 55,940 (0.3 mg/kg) and spiperone (4 mg/kg) increased the time that mice spent on the bar compared to predrug levels (P < 0.01). Spiperone produced a greater level of catalepsy than that produced by CP 55,940 (P < 0.05).
Fig. 5.
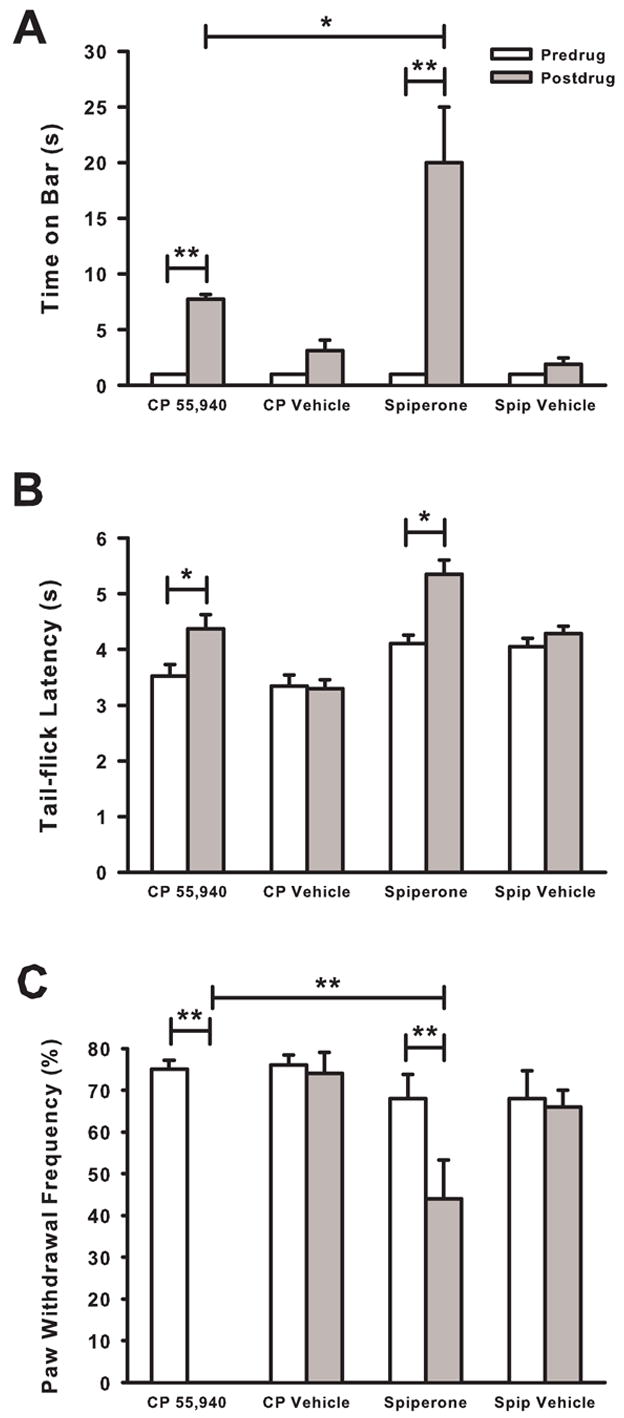
Effect of spiperone on bar test times, tail-flick latencies, and frequency of paw withdrawals. (A) Spiperone (4 mg/kg) produced an increase in time spent on the bar 30 min after administration. This increase was greater than that produced by CP 55,940 (0.3 mg/kg). (B) Spiperone and CP 55,940 increased tail-flick latencies. (C) Spiperone decreased the frequency of paw withdrawals by ~35% in mice with tumor-evoked hyperalgesia. In contrast, CP 55,940 completely attenuated tumor-evoked mechanical hyperalgesia. N=5–6 mice/group. *P < 0.05 and **P < 0.01.
Administration of the vehicles for CP 55,940 or spiperone did not increase tail-flick latencies compared to predrug latencies (Fig. 5B). However, CP 55,940 and spiperone increased tail-flick latency when compared to predrug levels (P < 0.05).
Administration of the vehicles for CP 55,940 or spiperone did not decrease the frequency of paw withdrawals compared to predrug levels (Fig. 5C). CP 55,940 produced a decrease in the frequency of paw withdrawals when compared to predrug values (P < 0.01), indicating that tumor-evoked mechanical hyperalgesia was completely reversed. Spiperone also decreased the frequency of paw withdrawals from predrug levels (P < 0.05). However, the frequency of paw withdrawals after the administration of spiperone was greater than that after the administration of CP 55,940 (P < 0.01). Thus, although spiperone produced more catalepsy than did CP 55,940, CP 55,940 attenuated tumor-evoked mechanical hyperalgesia completely whereas spiperone was only ~35% effective. The results of these studies using apomorphine to attenuate CP 55,940-induced catalepsy and spiperone to produce catalepsy in mice with tumor-evoked mechanical hyperalgesia suggest that the decrease in the frequency of paw withdrawals produced by CP 55,940 was not due only to its cataleptic effect.
3.5. Effect of motor dysfunction on CP 55,940-induced antihyperalgesia
We determined whether motor dysfunction contributed to the antihyperalgesic effect of CP 55,940 using a Rota-rod apparatus. In nontumor-bearing mice, CP 55,940 (1 and 3 mg/kg) decreased the time to fall compared to predrug levels (P < 0.05; Fig. 6A). Lower doses of CP 55,940 (0.03, 0.1, 0.3 mg/kg) and vehicle did not decrease the time to fall from the predrug levels. The dose-response relationship for CP 55,940-induced motor dysfunction revealed an ED50 of 1.27 mg/kg (95% CI = 0.78 to 2.10 mg/kg; Fig. 6B). Thus, CP 55,940 produced motor dysfunction at doses at or above 1 mg/kg in nontumor-bearing mice. Since the antihyperalgesic effect of CP 55,940 was almost 100% at 0.3 mg/kg in tumor-bearing mice, it is unlikely that motor dysfunction contributed to the antihyperalgesic effects of CP 55,940 in this model of cancer pain.
Fig. 6.
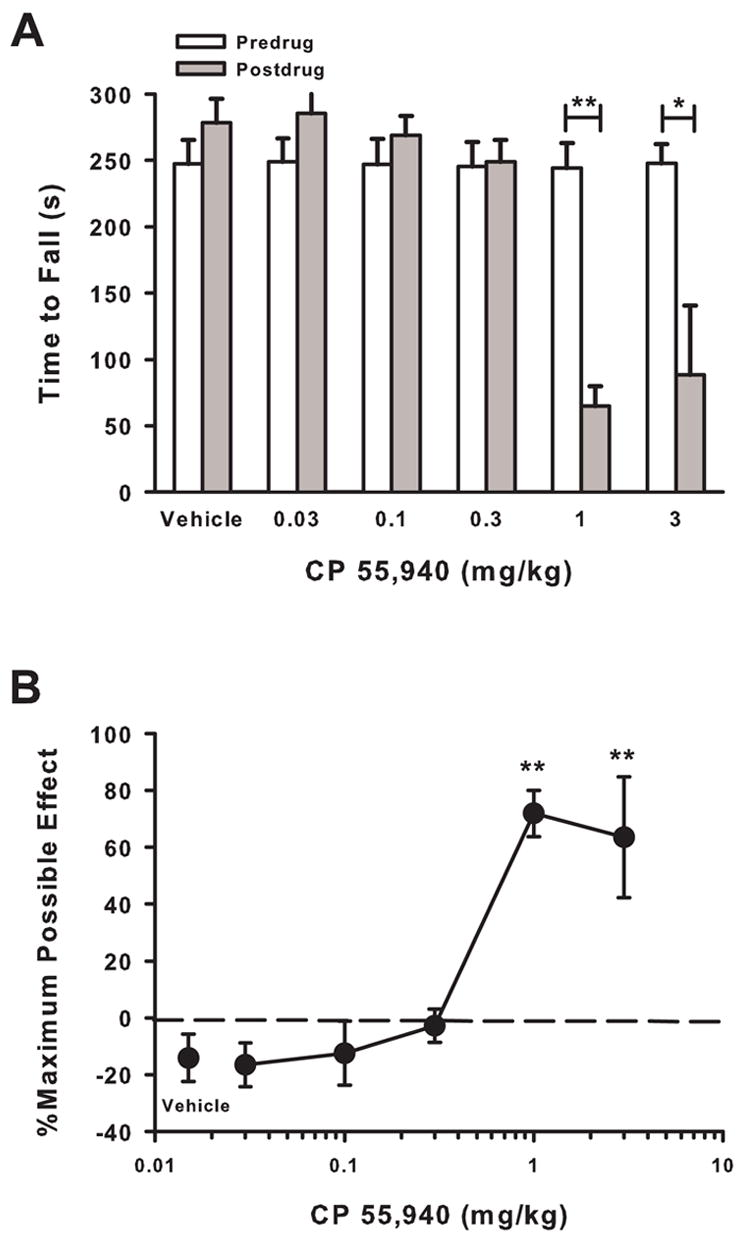
Effect of CP 55,940 on motor function in nontumor-bearing mice. (A) Thirty minutes after administration, CP 55,940 (1 and 3 mg/kg) decreased the time mice stayed on the accelerating Rota-rod apparatus. *P < 0.05 and **P < 0.01. (B) The effect of CP 5,940 on motor dysfunction was dose-dependent. N=5–6 mice/group. **P < 0.01 for dose of CP 55,940 compared to vehicle.
3.6. Role of cannabinoid CB1 and CB2 receptors in the effects of CP 55,940
Pretreatment with vehicle did not attenuate the antihyperalgesic effect of CP 55,940 (1 mg/kg) as the frequency of paw withdrawals decreased (P < 0.01; Fig. 7A). In contrast, pretreatment with the cannabinoid CB1 receptor selective antagonist (SR 141716A; 10 mg/kg) completely blocked the antihyperalgesic effect of CP 55,940; the frequency of paw withdrawals did not change from predrug values. However, the cannabinoid CB2 receptor selective antagonist (SR 144528; 3 mg/kg) did not attenuate the antihyperalgesic effect of CP 55,940 as the frequency of paw withdrawals decreased (P < 0.01). Pretreatment with SR 141716A followed by the administration of CP vehicle had no effect on the frequency of paw withdrawals indicating that SR 141716A itself did not alter mechanical hyperalgesia. Thus, the antihyperalgesic effect of CP 55,940 appeared to be mediated by the cannabinoid CB1 receptor subtype.
Fig. 7.
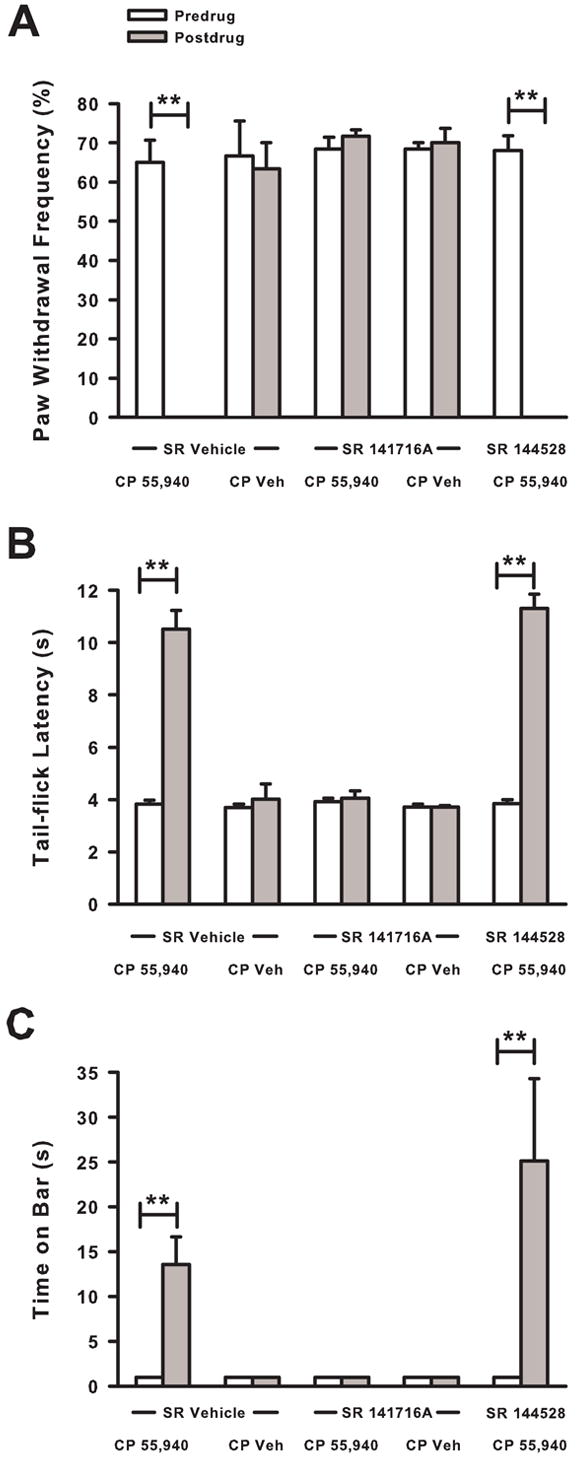
Effect of pretreatment with cannabinoid CB1 and CB2 receptor subtype selective antagonists on CP 55,940 induced antihyperalgesia, antinociception, and catalepsy. Pretreatment with the CB1 receptor subtype selective antagonist (SR 141716A, 10 mg/kg), but not the CB2 receptor subtype selective antagonist (SR 144528, 3 mg/kg), completely blocked the (A) antihyperalgesic, (B) antinociceptive, and (C) cataleptic effects of CP 55,940 (1 mg/kg). N=5–6 mice/group. **P < 0.01
Pretreatment with vehicle did not attenuate the increase in tail-flick latency produced by CP 55,940 (P < 0.01; Fig. 7B). In contrast, pretreatment with SR 141716A blocked the antinociceptive effect of CP 55,940; tail-flick latency after administration of CP 55,940 did not differ from predrug values. Pretreatment with SR 144528 did not attenuate the antinociceptive effect produced by CP 55,940 as tail-flick latency increased (P < 0.01). Thus, the antinociceptive effect of CP55,940 appeared to be mediated by the cannabinoid CB1, but not CB2, receptor subtype.
Pretreatment with vehicle did not attenuate the level of catalepsy produced by CP 55,940 (Fig. 7C) as the time on the bar increased from before administration of vehicle and CP 55,940 to after administration (P < 0.01). Pretreatment with SR 141716A completely blocked the catalepsy produced by CP 55,940 such that the time on the bar after administration of CP 55,940 was not was not different from predrug values. In contrast, pretreatment with SR 144528 failed to attenuate the cataleptic effect of CP 55,940 (P < 0.01). Thus, the cataleptic effect of CP 55,940, similar to its antihyperalgesic and antinociceptive effects, was mediated by the cannabinoid CB1, and not CB2, receptor subtype.
3.7. Effect of repeated administration of CP 55,940 on tumor-evoked mechanical hyperalgesia
We examined whether CP 55,940 would continue to attenuate tumor-evoked mechanical hyperalgesia when given repeatedly, as would likely be the case for patients with cancer pain. On day 9 after implantation, mice exhibited paw withdrawal frequencies of ~70% (Fig. 8). Then, mice received twice-daily administration of CP 55,940 (0.1 mg/kg) or vehicle for 6.5 days. By the next predrug testing time (Day 11), mice that received CP 55,940 exhibited a lower paw withdrawal frequency than on day 9 (P < 0.005) and less than the paw withdrawal frequency on day 11 of mice that received vehicle (P < 0.001). This effect was due to the repeated administration of CP 55,940, and not to that morning’s administration, because this paw withdrawal frequency was determined before mice received the morning dose of CP 55,940. Repeated administration of CP 55,940 continued to attenuate tumor-evoked mechanical hyperalgesia until the end of the study on day 15 as predrug paw withdrawal frequencies remained below predrug paw withdrawal frequency on day 9 (P < 0.001) and below predrug paw withdrawal frequencies of mice that received vehicle (P < 0.001).
Fig. 8.
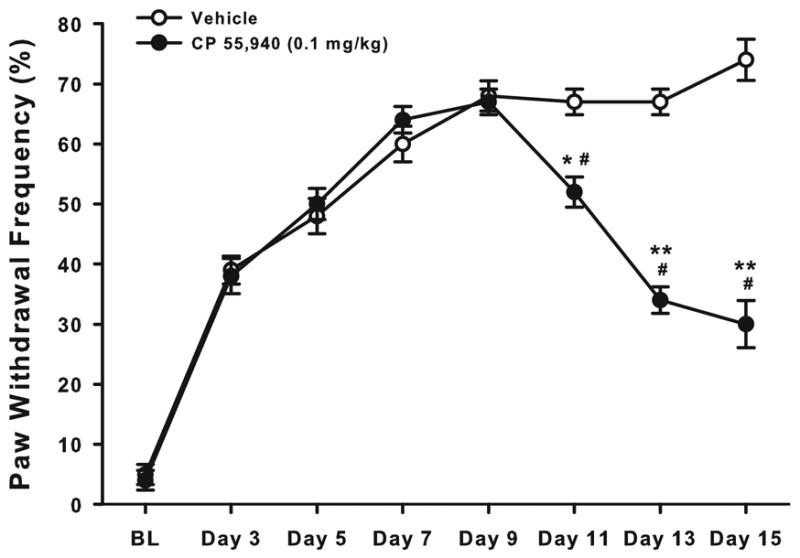
Effect of repeated administration of CP 55,940 on tumor-evoked mechanical hyperalgesia. Mice with tumor-evoked mechanical hyperalgesia received twice-daily administration of CP 55,940 or vehicle beginning on day 9. By the next behavioral testing time (Day 11), mice that received CP 55,940 exhibited less tumor-evoked mechanical hyperalgesia than they did on day 9 and less on day 11 than mice that received vehicle. Repeated administration of CP 55,940 continued to attenuate tumor-evoked mechanical hyperalgesia over the 6.5 day treatment. N=10 mice/group. *P ≤ 0.005; **P ≤ 0.001 for postimplantation days 11, 13, & 15 compared to day 9. #P < 0.001 for CP 55,940 compared to vehicle at days 11, 13, & 15.
CP 55,940 continued to attenuate tumor-evoked mechanical hyperalgesia 30 min after administration of the morning dose for the duration of the study (Fig. 9). On day 9 after implantation of fibrosarcoma cells, the morning dose of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia by ~40% (P < 0.001). Following twice-daily administration of CP 55,940, the morning dose of CP 55,940 continued to attenuate tumor-evoked mechanical hyperalgesia as postdrug paw withdrawal frequencies were lower than predrug levels on days 11 (P < 0.001), 13 (P < 0.001) and 15 (P < 0.005). Thus, repeated administration of CP 55,940 for 6.5 days continued to provide both short-term (i.e., 30 min) and longer term (i.e., days) attenuation of tumor-evoked mechanical hyperalgesia.
Fig. 9.
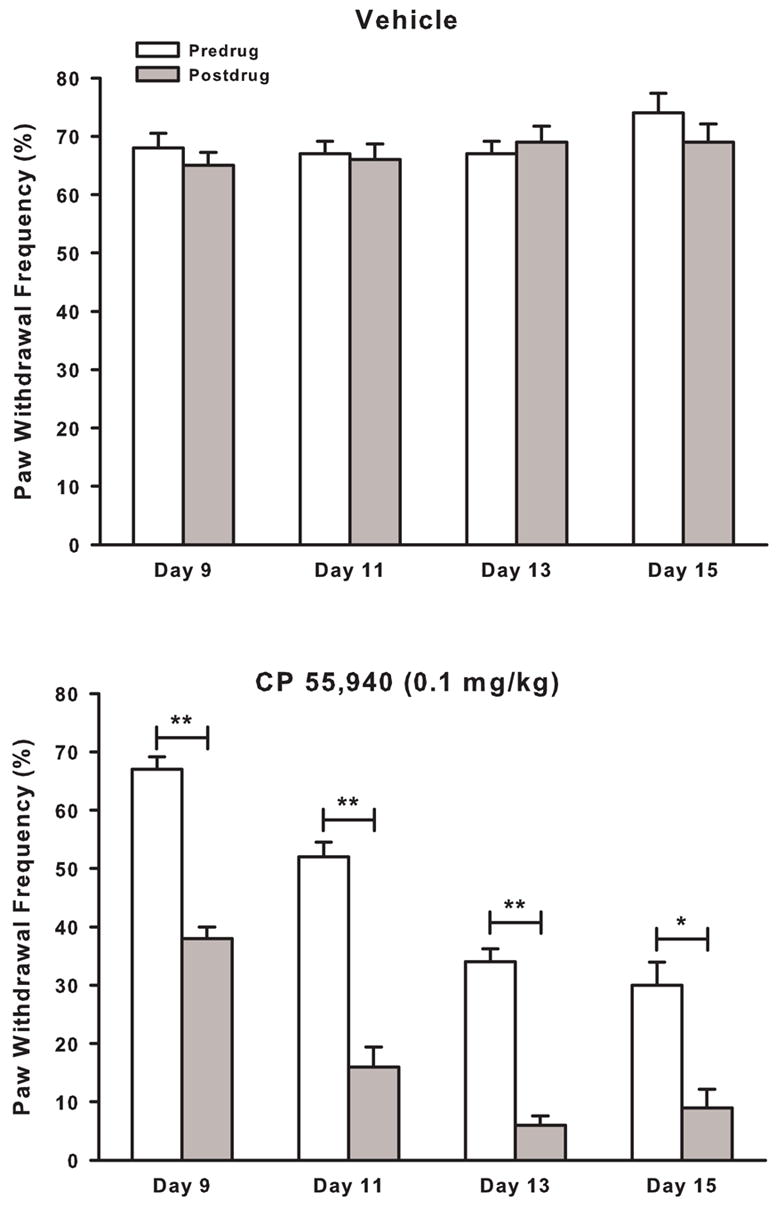
On day 9 after implantation of fibrosarcoma cells, mice exhibited paw withdrawal frequencies of ~70% before administration (i.e., Predrug) of the morning dose of A) vehicle or B) CP 55,940 (0.1 mg/kg). Thirty minutes after administration (i.e., Postdrug), CP 55,940 reduced paw withdrawal frequency. This short-term antihyperalgesic effect of CP 55,940 was also present on days 11, 13, and 15 as CP 55,940 continued to decrease paw withdrawal frequencies 30 min after administration. Thus, CP 55,940 continued to attenuate tumor-evoked mechanical hyperalgesia over the 6.5 day treatment period. *P ≤ 0.005; **P ≤ 0.001 for predrug vs postdrug values.
4. Discussion
These results demonstrate that the cannabinoid, CP 55,940, dose-dependently attenuated tumor-evoked mechanical hyperalgesia in this model of cancer pain. The antihyperalgesic effect of CP 55,940 was not due to its cataleptic effect or its effects on motor coordination. Our studies using the cannabinoid receptor subtype selective antagonists indicate that the antihyperalgesic, antinociceptive, and cataleptic effects of CP 55,940 were mediated by the cannabinoid CB1 and not the cannabinoid CB2 receptor. During the development of tumor-evoked mechanical hyperalgesia and its attenuation by CP 55,490, threshold force for evoking a paw withdrawal was strongly negatively correlated to the frequency of paw withdrawals to a standard force. These methods of assessing mechanical hyperalgesia yielded similar results. Repeated administration of CP 55,940 to mice with established tumor-evoked mechanical hyperalgesia produced two effects. First, the antihyperalgesic effect of CP 55,940 at 30 min was present even after 6 days of twice-daily administrations. Second, repeated administration of CP 55,940 had a longer lasting effect as predrug paw withdrawal frequencies decreased over the 6 days.
Animal models of cancer pain have been developed to examine the mechanisms that underlie tumor-evoked pain and hyperalgesia. Using models in which osteolytic fibrosarcoma cells are implanted into the humerus, femur, tibia, or calcaneous bone in mice, investigators have begun to elucidate the pathophysiological processes by which cancer produces pain (Menendez et al., 2003; Schwei et al., 1999; Wacnik et al., 2001; Wacnik et al., 2000). These models have also been used to evaluate novel approaches for treating cancer pain (Honore et al., 2000; Sabino et al., 2002; Sevcik et al., 2005). We used the model in which fibrosarcoma cells are implanted into and around the calcaneous bone for several reasons. First, this model produces mechanical hyperalgesia that can be easily detected on the plantar surface of the hind paw. The hind paw is perhaps the most common site for testing nociception and hyperalgesia in rodents and therefore, results can be compared to other studies and other models. Second, this model allows examination of the electrophysiological responses of primary afferent fibers in mice with tumor-evoked hyperalgesia. We have determined that implantation of fibrosarcoma cells results in the sensitization of a subpopulation of C-nociceptors that innervate the plantar surface of the hind paw (Cain et al., 2001). Furthermore, the peripheral terminals of primary afferent fibers innervating the skin overlying the tumor show morphological changes such as an increase in epidermal nerve fibers during the early development of tumor-evoked hyperalgesia, up-regulation of purinergic P2X3 receptors on these fibers, and subsequent decrease in epidermal innervation (Gilchrist et al., 2005). Third, transfection of the fibrosarcoma cells with a fluorescent protein allows examination of the morphology of nerve terminals that contact the tumor cells (Wacnik et al., 2005). Finally, this model will allow us to evaluate the effect of peripheral administration of drugs on tumor-evoked hyperalgesia.
Catalepsy is a state associated with rigidity of the extremities (Sanudo-Pena et al., 2000) in which a rodent remains in an externally imposed position (Sanberg et al., 1988). Rigidity of the hind limb may decrease a mouse’s ability to withdraw its hind paw. Thus, catalepsy may confound the results of behavioral studies that assess motor responses to noxious stimuli (Hohmann, 2002; Walker and Huang, 2002).
Our results indicate that the antihyperalgesic effect of CP 55,940 was not due entirely to its cataleptic effect. First, the ED50 for the antihyperalgesic effect of CP 55,940 was ~15-fold lower than for its cataleptic effect. CP 55,940 (0.3 mg/kg) almost completely (92% MPE) reversed tumor-evoked mechanical hyperalgesia but produced only minimal (~ 6 s (9% MPE)) catalepsy. Cannabinoids are more potent in producing antinociception than catalepsy (Little et al., 1988) and were more potent in producing antihyperalgesia in the present study. Second, pretreatment with the dopamine agonist, apomorphine, attenuated the cataleptic effect of CP 55,940 by ~60% but did not attenuate its antihyperalgesic effect. However, dopamine agonists attenuate inflammatory hyperalgesia (Gao et al., 2001) and potentiate cannabinoid antinociception (Carta et al., 1999). Therefore, although the cataleptic effect of CP 55,940 on paw withdrawal may have been blocked by apomorphine, apomorphine may have decreased the frequency of paw withdrawals through its antihyperalgesic effect. However, pretreatment with apomorphine followed by administration of the vehicle for CP 55,940 did not reduce the frequency of paw withdrawals in mice with tumor-evoked mechanical hyperalgesia indicating that apomorphine did not exhibit an antihyperalgesic effect.
Apomorphine did not completely block the cataleptic effect of CP 55,490. The remaining level of catalepsy (i.e., 3.5 s) may have been enough to affect the ability of a mouse to withdraw its hind paw. When we produced a greater level of catalepsy (~20 s) using the dopamine antagonist, spiperone, the frequency of paw withdrawals in mice with tumor-evoked hyperalgesia only decreased by ~35% compared to complete attenuation by CP 55,940. Spiperone administered intrathecally or intracerebroventricularly does not attenuate inflammatory hyperalgesia (Gao et al., 2000, 2001) and systemically administered spiperone does not exhibit antinociceptive effects (Cook et al., 2000). In the present study, spiperone increased tail-flick latencies by ~30% and thus may have also had an antinociceptive effect. Even if the reduction in the frequency of paw withdrawals produced by spiperone was due to its cataleptic effect, catalepsy alone could not account for the antihyperalgesic effect of CP 55,940 as the higher level of catalepsy produced by spiperone did not attenuate tumor-evoked mechanical hyperalgesia to the same level.
Further evidence that catalepsy and motor dysfunction were not the only mechanisms by which CP 55,940 induced the decrease in paw withdrawal frequency is that cannabinoids suppress nociceptive neurotransmission by acting on neurons in both the central and peripheral nervous systems. Cannabinoids decrease noxious stimulus-evoked excitation of nociceptive neurons located in the thalamus (Martin et al., 1996) and spinal cord (Hohmann et al., 1999) and modulate the activity of neurons in the periaqueductal grey (Vaughan et al., 2000) and rostral ventromedial medulla (Meng et al., 1998) associated with descending pain modulation (Lichtman and Martin, 1991). Cannabinoids also attenuate depolarization-evoked increase in intracellular Ca2+ in a subpopulation of primary afferent neurons with myelinated axons (Khasabova et al., 2002), providing a mechanism by which peripheral administration of cannabinoids may attenuate hyperalgesia (Johanek et al., 2001; Johanek and Simone, 2004).
Cannabinoids attenuate hyperalgesia evoked by capsaicin (Li et al., 1999), neuropathy (Bridges et al., 2001; Dyson et al., 2005; Fox et al., 2001; Herzberg et al., 1997) and inflammation (Clayton et al., 2002; Dyson et al., 2005; Jaggar et al., 1998; Martin et al., 1999; Richardson et al., 1998). We previously found that the cannabinoid, WIN 55,212-2, attenuated tumor-evoked hyperalgesia in a murine model of deep-tissue cancer pain (Kehl et al., 2003). However, at the highest dose that we could get into solution, WIN 55,212-2 was only partially (~50%) effective. In the present study, we used CP 55,940 because it has higher affinity (~20-fold) for both cannabinoid receptor subtypes than WIN 55,212-2 (Felder and Glass, 1998), and higher affinity at these receptors correlates well with increased behavioral effects (Compton et al., 1993). Indeed, CP 55,40 is more potent than WIN 55,212-2 in the tail-flick assay (Fox et al., 2001). In contrast to the partial effectiveness of WIN 55,212-2 in the model of tumor-evoked deep tissue hyperalgesia, CP 55,940 completely attenuated tumor-evoked cutaneous mechanical hyperalgesia in the present study. The ED50 of the antihyperalgesic effect of CP 55,940 (0.089 mg/kg; 95% CI=0.064 to 0.124 mg/kg) was ~250-fold lower than the ED50 of the antihyperalgesic effect of WIN 55,212-2 (23.3 mg/kg; 95% CI=13.6 to 40.0 mg/kg) (Kehl et al., 2003). These differences in effectiveness and potency of CP 55,940 and WIN 55,212-2 were likely due to differences in their pharmacologic properties, including binding affinities, as well as differences in characteristics of the models used.
The antihyperalgesic effects of CP 55,940 and WIN 55,212-2 on tumor-evoked hyperalgesia were mediated by the cannabinoid CB1 receptor as the cannabinoid CB1 receptor selective antagonist, SR 141716A, but not the cannabinoid CB2 receptor selective antagonist, SR 144528, completely blocked the antihyperalgesic effect of each cannabinoid receptor agonist. In models of neuropathic pain, the antihyperalgesic effect of cannabinoids is mediated by cannabinoid CB1 receptors (Bridges et al., 2001; Dyson et al., 2005; Fox et al., 2001; Herzberg et al., 1997). However, recent evidence suggests that the cannabinoid CB2 receptor may contribute to the antihyperalgesic effect of cannabinoids in capsaicin-induced hyperalgesia and neuropathic pain (Hohmann et al., 2004; Scott et al., 2004). In models of inflammatory pain, cannabinoid CB1 and CB2 receptors are involved in cannabinoid induced antihyperalgesia (Clayton et al., 2002; Kehl et al., 2003; Nackley et al., 2003; Quartilho et al., 2003). This finding is consistent with the presence of cannabinoid CB2 receptors on immune cells (Munro et al., 1993). Thus, the contribution of cannabinoid CB1 or CB2 receptors to cannabinoid induced antihyperalgesia differs depending upon the etiology of the hyperalgesia (i.e., neuropathic or inflammatory).
Patients with cancer pain may require treatments that provide analgesia over weeks or longer. Thus, we examined whether twice-daily administration of CP 55,940 for 6.5 days would continue to attenuate tumor-evoked mechanical hyperalgesia. Repeated administration of cannabinoids, including CP 55,940, have been shown to produce tolerance to their antinociceptive effects in animal models of acute pain (Gonzalez et al., 2005). However, tolerance to the antihyperalgesic effects of cannabinoids may not occur (De Vry et al., 2004) In the present study, repeated administration of CP 55,940 did not produce tolerance to its antihyperalgesic effect as paw withdrawal frequencies decreased 30 min after administration of CP 55,940 on each day of testing (days 9, 11, 13, and 15). Interestingly, predrug levels of tumor-evoked mechanical hyperalgesia decreased over the treatment period indicating that CP 55,940 also had a longer lasting effect.
There are several possible mechanisms that could underlie the longer lasting antihyperalgesic effect of CP 55,940. First, twice-daily administration could result in a build-up of CP 55,940 in fat stores because cannabinoids are lipid soluble (Agurell et al., 1986). Then, CP 55,940 might be released from the fat stores into the blood stream to keep the serum concentration of CP 55,940 high enough to inhibit excitation of nociceptive neurons. Cannabinoids exhibit antiinflammatory effects through the cannabinoid CB2 receptors located on immune cells (Mbvundula et al., 2004). Repeated administration of CP 55,940 could have reduced the inflammation induced by the fibrosarcoma cells. Finally, CP 55,940 could have attenuated proliferation and/or function of the fibrosarcoma cells (Munson et al., 1975). THC induces apoptosis of tumor cells, reduces tumor burden, and increases survival of mice implanted with lymphoma cells (McKallip et al., 2002). Interestingly, intratumoral administration of THC induces regression of malignant gliomas in vivo (Galve-Roperh et al., 2000). Further studies are needed to determine whether these or other mechanisms underlie the antihyperalgesic effect of repeated administration of CP 55,940.
We administered CP 55,940 systemically and therefore, we do not know whether its antihyperalgesic effects were mediated in the central nervous system or the periphery. Peripheral administration of cannabinoids produces antinociception (Calignano et al., 1998; Fox et al., 2001) and antihyperalgesia (Fox et al., 2001; Johanek et al., 2001; Johanek and Simone, 2004; Ko and Woods, 1999). Because cannabinoids have antiinflammatory and antineoplastic effects, administration of cannabinoids to the site of a tumor may decrease tumor growth and tumor-evoked pain while avoiding the unwanted side-effects associated with activation of cannabinoid receptors in the central nervous system.
In summary, the cannabinoid receptor agonist CP 55,940 attenuated tumor-evoked mechanical hyperalgesia through a mechanism involving the cannabinoid CB1 receptor. The ED50 for the antihyperalgesic effect of systemically administered CP 55,940 was lower than the ED50s for its effects on nociception, catalepsy, and motor function. Finally, repeated administration of CP 55,940 attenuated tumor-evoked mechanical hyperalgesia over the 6.5 day treatment period. Thus, cannabinoids may be clinically useful for treating pain associated with cancer.
Acknowledgments
The authors would like to thank Dr. Carolyn Fairbanks for her technical help with this study. ED50 values were calculated using an Excel macro programmed by Dr. Michael Ossipov. This study was supported by grants from the National Institutes of Health; DA18231 (DTH), DA11471 and CA91007 (DAS).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Agurell S, Halldin M, Lindgren JE, Ohlsson A, Widman M, Gillespie H, Hollister L. Pharmacokinetics and metabolism of delta 1-tetrahydrocannabinol and other cannabinoids with emphasis on man. Pharmacol Rev. 1986;38:21–43. [PubMed] [Google Scholar]
- Ahles TA, Ruckdeschel JC, Blanchard EB. Cancer-related pain--I. Prevalence in an outpatient setting as a function of stage of disease and type of cancer. J Psychosom Res. 1984;28:115–119. doi: 10.1016/0022-3999(84)90003-5. [DOI] [PubMed] [Google Scholar]
- Beck SL, Falkson G. Prevalence and management of cancer pain in South Africa. Pain. 2001;94:75–84. doi: 10.1016/S0304-3959(01)00343-8. [DOI] [PubMed] [Google Scholar]
- Bridges D, Ahmad K, Rice AS. The synthetic cannabinoid WIN55,212-2 attenuates hyperalgesia and allodynia in a rat model of neuropathic pain. Br J Pharmacol. 2001;133:586–594. doi: 10.1038/sj.bjp.0704110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cain DM, Wacnik PW, Turner M, Wendelschafer-Crabb G, Kennedy WR, Wilcox GL, Simone DA. Functional interactions between tumor and peripheral nerve: changes in excitability and morphology of primary afferent fibers in a murine model of cancer pain. J Neurosci. 2001;21:9367–9376. doi: 10.1523/JNEUROSCI.21-23-09367.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calignano A, La Rana G, Giuffrida A, Piomelli D. Control of pain initiation by endogenous cannabinoids. Nature. 1998;394:277–281. doi: 10.1038/28393. [DOI] [PubMed] [Google Scholar]
- Campbell JN, Meyer RA, LaMotte RH. Sensitization of myelinated nociceptive afferents that innervate monkey hand. J Neurophysiol. 1979;42:1669–1679. doi: 10.1152/jn.1979.42.6.1669. [DOI] [PubMed] [Google Scholar]
- Carta G, Gessa GL, Nava F. Dopamine D(2) receptor antagonists prevent delta(9)-tetrahydrocannabinol-induced antinociception in rats. Eur J Pharmacol. 1999;384:153–156. doi: 10.1016/s0014-2999(99)00696-2. [DOI] [PubMed] [Google Scholar]
- Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53:55–63. doi: 10.1016/0165-0270(94)90144-9. [DOI] [PubMed] [Google Scholar]
- Cherny NI. The pharmacologic management of cancer pain. Oncology (Williston Park) 2004;18:1499–1515. [PubMed] [Google Scholar]
- Clayton N, Marshall FH, Bountra C, O’Shaughnessy CT. CB1 and CB2 cannabinoid receptors are implicated in inflammatory pain. Pain. 2002;96:253–260. doi: 10.1016/S0304-3959(01)00454-7. [DOI] [PubMed] [Google Scholar]
- Clohisy DR, Ogilvie CM, Carpenter RJ, Ramnaraine ML. Localized, tumor-associated osteolysis involves the recruitment and activation of osteoclasts. J Orthop Res. 1996;14:2–6. doi: 10.1002/jor.1100140103. [DOI] [PubMed] [Google Scholar]
- Compton DR, Rice KC, De Costa BR, Razdan RK, Melvin LS, Johnson MR, Martin BR. Cannabinoid structure-activity relationships: correlation of receptor binding and in vivo activities. J Pharmacol Exp Ther. 1993;265:218–226. [PubMed] [Google Scholar]
- Cook CD, Barrett AC, Syvanthong C, Picker MJ. Modulatory effects of dopamine D3/2 agonists on kappa opioid-induced antinociception and diuresis in the rat. Psychopharmacology (Berl) 2000;152:14–23. doi: 10.1007/s002130000519. [DOI] [PubMed] [Google Scholar]
- Daut RL, Cleeland CS. The prevalence and severity of pain in cancer. Cancer. 1982;50:1913–1918. doi: 10.1002/1097-0142(19821101)50:9<1913::aid-cncr2820500944>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- De Vry J, Denzer D, Reissmueller E, Eijckenboom M, Heil M, Meier H, Mauler F. 3-[2-cyano-3-(trifluoromethyl)phenoxy]phenyl-4,4,4-trifluoro-1-butanesulfonate (BAY 59–3074): a novel cannabinoid Cb1/Cb2 receptor partial agonist with antihyperalgesic and antiallodynic effects. J Pharmacol Exp Ther. 2004;310:620–632. doi: 10.1124/jpet.103.062836. [DOI] [PubMed] [Google Scholar]
- Dunham NW, Miya TS. A note on a simple apparatus for detecting neurological deficit in rats and mice. J Am Pharm Assoc Am Pharm Assoc (Baltim) 1957;46:208–209. doi: 10.1002/jps.3030460322. [DOI] [PubMed] [Google Scholar]
- Dyson A, Peacock M, Chen A, Courade JP, Yaqoob M, Groarke A, Brain C, Loong Y, Fox A. Antihyperalgesic properties of the cannabinoid CT-3 in chronic neuropathic and inflammatory pain states in the rat. Pain. 2005;116:129–137. doi: 10.1016/j.pain.2005.03.037. [DOI] [PubMed] [Google Scholar]
- Erzin-Waters C, Muller P, Seeman P. Catalepsy induced by morphine or haloperidol: effects of apomorphine and anticholinergic drugs. Can J Physiol Pharmacol. 1976;54:516–519. doi: 10.1139/y76-071. [DOI] [PubMed] [Google Scholar]
- Felder CC, Glass M. Cannabinoid receptors and their endogenous agonists. Ann Rev Pharmacol Toxicol. 1998;38:179–200. doi: 10.1146/annurev.pharmtox.38.1.179. [DOI] [PubMed] [Google Scholar]
- Foley KM. Controlling cancer pain. Hosp Pract (Off Ed) 2000;35:101–108. 111–102. doi: 10.3810/hp.2000.04.193. [DOI] [PubMed] [Google Scholar]
- Fox A, Kesingland A, Gentry C, McNair K, Patel S, Urban L, James I. The role of central and peripheral Cannabinoid1 receptors in the antihyperalgesic activity of cannabinoids in a model of neuropathic pain. Pain. 2001;92:91–100. doi: 10.1016/s0304-3959(00)00474-7. [DOI] [PubMed] [Google Scholar]
- Galve-Roperh I, Sanchez C, Cortes ML, del Pulgar TG, Izquierdo M, Guzman M. Anti-tumoral action of cannabinoids: involvement of sustained ceramide accumulation and extracellular signal-regulated kinase activation. Nat Med. 2000;6:313–319. doi: 10.1038/73171. [DOI] [PubMed] [Google Scholar]
- Gao X, Zhang Y, Wu G. Effects of dopaminergic agents on carrageenan hyperalgesia in rats. Eur J Pharmacol. 2000;406:53–58. doi: 10.1016/s0014-2999(00)00649-x. [DOI] [PubMed] [Google Scholar]
- Gao X, Zhang Y, Wu G. Effects of dopaminergic agents on carrageenan hyperalgesia after intrathecal administration to rats. Eur J Pharmacol. 2001;418:73–77. doi: 10.1016/s0014-2999(01)00930-x. [DOI] [PubMed] [Google Scholar]
- Gilchrist LS, Cain DM, Harding-Rose C, Kov AN, Wendelschafer-Crabb G, Kennedy WR, Simone DA. Re-organization of P2X3 receptor localization on epidermal nerve fibers in a murine model of cancer pain. Brain Res. 2005;1044:197–205. doi: 10.1016/j.brainres.2005.02.081. [DOI] [PubMed] [Google Scholar]
- Gonzalez S, Cebeira M, Fernandez-Ruiz J. Cannabinoid tolerance and dependence: a review of studies in laboratory animals. Pharmacol Biochem Behav. 2005;81:300–318. doi: 10.1016/j.pbb.2005.01.028. [DOI] [PubMed] [Google Scholar]
- Harris LS, Dewey WL, Howes JF, Kennedy JS, Pars H. Narcotic-antagonist analgesics: interactions with cholinergic systems. J Pharmacol Exp Ther. 1969;169:17–22. [PubMed] [Google Scholar]
- Herzberg U, Eliav E, Bennett GJ, Kopin IJ. The analgesic effects of R(+)-WIN 55,212-2 mesylate, a high affinity cannabinoid agonist, in a rat model of neuropathic pain. Neurosci Lett. 1997;221:157–160. doi: 10.1016/s0304-3940(96)13308-5. [DOI] [PubMed] [Google Scholar]
- Hohmann AG. Spinal and peripheral mechanisms of cannabinoid antinociception: behavioral, neurophysiological and neuroanatomical perspectives. Chem Phys Lipids. 2002;121:173–190. doi: 10.1016/s0009-3084(02)00154-8. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Farthing JN, Zvonok AM, Makriyannis A. Selective activation of cannabinoid CB2 receptors suppresses hyperalgesia evoked by intradermal capsaicin. J Pharmacol Exp Ther. 2004;308:446–453. doi: 10.1124/jpet.103.060079. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Tsou K, Walker JM. Cannabinoid suppression of noxious heat-evoked activity in wide dynamic range neurons in the lumbar dorsal horn of the rat. J Neurophysiol. 1999;81:575–583. doi: 10.1152/jn.1999.81.2.575. [DOI] [PubMed] [Google Scholar]
- Honore P, Luger NM, Sabino MA, Schwei MJ, Rogers SD, Mach DB, O’Keefe PF, Ramnaraine ML, Clohisy DR, Mantyh PW. Osteoprotegerin blocks bone cancer-induced skeletal destruction, skeletal pain and pain-related neurochemical reorganization of the spinal cord. Nat Med. 2000;6:521–528. doi: 10.1038/74999. [DOI] [PubMed] [Google Scholar]
- Jaggar SI, Hasnie FS, Sellaturay S, Rice AS. The anti-hyperalgesic actions of the cannabinoid anandamide and the putative CB2 receptor agonist palmitoylethanolamide in visceral and somatic inflammatory pain. Pain. 1998;76:189–199. doi: 10.1016/s0304-3959(98)00041-4. [DOI] [PubMed] [Google Scholar]
- Jemal A, Siegel R, Ward E, Murray T, Xu J, Smigal C, Thun MJ. Cancer statistics, 2006. CA Cancer J Clin. 2006;56:106–130. doi: 10.3322/canjclin.56.2.106. [DOI] [PubMed] [Google Scholar]
- Johanek LM, Heitmiller DR, Turner M, Nader N, Hodges J, Simone DA. Cannabinoids attenuate capsaicin-evoked hyperalgesia through spinal and peripheral mechanisms. Pain. 2001;93:303–315. doi: 10.1016/S0304-3959(01)00336-0. [DOI] [PubMed] [Google Scholar]
- Johanek LM, Simone DA. Activation of peripheral cannabinoid receptors attenuates cutaneous hyperalgesia produced by a heat injury. Pain. 2004;109:432–442. doi: 10.1016/j.pain.2004.02.020. [DOI] [PubMed] [Google Scholar]
- Karst M, Salim K, Burstein S, Conrad I, Hoy L, Schneider U. Analgesic effect of the synthetic cannabinoid CT-3 on chronic neuropathic pain: a randomized controlled trial. Jama. 2003;290:1757–1762. doi: 10.1001/jama.290.13.1757. [DOI] [PubMed] [Google Scholar]
- Kehl LJ, Hamamoto DT, Wacnik PW, Croft DL, Norsted BD, Wilcox GL, Simone DA. A cannabinoid agonist differentially attenuates deep tissue hyperalgesia in animal models of cancer and inflammatory muscle pain. Pain. 2003;103:175–186. doi: 10.1016/s0304-3959(02)00450-5. [DOI] [PubMed] [Google Scholar]
- Khasabova IA, Simone DA, Seybold VS. Cannabinoids attenuate depolarization-dependent Ca2+ influx in intermediate-size primary afferent neurons of adult rats. Neuroscience. 2002;115:613–625. doi: 10.1016/s0306-4522(02)00449-9. [DOI] [PubMed] [Google Scholar]
- Ko MC, Woods JH. Local administration of delta9-tetrahydrocannabinol attenuates capsaicin-induced thermal nociception in rhesus monkeys: a peripheral cannabinoid action. Psychopharmacology (Berl) 1999;143:322–326. doi: 10.1007/s002130050955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuschinsky K, Hornykiewicz O. Morphine catalepsy in the rat: relation to striatal dopamine metabolism. Eur J Pharmacol. 1972;19:119–122. doi: 10.1016/0014-2999(72)90086-6. [DOI] [PubMed] [Google Scholar]
- Li J, Daughters RS, Bullis C, Bengiamin R, Stucky MW, Brennan J, Simone DA. The cannabinoid receptor agonist WIN 55,212-2 mesylate blocks the development of hyperalgesia produced by capsaicin in rats. Pain. 1999;81:25–33. doi: 10.1016/s0304-3959(98)00263-2. [DOI] [PubMed] [Google Scholar]
- Lichtman AH, Martin BR. Spinal and supraspinal components of cannabinoid-induced antinociception. J Pharmacol Exp Ther. 1991;258:517–523. [PubMed] [Google Scholar]
- Little PJ, Compton DR, Johnson MR, Melvin LS, Martin BR. Pharmacology and stereoselectivity of structurally novel cannabinoids in mice. J Pharmacol Exp Ther. 1988;247:1046–1051. [PubMed] [Google Scholar]
- Martin BR, Lichtman AH. Cannabinoid transmission and pain perception. Neurobiol Dis. 1998;5:447–461. doi: 10.1006/nbdi.1998.0218. [DOI] [PubMed] [Google Scholar]
- Martin WJ, Hohmann AG, Walker JM. Suppression of noxious stimulus-evoked activity in the ventral posterolateral nucleus of the thalamus by a cannabinoid agonist: correlation between electrophysiological and antinociceptive effects. J Neurosci. 1996;16:6601–6611. doi: 10.1523/JNEUROSCI.16-20-06601.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin WJ, Loo CM, Basbaum AI. Spinal cannabinoids are anti-allodynic in rats with persistent inflammation. Pain. 1999;82:199–205. doi: 10.1016/S0304-3959(99)00045-7. [DOI] [PubMed] [Google Scholar]
- Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature. 1990;346:561–564. doi: 10.1038/346561a0. [DOI] [PubMed] [Google Scholar]
- Mbvundula EC, Rainsford KD, Bunning RA. Cannabinoids in pain and inflammation. Inflammopharmacology. 2004;12:99–114. doi: 10.1163/1568560041352275. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Lombard C, Fisher M, Martin BR, Ryu S, Grant S, Nagarkatti PS, Nagarkatti M. Targeting CB2 cannabinoid receptors as a novel therapy to treat malignant lymphoblastic disease. Blood. 2002;100:627–634. doi: 10.1182/blood-2002-01-0098. [DOI] [PubMed] [Google Scholar]
- McMillan SC, Tittle M, Hagan S, Laughlin J. Management of pain and pain-related symptoms in hospitalized veterans with cancer. Cancer Nurs. 2000;23:327–336. doi: 10.1097/00002820-200010000-00001. [DOI] [PubMed] [Google Scholar]
- Menendez L, Lastra A, Fresno MF, Llames S, Meana A, Hidalgo A, Baamonde A. Initial thermal heat hypoalgesia and delayed hyperalgesia in a murine model of bone cancer pain. Brain Res. 2003;969:102–109. doi: 10.1016/s0006-8993(03)02284-4. [DOI] [PubMed] [Google Scholar]
- Meng ID, Manning BH, Martin WJ, Fields HL. An analgesia circuit activated by cannabinoids. Nature. 1998;395:381–383. doi: 10.1038/26481. [DOI] [PubMed] [Google Scholar]
- Miser AW, McCalla J, Dothage JA, Wesley M, Miser JS. Pain as a presenting symptom in children and young adults with newly diagnosed malignancy. Pain. 1987;29:85–90. doi: 10.1016/0304-3959(87)90181-3. [DOI] [PubMed] [Google Scholar]
- Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature. 1993;365:61–65. doi: 10.1038/365061a0. [DOI] [PubMed] [Google Scholar]
- Munson AE, Harris LS, Friedman MA, Dewey WL, Carchman RA. Antineoplastic activity of cannabinoids. J Natl Cancer Inst. 1975;55:597–602. doi: 10.1093/jnci/55.3.597. [DOI] [PubMed] [Google Scholar]
- Nackley AG, Makriyannis A, Hohmann AG. Selective activation of cannabinoid CB(2) receptors suppresses spinal fos protein expression and pain behavior in a rat model of inflammation. Neuroscience. 2003;119:747–757. doi: 10.1016/s0306-4522(03)00126-x. [DOI] [PubMed] [Google Scholar]
- Noyes R, Jr, Brunk SF, Avery DAH, Canter AC. The analgesic properties of delta-9-tetrahydrocannabinol and codeine. Clin Pharmacol Ther. 1975;18:84–89. doi: 10.1002/cpt197518184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pecherstorfer M, Vesely M. Diagnosis and monitoring of bone metastases: clinical means. In: Body J-J, editor. Tumor Bone Diseases and Osteoporosis in Cancer Patients. Marcel Dekker, Inc; New York, NY: 2000. pp. 97–129. [Google Scholar]
- Pertwee RG. Cannabinoid receptors and pain. Prog Neurobiol. 2001;63:569–611. doi: 10.1016/s0301-0082(00)00031-9. [DOI] [PubMed] [Google Scholar]
- Portenoy RK. Cancer pain. Epidemiology and syndromes Cancer. 1989;63:2298–2307. doi: 10.1002/1097-0142(19890601)63:11<2298::aid-cncr2820631140>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- Quartilho A, Mata HP, Ibrahim MM, Vanderah TW, Porreca F, Makriyannis A, Malan TP., Jr Inhibition of inflammatory hyperalgesia by activation of peripheral CB2 cannabinoid receptors. Anesthesiology. 2003;99:955–960. doi: 10.1097/00000542-200310000-00031. [DOI] [PubMed] [Google Scholar]
- Richardson JD, Kilo S, Hargreaves KM. Cannabinoids reduce hyperalgesia and inflammation via interaction with peripheral CB1 receptors. Pain. 1998;75:111–119. doi: 10.1016/S0304-3959(97)00213-3. [DOI] [PubMed] [Google Scholar]
- Rubens RD. Bone metastases--the clinical problem. Eur J Cancer. 1998;34:210–213. doi: 10.1016/s0959-8049(97)10128-9. [DOI] [PubMed] [Google Scholar]
- Sabino MA, Ghilardi JR, Jongen JL, Keyser CP, Luger NM, Mach DB, Peters CM, Rogers SD, Schwei MJ, de Felipe C, Mantyh PW. Simultaneous reduction in cancer pain, bone destruction, and tumor growth by selective inhibition of cyclooxygenase-2. Cancer Res. 2002;62:7343–7349. [PubMed] [Google Scholar]
- Sanberg PR, Bunsey MD, Giordano M, Norman AB. The catalepsy test: its ups and downs. Behav Neurosci. 1988;102:748–759. doi: 10.1037//0735-7044.102.5.748. [DOI] [PubMed] [Google Scholar]
- Sanudo-Pena MC, Romero J, Seale GE, Fernandez-Ruiz JJ, Walker JM. Activational role of cannabinoids on movement. Eur J Pharmacol. 2000;391:269–274. doi: 10.1016/s0014-2999(00)00044-3. [DOI] [PubMed] [Google Scholar]
- Schwei MJ, Honore P, Rogers SD, Salak-Johnson JL, Finke MP, Ramnaraine ML, Clohisy DR, Mantyh PW. Neurochemical and cellular reorganization of the spinal cord in a murine model of bone cancer pain. J Neurosci. 1999;19:10886–10897. doi: 10.1523/JNEUROSCI.19-24-10886.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott DA, Wright CE, Angus JA. Evidence that CB-1 and CB-2 cannabinoid receptors mediate antinociception in neuropathic pain in the rat. Pain. 2004;109:124–131. doi: 10.1016/j.pain.2004.01.020. [DOI] [PubMed] [Google Scholar]
- Sevcik MA, Ghilardi JR, Peters CM, Lindsay TH, Halvorson KG, Jonas BM, Kubota K, Kuskowski MA, Boustany L, Shelton DL, Mantyh PW. Anti-NGF therapy profoundly reduces bone cancer pain and the accompanying increase in markers of peripheral and central sensitization. Pain. 2005;115:128–141. doi: 10.1016/j.pain.2005.02.022. [DOI] [PubMed] [Google Scholar]
- Vaughan CW, Connor M, Bagley EE, Christie MJ. Actions of cannabinoids on membrane properties and synaptic transmission in rat periaqueductal gray neurons in vitro. Mol Pharmacol. 2000;57:288–295. [PubMed] [Google Scholar]
- Wacnik PW, Baker CM, Herron MJ, Kren BT, Blazar BR, Wilcox GL, Hordinsky MK, Beitz AJ, Ericson ME. Tumor-induced mechanical hyperalgesia involves CGRP receptors and altered innervation and vascularization of DsRed2 fluorescent hindpaw tumors. Pain. 2005;115:95–106. doi: 10.1016/j.pain.2005.02.024. [DOI] [PubMed] [Google Scholar]
- Wacnik PW, Eikmeier LJ, Ruggles TR, Ramnaraine ML, Walcheck BK, Beitz AJ, Wilcox GL. Functional interactions between tumor and peripheral nerve: morphology, algogen identification, and behavioral characterization of a new murine model of cancer pain. J Neurosci. 2001;21:9355–9366. doi: 10.1523/JNEUROSCI.21-23-09355.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wacnik PW, Wilcox GL, Clohisy DR, Ramnaraine ML, Eikmeier LJ, Beitz AJ. Cancer pain mechanisms and animal models of cancer pain. In: Devor M, Rowbotham MC, Wiesenfeld-Hallin Z, editors. Proceedings of the 9th World Congress on Pain. Vol. 16. IASP Press; Seattle, WA: 2000. pp. 615–637. [Google Scholar]
- Walker JM, Hohmann AG, Martin WJ, Strangman NM, Huang SM, Tsou K. The neurobiology of cannabinoid analgesia. Life Sci. 1999;65:665–673. doi: 10.1016/s0024-3205(99)00289-1. [DOI] [PubMed] [Google Scholar]
- Walker JM, Huang SM. Cannabinoid analgesia. Pharmacol Ther. 2002;95:127–135. doi: 10.1016/s0163-7258(02)00252-8. [DOI] [PubMed] [Google Scholar]
- Yurek DM, Randall PK. Simultaneous catalepsy and apomorphine-induced stereotypic behavior in mice. Life Sci. 1985;37:1665–1673. doi: 10.1016/0024-3205(85)90293-0. [DOI] [PubMed] [Google Scholar]
- Zimmermann M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain. 1983;16:109–110. doi: 10.1016/0304-3959(83)90201-4. [DOI] [PubMed] [Google Scholar]