

Abstract
Cycloheximide (CyX), a toxic antibiotic with a unique chemical structure generated by the actinomycete, Streptomyces griseus, has emerged as a primary focus of studies on mammalian bitter taste. Rats and mice avoid it at concentrations well below the thresholds for most bitter stimuli and T2R G-protein-coupled receptors specific for CyX with appropriate sensitivity are identified for those species. Like mouse and rat, golden hamsters, Mesocricetus auratus, also detected and rejected micromolar levels of CyX, although 1 mM CyX failed to activate the hamster chorda tympani nerve. Hamsters showed an initial tolerance for 500 μM CyX, but after that, avoidance of CyX dramatically increased, plasticity not reported for rat or mouse. As the hamster lineage branches well before division of the mouse-rat lineage in evolutionary time, differences between hamster and mouse-rat reactions to CyX are not surprising. Furthermore, unlike hamster LiCl-induced learned aversions, the induced CyX aversion neither specifically nor robustly generalized to other non-ionic bitter stimuli; and unlike adverse reactions to other chemosensory stimuli, aversions to CyX were not mollified by adding a sweetener. Thus, CyX is unlike other bitter stimuli. The gene for the high-affinity CyX receptor is a member of a cluster of 5 orthologous T2R genes that are likely rodent specific; this “CyX clade” is found in the mouse, rat and probably hamster, but not in the human or rabbit genome. The rodent CyX-T2R interaction may be one of multiple lineage-specific stimulus-receptor interactions reflecting a response to a particular environmental toxin. The combination of T2R multiplicity, species divergence and gene duplication results in diverse ligands for multiple species-specific T2R receptors, which confounds definition of ‘bitter’ stimuli across species.
1. Introduction
Bitter taste is difficult to study because many compounds with diverse chemical structures are bitter and species differences are great. Discovery of a set of about 30, appropriately localized, T2R candidate bitter receptors [1] inspired investigation of T2R structure, function and evolution; but it soon became evident that T2R species differences remained problematic [17, 94].
Cycloheximide (CyX), an antibiotic produced by Streptomyces griseus, is a rodent repellent, “rats preferring to die from thirst” rather than ingest a 15-μM CyX solution [50]. However, determinants of rodent behavior toward CyX, a compound with a unique structure (Fig. 1), are poorly defined. Like many aversive “taste” stimuli [31, 97], CyX has multiple chemosensory effects and its intake leads to a complex of systemic consequences [10, 57, 84]. Nevertheless, CyX, used extensively experimentally to disrupt protein synthesis [115], has become a focus of work on taste transduction and coding mechanisms for bitter taste [19, 20, 42, 52, 74,113]. This is, in part, because CyX sensitivity, in contrast to the human bitter prototype quinine, has been associated with a single gene, likely at the chromosome 6 Cyx locus [68], coding for a specific T2R receptor in mice [20, 74, 79] and an orthologous gene in rats [17]. Practically tasteless to humans, CyX is avoided by rats and mice even at concentrations less than 1 μM, much lower than thresholds for ‘typical’ bitter stimuli [11, 12, 68, 101]. Also, CyX, very toxic to rats, is much less toxic to several mouse species [102,106]. Species differences are useful for establishing identities of specific bitter-stimulus receptors identified with lineage-specific T2R genes [94], as well as providing insight into the evolutionary significance of diverse behaviors toward toxic substances.
Figure 1.
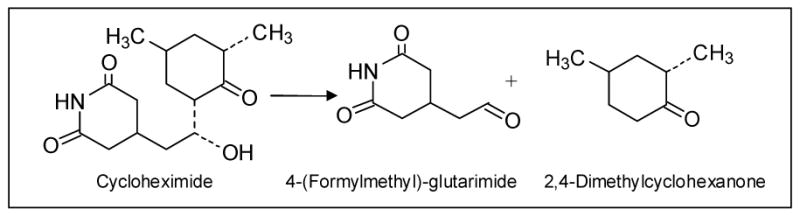
Cycloheximide Decomposition in Alkaline Solution. The major route of decomposition is shown. In neutral aqueous solution, cycloheximide is initially non-odorous to humans. It develops a detectable ketone-like odor similar to its breakdown products within 24 hrs (unpublished observations).
The gustatory system of golden hamsters, Mesocricetus auratus, is well characterized [5, 9, 35, 40, 64], yet hamster reactions to CyX are unknown. We took multiple approaches: behavioral, electrophysiological and comparative genomic, to appraise CyX as a bitter gustatory stimulus in golden hamsters.
Forty-eight hour, 2-bottle tests are used to test behavioral reactions to taste stimuli, although daily intake of an aversive compound cannot itself identify a gustatory stimulus because of the potential influence of post-ingestive effects on long-term intake. However, 2-bottle long-term tests and short-access tests reveal micromolar CyX aversion thresholds for other rodent species; but hamsters' CyX behavioral thresholds are unknown. Hamsters' 2-bottle aversion thresholds for other bitter stimuli [31], as well as hamsters' increased bitter-stimulus acceptability with addition of a sweetener [87] are documented. Amelioration of CyX aversions with sweetening is untested.
Unexpectedly, unlike rats or mice, hamsters initially ingested significant amounts of CyX, which made it possible to test whether CyX exposure and intake reduces subsequent intake of CyX and other stimuli [30, 36]. Many bitter compounds have systemic effects post-ingestion leading to increased behavioral sensitivity. Intake or intraperitoneal injection of CyX may induce learned aversions [8, 57] or, possibly, enhance sensitivity genetically by up-regulating expression of chemosensory receptors [103]. Well-studied LiCl-induced learned aversions are not quickly forgotten [75,76], and hamsters do not generalize LiCl-induced aversions learned to caffeine or SOA to the ionic bitter stimuli: quinine, denatonium or MgSO4 [31]. Neither the induction of CyX-aversions nor the specificity of generalizations of CyX-induced aversions to other stimuli has been tested.
Taste buds on the anterior mammalian tongue are innervated by the chorda tympani nerve (CT), which may specialize in taste quality discrimination [92, 98]. Taste buds on the posterior tongue, innervated by the glossopharyngeal nerve (GL), are more involved in oral reflexive behaviors [58]. Sweet, salty, sour and ionic bitter stimuli activate the hamster CT [30, 31]. However, non-ionic stimuli such as caffeine and sucrose octaacetate (SOA), bitter to humans and aversive to hamsters, fail to activate the CT [31]. It is unknown whether the hamster CT responds to non-ionic CyX or n-propylthiouracil (PROP), an identified human T2R ligand with dimorphic bitter taste [16].
The CyX-sensitive receptor protein falls among rapidly evolving T2R receptors [17, 93, 94]. Species differences in CyX-sensitive and other T2R-receptors need to be evaluated to make sense of bitter taste. Gene and protein sequences for hamster, human, rat, mouse and rabbit are available in NCBI and Ensembl databases for genomic analysis. Synonymous codon substitutions, functioning as molecular evolutionary clocks within orthologous genes, can reveal how closely species are related to each other, and protein sequences allow assessment of T2R species diversity. CyX receptor evolution can be better characterized by comparing rat–mouse and human–mouse structural similarities in T2R receptor proteins, more slowly evolving T1R receptor proteins and other highly “conserved” proteins. Finally, study of sequence similarities of mouse and rat CyX receptors, members of species-specific clades of 5 paralogous receptors [93], and orthologous receptors of humans and rabbits will help identify associations between T2Rs and species-distinctive behaviors toward CyX and other bitter stimuli.
Our data on CyX in hamsters and associated behavioral, physiological and genomic data for other species reveal fundamental species diversity in bitter taste and other determinants of dislike that have evolved to protect mammals against toxins, and support the view that all bitter-tasting compounds do not elicit a “unitary” bitter quality across mammals.
2. Materials and Methods
2.1 Subjects
Male LVG golden hamsters (Mesocricetus auratus), purchased from Charles River Laboratories, weighing >100 g were the subjects of all experiments. A total of 12 were used for 2-bottle testing, 6 for nerve recordings, 28 to measure effects of multiple 1-hr presentations of 500 μM CyX on subsequent intake of products of NaOH-treated CyX, and 29 to measure effects of a 1-hr presentation of 500 μM CyX on subsequent 1-hr intake of a variety of solutions. At 2 months of age, CyX study animals weighed 117 ± 7 g (mean ±SD), at 3 months subsequent to intake of CyX they weighed 129 ± 7 g; thus, they did not lose weight due to CyX intake. The nocturnal rodents were maintained and tested in individual cages in a 21-22°C room on a reverse daylight cycle: lights off: 7 am, lights on: 7 pm. The University of Connecticut Health Center's Animal Care Committee approved all protocols.
2.2. Two-bottle 48-hour intake: Stimuli and preference-aversion
48-hr, 2-bottle intake tests on 5 to 8 hamsters were conducted to establish stimulus preference vs. water without water deprivation. Stimuli tested were CyX (0.3, 3, 30 μM) and 2 other aversive stimuli: PROP (1, 5 mM) and chlorhexidine (0.2 mM) (Sigma, St Louis MO USA). Chlorhexidine is a bitter-tasting bis-biguanide antiseptic with taste-altering properties in humans [14, 34]. Binary mixtures of equally aversive 30 μM CyX or 0.2 mM chlorhexidine with the highly preferred 5 mM dulcin (Sigma, St Louis MO USA), a sweetener with properties quite similar to sucrose in hamsters [33], were also tested to see if adding a sweetener decreases negative reactions to CyX and chlorhexidine as it does other aversive stimuli [87]. Finally, NaOH-treated CyX (for description of treatment, see section 2.4, below) was tested to determine any effect of a fragrant ketone breakdown product: 2, 4-dimethylcyclohexanone (Fig. 1) on intake behavior. Positions of water and stimulus bottles were exchanged after 24 hrs. 2-day intake of stimulus and water and percent preference scores (100 × stimulus intake (mL) ÷ (stimulus + water intake (mL)) were computed. Statistical analysis used t-tests (stimulus vs. water intake) and analysis of variance (ANOVA) with Neuman-Keuls (NK) tests for post hoc analysis; α was set at .05.
2.3. Nerve recordings: Surgery, electrophysiology and stimuli
Hamsters (N = 6) were injected intraperitoneally with sodium pentobarbital (Abbot Labs, N. Chicago IL USA) to establish and maintain a surgical level of anesthesia (initial dose: 80 mg/kg, maintenance doses: 40 mg/kg) and sacrificed at the end of an experiment. Body temperature (∼ 37°C) was maintained with a Deltaphase® isothermal pad. A tracheal cannula was inserted and the hypoglossal nerve bilaterally transected to prevent inadvertent tongue movement. The hamster's head was imobilized with a head holder, the right CT nerve exposed using a mandibular approach [29] and cut where it exits the tympanic bulla. A desheathed nerve was placed on a nichrome-wire recording electrode and an indifferent electrode positioned in nearby tissue.
Neural activity was differentially amplified, rectified, integrated (200 ms time-constant), digitized with a Cambridge Electronic Design (CED) Micro 1401 II analog to digital converter (Cambridge UK) and displayed on a PC. Ascending concentration series (0.01, 0.1, 1.0, 10 mM) of CyX, PROP, and quinine hydrochloride (Sigma, St Louis MO USA) were applied to the anterior portion of the tongue via gravity flow at a rate of 1.5 – 2.0 mL/sec. Responses were quantified as 5-sec post-stimulus minus 5-sec pre-stimulus areas under the integrated response curves using CED's Spike2 (v.5.12) software. Consistency of nerve preparations was monitored with responses to 500 mM NH4Cl (Sigma, St Louis MO USA) applied at least once at the beginning and end of each concentration series. The NH4Cl response was also used as a standard to normalize data across nerves. Relative response magnitudes (percent of NH4Cl response) were statistically evaluated using repeated-measures ANOVAs for 4 concentrations of each compound, with α set at .05. Subsequent analyses used NK tests.
2.4. Cycloheximide exposure: Test stimuli and measuring 1-hour, 1-bottle intake
Before starting 1-bottle testing, hamsters were accustomed to obtaining all of their daily fluid from one bottle in two 1-hr periods: a 10 to 11 am testing period and a 4 to 5 pm hydration period, when they were always presented water.
To test effects of an initial exposure to 500 μM CyX on subsequent intake, each of 28 animals was presented with 500 μM CyX for the first time for 1 hr on day 0 (Table 1). The animals were divided into 3 subgroups in which day 0 was offset by 0 (N =12), 1(N = 8) or 2 (N = 8) weeks to allow use of some animals for daily water-intake normalization (see below). To establish the effect of a first exposure to CyX on subsequent CyX intake with the passage of time, intake for a 2nd 1-hr presentation of CyX at 1-2 (N = 10), 7 (N = 6), 15 (N = 6), and 22 (N = 6) days post-exposure was measured in separate subsets of animals. Intake on a 3rd 1-hr CyX presentation on days 23 to 50 (N = 23), and a 4th CyX presentation on days 43 to 50 (N = 8) were also measured (Table 1).
Table 1.
Schedule for 1-hour intake testing in cycloheximide (CyX) exposure study
| TEST STIMULI | CyX | P0ST-CyX TESTING | |||||
|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | ||||
| Day 0 | Day 1-2 | Day 7 | Day 15 | Day 22 | Day 23+ | Day 43+ | |
| Cycloheximide | X | X | X | X | X | X | X |
| Water | X | ||||||
| Quinine | X | ||||||
| Caffeine | X | ||||||
| SOA | X | ||||||
| PROP | X | ||||||
| Sucrose | X | ||||||
| NaCl | X | ||||||
X: Test stimulus (TS) timing.
Day 0: 28 hamsters presented with CyX to drink for 1st time for 1 hr.
Day 1-2 column: Each hamster gets 2 TS; if not tested with CyX get later 2nd CyX test.
CyX test row: Subgroups of 28 hamsters get a 2nd test on day 1-2 (N = 10) or later days (7, 15, 22, Ns = 6); 23 hamsters get a 3rd test, and 8 a 4th test.
Water intake was measured as a control on each day of testing.
Days 1 and 2 post-CyX exposure were used to test CyX effects on 1-hr intake of a variety of compounds besides CyX, non-bitter stimuli: 0.1 M NaCl and 0.1 M sucrose (Baker, Phillipsburg NJ USA); non-ionic bitter stimuli: 1 mM SOA, 5 mM PROP and 30 mM caffeine; ionic-bitter stimulus: 1 mM quinine·HCl (Sigma, St Louis MO USA) and water (Table 1). On day 1, each animal was presented with one test solution; N = 5 for water and CyX; N = 3 for the other 6 stimuli. On day 2, each animal was offered a different solution from the one offered on day 1, doubling N for each stimulus. Day 2 stimulus identity was counterbalanced. Before the first CyX exposure on day 0, baseline, 1-hr intake of the 6 test solutions and water, was measured in a counterbalanced order.
A 2nd group of 28 animals was used to compare 1-hr intake of 500 μM CyX to 1-hr intake of CyX breakdown products (see section 2.2, above) after 2 or more presentations of 500 μM CyX. To obtain the breakdown products, cycloheximide (28 mg, 0.1 mmole) was dissolved in 1 mL of 1 M NaOH, then heated at 100°C for 5 min. After neutralization with 1 mL of 1 M HCl to pH 6.3, the solution was diluted with water to give 200 mL. A ketone odor was noted. The resulting solution is equivalent to the original 500 μM cycloheximide + 5 mM NaCl. Ten animals were tested with CyX multiple times before being tested with the breakdown products; 18 animals were used to establish same-day water intakes.
Intakes (mL) of test stimuli (TS) were normalized to mean water (H2O) intake on the day of solution presentation for analyses of TS intake. The TS intake/H2O ratio (TS intake ÷ mean H2O intake) is less than 1 for aversive compounds. The TS post/pre-CyX intake ratio (post-CyX TS intake/H2O ratio ÷ mean pre-CyX TS intake/H2O ratio), used to assess effects of CyX exposure on TS intake, is less than 1 if CyX exposure suppressed intake below mean baseline intake. Mean percent suppression of TS intake was computed as 100 × (1 – post/pre-CyX) [36, 83]. Statistical analysis of TS intake/H2O and post/pre-CyX intake ratios used t-tests and ANOVA with NK post-hoc testing; α was set at .05.
2.5. T2R receptor species diversity; hamster molecular clock
Protein and nucleotide sequences of 10 sets of orthologous genes representing a range of evolutionary diversity and function in mouse, rat, human and hamster were examined. Table 2 lists the 10 proteins and NCBI genomic database accession numbers. The proteins were put in four groups: 1) 2 representative T2R bitter receptors (mouse CyX receptor mT2R5 and human hT2R1, ligand unknown) [1, 20, 93]; 2) the 3 T1R amino acid (umami)/ sweet receptors (T1R1, T1R2 and T1R3) [114]; 3) 2 cytochrome P450 enzymes (human hCyp1a2 and polymorphic human hCyp2d6, both involved in metabolism of endogenous substrates and xenobiotics) [77]; and 4) 3 other proteins (a G-protein coupled, inwardly rectifying K+ channel protein (GIRK): KCNJ6, involved in neural excitability [70]; the olfactory receptor OR I7 [2, 112], a member of a large G-protein coupled receptor (GPCR) family with molecular domains similar to bitter receptors; and metabotropic glutamate GPCR mGluR1 [46], with molecular domains similar to candidate sweet receptors). We chose receptors for which candidate human, mouse and rat orthologues could be identified (see below) to assess species diversity for the 4 groups of proteins. Group 1 was expected to show the most species diversity [94] and group 4 was expected to be the most conserved across species [60, 69]. The genome, mostly sequenced for human, rat and mouse, is hardly sequenced for the golden hamster. The 2 cytochrome P450s and K+ channel sequences, available for the hamster, allowed us to estimate evolutionary relationships between hamsters and other species; percent synonymous codon substitutions in orthologous proteins serve as a molecular clock.
Table 2.
Comparison of Protein and Nucleotide Sequences in Genomic Database.
| Protein | Species | NCBI Accession Number | Amino Acids Compared |
|---|---|---|---|
| 1) T2R Bitter Receptors | |||
| mT2R5 | Mouse | AF227147 | - |
| rT2R9 | Rat | AF227146 | 300 |
| hT2R10 | Human | NM_023921 | 300 |
| mT2R19 | Mouse | AF227149 | - |
| rT2R1 | Rat | AF227140 | 335 |
| hT2R1 | Human | AF227129 | 299 |
| 2) T1R Taste Receptors | |||
| T1R1 | Mouse | NM_031867 | - |
| T1R1 | Rat | XM_342986 | 842 |
| T1R1 | Human | BK000153 | 842 |
| T1R2 | Mouse | NM_031873 | - |
| T1R2 | Rat | XM_342960 | 755 |
| T1R2 | Human | BK000151 | 843 |
| T1R3 | Mouse | NM_031872 | - |
| T1R3 | Rat | NM_130818 | 858 |
| T1R3 | Human | BK000152 | 861 |
| 3) Cytochrome P450s | |||
| Cyp1a2 | Mouse | NM_009993 | - |
| Cyp1a2 | Rat | NM_012541 | 513 |
| Cyp1a2 | Hamster | D10252 | 513 |
| Cyp1a2 | Human | NM_000761 | 515 |
| Cyp2d26 | Mouse | NM_029562 | - |
| Cyp2d2 | Rat | M22330 | 500 |
| Cyp2d20 | Hamster | AB031864 | 500 |
| Cyp2d6 | Human | NM_000106 | 500 |
| 4) Other Channels/Receptors | |||
| KCNJ6 | Mouse | NM_010601 | - |
| KCNJ6 | Rat | NM_013192 | 425 |
| KCNJ6 | Hamster | U21937 | 425 |
| KCNJ6 | Human | NM_002240 | 423 |
| OR-I7 | Mouse | AF106007 | - |
| OR-I7 | Rat | M64386 | 327 |
| OR-I7 | Human | NM_003696 | 327 |
| mGluR1 | Mouse | NM_016976 | - |
| mGluR1 | Rat | NM_017011 | 1200 |
| mGluR1 | Human | NM_000838 | 1202 |
Sequences of each species were compared to mouse sequences thereby estimating diversity compared to a mouse standard. The proteins varied in size from ∼300 (OR and T2Rs) to ∼1200 (mGluR1) amino acids. Sequence data, obtained from the NCBI database, were aligned pair-wise using the BLASTp and BLASTn programs on the NCBI website. Amino acid (AA) and codon comparisons were performed with an Excel program. Genes were recognized as orthologous when “best hits” were reciprocal and proportions of synonymous codon substitutions were similar to those of other orthologous proteins for the compared species.
Mixed ANOVAs with subsequent NK tests were used for statistical analysis of percent rat-mouse, hamster-mouse and human-mouse AA identities in a protein sequence and percent rat-mouse, hamster-mouse and human-mouse synonymous substitutions in nucleotide sequences for conserved AA; α was set at .05.
Throughout the text we use the nomenclature of Shi and Zhang [93] who first described gene clusters containing the candidate genes for CyX receptors. In parentheses, below, is the nomenclature for the coded receptors currently endorsed by the Mouse Genome Database (MGD, http://www.informatics.jax.org/). BLASTp and BLASTn were used to examine proteins and genes for the 2 syntenic clusters of the 5 mouse genes, mT2R5 (mTas2r105), mT2R43 (mTas2r107), mT2R44 (mTas2r106), mT2R45 (mTas2r104) and mT2R46 (mTas2r114); and their respective rat orthologues, rT2R9 (rTas2r105), rT2R4 (rTas2r10), rT2R19 (rTas2r19), rT2R21 (rTas2r21) and rT2R5 (rTas2r5)) [93], and to search the Ensembl rabbit (Oryctolagus cuniculus) database for mT2R5 orthologues.
3. Results
3.1. 2-bottle intake of cycloheximide and other aversive stimuli
Results for 2-bottle tests with CyX, chlorhexidine, dulcin and PROP are ordered from left to right in Fig. 2. Hamster 2-day intake of each of 4 pure CyX solutions and water are to the left. They appear in the figure in the order that they were presented to the hamsters. The first presentation of 30 μM CyX yielded a day 1, 38.6 ± 7.6 % (p = .10) and a day 2, 24.8 ± 9.0 % (p = .015) % preference; the 2-day, 28.9 ± 3.1 % preference is plotted. 2-day, % preference scores for the second and subsequent CyX presentations, consistent on days 1 and 2, were for 0.3 μM: 36.4 ± 4.8 % (p =.05), for 3 μM: 19.5 ± 4.5 % (p = .01) and for 30 μM: 17.3 ± 6.0 % (p = .01). A within-subjects ANOVA, F (1,7) = 13.3, p = .01, revealed that each CyX concentration in the series was less palatable than water (NK p < .001). ANOVA also revealed that % preference scores dropped with concentration, F (3, 14) = 6.96, p =.008: 0.3 μM was less aversive (NK p = .01) than equally aversive 3 μM and 30 μM CyX. Thus, the CyX aversion threshold falls near 0.3 μM and the adverse reaction to CyX plateaus around 3 μM in hamsters with previous CyX experience.
Figure 2.
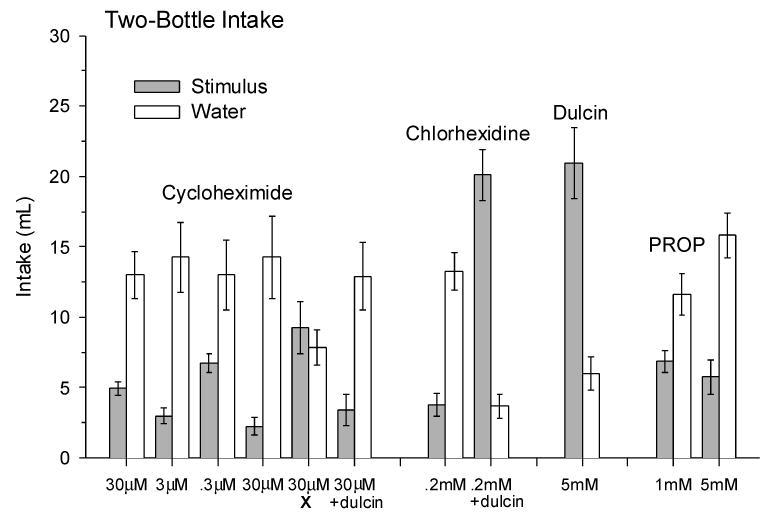
Two-Day, 2-Bottle Mean Intake (±se) of Cycloheximide (CyX), Chlorhexidine, Dulcin and n-Propylthiouracil (PROP). CyX solutions include pure 0.3 - 30 μM CyX, as well as 30 μM CyX treated with NaOH (X), and 30 μM CyX + dulcin. The data for the pure CyX solutions are plotted in the order that they were presented; i.e., the leftmost 30 μM CyX bar represents the hamsters' initial access to CyX. Other solutions are 0.2 mM chlorhexidine, the 0.2 mM chlorhexidine + dulcin mixture, 5 mM dulcin, and 1 mM and 5 mM PROP. Means ± se are plotted. 2-day water and stimulus intake differed for all stimuli except alkali-treated 30 μM cycloheximide X.
The final two sets of bars under “Cycloheximide” in Fig. 2 show hamsters were indifferent to odorous dimethylcyclohexanone (Fig. 1) and/or other breakdown products of alkalization of CyX (X), 52.6 ± 7.5 % preference; and hamsters' adverse reaction to 30 μM CyX, 21.7 ± 9.1 % preference, was not ameliorated by adding 5 mM dulcin (+ dulcin), 23.7 ± 9.9 % preference. The 2 sets of bars under “Chlorhexidine” in Fig. 2 show adding dulcin to 0.2 mM chlorhexidine, 22.1 ± 4.3 % preference, t (7) = 5.44, p = .001, created a more palatable mixture, 84.2 ± 3.4% preference, t (7) = 7.65, p = .0001. Dulcin, itself, was highly preferred, 77.1 ± 5.0 %, t (7) = 5.15, p = .001. Thus, CyX odorous breakdown products were neutral to hamsters and, unlike equi-aversive chlorhexidine, CyX palatability failed to increase when mixed with a sweetener.
Finally, the two sets of bars at the right in Fig. 2 show hamsters found 1 mM PROP, 38.3 ± 5.1 % preference, t (7) = 2.42, p = .05, and 5 mM PROP, 25.8 ± 2.7 % preference, t (7) = 6.66, p = .0003, increasingly less attractive than water. Thus, hamsters' PROP aversion threshold is <1 mM, a concentration that human non-tasters can hardly detect [16].
3.2. Chorda tympani (CT) nerve recordings
Fig. 3 presents responses to 10 mM CyX, PROP and quinine recorded from one hamster CT nerve. A small response to 10 mM CyX, as shown here, was detectable in 4 of the 6 preparations tested; whereas all 6 nerves responded to quinine. Fig. 4 graphs mean CT responses to concentration series of CyX, quinine and PROP. There was no CT response to PROP, just as there is no CT response to caffeine or SOA in hamsters [31]. CT responses to quinine increased with concentration, F (3, 15) = 75.3, p < .000001), with mean relative responses of 13.9 ± 8.7 % to 1 mM quinine and 41.5 ± 7.9 % to 10 mM quinine. No significant responses were recorded to lower concentrations of quinine. A significant increase in CT response to CyX with concentration, F (3, 15) = 6.84, p =.004, was due to the small mean response, 5.3 ± 3.2 %, to 10 mM CyX; a response smaller than the responses to either 1 or 10 mM quinine, F (2, 10) = 48.9, p = .00001. No CT responses occurred to 1 mM or lower CyX concentrations.
Figure 3.

Data Traces of Responses to 10 mM Solutions by Hamster Chorda Tympani Nerve. Recordings from 1 nerve to cycloheximide (CyX), propylthiouracil (PROP) and quinine·HCl: 5-sec of neural activity recorded before and 10 sec after stimulus application (onset indicated by vertical lines aligned for the 3 records) is shown in each trace. Major divisions on the ordinate are 0.01 volts. Numerals at major divisions on the abscissa indicate stimuli were applied between 13 and 16 seconds after starting the recording of the 3 separate epochs.
Figure 4.
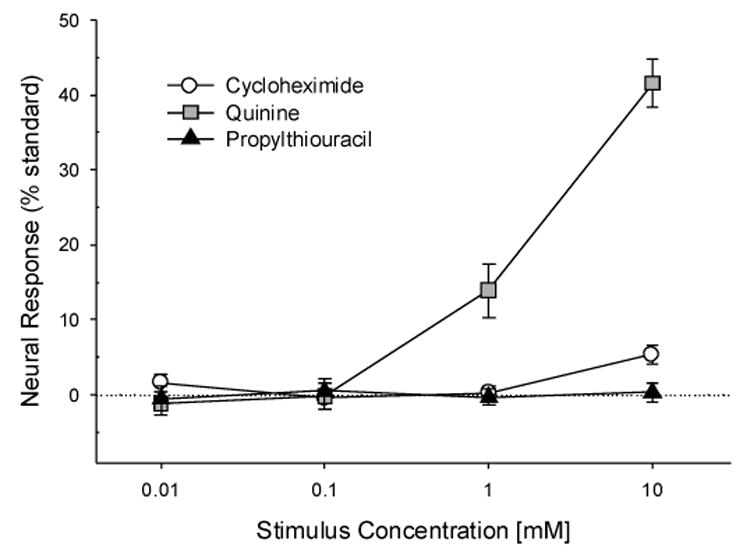
Chorda Tympani Nerve Responses to Cycloheximide, Quinine·HCl and n-Propylthiouracil Concentration Series. Responses, measured as 5-sec post-stimulus minus 5-sec pre-stimulus areas beneath integrated response curves, are given as mean ±se percentages of the standard response to 500 mM NH4Cl.
Thus, 10 mM CyX, which also activates nasal trigeminal polymodal nociceptors and depresses respiration in rats and mice [26], activates the CT. However, the 3 μM CyX that the hamsters avoided in 2-bottle tests (see above) and the 500 μM CyX used in the exposure protocol (see below) had no measurable effect on hamster CT neural activity.
3.3. Effect of a 1-hr cyloheximide presentation on its subsequent intake
Fig. 5 plots intake of 500 μM CyX relative to same-day water intake, intake/H2O, on a first 1-hr presentation (day 0) and on a second presentation, 1-2 days to 22 days later (Table 1). On first exposure, hamsters (N = 28) drank 1.45 ± 0.09 mL of CyX, much less than water, t (29) = 7.8, p < .000001. They consumed 0.38 ± 0.10 mL on the next 1-hr presentation (N=10) 1 or 2 days later, also much less than water, t (14) = 19.42, p<.000001. Mean water intake was 3.62 ± 0.10 mL before exposure to CyX, a mean of means for 23 groupings of 7-29 hamsters used to establish daily water intake (see Methods), and 3.47 ± 0.32 mL (N = 10) after exposure to CyX. All 28 hamsters, averse to CyX on presentation 1, mean CyX1 intake/H2O = 0.383 ± 0.02, were more averse to CyX on presentation 2, F (1, 27) = 140.2, p < .000001, whether it was 1-2, 7, 15, or 22 days later (Table 1). Intakes of the 4 separate groups (N = 6 -10) tested with CyX for a 2nd time after different numbers of days had passed were equivalent, mean CyX2 intake/H2O = 0.081 ± 0.01. Furthermore, 23 animals retested a 3rd time at 23-50 days later were just as averse, mean CyX3 intake/H2O = 0.103 ± 0.01, as on their 2nd exposure, mean CyX2 intake/H2O = 0.083 ±0.01. Eight animals retested a 4th time 43-50 days later, mean CyX4 intake/H2O = 0.016 ± 0.004, found CyX even more aversive, t (7) = 3.93, p = .006, than on their 2nd exposure, mean CyX2 intake/H2O = 0.087 ±0.02. Thus, a single 1-hr exposure to 500 μM CyX suppressed CyX intake by 79 % for weeks, and an additional 3 CyX presentations, with minimal further consumption, suppressed CyX intake by 96%.
Figure 5.
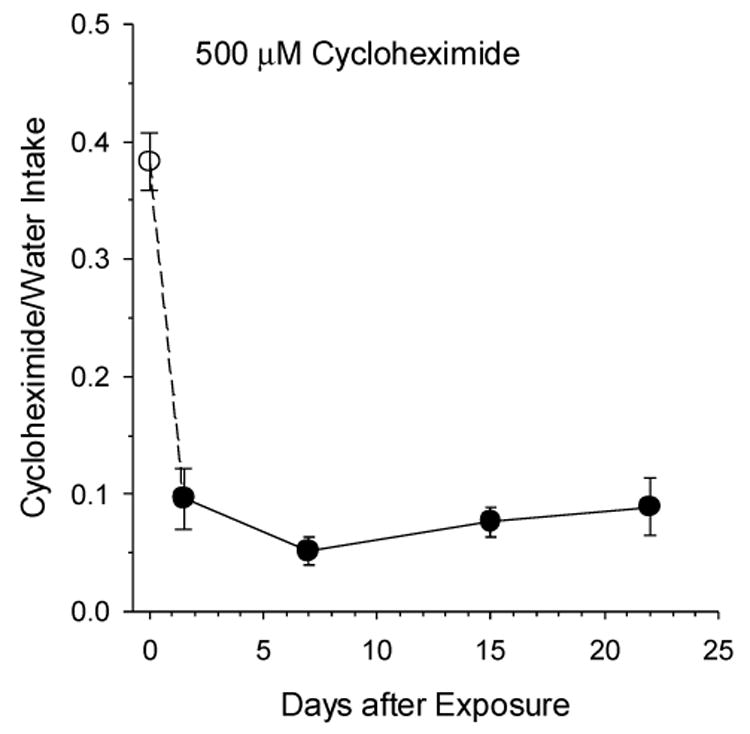
Effect of Presenting 500 μM Cycloheximide (CyX) for 1 Hr on Subsequent CyX Intake. Mean ± se CyX/water intake ratios for the 1st presentation, open point (N=28), and 2nd presentation, filled points for separate subgroups of hamsters (N = 6-10), are plotted. No matter the time after 1st exposure, CyX intake was much diminished on the 2nd offering
Ten animals were tested with NaOH-treated CyX breakdown products (XCyX) after a strong aversion to CyX had been established by 2 to 4 exposures to 500 μM CyX. Mean normalized intake of the breakdown products, XCyX intake/H2O = 0.68 ± 0.11, including the volatile 2,4-dimethylcyclohexanone (Fig. 1), far exceeded CyX2-4 intake/H2O = 0.06 ± 0.02, t (9) = 5.19, p = .0006. However, the 2.71 ± 0.45 mL XCyX intake was significantly lower, t(26) = 2.59, p = 0.02), than same-day water intake, 4.00 ± 0.28 mL (N = 18).
Thus, 1-2 days after one exposure, CyX's aversive potency quadrupled, remaining enhanced for at least 22 days without additional exposure or intake above the ∼0.75 μmole per 120-g animal self-administered. This amount, <1% of the oral LD50 for mice, approximates the oral LD50 for rats [72, 102]. High intake of odorous breakdown products after multiple CyX exposures suggests that the hamsters were not primarily identifying CyX by the odor of CyX breakdown products.
3.4. Taste-stimulus intake suppression after a 1-hr exposure to cycloheximide
Fig. 6 presents results on the effects of CyX presentation and self-administration on intake of other solutions. Each pair of bars compares mean intake before and 1-2 days after the initial 1-hr presentation of 500 μM CyX to the hamsters. The leftmost pair of bars presents pre- and post-CyX water intake.
Figure 6.
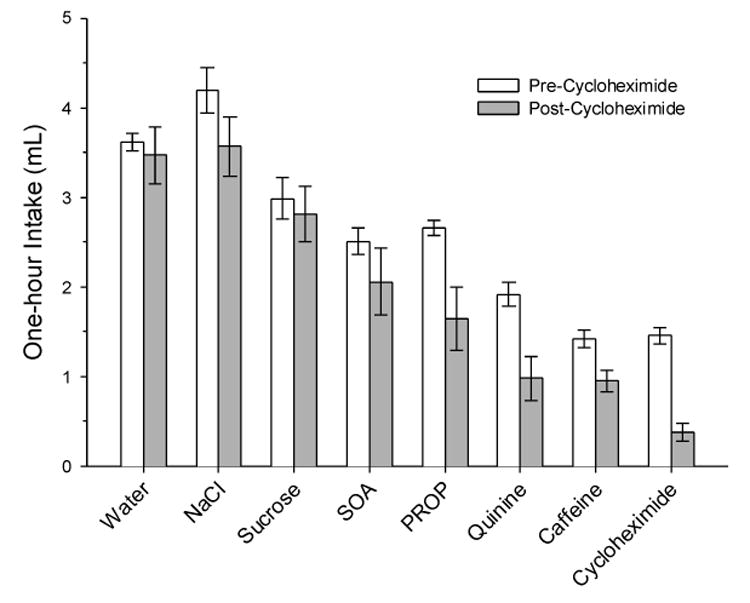
Effect of One 1-Hr Exposure to 500 μM Cycloheximide (CyX) and Self-Administration of 0.75 μMoles of CyX on Intake of Solutions. Means (± se) are plotted. After a single CyX presentation, CyX intake was much reduced compared to the reduction in water intake (p<.0005); drinking of other bitter solutions was reduced to a lesser extent, while drinking of NaCl and sucrose was unaffected.
The pre-exposure results, clear bars in Fig. 6, suggest that 1 mM quinine, 30 mM caffeine and 500 μM CyX were equi-aversive to hamsters. ANOVA of TS intake/H2O ratios before CyX exposure, F(6,83) = 41.2, p <.000001, revealed mean intake (± se) of NaCl, 1.10 ±0.07, sucrose, 0.78 ±0.06, SOA, 0.70 ±0.04, and PROP, 0.66 ±0.02, were higher than CyX intake (NK ps = .0001); but the intake of quinine, 0.48 ±0.04, and caffeine, 0.35 ±0.02, approximated CyX intake, 0.383 ± 0.02, and, thereby, may be considered as aversive as CyX.
CyX exposure had general effects on intake (filled bars in Fig. 6): mean post-exposure intake fell below mean pre-exposure intake for all TS, including water and non-bitter stimuli. Thus, TS suppression was considered significant if it reliably exceeded water-intake suppression. ANOVA, F (7, 48) = 6.87, p = 00001, revealed that TS post/pre-CyX ratios differed across individual solutions; but only the CyX ratio of 0.25 ±0.06, lower than ratios for all other solutions (NK p <.05), differed significantly from water post/pre-CyX, 0.90 ±0.08 (NK p =.0005) (Fig. 6). However, an ANOVA done after grouping data for the 2 non-bitter, N = 12, and 4 bitter, N = 24, stimuli, F (3, 52) = 15.33, p<.000001, revealed a 0.653 ± 0.06 bitter-stimulus post/pre-CyX ratio suggestive of taste-stimulus suppression (NK p < .05). Fig. 7 shows results for combined stimuli converted to percent suppressions, with general suppression, estimated by the 10.2 % water-intake suppression, subtracted. The conversion allows quantitative comparison to published data on LiCl-induced aversions, studies in which general effects due to toxin injection were controlled with a water-conditioned group [31]. Exposure to CyX suppressed intake of the combined bitter stimuli, including stimuli as aversive as CyX (quinine, 37.1 ± 12.7 %; caffeine, 20.9 ± 7.3 %) and less aversive than CyX (SOA, 14.5 ± 13.5 %; PROP, 25.6 ± 12.6 %), but not nearly as much as it suppressed intake of CyX itself. Average bitter-stimulus suppression, 24.5 ± 5.8 %, compared to 64.7 ± 6.4 % for CyX (NK p = .0002). Thus, in the context of ∼75 % suppression of intake of other ionic bitter stimuli stemming from LiCl-induced aversions to quinine (31), we conclude that CyX-induced intake suppression to aversive stimuli other than CyX was weak.
Figure 7.

Mean (±se) Percent Suppression of Taste Stimulus Intake following One Exposure to 500 μM Cycloheximide (CyX). Like CyX (p = 0002), bitter stimuli [1 mM quinine, 1 mM SOA, 5 mM PROP, 30 mM caffeine] were suppressed (p = 05); non-bitter stimuli [.1 M NaCl, .1 M sucrose] were not.
3.5. Candidate bitter-receptor genomics: a genetic context for cycloheximide
Fig. 8 presents results of comparing receptor-protein amino acid (AA) sequences of rat to mouse and human to mouse. A 2-way ANOVA of percent AA identity in the sequences of the 4 groups of proteins listed in Table 2 revealed significant effects of protein group, F (3,6) = 61.1, p = .0001; species, F (1, 6) = 461.9, p = .000001, and a protein-species interaction, F (3,6) = 46.8, p = .0001.
Figure 8.

Percent Amino Acid Identity (± se) of Orthologous Proteins of Rat and Mouse or Human and Mouse. The 4 groups of proteins (see Table 2) from left to right are: 2 T2R receptors, 3 T1R receptors, 2 cytochrome P450 enzymes and (combined) potassium channel protein KCNJ6, olfactory receptor I7 and metabotropic glutamate receptor mGluR1. Amino acid (AA) identity is greater for the rat-mouse comparison than the human-mouse comparison (p = .000001); and the T2R candidate bitter receptors have the lowest AA identity of the 4 groups of proteins (α = .05).
Candidate T2R bitter-taste receptor homologous pairs for rat and mouse (rT2R9–mT2R5, rT2R1–mT2R19) shared 84–85 %, whereas, human–mouse pairs (hT2R10–mT2R5, hT2R1–mT2R19) shared only 54–52 % AA identity; a large average 32 % rat-human species difference (NK p = .0002). Mouse mT2R5 and rat rT2R9 receptors are activated by μM CyX but the human best-match, hT2R10, is not [17, 20]. The hT2R10 and mT2R5 orthologous receptors may share descent but not ligands because of rapid T2R evolution.
To further explore evolution of CyX receptors of the 2 rodents, we compared protein sequences coded for by 5-gene, syntenic clusters in the rat and mouse genomes that include the genes associated with high CyX sensitivity. The cluster is not found in the human genome [93]. Results showed a rat-mouse mean 84.8 ± 0.4 % AA identity for orthologous proteins, consistent with related function for each of the 5 across-species pairs of genes. Means for within-cluster, within-species paralogous proteins were 56.4 ± 1.1% AA identity for mouse and 55.7 ±1.3 % AA identity for rat, consistent with the idea that each of the 5 genes has a distinct function within each species. Thus, based on gene duplication, mouse and rats share a group of 5 T2R receptors with distinct ligands, none of which is a ligand for the single human orthologue.
In order to better estimate the point in evolutionary time of this rat-mouse “CyX clade” radiation, we probed the Ensembl database on the European rabbit, Oryctolagus cuniculus, with the entire nucleotide sequence of mouse mT2R5. The single hit was a gene coding for a 316 AA candidate rabbit T2R receptor that was best matched, 63% AA identity, by human hT2R10, not any member of either rat or mouse 5-gene “CyX clade.” The % AA identities of rabbit ‘oT2R10’ with receptors within mouse and rat clades averaged 53.1 ±0.40 %; the human hT2R10 to mouse and rat clade members averaged 55.5 ±0.39 %. Thus, the rabbit genome, like the human, has neither a specific CyX receptor nor a “CyX clade” of genes. The CyX receptor must have evolved after the rodents diverged from human and lagomorph species.
As predicted, candidate sweet-umami taste receptors, T1R1, T1R2 and T1R3, and cytochrome P450 proteins, Cyp1a2 and Cyp2d6 (see Table 2), had intermediate AA identities for orthologous pairs (Fig. 8); rat-mouse ranging from 88 to 93 % and human-mouse from 69 to 74 %, demonstrating a significant ∼20% rat-human difference (NK p = .0003). AA sequences of the other channels/receptors, K+ channel protein KCNJ6, olfactory octanal receptor OR-I7 and metabotropic glutamate receptor mGluR1, in contrast, are, as expected, more homogeneous (conserved) across the species, with 90 to 99 % rat-mouse and human-mouse AA identity. For OR-I7, human-mouse AA identity was 90 % and rat-mouse was 95 % [60]. Thus, T1R candidate sweet and glutamate receptors show intermediate divergence across species.
Also, consistent with predictions based on other data (93), protein sequence results have T2R receptor proteins evolving more rapidly than T1Rs, validating the approach of comparing AA sequences in best-fit, orthologous proteins to sequences of a standard species, the mouse.
Rates of synonymous substitutions (SS) in nucleotide codons for conserved AAs, “a molecular clock,” were studied with the goal of estimating evolutionary relationships between hamsters and other species. ANOVA on rat-mouse vs. human-mouse percent SS of genes for the full set of 10 proteins listed in Table 2 revealed an expected evolutionary-time rat-human species effect, F (1, 6) = 127.7, p = .00003. Mean SS percentages were 14.0 ± 0.72 % for rat-mouse and 31.0 ± 0.45 % for human-mouse, indicating less evolutionary time between the 2-rodent than the human-rodent divergences. An ANOVA comparing rat-mouse, hamster-mouse and human-mouse % SS for genes of a reduced set of proteins, the 2 cytochrome P450s and the K+ channel, also revealed a significant species effect, F (2,4) = 52.2, p = .001, with distinct means: 14.2 ± 1.5 % for rat-mouse, 19.6 ± 1.4 % for hamster-mouse and 28.9 ± 1.2 % for human-mouse (NK ps ≤ .02). Similar SS percentages in the full and reduced set for rat-mouse and human-mouse validate the hamster-mouse SS value. Finally, we also determined % SS for a more distant non-mammalian vertebrate puffer fish to mouse for 2 orthologous genes, mGluR1 and KCNJ6; the SS values, 49.9 and 48.5% respectively, are close to estimated % SS upper limits. Our results are consistent with hamsters being genetically more distinct from mice and rats than mice and rats are from each other. Thus, hamsters separated before mouse and rat in the rodent radiation.
Our genomic comparisons of rat and human to mouse attest to species diversification being greater for bitter receptor proteins than sweet receptor or cytochrome P450 proteins, which are themselves quite diverse across species compared to highly “conserved” proteins. Also, our results are consistent with a rodent-specific, high-affinity CyX receptor, evolving after the rodent radiation, wherein cricetid rodents separated from murid rodents well before the rat and mouse murids diverged. These results provide a genetic context for behavioral and neural species differences in CyX sensitivity.
4. Discussion
4.1. Cycloheximide: No ordinary bitter stimulus
Although nearly tasteless to humans, rats and mice avoid cycloheximide (CyX) at concentrations much lower than thresholds for most bitter stimuli [12, 68,101]. Hamsters also found this antibiotic uniquely aversive with a ∼1 μM detection threshold (Fig. 2). However, unlike other aversive chemosensory stimuli: tastes, odors, and irritants, polymodal ethanol or an equally aversive antiseptic chlorhexidine [Fig. 2, 23, 87, 90], CyX was not better tolerated when mixed with a sweetener. Thus, hamsters, like other rodents, are highly sensitive to CyX and that CyX aversion persists, unlike aversions to other chemosensory stimuli, when mixed with sweet stimuli.
Like other non-ionic bitter stimuli to which hamsters are averse, 1 mM SOA and 1 mM PROP [31, Fig. 2], 1 mM CyX failed to activate the hamster chorda tympani (CT) nerve. Unlike those compounds for which solubility limits were attained without CT response, CyX elicited a small consistent CT response at 10 mM (Figs. 3, 4), suggesting activation of low-affinity receptors. Like hamster, B6 mouse CT neurons and rat gustatory neurons (CT and neurons in CT brainstem projection sites within the rat nucleus of the solitary tract, NST) respond weakly to CyX, but CTs of SWR and SWR-derived mice are more sensitive [22, 63 74, 84,100,113]. The SWR mouse strain has a unique neural and behavioral sensitivity to SOA [49, 66] but is not necessarily behaviorally more sensitive to CyX than other mouse strains [11, 12, 68, 78]. The CyX-specific, mT2R5 receptor has a Kd of ∼1 μM CyX [20] and CT responses and behavioral aversions to CyX are abolished in SWR T2R5 knockouts, which remain sensitive to other bitter stimuli, including non-ionic PROP and SOA, and ionic quinine·HCl [74]. Unlike the SWR CT, which responds to 0.1-1.0 mM CyX, the 10 mM CyX threshold of the hamster CT is 10,000 times higher than the hamster's behavioral threshold. Therefore, the hamsters' aversions to micromolar CyX are not tied to a CT response (Fig. 4).
The glossopharyngeal (GL) nerve may provide chemosensory cues to account for rodent behavior toward CyX and other nonionic bitter stimuli. The GL innervates posterior lingual taste papillae and posterior soft palatal mucosa [107]. The B6 mouse GL responds well to 100 μM CyX and 10 mM PROP, stimuli also well above behavioral aversion thresholds [11,22]. Furthermore, responses to 10 μM CyX dominated in situ Ca++ imaging results from GL-innervated rat foliate taste-bud cells [19] and single NST neurons activated by foliate stimulation [42]. The physiological data coincide with abundant localization of T2Rs in the GL receptive fields [1,71,74]. Moreover, only with the GL intact did CyX elicit profuse salivation. With the GL cut, most rats, after self-administration of up to 6 mL of a 2% CyX solution (well above the LD50), died; the rats' intestinal walls had hemorrhaged [84]. Besides afferents that initiate gustatory-salivary reflexes, the GL carries secreto-motor fibers that control parotid gland secretions [38, 53] and sensory components of quinine-elicited oral motor rejection reflexes [58, 59] from the rat posterior tongue. The vagus nerve may also provide chemosensory cues to account for rodent behavior toward CyX and other nonionic bitter stimuli. The vagus innervates laryngeal, pharyngeal and gastrointestinal mucosa [107]. The vagal laryngeal branch [27] innervates T2R receptors found in taste buds, gastrointestinal tract enteroendocrine cells [109,110], and solitary chemosensory cells (SCC), thought to be a chemosensory “alarm system” [91]. Hamster laryngeal vagal nerve fibers respond strongly to the non-ionic bitter stimulus urea [24], a stimulus that hamsters fail to cross-generalize with multiple ionic bitter stimuli [36]. T2R5 CyX receptors initiate mouse neural circuits that project to posterior brainstem regions hardly overlapping regions receiving input from the CT and mT1R3 [99], one moiety of the candidate heterodimeric sweet receptor. Thus, neuronal circuits initiated by CyX T2R receptors provide visceral sensory-motor cues that rodents may use to identify CyX.
Mice and rats use odor cues to learn to avoid a bottle containing the “awful” 355 μM CyX, odor cues that are acutely sensitive to systematic disruption and restoration of the olfactory pathways [39, 57, 73]. As CyX itself is not volatile, odor of its fragrant dimethylcyclohexanone breakdown product may be involved (Fig. 1). We found hamsters drank CyX breakdown products like water in 2-bottle tests (Fig. 2). Thus, although CyX solutions may smell of breakdown products, those odors themselves did not suppress hamster intake. Also, hamsters found CyX quite aversive after multiple exposures but still readily drank a solution of the breakdown products. Thus, in contrast to mice and rats, we have little evidence for a significant role of odor in hamster behavior toward CyX, native or induced. Nasal irritation may possibly play a role; SCCs in the nasal cavity and vomeronasal ducts innervated by the trigeminal nerve express T2R receptors that respond to CyX [26].
Altogether, we can conclude that CyX, a potent toxin detected in solution at μmolar levels, is hardly simply a bitter taste stimulus to hamsters. Its aversive potency is not ameliorated by sweet and it is not sensed by the chorda tympani nerve; micromolar CyX is likely detected by afferents in glossopharngeal, vagal or trigeminal nerves that link to sites of CyX T2R receptor abundance.
4.2. Cycloheximide: Once is enough
Surprisingly, naïve hamsters drank 1.5 mL of 500 μM CyX when sampling for 1 hour from a single bottle despite detecting and avoiding 0.3 μM CyX, more than 1500 times more dilute, when having a choice of water in another bottle for 48 hrs (Fig. 2). In contrast, hamsters drink 0.8 mL of 3 mM quinine·HCl in 1 hr from a single bottle, when their 2-bottle aversion threshold for quinine is 0.3 mM, 10 times more dilute [31, 37]. Thus, CyX, unlike the bitter prototype, may not at first be highly aversive to hamsters. But it is highly aversive after one exposure, a plasticity apparently not seen in other rodents. CyX intake by mice approaches zero for 10 μM in either brief-access or 48-hr tests [11, 78]. Hamster voluntary consumption of a substantial amount of CyX was not fatal. Nevertheless, they drank more than 1 mL just once. When tested on two days in a row or with as many as 21 days intervening, intake in a 2nd sampling was minimal and the “acquired” aversion continued, without additional reinforcement, for at least 3 weeks (Fig. 5). It is unknown whether the enhanced avoidance of CyX is a learned [18] or is due to genetic induction.
Toxins that contaminate exclusive water or food supplies would devastate species without environmental response genes. Such genes can be induced to increase enzyme production beyond constitutive levels in order to breakdown/sequester toxins once ingested; or they, possibly, could induce receptor-protein expression upon ingestion to enhance aversion and prevent significant future intake [7]. A compelling example of genetic enzyme induction post-ingestion occurs in lepidopterans that use cytochrome P450 mono-oxygenases to rapidly breakdown lethal host-plant substances that, once ingested, instigate transcription of larval cytochrome P450 genes [6, 21, 86]. Intake of xenochemicals by humans also induces cytochrome P450 enzymes that break down toxins such as ethanol [77]. Empirical evidence for induction of chemoreceptor proteins is sparse; however, post-exposure functional changes in insect chemo-receptors [43, 88], mouse and human odor receptors [3, 103,104] and, possibly, hamster taste receptors [9] are suggestive.
It has been proposed that an experience with a bitter taste per se may induce aversion learning [18]. Rodent behavior toward chemosensory stimuli changes after experience with safe or dangerous compounds to which rats and mice are averse. For example, benign aversive SOA is avoided more by naïve rats when measured for a 2nd time, compared to a 1st [23]; and avoided more by mice when a concentration series is presented in descending compared to ascending order [12]. Intake is also lower for descending, compared to ascending, cyclamate concentration series for mice trained to avoid sodium cyclamate [45]. Thus, some post-exposure intake suppression via unknown mechanisms is seen for systemically benign aversive stimuli, whether aversions are native or learned by association with a toxin.
Experiential effects on intake are also documented for bitter compounds that, like CyX, are far from benign. Phenylthiocarbamide (PTC), for which a gene coding a human T2R receptor is well characterized [54, 55], is an odorous bitter compound with prominent adverse systemic effects [44]. In fact, the strongest evidence for T2R receptors mediating human bitter taste is the association of hT2R38 sequence variation with human polymorphisms in tasting PTC [16], although PTC receptor ligand binding sites may not contribute to the variation [28]. Mice, especially inbred strains that avoid PTC initially, become more averse to PTC with repeated testing [67]. In brief-access tests, however, PTC intake hardly differs from water intake, suggesting PTC has little, or an initially neutral, taste to the mice [78]. Like traditional toxins, PTC [97], or even ethanol [15, 89], when injected IP induces learned aversions to associated taste stimuli. Thus, post-exposure intake suppression is seen for systemically harmful stimuli, whether self-administered or introduced experimentally.
Like naïve hamster consumption of CyX, rats initially readily consume LiCl, which has a salty taste to humans and tastes like NaCl to rats. Self-intake or IP injection of LiCl after tasting LiCl induces strong aversions not only to LiCl itself [75, 76] but to similar-tasting NaCl. LiCl's adverse post-ingestional effects [44] are associated with activation of visceral nociceptors [111] and neither the LiCl nor benign NaCl taste aversion is readily forgotten. The post-exposure, persistent 65% decrease in CyX intake shown in Fig. 5 is reminiscent of the effects of LiCl intake on consumption. Like LiCl intake, CyX intake resulted in a strong aversion to the consumed toxic compound itself.
Generalizations of LiCl-induced conditioned taste aversions (CTA) have identified quality-similarities of multiple bitter stimuli in hamsters and other rodents [31,47,81]. The taste quality and intensity of conditioned stimuli (CS) are not altered; rather, CS central negative hedonic signs are strengthened [48, 111]. However, unlike aversions to LiCl after intake or IP injection, induced CyX aversions did not generalize strongly to other compounds. By combining our data on animals tested on the 4 bitter stimuli, essentially quadrupling N, a reliable 25 % bitter-stimulus intake-suppression was revealed (Fig. 7). However, the stimulus intake suppressions following CYX exposure, 37 % for quinine and 21 % for caffeine, were weak; much lower than ∼75% LiCl-induced generalizations to multiple ionic bitter stimuli [31]. The weakness of the generalizations from CyX is not likely due to faint test stimuli [82]; before CyX exposure, the hamsters found quinine and caffeine as averse as CyX. The weak generalizations suggest that hamsters, like rats, perceive some similarities in CyX and quinine [13], as well as caffeine. As hamsters do not cross-generalize learned aversions from quinine to caffeine [31], CyX may have a heterogeneous taste with several distinct qualities. LiCl-induced CTAs hamsters learned to binary, ternary and quaternary heterogeneous taste mixtures were generalized to single-quality components [32].
Therefore, unlike prototype bitter quinine, CyX was not at first strongly avoided, and, unlike LiCl-induced learned aversions to quinine, which generalize specifically to ionic bitter stimuli, CyX-induced aversions generalized weakly and indiscriminately to ionic and non-ionic bitter stimuli. Perhaps, neither odor nor bitter taste but activation of non-taste oral or gastrointestinal reactions cue learning or, by unknown mechanisms, induce CyX T2R receptors to levels above constitutive levels. Receptor induction would provide the hamsters with a sensitizable, second-tier toxin defense and explain why, for the very dangerous toxin CyX, once is enough.
4.3. Cycloheximide: Microbe-rodent co-evolution?
Species diversity in taste is associated with varied ecological niches with restricted food choices and/or water access. Bitter taste, one of numerous strategies evolved for dealing with dangerous compounds [7, 91,105], is especially diverse (Fig. 8); albeit human-rodent diversity for salt taste, which assists in ion-water homeostasis, is also notable [30, 65]. Hamsters, like other granivores or herbivores, are insensitive to stimuli that are quite bitter to humans. For example hamsters fail to detect denatonium [108] unless concentration is raised 100,000 times above the human threshold [31]. In contrast, CyX, a naturally occurring antibiotic with unique structure produced by the actinomycete Streptomyces griseus [102], is detected at <1 μM by hamsters, as well as lab mice and rats; but has uncertain bitterness at 1 mM to people [35, 68, 101].
Vertebrate species vary a great deal in T2R gene numbers and sequences [93]. Some T2Rs may be conserved from mouse to human [94], such as the homologous pairs mT2R1–hT2R1 and mT2R31–hT2R38, the latter accounting for human polymorphisms in tasting PTC as bitter [54, 56]. But T2Rs hardly match the sequence preservation in most orthologous protein sequences [69], not even T1R candidate sweet receptors [114], which are themselves as diverse across species as cytochrome P450s (Fig. 8), enzymes known for sequence polymorphisms [77]. Although T2R candidate bitter receptors [1] were discovered before the T1R receptors, our results, consistent with the conclusion that species diversification is greater for bitter receptors than sweet receptors [93], help explain the difficulty in studying the bitter taste across species. The few T2R ligands known are species specific [17, 20, 54, 55, 114]. Our data on hamsters and cycloheximide further substantiate that the “bitterness” people perceive is not likely a single quality in other species, which have distinct aversive stimulus classes, none “a close homologue of the human bitter taste” [31].
Rodent-human comparisons put T2Rs among the most diverse orthologues, a species divergence that, along with T2R receptor multiplicity, makes it extremely difficult to recognize functional relationships across T2R receptor proteins. The 53% AA identity for “conserved” T2R1 and lineage-specific CyX receptors [94] with human othologues (Fig. 8) fell among the lowest 3% of values for all identified mouse-human orthologous protein sequences [69]. In fact, the % AA identities of these human-mouse candidate orthologues, receptors with presumed related function across species, approximated % AA identities of within-mouse T2R paralogues, receptors with distinct functions within a single species.
The mouse CyX receptor gene, mT2R5, and its rat orthologue code for rodent lineage-specific bitter receptors [94]. CyX is neither an effective ligand for hT2R10, the human orthologue [17], nor is it likely to be for the rabbit T2R orthologue ‘oT2R10’, first described here. The rabbit findings suggest that the CyX mT2R5 gene, along with other members of the “CyX clade” of 5 quite distinct paralogous genes, evolved some time after the lagomorph-rodent divergence 55 million years ago (mya), well before cricetid (hamsters) and murid (mus, rattus) rodent families diverged 24 mya and, consistent with our molecular clock data, before the murid mus-rattus split 11 mya [25, 80, 95, 96]. A rough estimate, based on the premise that nucleotide differences reflect merely the passage of time, has rodent “CyX clades” evolving about 33 mya: mouse-rat across-species clade-member orthologous nucleotide sequences differ by an average 10 % but within-species paralogous clade nucleotide sequences differ by an average 30 %. Multiple paralagous rodent genes in “CyX clades” with distinct, perhaps specific, ligands portend a difficult future for study of lineage-specific T2R receptors, which may well have ligands that specifically threaten rodent survival. The fortunate, fortuitous discovery of CyX as the specific ligand for mT2R5 reflects its ubiquitous laboratory use and the specific CyX-TR2 receptor interaction may be helpful for determining the nature of T2R ligand-receptor binding sites. Nonetheless, species diversity in T2Rs leaves humans and rabbits without the unusually sensitive CyX receptor or likely any of another 4 receptors coded for by the “CyX clade” of genes.
CyX T2R receptor structure may be controlled by diversifying adaptive evolution, with selection driven by a dangerous protein-synthesis inhibiting antibiotic inherent in the ecological niche. Golden hamsters likely have a CyX gene given their < 1 μM CyX behavioral sensitivity. Hamsters are fossorial rodents that construct deep burrows with multiple chambers [41], storing large grain caches underground in intimate contact with soil, an ideal habitat for the micobe that makes CyX, S. griseus. Furthermore, hamsters diverged from mouse/rat more than 10 million years before the mouse and rat diverged. Cricetids, unlike murids, carry grain in large, expandable external pouches [85] and, among rodents, use a distinct strategy to control ethanol toxicity [4, 51, 61, 62]. Consistent with the plasticity that we report here, hamsters may also use an inducible mechanism to defend against CyX toxicity. Although well-studied, rats and mice are not reported to decrease intake after one tolerable contact with CyX but appear to avoid it on first encounter. This argues for sequencing T2R genes and chemical ecological studies of hamsters to explore co-evolution of ligands and receptors for the likely, mostly species-specific, bitter tastes.
4.4. Conclusions
Cycloheximide (CyX), an exceptionally strong aversive stimulus in mice and rats, is especially potent in golden hamsters after experience, a behavioral plasticity, possibly learning based on consequences of intake, but not reported for mouse or rat. Ready initial intake of CyX at high concentrations, failure of a sweet stimulus to alleviate CyX avoidance, and weak, less specific generalization of induced CyX aversions are each unlike hamsters' reactions towards other bitter taste stimuli, suggesting bitter is not a unitary quality conserved across mammalian species. In contrast to mice and rats, odors of CyX breakdown products are not cues for a learned induced CyX sensitivity in hamsters. Mouse CyX potency is associated with candidate bitter receptor mT2R5 for which there is a rat orthologue, but the hamster lineage branches before mouse and rat in evolutionary time and the closest human homologue is insensitive to CyX as may be its rabbit orthologue. Furthermore, a CyX-insensitive hamster anterior taste-bud field is compatible with a preponderance of CyX sensitivity and T2R5-receptor localization to posterior lingual taste buds, laryngeal epithelium, gastrointestinal tract and elsewhere in rodents. Any “bitter taste” of CyX shared with other bitter stimuli is likely transmitted by sensory nerves other than the chorda tympani. Thus, CyX has complex species-specific effects unlike other aversive/bitter stimuli and the mouse CyX stimulus–T2R receptor interaction may be one of multiple specific interactions evolved in one species for protection from toxins generated by other species.
Acknowledgments
This work was supported by an NIH grant from NIDCD: DC004099.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJP, Zuker CS. A novel family of mammalian taste receptors. Cell. 2000;100:693–702. doi: 10.1016/s0092-8674(00)80705-9. [DOI] [PubMed] [Google Scholar]
- 2.Araneda RC, Kini AD, Firestein SK. The molecular receptive range of an odorant receptor. Nat Neurosci. 2000;3:1248–1255. doi: 10.1038/81774. [DOI] [PubMed] [Google Scholar]
- 3.Araneda RC, Firestein S. The scents of androstenone in humans. J Physiol. 2003;554:1. doi: 10.1113/jphysiol.2003.057075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Arvola A, Forsander O. Comparison between water and alcohol consumption in six animal species in free-choice experiments. Nature. 1961;191:819–820. doi: 10.1038/191819a0. [DOI] [PubMed] [Google Scholar]
- 5.Beidler LM, Fishman IY, Hardiman CW. Species differences in taste responses. Am J Physiol. 1955;181:235–239. doi: 10.1152/ajplegacy.1955.181.2.235. [DOI] [PubMed] [Google Scholar]
- 6.Berenbaum MR. The chemistry of defense: theory and practice. Proc Natl Acad Sci U S A. 1995;92:2–8. doi: 10.1073/pnas.92.1.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Berenbaum MR. Postgenomic chemical ecology: from genetic code to ecological interactions. J Chem Ecol. 2002;28:873–896. doi: 10.1023/a:1015260931034. [DOI] [PubMed] [Google Scholar]
- 8.Bermúdez-Rattoni F, Bures J, Yamamoto T. Paradoxes, projections, and perspectives of CTA research. In: Bures J, Bermúdez-Rattoni F, Yamamoto T, editors. Conditioned taste aversion: memory of a special kind. New York: Oxford University Press; 1998. pp. 121–131. [Google Scholar]
- 9.Berterethce MV, Boireau-Ducept N, Pillias AM, Faurion A. Stimulus-induced increase of taste responses in the hamster chorda tympani by repeated exposure to ‘novel’ tastants. Appetite. 2005;45:324–333. doi: 10.1016/j.appet.2005.06.002. [DOI] [PubMed] [Google Scholar]
- 10.Booth DA, Simson PC. Aversions to a cue acquired by its association with effects of an antibiotic in the rat. J Comp Physiol Psychol. 1973;63:101–108. doi: 10.1037/h0035293. [DOI] [PubMed] [Google Scholar]
- 11.Boughter JD, Jr, Raghow S, Nelson TM, Munger SD. Inbred mouse strains C57Bl/6J and DBA/2j vary in sensitivity to a subset of bitter stimuli. BMC Genet. 2005;6:36. doi: 10.1186/1471-2156-6-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boughter JD, Jr, Whitney G. Behavioral specificity of the bitter taste gene Soa. Physiol Behav. 1998;63:101–108. doi: 10.1016/s0031-9384(97)00398-3. [DOI] [PubMed] [Google Scholar]
- 13.Brasser SM, Mozhui K, Smith DV. Differential correlation in taste responsiveness to bitter stimuli in rats. Chem Senses. 2005;30:793–799. doi: 10.1093/chemse/bji071. [DOI] [PubMed] [Google Scholar]
- 14.Breslin PAS, Tharp CD. Reduction of saltiness and bitterness after a chlorhexidine rinse. Chem Senses. 2001;26:105–116. doi: 10.1093/chemse/26.2.105. [DOI] [PubMed] [Google Scholar]
- 15.Broadbent J, Muccino KJ, Cunningham CL. Ethanol-induced conditioned taste aversion in 15 inbred mouse strains. Behav Neurosci. 2002;116:138–148. [PubMed] [Google Scholar]
- 16.Bufe B, Breslin PAS, Kuhn C, Reed DR, Tharp CD, Slack JP, Kim UK, Drayna D, Meyerhof W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Cur Biol. 2005;15:322–327. doi: 10.1016/j.cub.2005.01.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bufe B, Hofmann T, Krautworst D, Raguse JD, Meyerhof W. The human TAS2R16 receptor mediates bitter taste in response to β-glucopyranosides. Nature Gen. 2002;32:397–401. doi: 10.1038/ng1014. [DOI] [PubMed] [Google Scholar]
- 18.Bures J. Ethology, physiological psychology, and neurobiology of CTA. In: Bures J, Bermúdez-Rattoni F, Yamamoto T, editors. Conditioned taste aversion: memory of a special kind. New York: Oxford University Press; 1998. pp. 1–13. [Google Scholar]
- 19.Caicedo A, Roper SD. Taste receptor cells that discriminate between bitter stimuli. Science. 2001;291:1557–1560. doi: 10.1126/science.291.5508.1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chandrashekar J, Mueller KL, Hoon MA, Adler E, Feng L, Guo W, Zuker CS, Ryba NJ. T2Rs function as bitter taste receptors. Cell. 2000;100:703–711. doi: 10.1016/s0092-8674(00)80706-0. [DOI] [PubMed] [Google Scholar]
- 21.Cohen MB, Schuler MA, Berenbaum MR. A host inducible cytochrome P-450 from host-specific caterpillar: Molecular cloning and evolution. Proc Natl Acad Sci USA. 1992;89:10920–10924. doi: 10.1073/pnas.89.22.10920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Danilova V, Hellekant G. Comparison of the responses of the chorda tympani and glossopharyngeal nerves to taste stimuli in C57BL/6J mice. BMC Neurosci. 2003;4:5. doi: 10.1186/1471-2202-4-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dess NK. Responses to basic taste qualities in rats selectively bred for high versus low saccharin intake. Physiol Behav. 2000;69:247–257. doi: 10.1016/s0031-9384(99)00246-2. [DOI] [PubMed] [Google Scholar]
- 24.Dickman JD, Smith DV. Response properties of fibers in the hamster superior laryngeal nerve. Brain Res. 1988;450:25–38. doi: 10.1016/0006-8993(88)91541-7. [DOI] [PubMed] [Google Scholar]
- 25.Douzery EJP, Huchon D. Rabbits, if anything, are likely Glires. Mol Phylogenet Evol. 2004;33:922–935. doi: 10.1016/j.ympev.2004.07.014. [DOI] [PubMed] [Google Scholar]
- 26.Finger TE, Bottger B, Hanson A, Anderson KT, Alimohammadi H, Silver WL. Solitary chemoreceptor cells in the nasal cavity serve as sentinels of respiration. Proc Natl Acad Sci USA. 2003;100:8981–8986. doi: 10.1073/pnas.1531172100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Finger TE, Danilova V, Barrows J, Bartel DL, Vigers AJ, Stone L, Hellekant G, Kinnamon SC. ATP signaling is crucial for communication from taste buds to gustatory nerves. Science. 2005;310:1495–1499. doi: 10.1126/science.1118435. [DOI] [PubMed] [Google Scholar]
- 28.Floriano WB, Hall S, Vaidehi N, Kim U, Drayna D, Goddard WA., 3rd Modeling the human PTC bitter-taste receptor interactions with bitter tastants. J Mol Model. 2006;12:931–941. doi: 10.1007/s00894-006-0102-6. [DOI] [PubMed] [Google Scholar]
- 29.Frank ME. Taste nerve recordings in rodents. In: Spielman AI, Brand JG, editors. Experimental cell biology of taste and olfaction. Boca Raton: CRC Press; 1995. pp. 263–270. [Google Scholar]
- 30.Frank ME. Neuron types, receptors, behavior, and taste quality. Physiol Behav. 2000;69:53–62. doi: 10.1016/s0031-9384(00)00188-8. [DOI] [PubMed] [Google Scholar]
- 31.Frank ME, Bouverat BP, MacKinnon BI, Hettinger TP. The distinctiveness of ionic and nonionic bitter stimuli. Physiol Behav. 2004;80:421–431. doi: 10.1016/j.physbeh.2003.09.009. [DOI] [PubMed] [Google Scholar]
- 32.Frank ME, Formaker BK, Hettinger TP. Taste responses to mixtures: Analytic processing of quality. Behav Neurosci. 2003;117:228–235. doi: 10.1037/0735-7044.117.2.228. [DOI] [PubMed] [Google Scholar]
- 33.Frank ME, Formaker BK, Hettinger TP. Peripheral gustatory processing of sweet stimuli by golden hamsters. Brain Res Bull. 2005;66:70–84. doi: 10.1016/j.brainresbull.2005.04.004. [DOI] [PubMed] [Google Scholar]
- 34.Frank ME, Gent JF, Hettinger TP. Effects of chlorhexidine on human taste perception. Physiol Behav. 2001;74:85–99. doi: 10.1016/s0031-9384(01)00558-3. [DOI] [PubMed] [Google Scholar]
- 35.Frank ME, Hettinger TP. What the tongue tells the brain about taste. Chem Senses. 2005;30 1:i68–i69. doi: 10.1093/chemse/bjh117. [DOI] [PubMed] [Google Scholar]
- 36.Frank ME, Nowlis GH. Learned aversions and taste qualities in hamsters. Chem Senses. 1989;14:379–394. [Google Scholar]
- 37.Frank ME, Wada Y, Makino J, Mizutani M, Umezawa H, Katsuie Y, Hettinger TP, Blizard DA. Variation in intake of sweet and bitter solutions by inbred strains of hamsters. Behav Genetics. 2004;34:465–476. doi: 10.1023/B:BEGE.0000023651.99481.d5. [DOI] [PubMed] [Google Scholar]
- 38.Fukami H, Bradley RM. Biophysical and morphological properties of parasympathetic neurons controlling the parotid and von Ebner salivary glands in rats. J Neurophysiol. 2005;93:678–686. doi: 10.1152/jn.00277.2004. [DOI] [PubMed] [Google Scholar]
- 39.Fukushima N, Oikawa S, Yokouchi K, Kawagishi K, Moriizumi T. The minimum number of neurons in the central olfactory pathway in relation to its function: a retrograde fiber tracing study. Chem Senses. 2002;27:1–6. doi: 10.1093/chemse/27.1.1. [DOI] [PubMed] [Google Scholar]
- 40.Ganchrow D, Ganchrow JR, Verdin-Alcazar M, Whitehead MC. Brain-derived neurotrophic factor-, neurothrophin-3-, and tyrosine kinase receptor-like immunoreactivity in lingual taste bud fields of mature hamster after sensory denervation. J Comp Neurol. 2003;455:25–39. doi: 10.1002/cne.2164. [DOI] [PubMed] [Google Scholar]
- 41.Gattermann R, Fritzsche P, Neumann K, Al-Hussein I, Kayser A, Abiad M, Yakti R. Notes on the current distribution and the ecology of wild golden hamsters (Mesocricetus auratus) J Zool Lond. 2001;254:359–365. [Google Scholar]
- 42.Geran LC, Travers SP. Single neurons in the nucleus of the solitary tract respond selectively to bitter taste stimuli. J Neurophysiol. 2006;96:2513–2527. doi: 10.1152/jn.00607.2006. [DOI] [PubMed] [Google Scholar]
- 43.Glendinning JI, Brown H, Capoor M, Davis A, Gbenemah A, Long E. A peripheral mechanism for behavioral adaptation to specific “bitter” taste stimuli in an insect. J Neurosci. 2001;21:3688–3696. doi: 10.1523/JNEUROSCI.21-10-03688.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gosselin RE, Smith RP, Hodge HC. Clinical toxicology of commercial products. Baltimore: Williams & Witkins; 1984. [Google Scholar]
- 45.Harder DB, Maggio JC, Whitney G. Assessing gustatory detection capabilities using preference procedures. Chem Senses. 1989;14:547–564. [Google Scholar]
- 46.Hermans E, Challis RAJ. Structural, signaling and regulatory properties of the group I metabotropic glutamate receptors: prototypic family C G-protein-coupled receptors. Biochem J. 2001;359:465–484. doi: 10.1042/0264-6021:3590465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hill DL, Formaker BK, White KS. Perceptual characteristics of the amiloride-suppressed sodium chloride taste response in the rat. Behav Neurosci. 1990;104:734–741. doi: 10.1037//0735-7044.104.5.734. [DOI] [PubMed] [Google Scholar]
- 48.Houpt TA, Philopena JM, Joh TH, Smith GP. c-Fos induction in the rat nucleus of the solitary tract by intraoral quinine infusion depends on prior contingent pairing of quinine with lithium chloride. Physiol Behav. 1996;60:1535–1541. doi: 10.1016/s0031-9384(96)00326-5. [DOI] [PubMed] [Google Scholar]
- 49.Inoue M, Li X, McCaughey SA, Beauchamp GK, Bachmanov AA. Soa genotype selectively affects mouse gustatory neural responses to sucrose octaacetate. Physiol Genomics. 2001;5:181–186. doi: 10.1152/physiolgenomics.2001.5.4.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Johnson F, Starkovsky NA, Paton AC, Carlson AA. The total synthesis of cycloheximide. J Amer Chem Soc. 1966;88:149–159. doi: 10.1021/ja00953a028. [DOI] [PubMed] [Google Scholar]
- 51.Keung WM, Vallee BL. Kudzu root: an ancient Chinese source of modern antidipsotropic agents. Phytochem. 1998;47:499–506. doi: 10.1016/s0031-9422(97)00723-1. [DOI] [PubMed] [Google Scholar]
- 52.Kim JW, Robers C, Maruyama Y, Berg S, Roper S, Chaudhari N. Faithful expression of GFP from the PLCB2 promoter in a functional class of taste receptor cells. Chem Senses. 2006;31:213–219. doi: 10.1093/chemse/bjj021. [DOI] [PubMed] [Google Scholar]
- 53.Kim M, Chiego DJ, Jr, Bradley RM. Morphology of parasympathetic neurons innervating rat lingual salivary glands. Autonom Neurosci. 2004;111:27–36. doi: 10.1016/j.autneu.2004.01.006. [DOI] [PubMed] [Google Scholar]
- 54.Kim UK, Drayna D. Genetics of individual differences in bitter taste perception: lessons from the PTC gene. Clin Genet. 2004;67:275–280. doi: 10.1111/j.1399-0004.2004.00361.x. [DOI] [PubMed] [Google Scholar]
- 55.Kim U, Jorgenson E, Coon H, Leppert M, Risch N, Drayna D. Positional cloning of the human quantitative trait locus underlying taste sensitivity to phenylthiocarbamide. Science. 2003;299:1221–1225. doi: 10.1126/science.1080190. [DOI] [PubMed] [Google Scholar]
- 56.Kim U, Wooding S, Ricci D, Jorde LB, Drayna D. Worldwide haplotype diversity and coding sequence variation at human bitter taste receptor loci. Human Mutation. 2005;26:199–204. doi: 10.1002/humu.20203. [DOI] [PubMed] [Google Scholar]
- 57.Kimura Y, Miwa T, Furakawa M, Umeda R. Effect of topical application of steroid on olfactory disturbance in mice. Chem Senses. 1991;16:297–302. [Google Scholar]
- 58.King CT, Garcea M, Spector AC. Glossopharyngeal nerve regeneration is essential for the complete recovery of quinine-stimulated oromotor rejection behaviors and central patterns of neuronal activity in the nucleus of the solitary tract in the rat. J Neurosci. 2000;20:8426–8434. doi: 10.1523/JNEUROSCI.20-22-08426.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.King CT, Garcea M, Stolzenberg DS, Spector AC. The consequences of cross-regeneration of the lingual gustatory nerves on quinine-stimulated gaping and Fos-like immunoreactivity in the nucleus of the solitary tract in rats. ISOT/JASTS. 2004;P-154:118. [Google Scholar]
- 60.Krautwurst D, Yau KW, Reed RR. Identification of ligands for olfactory receptors by functional expression of a receptor library. Cell. 1998;95:917–926. doi: 10.1016/s0092-8674(00)81716-x. [DOI] [PubMed] [Google Scholar]
- 61.Kulkosky PJ, Cornell NW. Free-choice ethanol intake and ethanol metabolism in the hamster and rat. Pharmac Biochem Behav. 1979;11:439–444. doi: 10.1016/0091-3057(79)90121-7. [DOI] [PubMed] [Google Scholar]
- 62.Lee SF, Chen ZY, Fong WP. Gender difference in enzymes related with alcohol consumption in hamster, and avid consumer of alcohol. Comp Biochem Physiol C. 2001;129:285–293. doi: 10.1016/s1532-0456(01)00205-8. [DOI] [PubMed] [Google Scholar]
- 63.Lemon CH, Smith DV. Neural representation of bitter taste in the nucleus of the solitary tract. J Neurophysiol. 2005;94:3719–3729. doi: 10.1152/jn.00700.2005. [DOI] [PubMed] [Google Scholar]
- 64.Li CS, Cho YK, Smith DV. Modulation of parabrachial taste neurons by electrical and chemical stimulation of the lateral hypothalamus and amygdala. J Neurophysiol. 2005;93:1183–1196. doi: 10.1152/jn.00828.2004. [DOI] [PubMed] [Google Scholar]
- 65.Lindemann B. Receptors and transduction in taste. Nature. 2001;413:219–225. doi: 10.1038/35093032. [DOI] [PubMed] [Google Scholar]
- 66.Lush IE. The genetics of tasting in mice. I. Sucrose octaacetate. Genet Res. 1981;38:93–95. doi: 10.1017/s0016672300020425. [DOI] [PubMed] [Google Scholar]
- 67.Lush IE. Differences between mouse strains in their consumption of phenylthiourea (PTC) Heredity. 1986;57:319–323. doi: 10.1038/hdy.1986.129. [DOI] [PubMed] [Google Scholar]
- 68.Lush IE, Holland G. The genetics of tasting in mice. V. Glycine and cycloheximide. Genet Res. 1988;52:207–212. doi: 10.1017/s0016672300027671. [DOI] [PubMed] [Google Scholar]
- 69.Makalowski W, Zhang J, Boguski MS. Comparative analysis of 1196 orthologous mouse and human full-length mRNA and protein sequences. Genome Res. 1996;6:846–857. doi: 10.1101/gr.6.9.846. [DOI] [PubMed] [Google Scholar]
- 70.Mark M, Herlitze S. G-protein mediated gating of inward-rectifier K+ channels. Eur J Biochem. 2000;267:5830–5836. doi: 10.1046/j.1432-1327.2000.01670.x. [DOI] [PubMed] [Google Scholar]
- 71.Matsunami H, Montmayeur JP, Buck LB. A family of candidate taste receptors in human and mouse. Nature. 2000;404:601–604. doi: 10.1038/35007072. [DOI] [PubMed] [Google Scholar]
- 72.Windholz M, Budavari S, Stroumtsos LY, Fertig MN, editors. Merck Index. 9. Vol. 2733. Merck & Co; 1976. p. 357. [Google Scholar]
- 73.Moriizumi T, Tsukatani T, Sakashita H, Miwa T. Olfactory disturbance induced by deafferentation of serotonergic fibers in the olfactory bulb. Neurosci. 1994;61:733–788. doi: 10.1016/0306-4522(94)90396-4. [DOI] [PubMed] [Google Scholar]
- 74.Mueller KL, Hoon MA, Erlenbach I, Chandrashekar J, Zuker CS, Ryba NJP. The receptors and coding logic for bitter taste. Nature. 2005;434:225–229. doi: 10.1038/nature03352. [DOI] [PubMed] [Google Scholar]
- 75.Nachman M. Learned aversions to the taste of lithium chloride and generalization to other salts. J Comp Physiol Psychol. 1963;56:343–349. doi: 10.1037/h0046484. [DOI] [PubMed] [Google Scholar]
- 76.Nachman M, Ashe JH. Learned taste aversions in rats as a function of dosage, concentration, and route of administration of LiCl. Physiol Behav. 1973;10:73–78. doi: 10.1016/0031-9384(73)90089-9. [DOI] [PubMed] [Google Scholar]
- 77.Nebert DW, Russell DW. Clinical importance of the cytochromes P450. Lancet. 2002;360:1155–1162. doi: 10.1016/S0140-6736(02)11203-7. [DOI] [PubMed] [Google Scholar]
- 78.Nelson TM, Munger SD, Boughter JD., Jr Taste sensitivities to PROP and PTC vary independently in mice. Chem Senses. 2003;28:695–704. doi: 10.1093/chemse/bjg062. [DOI] [PubMed] [Google Scholar]
- 79.Nelson TM, Munger SD, Boughter JD., Jr Haplotypes at the Tas2r locus on chromosome 6 vary with quinine taste sensitivity in inbred mice. BMC Gen. 2005;6:32. doi: 10.1186/1471-2156-6-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Neumann K, Michaux J, Ledbedev V, Yigit N, Colak E, Ivanova N, Poltoraus A, Surov A, Markov G, Maak S, Neumann S, Gattermann R. Molecular phylogeny of the Cricetinae subfamily based on the mitochondrial cytochrome b and 12S rRNA genes and the nuclear vWF gene. Mol Phylogenet Evol. 2006;29:135–148. doi: 10.1016/j.ympev.2006.01.010. [DOI] [PubMed] [Google Scholar]
- 81.Ninomiya Y, Higashi T, Katsukawa H, Mizukoshi T, Funakoshi M. Qualitative discrimination of gustatory stimuli in three different strains of mice. Brain Res. 1984;322:83–92. doi: 10.1016/0006-8993(84)91183-1. [DOI] [PubMed] [Google Scholar]
- 82.Nowlis GH. Conditioned stimulus intensity and acquired alimentary aversions in the rat. J Comp Physiol Psychol. 1974;86:1173–1184. doi: 10.1037/h0037644. [DOI] [PubMed] [Google Scholar]
- 83.Nowlis GH, Frank ME, Pfaffmann C. Specificity of acquired aversions to taste qualities in hamsters and rats. J Comp Physiol Psychol. 1980;94:932–942. doi: 10.1037/h0077809. [DOI] [PubMed] [Google Scholar]
- 84.Omura K, Takagi SF, Harada O. Mechanism of the repellent action of naramycin to rats. Gunma J Med Sci. 1961;10:217–227. [Google Scholar]
- 85.Poor A. Cricetinae. [accessed 03-24-2006];(On-line) Animal Diversity Web. 2005 http://animal diversity.ummz.umich.edu.
- 86.Prapaipong H, Berenbaum MR, Schuler MA. Transcriptional regulation of the Papilio polyxenes CYPB1 gene. Nuc Acids Res. 1994;22:3210–3217. doi: 10.1093/nar/22.15.3210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Rehnberg BG, Hettinger TP, Frank ME. The role of sucrose-sensitive neurons in ingestion of sweet stimuli by hamsters. Physiol Behav. 1990;48:459–466. doi: 10.1016/0031-9384(90)90344-4. [DOI] [PubMed] [Google Scholar]
- 88.Renwick JAA. Variable diets and changing taste in plant-insect relationships. J Chem Ecol. 2001;27:1063–1076. doi: 10.1023/a:1010381509601. [DOI] [PubMed] [Google Scholar]
- 89.Risinger FO, Boyce JM. Conditioning tastant and the acquisition of conditioned taste avoidance to drugs of abuse in DBA/2J mice. Psychopharm. 2002;160:225–232. doi: 10.1007/s00213-001-0973-2. [DOI] [PubMed] [Google Scholar]
- 90.Rodgers DA, McClearn GE. Sucrose versus ethanol appetite in inbred strains of mice. Quart J Stud Alc. 1964;25:26–35. [PubMed] [Google Scholar]
- 91.Sbarati A, Osculati F. The taste cell-related diffuse chemosensory system. Prog Neurobiol. 2005;75:295–307. doi: 10.1016/j.pneurobio.2005.03.001. [DOI] [PubMed] [Google Scholar]
- 92.Sewards TV. Dual sensory pathways for sensory and hedonic aspects of taste. Brain Res Bull. 2004;62:271–283. doi: 10.1016/j.brainresbull.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 93.Shi P, Zhang J. Contrasting modes of evolution between vertebrate sweet/umami receptor genes and bitter receptor genes. Mol Biol Evol. 2006;23:292–300. doi: 10.1093/molbev/msj028. [DOI] [PubMed] [Google Scholar]
- 94.Shi P, Zhang J, Yang H, Zhang Y. Adaptive diversification of bitter taste receptor genes in mammalian evolution. Mol Biol Evol. 2003;20:805–814. doi: 10.1093/molbev/msg083. [DOI] [PubMed] [Google Scholar]
- 95.Steppan SJ. Murinae v1. [accessed 03-23-2006];(On-line) Tree of Life Web. 2005 http://tol.org.
- 96.Steppan SJ, Adkins RM, Anderson J. Phylogeny and divergence-date estimates of rapid radiations in muroid rodents based on multiple nuclear genes. Systematic Biol. 2004;53:533–553. doi: 10.1080/10635150490468701. [DOI] [PubMed] [Google Scholar]
- 97.St John SJ, Pour L, Boughter JD., Jr Phenylthiocarbamide produces conditioned taste aversions in mice. Chem Senses. 2005;30:377–382. doi: 10.1093/chemse/bji032. [DOI] [PubMed] [Google Scholar]
- 98.St John SJ, Spector AC. Behavioral discrimination between quinine and KCl is dependent on the seventh cranial nerve: implications for the functional roles of the gustatory nerves in rats. J Neurosci. 1998;18:4353–4362. doi: 10.1523/JNEUROSCI.18-11-04353.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Sugita M, Shiba Y. Genetic tracing shows segregation of taste neuronal circuitries for bitter and sweet. Science. 2005;309:781–785. doi: 10.1126/science.1110787. [DOI] [PubMed] [Google Scholar]
- 100.Taketo M, Schroeder AC, Mobraaten LE, Gunning KB, Hanten G, Fox RR, Roderick TH, Stewart CL, Lilly F, Hansen CT, Overbeek PA. FVB/N: An inbred mouse strain preferable for transgenic analyses. 1991;88:2065–2069. doi: 10.1073/pnas.88.6.2065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Tobach E, Bellin JS, Das DK. Differences in bitter taste perception in three strains of rats. Behav Genet. 1974;4:405–410. doi: 10.1007/BF01066160. [DOI] [PubMed] [Google Scholar]
- 102.Waksman SA, Lechavalier HA. The actinomycetes, vol III, antibiotics of actinomycetes. Baltimore: Williams & Wilkins; 1962. pp. 248–252. [Google Scholar]
- 103.Wang L, Chen L, Jacob T. Evidence for peripheral plasticity in human odour response. J Physiol. 2003;554:236–244. doi: 10.1113/jphysiol.2003.054726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Wang H, Wysocki CJ, Gold JH. Induction of olfactory receptor sensitivity in mice. Science. 1993;260:998–1000. doi: 10.1126/science.8493539. [DOI] [PubMed] [Google Scholar]
- 105.Weir TL, Park SW, Vivanco JM. Biochemical and physiological mechanisms mediated by allelochemicals. Cur Opin Plant Biol. 2004;7:472–479. doi: 10.1016/j.pbi.2004.05.007. [DOI] [PubMed] [Google Scholar]
- 106.Welch JF. Rodent control: a review of chemical repellents for rodents. J Agric Food Chem. 1954;2:142–149. [Google Scholar]
- 107.Wilson-Pauwels L, Akesson EJ, Stewart PA. Cranial nerves, anatomical and clinical comments. Philadelphia: BC Decker Inc; 1988. [Google Scholar]
- 108.Wong R. Response latency of gerbils and hamsters to nuts flavoured with bitter tasting substances. Q J Exp Psychol B. 1994;47:173–186. [PubMed] [Google Scholar]
- 109.Wu SV, Chen MC, Rozengurt E. Genomic organization, expression, and function of bitter taste receptors (T2R) in mouse and rat. Physiol Genomics. 2005;22:139–149. doi: 10.1152/physiolgenomics.00030.2005. [DOI] [PubMed] [Google Scholar]
- 110.Wu SV, Rozengurt N, Yang M, Young SH, Sinnett-Smith J, Rozengurt E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc Natl Acad Sci (USA) 2002;99:2392–2397. doi: 10.1073/pnas.042617699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Yamamoto T. Electrophysiology of CTA. In: Bures J, Bermudez-Rattoni F, Yamamoto T, editors. Conditioned taste aversion: memory of a special kind. New York: Oxford University Press; 1998. pp. 76–91. [Google Scholar]
- 112.Zhang X, Firestein S. The olfactory receptor gene superfamily of the mouse. Nature Neurosci. 2002:124–133. doi: 10.1038/nn800. [DOI] [PubMed] [Google Scholar]
- 113.Zhang Y, Hoon MA, Chandrashekar J, Mueller KL, Cook B, Wu D, Zuker CS, Ryba NJP. Coding of sweet, bitter, and umami tastes: different receptor cells sharing similar signaling pathways. Cell. 2003;112:293–301. doi: 10.1016/s0092-8674(03)00071-0. [DOI] [PubMed] [Google Scholar]
- 114.Zhao GG, Zhang Y, Hoon MA, Chandrashekar J, Erlenbach I, Ryba NJP, Zuker CS. The receptors for mammalian sweet and umami taste. Cell. 2003;115:255–266. doi: 10.1016/s0092-8674(03)00844-4. [DOI] [PubMed] [Google Scholar]
- 115.Zhou P. Determining protein half-lives. Methods Mol Biol. 2004;284:67–77. doi: 10.1385/1-59259-816-1:067. [DOI] [PubMed] [Google Scholar]