

Abstract
β-1,4-Endoglucanases (EGases, EC 3.2.1.4) degrade polysaccharides possessing β-1,4-glucan backbones such as cellulose and xyloglucan and have been found among extremely variegated taxonomic groups. Although many animal species depend on cellulose as their main energy source, most omnivores and herbivores are unable to produce EGases endogenously. So far, all previously identified EGase genes involved in the digestive system of animals originate from symbiotic microorganisms. Here we report on the synthesis of EGases in the esophageal glands of the cyst nematodes Globodera rostochiensis and Heterodera glycines. From each of the nematode species, two cDNAs were characterized and hydrophobic cluster analysis revealed that the four catalytic domains belong to family 5 of the glycosyl hydrolases (EC 3.2.1, 3.2.2, and 3.2.3). These domains show 37–44% overall amino acid identity with EGases from the bacteria Erwinia chrysanthemi, Clostridium acetobutylicum, and Bacillus subtilis. One EGase with a bacterial type of cellulose-binding domain was identified for each nematode species. The leucine-rich hydrophobic core of the signal peptide and the presence of a polyadenylated 3′ end precluded the EGases from being of bacterial origin. Cyst nematodes are obligatory plant parasites and the identified EGases presumably facilitate the intracellular migration through plant roots by partial cell wall degradation.
Keywords: endoglucanase, Globodera, Heterodera, in situ hybridization, secretory protein
The most abundant carbohydrate polymer on earth, cellulose, is mainly produced by terrestrial plants and marine algae. This recalcitrant polymer is used as a food source by many organisms. In plants, cellulose is a main component of the cell walls, in which it predominantly occurs in long rigid microfibrils. These microfibrils each consist of several dozen parallelly oriented linear chains of (1 → 4) β-linked d-glucose showing a varying and plant species-dependent degree of crystallinity. The cellulose microfibrils are embedded in a matrix of lignin and hemicellulose. The latter includes either xyloglucans (type I primary cell walls) or glucuronoarabinoxylans (type II primary cell walls) (1). To exploit the energy and carbon available in cellulose, organisms such as fungi and bacteria produce mixtures of synergistically acting cellulases (2, 3). Many cellulases share a common basic architecture composed of a catalytic domain linked to a cellulose binding domain by a glycosylated, Pro/Thr/Ser-rich peptide (4). The noncatalytic cellulose-binding domain (CBD) determines the efficiency of degradation of insoluble cellulose (5).
Although cellulose is a major food source for many animal species, most omnivores and herbivores do not produce cellulases themselves and live in symbiosis with cellulolytic microorganisms. For example, ruminants have highly specialized digestive tracts where mixtures of bacteria and protozoa degrade cellulose under anaerobic conditions (6). In most nonruminant herbivous mammals and birds, cellulose digestion is mediated by microorganisms, too. The ability of animals to hydrolyze cellulose in the absence of microorganisms has been the subject of various studies. However, it is often difficult to establish whether cellulase is synthesized by an animal or by microorganisms without isolating the corresponding genes. To date, no animal cellulase genes have been identified and endogenous cellulase production by animals seems exceptional. Indications for the existence of cellulase genes within the animal kingdom have been obtained by measuring cellulase activity in microbe-free digestive tracts of cockroaches, higher termites, grass carps, eri silkworms, and earthworms (7–11).
This paper describes the characterization of four cellulase genes from obligatory plant–parasitic cyst nematodes of the genera Globodera and Heterodera. These cellulases are thought to play a role during intracellular migration of the larvae through the root cortex to the vascular cylinder where they modify selected plant cells into a multinucleate feeding cell (12). Nematodes are equipped with a protrusible hollow stylet through which secretions from the esophageal glands are released. These secretions are the subject of intensive study because of their putative roles in plant pathogenesis (13). A mAb (MGR 48), recognizing an antigen(s) within the subventral esophageal glands from the cyst nematodes Globodera rostochiensis and Heterodera glycines (14, 15), was employed to immunopurify endoglucanases (EGases). Evidence is provided that both in G. rostochiensis and H. glycines EGases are produced endogenously.
MATERIALS AND METHODS
Materials.
Nematodes were cultured on host plant roots and preparasitic second stage juveniles (J2) were extracted as described (14, 16). All reagents were of analytical grade.
Purification and N-Terminal Sequencing.
In homogenates of G. rostochiensis mAb MGR48 reacted on Western blots with four protein bands (31, 32, 39, and 49 kDa; ref. 14), whereas in H. glycines only one protein band (49 kDa) was recognized. From each nematode species, one protein band was selected for purification and N-terminal sequencing. Approximately 1.0 ml of preparasitic J2 (≈2.106 J2/ml) of each G. rostochiensis and H. glycines were homogenized in 10 mM Tris⋅Cl (pH 7.4) including 3 mM Pefabloc (Boehringer Mannheim) and the supernatant was separated by preparative continuous flow SDS/PAGE (Bio-Rad) on a 10% denaturing polyacrylamide gel (14). Immunoblotting with mAb MGR48 was used to identify successive fractions containing the 39-kDa subventral gland protein of G. rostochiensis and the 49-kDa subventral gland protein of H. glycines. For each protein, these fractions were pooled and subsequently purified on a MGR48 immunoaffinity chromatography column (17). The immunopurified proteins (svp39 and svp49) were dialyzed in 0.1 M NH4HCO3 and lyophilized. The purified proteins were subjected to SDS/PAGE by using a Tris–tricine buffer system and electroblotted on a polyvinylidene fluoride membrane (18). Blotted protein bands were stained with Coomassie brilliant blue R-250 and cut out for protein sequencing (ARIAD Pharmaceuticals, Cambridge, MA). N-terminal blocking of the proteins was circumvented by o-phthalaldehyde treatment and CNBr digestion.
cDNA Isolation and Characterization.
A degenerative gene-specific primer (HG1f) designed from the N-terminal amino acid sequence of the 49-kDa subventral gland protein of H. glycines (Table 1) was used in 3′ rapid amplification of cDNA ends (3′ RACE; GIBCO/BRL). Template RNA for 3′ RACE was extracted from J2 in 4 M guanidinium isothiocyanate buffer (19) and purified on a cesium trifluoroacetate cushion (Pharmacia). Gene-specific primers to the initial 3′ RACE products were designed and used in a 5′ RACE protocol (GIBCO/BRL). Full-length cDNA clones were obtained by using the 5′ RACE products to probe plaque lifts of a H. glycines (J2) oligo-dT-primed cDNA library (Uni-Zap; Strategene, ref. 19). Two full-length cDNAs named HG-eng-1 and HG-eng-2 were isolated from Escherichia coli and sequenced on an Automated Laser Fluorescent DNA sequencer (Pharmacia).
Table 1.
Partial amino acid sequences of purified nematode secretory proteins and deduced oligonucleotide primers used to amplify and clone EGase genes from G. rostochiensis and H. glycines
| Nematode/primer* | Protein/DNA | Sequence |
|---|---|---|
| Amino acid sequence† | ||
| H. glycines | svp49 (N terminal) | ASAVAPPFGQLSVSGSNLVGANKQPVQLISNSLFEL |
| G. rostochiensis | svp39 (N terminal) | LTATPPPYGQLSVSGTKLVGSMGQPVQLIGNSLFWPQFQGQYWNA |
| G. rostochiensis | svp39 (internal) | PTATYNLAVAVIEAAISQGMYVIVDTHSTEAHAD |
| DNA sequence of deduced oligonucleotide primer‡ | ||
| HG1f | svp49 | 5′-GCIGTIGCICCICCITTYGGICA-3′ |
| GR1f | svp39 (N terminal) | 5′-ACIGCIACICCICCICCITAYGGNCA-3′ |
| GR2f | 5′-CAGTGAAGGCRCTCAAATGCAATTGG-3′ | |
| GR3f | 5′-CTTCCGTGTCTTCCTCCTCCATTG-3′ | |
| GR4f | 5′-TGGGATCCACTGCCACGCCTCCCCCA-3′ | |
| HG2r | 5′-ATAGCCGCCCTCGTCCAC-3′ | |
| GR5r | svp39 (internal) | 5′-TGCCARTCIACDATNACRTACTKCC-3′ |
| GR6r | 5′-CCAATTGCATTTGAGYGCCTTCACTG-3′ | |
| GR7r | 5′-GGAAGCTTGCGCAACCACTTTTTTATCATCATC-3′ | |
| GR8r | 5′-GGAAGCTTCGCAACAATTTTATCGTCAATAAATT-3′ | |
| pCDNA5f | pCDNAII | 5′-GGTGACACTATAGAATACTCAAGCTATGCA-3′ |
| pCDNA3r | pCDNAII | 5′-GACGGCCAGTGAATTGTAATACGACTCACT-3′ |
The forward (f) or reverse (r) orientation of the oligonucleotide primers is indicated in their names.
The single character amino acid code is used.
I - Inosine, K = G + T, N = A + C + G + T, R = A + G, Y = C + T; underlining indicates endonuclease restriction site.
Similarly, an oligo-dT-primed cDNA library from G. rostochiensis was prepared in the vector pcDNAII (Invitrogen). Two gene-specific primers derived from amino acid sequences obtained by protein sequencing were used for initial amplifications (Table 1). The 5′ and 3′ ends of the library inserts were isolated by using gene-specific primers combined with primers derived from flanking sequences in the library vector pCDNAII. Two cDNAs, GR-eng-1 and GR-eng-2, were isolated from a plasmid prep (Promega) of the cDNA library and cloned into pCR2.1 (Invitrogen).
Heterologous Expression in E. coli.
HG-eng-1 and HG-eng-2 were cloned into the expression vector pET-28c and pET28a, respectively, and overexpressed and purified according to the manufacturer (Novagen). GR-eng-1 and GR-eng-2 were cloned into pMAL-c2 expression vector by using adapter-primers (GR4f, GR7r, and GR8r; Table 1). Overexpression and affinity purification of the fusion proteins was done according to the manufacturer (New England Biolabs).
Carboxymethylcellulose (CMC) Assay.
CMC hydrolase activity in both G. rostochiensis and H. glycines J2 was assayed with a nonreducing SDS/PAGE gel overlay method (20). An identical gel was transferred to nitrocellulose membrane and probed with MGR48 for comparison of banding pattern (14). A semi-quantitative CMC cup plate assay was used to determine the CMC hydrolase activity of affinity-purified heterologous fusion proteins (21).
In Situ Hybridization.
Digoxigenin-11-UTP-labeled RNA probes were transcribed from the nucleotide 273–447 region of the HG-eng-2 cDNA. J2 of H. glycines were fixed overnight in 2% paraformaldehyde, cut into sections, and permeabilized as described (14). The J2 sections were then hybridized with sense or antisense riboprobe. Following digestion with RNAse A and stringency washings, bound riboprobe was detected by alkaline phosphatase staining (Genius kit, Boehringer Mannheim). The stained J2 were examined with differential interference contrast microscopy.
Production of Antisera.
For the subcutaneous immunization of BALB/c mice, partially purified fusion proteins were mixed 1:1 with Freund’s complete adjuvant (17). All four antisera raised against the fusion proteins were assayed with immunofluorescence microscopy and Western blotting for binding to nematode esophageal gland proteins (14, 16).
Hydrophobic Cluster Analysis (HCA).
HCA plots were made by using the program HCA-PLOT V3.0 (Doriane, Le Chesnay, France). In these plots, the amino acid sequence of the proteins is drawn on a duplicated helical net using the standard one-letter code except for Pro, Gly, Thr, and Ser, which are represented by stars, diamonds, squares, and pointed stars, respectively. Clusters of hydrophobic residues (V, I, L, F, W, M, and Y) are automatically drawn on the bidimensional helical representation. Analysis of the plots was performed as described in Results (22).
RESULTS
mAb MGR48 was used to immunopurify the subventral gland secretory proteins svp39 and svp49 from G. rostochiensis (14) and H. glycines, respectively. Five to ten micrograms of these proteins was used for amino acid sequencing (Table 1). Two sequences (one N terminal and one internal) were determined for the G. rostochiensis protein svp39. An N-terminal amino acid sequence was obtained with svp49 purified from H. glycines. The amino acid sequences presented in Table 1 were used to design gene-specific PCR primers.
Cloning of Two EGases from H. glycines.
PCR products from 3′ RACE using forward primer HG1f (Table 1) and 5′ RACE using reverse primer HG2r were used for screening of a cDNA library from preparasitic J2 of H. glycines. A large cDNA clone obtained (HG-eng-1) comprised 1,615 bp with an ORF of 1,428 bp, encoding a putative 49,800 Da protein (HG-ENG-1). A smaller cDNA clone comprised 1,191 bp (HG-eng-2) with a 957-bp ORF encoding a putative 34,700 Da protein (HG-ENG-2). A predicted amino acid segment nearly identical to the N-terminal sequence of the purified svp49 protein was identified downstream of the alanines at the positions 108 and 121 in HG-eng-1 and HG-eng-2, respectively. The regions between the putative ATG start codons (positions 60 and 64 in HG-eng-1 and HG-eng-2, respectively) and the N termini were identified as signal peptides (23). In HG-ENG-1, the most likely cleavage site was between amino acids Ser–Leu at the amino acid positions 26 and 27 and between Val–Asp at positions 16 and 17 in HG-ENG-2.
Cloning of Two EGases from G. rostochiensis.
Two primers (GR1f and GR5r; Table 1) were designed from the N-terminal and internal amino acid sequences obtained by protein sequencing of svp39. By using these primers, a 300-bp DNA fragment was amplified from a cDNA library. Based on its sequence, two additional gene-specific primers (GR2f and GR6r) were synthesized in opposite orientation. In theory, use of these primers should have resulted in two overlapping DNA fragments when combined with either a reverse (pCDNA3r) or a forward (pCDNA5f) primer specific for the flanking region in the vector pCDNAII. However, two products of 1,450 and 1,200 bp were amplified by using GR2f and pCDNA3r. In the opposite orientation, primers GR6r and pCDNA5f only resulted in a single 250-bp PCR fragment. Three additional oligonucleotide primers (Table 1) were synthesized with two cDNAs, 1,546 bp (GR-eng-1) and 1,300 bp (GR-eng-2) that were isolated. No putative ATG start codon at the 5′ end was observed in GR-eng-1; therefore, it is presumably not present in the cDNA library as a full-length mRNA. The largest ORFs in these two cDNA fragments were 1,414 bp and 1,179 bp, respectively, encoding two putative precursor proteins of 49,700 Da (GR-ENG-1) and 42,000 Da (GR-ENG-2). In both clones, a peptide sequence nearly identical to the N-terminal protein sequence of purified svp39 was present. Upstream of this amino acid sequence, a signal peptide was predicted for both proteins (23). The most likely cleavage site was between amino acids Ala and Leu at positions 17 and 18 in GR-ENG-1 and at Ala and Leu at positions 24 and 25. These predictions correspond to the results of the N-terminal amino acid sequence obtained by protein sequencing of the svp39 protein.
Sequence Comparison.
The two cDNA clones of G. rostochiensis shared a high degree of similarity at their 5′ ends (95% identity in the first 884 nucleotides). The two cDNA clones isolated from H. glycines were less similar at their 5′ ends (79% identity in the first 873 nucleotides). The four encoded proteins were compared with sequences currently available in protein databases by using the fasta search method (24). The search result showed that the nematode proteins shared an overall identity of 37–44% with β-1,4-endoglucanases of various bacterial origins including Erwinia chrysanthemi, C. acetobutylicum, and B. subtilis (Swiss-Prot accession nos. P07103, P15704, and P07983, respectively).
Fig. 1A presents the global architecture of the four nematode EGase precursor proteins with the amino acid positions of the functional domains. In HG-ENG-1 and GR-ENG-1, C-terminal extensions of 96 and 97 amino acid residues, which are missing in HG-ENG-2 and GR-ENG-2, respectively, show significant similarity to type II CBD (25). The alignment of these C-terminal extensions with two representatives of this bacterial type of cellulose binding domain is illustrated in Fig. 1B.
Figure 1.

(A) Global architecture of the four nematode β-1,4-endoglucanase precursor proteins. The amino acid positions of the functional domains are indicated above. (B) Sequence alignment of the C-terminal cellulose binding domains of G. rostochiensis ENG-1 and H. glycines ENG-1 with two representatives of the bacterial type of cellulose binding domains [Thermomonospora fusca EGase E-5 (THEFU), GenBank accession no. L01577, and Clostridium cellulovorans EGase D (CLOCL), GenBank accession no. M37434]. Residues conserved in all four cellulose binding domains are shaded.
EGase Activity Assays.
Fusion proteins of the predicted sizes, 54 kDa for HG-ENG-1 and 38 kDa for HG-ENG-2, expressed in E. coli were recognized by mAb MGR48 on Western blots. Similarly, fusion proteins of the predicted sizes, 90 kDa for GR-ENG-1 and 82 kDa for GR-ENG-2, expressed in E. coli were recognized on Western blots by a mixture of subventral gland protein-specific mAbs (14) (Fig. 2A).
Figure 2.

Detection of recombinant nematode EGases in E. coli lysates on Western blot (A) and CMC hydrolysis assays (B). (A) Immunodetection was done with subventral gland specific mAbs (17). Lanes: 1, HG-ENG-1 produced from pET28c; 2, pET28c control; 3, HG-ENG-2 produced from pET28a; 4, pET28a control; 5, GR-ENG-1 from vector pMAL-c2; 6, GR-ENG-2 from vector pMAL-c2; 7, pMAL-c2 control. (B) Detection of CMCase activity (halo) in affinity-purified heterologous cyst nematode EGases that correspond to lanes above.
Applying affinity-purified preparations of the four recombinant ENG proteins in a CMC plate assay showed significant hydrolytic activity (Fig. 2B). Similarly, homogenates of both nematode species were analyzed in a PAGE gel overlay on an agarose bed containing CMC (Fig. 3B). In G. rostochiensis homogenate, a distinct band of approximately 50 kDa showed CMC hydrolase activity, whereas in H. glycines, a slightly smaller protein hydrolyzed CMC. Both apparent CMC hydrolases aligned with one of the subventral gland specific antigens (≈49 kDa) recognized by MGR48. No CMCase activity was detected for svp39 in G. rostochiensis homogenates. This is presumably caused by the detrimental effects of SDS. When compared with a standard SDS/PAGE (14), the SDS concentration in the electrophoresis buffer and polyacrylamide gels was significantly reduced. This may account for the minor differences in the reaction pattern of MGR48 on Western blot (Fig. 3A).
Figure 3.
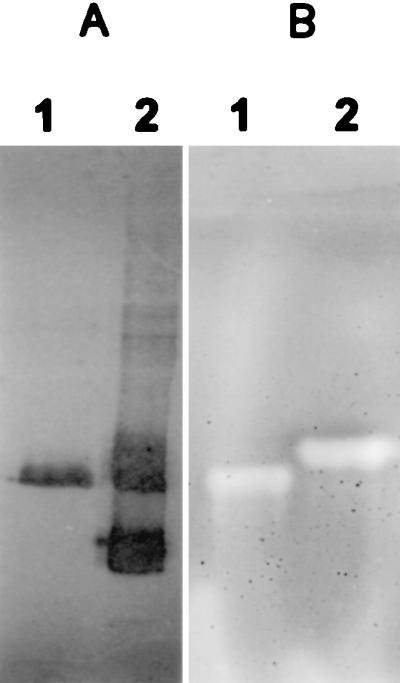
Comparative nonreducing SDS/PAGE of nematode homogenates following either Western blotting (A) or CMC overlay (B). (A) mAb MGR48 reacting with subventral gland protein of H. glycines (lane 1) and G. rostochiensis (lane 2). (B) Congo red stained agarose bed containing CMC showing hydrolysis activity (halo) by specific proteins of H. glycines (lane 1) and G. rostochiensis (lane 2).
Localization of EGase Transcripts and Protein.
EGase transcripts were detected by whole-mount in situ hybridization in the subventral gland cells of H. glycines. Antisense RNA probes transcribed from the HG-eng-2 cDNA bound specifically within the subventral glands of preparasitic J2s of H. glycines (Fig. 4 A and B). The transcripts were abundant within the subventral gland cell lobe, but were not detected in gland extensions and ampullae.
Figure 4.

Localization of EGase transcripts and proteins within cyst nematodes. (A and B) Anterior sections of preparasitic J2 of H. glycines hybridized with digoxigenin-labeled RNA probes transcribed from the HG-eng-2 cDNA. (A) No staining is observed with the sense probe. (B) Specific binding of the antisense probe to the posterior region of the subventral gland cells (g). The gland extensions (e) remain unstained. n, nucleolus of dorsal gland; m, metacorpal pump chamber. (C and D) Immunofluorescence of H. glycines using antiserum raised against recombinant nematode HG-ENG2. (C) No specific labeling with the preimmune serum. (D) Intense labeling of subventral glands with HG-ENG-2-specific immune serum. (Bar = 20 μm.)
Antisera raised against the four recombinant ENG proteins bound specifically on Western blots to the purified recombinant proteins that were used as immunogens and the subventral gland proteins in nematode homogenates (data not shown). Immunofluorescence microscopy revealed a clear binding of these antisera to the subventral glands of preparasitic J2 of the two cyst nematode species (shown for H. glycines only, Fig. 4 C and D).
HCA.
For further characterization, HCA plots of the catalytic domains of four nematode EGases were compared with various other cellulases. The nematode EGases showed the highest similarity to cellulases from the glycosyl hydrolase family 5 (26–28). Fig. 5 shows the HCA plots of HG-ENG-1 and GR-ENG-1 together with two representatives of glycosyl hydrolase family 5, namely the EGases of B. subtilis (Swiss-Prot accession no. P07983) and EGase A of C. cellulolyticum (Swiss-Prot accession no. P17901). The latter, whose three-dimensional structure has been determined (29), was used to identify the catalytic machinery and the secondary structure elements constituting the (α/β)8 barrel structure in the corresponding nematode EGases. Thus, the acid/base and the nucleophilic residue can be assigned to Glu-160 and Glu-249 in HG-ENG-1, Glu-149 and Glu-238 in HG-ENG-2, and Glu-153 and Glu-243 in both GR-ENG-1 and GR-ENG-2 (see also Fig. 1A).
Figure 5.

Hydrophobic cluster analysis of the N-terminal catalytic domains of (A) HG-ENG-1, (B) GR-ENG-1, (C) EGase of B. subtilis (Swiss–Prot accession no. P07983), and (D) EGase A of C. cellulolyticum (Swiss–Prot accession no. P17901). The assignment of the secondary structure in C. cellulolyticum EGase A is according to Ducros et al. (29). The correspondence between the β strands in C. cellulolyticum EGase A and the equivalent strands in the other sequences is shown. The two catalytic residues in family 5 glycosyl hydrolases are shown as white letters in solid circles.
DISCUSSION
Cyst Nematodes Produce β-1,4-Endoglucanases Endogenously.
Several lines of evidence demonstrate the endogenous origin of the four β-1,4-endoglucanase genes isolated from G. rostochiensis and H. glycines. First, the leucine-rich hydrophobic core (h-region) of the signal peptides (23) and the polyadenylated 3′ cDNA ends show that the nematode EGases are of eukaryotic origin and do not originate from bacterial symbionts. Second, contributions from eukaryotic symbionts can be precluded because detailed electron microscopy studies have shown that mAb MGR48 antigens, now identified as EGases, are localized within the secretory granules of the nematode’s subventral glands and no eukaryotic symbionts were observed in these glands (14, 15, 30). Third, the nematode origin of the isolated EGase cDNAs was confirmed by specific binding within the subventral gland of antisera raised to all four active recombinant EGases and of an antisense riboprobe synthesized from the HG-eng-2.
What Kind of β-1,4-Endoglucanases Are Produced by Cyst Nematodes?
HCA of the catalytic domain allowed the assignment of the acid/base and nucleophilic residues involved in catalysis and revealed that all four EGases belong to the family 5 glycosyl hydrolases, the most numerous family to date. This family includes EGases, mannases, and β-1,3-exoglucanases from aerobic and anaerobic bacteria and fungi (26–28). The substrate specificities of family 5 members can be relatively broad and, apart from β-1,4-endoglucanase activity, GR-ENG-1 and -2 and HG-ENG-1 and -2 may have other glycosyl hydrolase activities as well. In two of the four EGases, namely GR- and HG-ENG-1, CBDs were identified. Hence, both cyst nematodes could also have the ability to degrade crystalline cellulose.
The EGases from G. rostochiensis and H. glycines show an intriguing homology with those from bacteria. The catalytic domains of the nematode EGases show 37–44% overall amino acid identity with EGases from E. chrysanthemi, C. acetobutylicum, and B. subtilis. In addition, the CBDs show significant sequence similarity to type II CBDs. Type II CBDs have only been encountered in bacteria (ref. 25; Prosite: PCDOC00485). The close relatedness of the four nematode EGases to those of bacteria raises the question whether nematodes have acquired their cellulases by horizontal gene transfer from a prokaryote. However, too few sequence data are currently available for plant parasitic nematodes to make a firm statement about possible differences in molar G+C content of nematode EGase genes (31) as compared with the hosting nematode chromosome.
Between the catalytic domains and the CBDs, linker sequences of 51 and 50 amino acid residues are present in HG-ENG-1 and GR-ENG-1, respectively. In HG-ENG-1 the linker consists almost completely of an imperfect (SGSS)12 repeat. A similar amino acid repeat of unknown function has been identified from Dictyostelium discoideum (GenBank accession no. U66523). In general, these Gly/Ser-rich regions in proteins may allow flexibility between two functional domains (32). At the C-terminal extension of GR-ENG-2, a sequence [PAAAKK(S)PPAK]3 was found with high similarity to parts of H1 histones of tomato involved in the protein–DNA interaction (33). Nematode EGases are functional outside the plant cell and it is hard to give a biological interpretation for this observation.
Putative Biological Functions of β-1,4-Endoglucanases Produced by Nematodes.
The intracellular migration of cyst nematodes within roots was thought to be facilitated by mechanical means only (34). Our present findings indicate that intracellular migration in host roots by G. rostochiensis and H. glycines involves a combination of mechanical force and enzymatic softening of plant cell walls. Recently we showed that the secretory proteins of the subventral glands, now identified as EGases, are expressed solely in the mobile stages of G. rostochiensis, namely in preparasitic and parasitic J2 and in adult males (15). Both cyst nematode species produce an EGase with a CBD, which could allow for the degradation of crystalline cellulose within the plant cell wall. The two nematode EGases lacking a CBD could enable hydrolysis of soluble polysaccharide fragments, which are products of the initial degradation of crystalline cellulose. These latter EGases could also be involved in the hydrolysis of xyloglucan, a polysaccharide interconnecting crystalline cellulose microfibrils in type I primary cell walls (35, 36). However, the recombinant nematode EGases did not show activity in a plate assay with tamarind xyloglucan (data not shown). In our view the enzymatic characteristics of the EGases described allow the nematode to weaken the mechanical strength of the cellulose microfibril network. It is likely that cyst nematodes produce other cell wall degrading enzymes to attack the plant cell wall in concert with cellulases, e.g., pectinases and proteases.
Distribution of Endogenous EGases Among Members of the Phylum Nematoda.
The mAb MGR48 employed to immunopurify the EGases from G. rostochiensis and H. glycines recognizes also proteins from the cyst nematodes Globodera pallida, Globodera tabacum, and Heterodera schachtii (14). In addition, cellulase activities have been detected in homogenates and secretions of a range of plant-parasitic and fungivorous nematodes (37–42). In view of these results, it is assumed that the ability to produce cellulases endogenously is widely distributed within the phylum Nematoda.
Acknowledgments
This work was supported by the Dutch Technology Foundation (STW) coordinated by the Life Science Foundation (J.M.dB. and J.P.W.G.S.), the Dutch Potato Board (G.S.), European Community Grants BIO2-CT92-0239 (A.S., F.J.G., and J.B.) and FAIR1-CT95-0905 (A.S. and J.B.), and North Atlantic Treaty Organization Award CRG931004 (F.J.G. and R.S.H.). In addition, E.L.D. was supported by the North Carolina Agricultural Research Service, U.S. Department of Agriculture–National Research Institute Project 95-37302-1918 and United Soybean Board Project 400-41-52. T.J.B. and J.M.dB. were supported by grants from the Iowa Soybean Promotion Board, the Iowa State University Biotechnology Council, and the Iowa Agriculture and Home Economics Experiment Station (J-17623, Project No. 3381).
ABBREVIATIONS
- EGase
endoglucanase
- CBD
cellulose-binding domain
- CMC
carboxymethylcellulose
- HCA
hydrophobic cluster analysis
- J2
second stage juvenile
Footnotes
References
- 1.Carpita N C, Gibeaut D M. Plant J. 1993;3:1–30. doi: 10.1111/j.1365-313x.1993.tb00007.x. [DOI] [PubMed] [Google Scholar]
- 2.Beguin P, Aubert J. FEMS Microbiol Rev. 1994;13:25–58. doi: 10.1111/j.1574-6976.1994.tb00033.x. [DOI] [PubMed] [Google Scholar]
- 3.Teeri T T. Trends Biotechnol. 1997;15:160–167. [Google Scholar]
- 4.Gilkes N R, Henrissat B, Kilburn D G, Miller R C, Jr, Warren R A. Microbiol Rev. 1991;55:303–315. doi: 10.1128/mr.55.2.303-315.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Klyosov A. Biochemistry. 1990;29:10577–10585. doi: 10.1021/bi00499a001. [DOI] [PubMed] [Google Scholar]
- 6.Attwood G T, Herrera F, Weissenstein L A, White B A. Can J Microbiol. 1996;42:267–278. doi: 10.1139/m96-039. [DOI] [PubMed] [Google Scholar]
- 7.Scrivener A M, Slaytor M, Rose H A. J Insect Physiol. 1989;35:935–941. [Google Scholar]
- 8.Hogan M, Veivers P C, Slaytor M, Czolu R T. J Insect Physiol. 1988;34:891–900. [Google Scholar]
- 9.Das K, Tripathi S. Aquaculture. 1991;92:21–32. [Google Scholar]
- 10.Pant R, Ramana D. Insect Biochem. 1989;19:269–276. [Google Scholar]
- 11.Whiston R A, Seal K J. Biol Wastes. 1988;25:239–242. [Google Scholar]
- 12.Jones M G K. Ann Appl Biol. 1981;97:353–372. [Google Scholar]
- 13.Hussey R S. Annu Rev Phytopathol. 1989;27:123–141. doi: 10.1146/annurev-phyto-073009-114414. [DOI] [PubMed] [Google Scholar]
- 14.De Boer J M, Smant G, Goverse A, Davis E L, Overmars H A, Pomp H, Van Gent-Pelzer M, Zilverentant J F, Stokkermans J W P G, Hussey R S, Gommers F J, Bakker J, Schots A. Mol Plant–Microbe Interact. 1996;9:39–46. doi: 10.1094/mpmi-9-0039. [DOI] [PubMed] [Google Scholar]
- 15.Smant G, Goverse A, Stokkermans J P W G, De Boer J M, Pomp H, Zilverentant J F, Overmars H A, Helder J, Schots A, Bakker J. Phytopathology. 1997;87:839–845. doi: 10.1094/PHYTO.1997.87.8.839. [DOI] [PubMed] [Google Scholar]
- 16.Goverse A, Davis E L, Hussey R S. J Nematol. 1994;26:251–259. [PMC free article] [PubMed] [Google Scholar]
- 17.Harlow E, Lane D. Antibodies: A Laboratory Manual. Plainview, NY: Cold Spring Harbor Lab. Press; 1988. [Google Scholar]
- 18.Schagger H, Von-Jagow G. Anal Biochem. 1987;166:368–379. doi: 10.1016/0003-2697(87)90587-2. [DOI] [PubMed] [Google Scholar]
- 19.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 20.Mateos P F, Jimenez Zurdo J I, Chen J, Squartini A S, Haack S K, Martinez Molina E, Hubbell D H, Dazzo F B. Appl Environ Microbiol. 1992;58:1816–1822. doi: 10.1128/aem.58.6.1816-1822.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bortoli German I, Haiech J, Chippaux M, Barras F. J Mol Biol. 1995;246:82–94. doi: 10.1006/jmbi.1994.0068. [DOI] [PubMed] [Google Scholar]
- 22.Lemesle Varloot L, Henrissat B, Gaboriaud C, Bissery V, Morgat A, Mornon J P. Biochimie. 1990;72:555–574. doi: 10.1016/0300-9084(90)90120-6. [DOI] [PubMed] [Google Scholar]
- 23.Nielsen H, Engelbrecht J, Brunak S, Von Heijne G. Protein Eng. 1997;10:1–6. doi: 10.1093/protein/10.1.1. [DOI] [PubMed] [Google Scholar]
- 24.Pearson W R, Lipman D J. Proc Natl Acad Sci USA. 1988;85:2444–2448. doi: 10.1073/pnas.85.8.2444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tomme P, Warren R A J, Gilkes N R. Adv Microb Physiol. 1995;37:1–81. doi: 10.1016/s0065-2911(08)60143-5. [DOI] [PubMed] [Google Scholar]
- 26.Henrissat B. Biochem J. 1991;280:309–316. doi: 10.1042/bj2800309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Henrissat B, Bairoch A. Biochem J. 1993;293:781–788. doi: 10.1042/bj2930781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Henrissat B, Bairoch A. Biochem J. 1996;316:695–696. doi: 10.1042/bj3160695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ducros V, Czjzek M, Belaich A, Gaudin C, Fierobe H P, Belaich J P, Davies G J, Haser R. Structure (London) 1995;3:939–949. doi: 10.1016/S0969-2126(01)00228-3. [DOI] [PubMed] [Google Scholar]
- 30.Endo B Y. Proc Helminthol Soc Wash. 1984;51:1–24. [Google Scholar]
- 31.Guiseppi A, Aymeric J L, Cami B, Barras F, Creuzet N. Gene. 1991;106:109–114. doi: 10.1016/0378-1119(91)90573-t. [DOI] [PubMed] [Google Scholar]
- 32.Huston J S, Levinson D, Mudgett Hunter M, Tai M S, Novotny J, Margolies M N, Ridge R J, Bruccoleri R E, Haber E, Crea R, et al. Proc Natl Acad Sci USA. 1988;85:5879–5883. doi: 10.1073/pnas.85.16.5879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jayawardene N, Riggs C. Eur J Biochem. 1994;223:693–699. doi: 10.1111/j.1432-1033.1994.tb19042.x. [DOI] [PubMed] [Google Scholar]
- 34.Wyss U, Zunke U. Rev Nematol. 1986;9:153–165. [Google Scholar]
- 35.Hayashi T. Annu Rev Plant Physiol Plant Mol Biol. 1989;40:139–168. [Google Scholar]
- 36.Vincken J P, Beldman G, Niessen W M A, Voragen A G. Carbohydr Polym. 1996;29:75–85. doi: 10.1016/s0008-6215(96)90801-0. [DOI] [PubMed] [Google Scholar]
- 37.Bird A F, Downton W J S, Hawker J S. Marcellia. 1975;38:165–169. [Google Scholar]
- 38.Deubert K H, Rohde R A. In: Plant Parasitic Nematodes: Cytogenetics, Host–Parasite Interactions and Physiology. Zuckerman B M, Mai W F, Rohde R A, editors. Vol. 2. New York: Academic; 1971. pp. 73–90. [Google Scholar]
- 39.Kojima K K, Kamijyo A, Masumori A, Sasaki S. J Jpn For Soc. 1994;76:258–262. [Google Scholar]
- 40.Tracey M V. Nematologica. 1958;3:179–183. [Google Scholar]
- 41.Dropkin V H. Nematologica. 1963;9:444–454. [Google Scholar]
- 42.Myers R F. Nematologica. 1965;11:441–448. [Google Scholar]