

Abstract
Cytogenetic analysis of maize landraces from northwestern Argentina has revealed an altitudinal cline in the mean number of B chromosomes (B's) per plant, with cultivars growing at higher altitudes exhibiting a higher number of B's. Altitudinal and longitudinal clines are frequently interpreted as evidence of selection, however, they can also be produced by the interplay between drift and spatially restricted gene flow or by admixture between previously isolated populations that have come into secondary contact. Here, we test the adaptive significance of the observed altitudinal gradient by comparing the levels of differentiation in the mean number of B's to those obtained from 18 selectively neutral loci [simple sequence repeats (SSRs)] among seven populations of the cline. The adequacy of alternative genetic-differentiation measures was determined, and associations between cytogenetic, genetic, and altitudinal distances were assessed by means of matrix- correspondence tests. No evidence for association between pairwise FST and altitudinal distance or B-chromosome differentiation was found. The contrasting pattern of altitudinal divergence between the mean number of B's per plant and the genetic differentiation at SSR loci indicates that demographic processes cannot account for the observed levels of divergence in the mean number of B's.
B chromosomes (B's) are nonessential, extra chromosomes that have been found in all major groups of plants and animals (Jones and Rees 1982). They are recognized by three criteria: (i) they are dispensable and can be present or absent from individuals within a population, (ii) they do not pair or recombine with any members of the standard diploid (or polyploid) set of A chromosomes at meiosis, and (iii) their inheritance is non-Mendelian and irregular (Beukeboom 1994).
B chromosomes' origin and maintenance in natural populations have been a matter of debate over the past 50 years (Blackwood 1956; Jones and Rees 1982; Carlson 1986; Porter and Rayburn 1990; Carlson and Roseman 1992; Jones 1995). According to the most recent views, these are selfish genetic elements that are derived from the A complement and that maintain their polymorphism by a series of accumulation mechanisms (e.g., mitotic nondisjunction, reduction of meiotic loss) or, much more rarely, by providing their carriers a selective advantage. They can attain extremely high frequencies in natural populations, depending both on the degree to which a particular species can tolerate these additional elements and on the strength of the accumulation mechanisms (Camacho et al. 2000). These interactions result in a dynamic system, which has been interpreted as a host–parasite relationship between the A and B chromosomes (Jones and Houben 2003).
B chromosomes are of particular interest in maize (Zea mays ssp. mays L.). Numerical polymorphisms have been described in a large number of maize landraces (Mcclintock et al. 1981; Chiavarino et al. 1995; Naranjo et al. 1995; Rosato et al. 1998), with differences in the number of B's being one of the main factors contributing to intraspecific genome-size variation. Although several accumulation mechanisms have been reported for maize B chromosomes (Roman 1947,1948; Carlson and Chou 1981; Carlson and Roseman 1992), the significance of the distinctive patterns of occurrence of these supernumerary elements in different native populations still remains unclear. Attempts to correlate B chromosome frequencies with cytogenetical and environmental variables have yielded somewhat contradictory results. No association was found between the number of B's and DNA content or the number of heterochromatic bands in 12 Arizona populations (Porter and Rayburn 1990), while negative correlations were reported between B's and the amount of heterochromatin in both North American and Italian populations (Longley 1938; Bianchi et al. 1963). Bretting and Goodman (1989) noted a generally negative correlation between B chromosomes and altitude in 300 Mesoamerican populations, but this association was not encountered by Porter and Rayburn (1990).
The cytogenetic studies conducted by Rosato et al. (1998) and Poggio et al. (1998) in 21 native populations from northwestern Argentina revealed a negative correlation between the mean number of B's per plant and the number of heterochromatic bands and a positive correlation between the mean number of B's per plant and altitude. According to these authors, B's would be tolerated at high frequencies in those populations with lower numbers of heterochromatic bands, so as to maintain a relatively constant amount of nuclear DNA or an “optimal” nucleotype (sensu Bennett 1987). Although selective pressures may indeed be acting on the conservation of total genomic DNA, this hypothesis does not account for the positive correlation between B's and altitude. Altitudinal and longitudinal clines of morphological traits, or allelic frequencies, have long been interpreted as evidence of selection. However, clines can also be produced by the interplay between drift and spatially restricted gene flow (isolation by distance) or by admixture between previously isolated populations that have come into secondary contact (Confalonieri et al. 1998; Storz and Dubach 2004). One means of distinguishing between adaptive vs. nonadaptive causes of clinal variation is to compare the relative levels of between-population divergence in the trait under study and in neutral DNA markers (Storz 2002). The rationale behind this approach is that the effects of selection are generally locus specific, whereas demographic processes are expected to have relatively uniform effects across the entire genome (Cavalli-Sforza 1966; Lewontin and Krakauer 1973).
Rosato et al. (1998) suggested that the altitudinal cline of B chromosomes found in maize landraces from northwestern Argentina could be the product of selective forces. However, no experimental data were provided to support this assertion. The aim of the present study is to test the adaptive significance of the altitudinal cline of B chromosomes by using neutral molecular markers [single sequence repeats (SSRs)] to assess the levels of genetic divergence of the populations studied by Rosato et al. (1998). If the B chromosome gradient is due to demographical processes related to the history of the populations, the levels of divergence estimated by the neutral markers should also exhibit a clinal pattern of variation. In contrast, if the B polymorphism is in fact subject to selection across the altitudinal gradient, the levels of divergence estimated by the neutral markers should show no association with altitude or B chromosome differentiation among populations.
MATERIALS AND METHODS
The seven populations included in this study are a subset of those previously analyzed by Rosato et al. (1998) and were selected on the basis of seed availability from the original collections performed by these authors during 1994–1995. Populations are distributed spanning an altitudinal distance of ∼2090 m and over a linear distance of 649 km. Voucher specimens, collection sites, racial identification, cultivar altitudes, mean number of B's per plant, and sample sizes are given in Table 1. The geographical distribution of populations is presented in Figure 1.
TABLE 1.
Maize landraces included in the present study
| Population identification | Vouchera | Collection site | Landraceb | Cultivar altitude (m.a.s.l.) | Mean no. of B's per plantc | Sample size |
|---|---|---|---|---|---|---|
| 6167 | VAV 6167 | El Puesto, Dpto. Santa Victoria, Salta, Argentina | Altiplano | 3000 | 1.294 | 14 |
| 6485 | VAV 6485 | Colonia San José, Dpto. Tilcara, Jujuy, Argentina | Blanco | 2670 | 1.795 | 13 |
| 6480 | VAV 6480 | La Ciénaga de Pumamarca, Dpto. Tumbaya, Jujuy, Argentina | Amarillo Grande | 2420 | 1.596 | 25 |
| 6484 | VAV 6484 | Tumbaya, Dpto. Tumbaya, Jujuy, Argentina | Amarillo Chico | 2010 | 0.204 | 20 |
| 6476 | VAV 6476 | Termas de Reyes, Dpto. Capital, Jujuy, Argentina | Amarillo Chico | 1690 | 0.255 | 14 |
| 6313 | VAV 6313 | Los Toldillos, Dpto. Ambato, Catamarca, Argentina | Pisingallo | 1600 | 0.685 | 16 |
| 6482 | VAV 6482 | La Candelaria, Dpto. Candelaria, Salta, Argentina | Orgullo Cuarentón | 910 | 0.122 | 24 |
m.a.s.l, meters above sea level; Dpto., Departamento.
Voucher specimens are deposited at the “Laboratorio de Recursos Genéticos Vegetales N.I. Vavilov” (VAV), Faculty of Agronomy, University of Buenos Aires.
Taxonomic identification based on morphological criteria.
Data from Rosato et al. (1998).
Figure 1.—

Geographic localities of the seven populations included in this study.
Selection of SSR loci:
Since simple-sequence-repeat variation is generally considered to be selectively neutral (Schlötterer and Wiehe 1999), a set of SSR loci distributed across the 10 maize chromosomes was used to assess population differentiation at neutral loci. Eighteen SSR were selected from a preliminary survey of 39, with only those loci with unambiguous interpretation being used for analysis. Each chromosome pair was represented by two unlinked loci, except for pairs 8 and 9, in which a single locus was studied. Details of the used loci, including chromosomal location (bin number), putative repeat motifs, and associated primers, are shown in Table 2. Additional information on these loci can be found at the Maize Genetics and Genomics Database (http://www.maizegdb.org/ssr.php).
TABLE 2.
Details of the 18 SSR loci used in this study
| Chromosome location (bin) | Repeat motif | Primers
|
||
|---|---|---|---|---|
| Locus | Forward 5′–3′ | Reverse 5′–3′ | ||
| bnlg1866 | 1.03 | AG | CCCAGCGCATGTCAACTCT | CCCCGGTAATTCAGTGGATA |
| phi037 | 1.08 | CT | CCCAGCTCCTGTTGTCGGCTCAGAC | TCCAGATCCGCCGCACCTCACGTCA |
| bnlg1018 | 2.04 | AG | CGAGGTTAGCACCGACAAAT | CGAGTAAATGCTCTGTGCCA |
| phi127 | 2.08 | GTCT | ATATGCATTGCCTGGAACTGGAAGGA | AATTCAAACACGCCTCCCGAGTGT |
| phi029 | 3.04 | CCCT-CT | TTGTCTTTCTTCCTCCACAAGCAGCGAA | ATTTCCAGTTGCCACCGACGAAGAACTT |
| bnlg1182 | 3.09 | AG | AGCCGAGTCAGTTCGAGGTA | CAGGGGCTTGAGGTGAGTTA |
| nc135 | 4.01 | AG | CACAAAGAGCAGCCCACTTT | AAGTTGCTGACATCGATCCA |
| bnlg252 | 4.06 | — | CGTTCTCCGTACAGCACAGACCAACGT | CTCAGATGAACTCCTCAGCAGCTGTAGCCT |
| bnlg1700 | 5.03 | AG | GTCACATCCATGTAGTGCACG | GGCACCCTTTTGAAACCTTT |
| bnlg1287 | 5.04 | AG | GCCCTACCTGTTCTGTCTCG | TGTCCCATACCTCAACGTGA |
| bnlg1165 | 6.01 | AG | CGCTTGCATCATCTCAAGAA | TTCAAGTTTAGCCACCCACC |
| bnlg1732 | 6.05 | AG | AACTTTTGGCATTGCACTGG | CGTAAGTGCACACGGCATTA |
| bnlg1070 | 7.03 | AG | TTCCAGTAAGGGAGGTGCTG | TAAGCAACATATAGCCGGGC |
| phi069 | 7.05 | GAC | AGACACCGCCGTGGTCGTC | AGTCCGGCTCCACCTCCTTC |
| phi121 | 8.03 | CCG | AGGAAAATGGAGCCGGTGAACCA | TTGGTCTGGACCAAGCACATACAC |
| bnlg1209 | 9.04 | AG | GTCCCGGGCAGAATAATACC | TTCCTCCTTGAAGTGCTCGT |
| phi059 | 10.02 | ACC | AAGCTAATTAAGGCCGGTCATCCC | TCCGTGTACTCGGCGGACTC |
| bnlg1360 | 10.07 | AG | TCTGCTCATCCACAACTTGC | AGAACGTGAAGCTGAGCGTT |
SSR typing:
DNA was extracted from 2- to 3-day-old seedlings according to Dellaporta et al. (1983). PCR reaction mixtures contained 1 μl of DNA extract (∼50 ng), 30 ng each primer, 125 μm each dNTP, 1.5 mm MgCl2, 0.5 unit Taq DNA polymerase (Promega, Madison, WI), 1× PCR buffer and sterile double-distilled water to a final volume of 25 μl. A touchdown cycling profile (annealing temperature 65°–55°) was used and PCR products were separated on a 6% denaturing polyacrylamide gel (8 m urea) following standard procedures. Gels were silver stained with Silver Sequence DNA Staining Reagents (Promega). Alleles were identified by comparison with products of known size using GelPro Analyzer 4.0 (Media Cybernetics, Bethesda, MD).
Data analysis:
Allele frequencies were calculated with the direct-count method using Fstat version 2.9.3.2 (Goudet 2001). Genotypic linkage disequilibrium was analyzed using Genepop version 3.4. (Raymond and Rousset 1995). Significance levels were modified according to Bonferroni procedures at an overall α = 0.05 (Rice 1989).
Genetic differentiation measures:
The analysis of SSR data usually entails the use of either of two classes of population differentiation measures: allele identity-based statistics derived from the infinite allele model (IAM) (Kimura and Crow 1964) or allele size-based statistics derived from the stepwise mutation model (SMM) (Kimura and Ohta 1978). SSR markers are frequently assumed to evolve under the SMM, however, the relative efficiency of both types of measures is not only dependent on the mutational process underlying SSR polymorphism, but also on the relative contributions of mutation, drift, and migration. Of the 18 loci included in this study, locus bnlg1018, bnlg1209, bnlg1287, bnlg1070, bnlg1182, bnlg1866, bnlg1732, bnlg1360, and bnlg252 were demonstrated to evolve in a stepwise fashion by Vigouroux et al. (2002); variation at loci bnlg1700 and bnlg1165 was shown to be consistent with the SMM and variation at loci phi121, phi059, nc135, phi037, phi069, phi127, and phi029 was found not to be consistent with this model (Lia et al. 2007).
Since the accurate estimation of population differentiation is fundamental in the present study, the allele-size randomization test of Hardy et al. (2003) was used to determine whether allele identity-based (e.g., FST, Wright 1978) or allele size-based statistics (e.g., RST, Slatkin 1995) were most adequate to analyze the stepwise evolving loci. The principle of the test is based on obtaining a distribution of a statistic under the null hypothesis that differences in allele sizes do not contribute to population differentiation. Throughout the randomization procedure, genotypes are defined in terms of allelic states and are not modified, but allele sizes are randomly reassigned among allelic states. The test can thus be designed by comparing the observed RST value to the distribution of permuted RST (pRST) values obtained for all possible configurations of allele-size permutations (or a representative subset of them). A significant test implies that RST performs better than FST, a nonsignificant test implies that FST performs better than RST. The test was conducted as implemented in the spatial pattern analysis of genetic diversity (SPAGeDi) software (Hardy and Vekemans 2002).
Global and pairwise θ, an unbiased estimate of FST (Weir and Cockerham 1984), were calculated over all loci with the Fstat software package (Goudet 2001). The significance of θ values, hereafter referred to as FST, was tested by permuting genotypes rather than alleles among samples, as this is the preferred method when the Hardy–Weinberg equilibrium is rejected within samples, and was corrected for multiple comparisons using Bonferroni procedures at an overall α = 0.05.
Correlation analyses:
All tests involving B chromosome comparisons were performed using the variable mean number of B's per plant ( ) as originally defined by Rosato et al. (1998), which corresponds to the number of B's per individual averaged across the total number of individuals evaluated in each population.
) as originally defined by Rosato et al. (1998), which corresponds to the number of B's per individual averaged across the total number of individuals evaluated in each population.
To analyze the relationship between allele frequencies and altitude, and allele frequencies and mean number of B's per plant, allele frequencies were calculated for each locus and population and used to compute Spearman's coefficient of rank correlation (rs) with the STATISTICA software package (StatSoft, Tulsa, OK). Significance levels were corrected for multiple comparisons using Bonferroni procedures at an overall α = 0.05.
The association between genetic differentiation at SSR loci and B chromosome differentiation, altitudinal distance, and geographic distance was determined using simple Mantel tests (Mantel 1967) as implemented in ISOLDE routine of Genepop version 3.4 (Raymond and Rousset 1995), with FST as a measure of genetic differentiation. Following the approach of Perfectti et al. (2004), B chromosome differentiation among populations was computed as  where
where 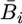 and
and  represent the mean number of B chromosomes per plant for the ith and jth populations. Altitudinal distances were computed as
represent the mean number of B chromosomes per plant for the ith and jth populations. Altitudinal distances were computed as  where xi and xj represent altitudes for the ith and jth populations.
where xi and xj represent altitudes for the ith and jth populations.
RESULTS
A total of 126 individuals were genotyped at the 18 SSR loci. Seventeen loci were polymorphic, with the only monomorphic locus being phi121. The total number of alleles detected was 184 and the mean number of alleles per locus and population was 5.032 (range: 4.333–6.055). No evidence of linkage disequilibrium was found among loci. Allele frequencies per locus and population are provided as supplemental Table S1 at http://www.genetics.org/supplemental/.
Estimates of genetic differentiation (FST vs. RST)
The adequacy of FST and RST statistics as measures of genetic differentiation was evaluated using the allele size permutation test of Hardy et al. (2003). Since deviations from the SMM at some loci can reduce the power of the test, only those loci known to conform to model assumptions were included in the analysis. To determine whether differences in the mutational behavior of the loci under study could influence the results of the test, two global estimates were computed: (a) a 9-loci estimate using those SSR found to evolve in a strictly stepwise fashion by mutation-accumulation experiments (Vigouroux et al. 2002) and (b) an 11-loci estimate including bnlg1165 and bnlg1700, whose allele-nucleotide substitution patterns are compatible with the SMM (Lia et al. 2007), although a fit to the model has not been tested experimentally. Allele permutations did not reveal any significant contribution of stepwise mutations to population differentiation with either of the two global estimates (Table 3).
TABLE 3.
Contribution of stepwise-like mutations to genetic differentiation as determined by the allele size permutation test of Hardy et al. (2003)
| RST | pRST (95% C.I.)a | P | |
|---|---|---|---|
| Bnlg1018 | 0.3266 | 0.2323 (0.0350–0.4661) | 0.1968 |
| Bnlg1209 | 0.0467 | 0.1326 (0.0169–0.3021) | 0.8901 |
| Bnlg1287 | 0.0578 | 0.1097 (0.0156–0.2250) | 0.7542 |
| Bnlg1070 | 0.0063 | 0.1186 (0.0013–0.2935) | 0.9670 |
| Bnlg1182 | 0.3256 | 0.2781 (0.0027–0.4790) | 0.4326 |
| Bnlg1866 | 0.3198 | 0.0911 (0–0.2811) | 0.0130* |
| Bnlg1732 | 0.0050 | 0.0567 (0–0.1794) | 0.9131 |
| Bnlg1360 | 0.2751 | 0.1017 (0.0022–0.2379) | 0.0080** |
| Bnlg252 | 0.2454 | 0.1826 (0.0339–0.3805) | 0.2298 |
| Bnlg1700 | 0.2937 | 0.1711 (0.0321–0.3373) | 0.0919 |
| Bnlg1165 | 0.1861 | 0.0730 (0.0016–0.1595) | 0.0060** |
| Global (9 loci) | 0.2309 | 0.1633 (0.0874–0.2455) | 0.0629 |
| Global (11 loci) | 0.2310 | 0.1587 (0.08345–0.2490 | 0.0559 |
Values are given for each locus and the multilocus estimates. *P < 0.05, **P < 0.01.
Central pRST and 95% confidence interval values obtained after random permutations of allele sizes.
Single-locus comparisons were conducted for the 11 loci, with the number of permutations ranging from 120 to 1000 depending on the number of alleles at each locus. The majority of loci (8/11) showed RST values that were not significantly different from pRST. However, RST was significantly larger than pRST at loci bnlg1866, bnlg1360, and bnlg1165 (Table 3), suggesting that mutation rates are not homogeneous among the 11 SSR loci studied.
Overall, the results of the permutation tests indicate that differences in allele sizes do not contribute to population differentiation and that divergence is caused mainly by drift, with gene flow, and/or the reciprocal of the divergence time, being large compared to the mutation rate. Under this scenario, FST should be preferred over RST (Gaggiotti et al. 1999; Hardy et al. 2003). In addition, given that 7 of the 18 SSRs included in this study do not conform to the SMM, the use of FST allows for the 18 loci to be pooled into a single measure of genetic differentiation, even though they have different evolutionary dynamics.
Population structure:
The analysis of SSR variation revealed a significant degree of genetic subdivision among populations (FST = 0.115, 95% C.I. = 0.094–0.142). FST values for pairwise comparisons and their significance levels are given in Table 4. A strong pattern of isolation by distance was evident, as revealed by a positive association between pairwise FST and pairwise geographic distances (rs = 0.861, P = 0.009, 5000 permutations) (Figure 2).
TABLE 4.
Genetic differentiation, altitudinal distance, geographic distance, and B chromosome differentiation for each pairwise combination of populations
| Population pair | FST | Altitudinal distance (m) | Geographic distance (km) | ΔB |
|---|---|---|---|---|
| 6167-6485 | 0.0381** | 330 | 151 | 0.501 |
| 6167-6480 | 0.0467** | 580 | 172 | 0.302 |
| 6167-6484 | 0.0380** | 990 | 184 | 1.090 |
| 6167-6476 | 0.0778** | 1310 | 220 | 1.039 |
| 6167-6313 | 0.2279** | 1400 | 649 | 0.609 |
| 6167-6482 | 0.1332** | 2090 | 429 | 1.172 |
| 6485-6480 | 0.0263** | 250 | 22.3 | 0.199 |
| 6485-6484 | 0.0408** | 660 | 32.4 | 1.591 |
| 6485-6476 | 0.0718** | 980 | 67.4 | 1.540 |
| 6485-6313 | 0.1956** | 1070 | 509 | 1.110 |
| 6485-6482 | 0.1164** | 1760 | 282 | 1.673 |
| 6480-6484 | 0.0480** | 410 | 13.2 | 1.392 |
| 6480-6476 | 0.0627** | 730 | 48.4 | 1.341 |
| 6480-6313 | 0.1951** | 820 | 491 | 0.911 |
| 6480-6482 | 0.1157** | 1510 | 265 | 1.474 |
| 6484-6476 | 0.0955** | 320 | 35.1 | 0.051 |
| 6484-6313 | 0.2407** | 410 | 477 | 0.481 |
| 6484-6482 | 0.1533** | 1100 | 253 | 0.082 |
| 6476-6313 | 0.1599** | 900 | 440 | 0.430 |
| 6476-6482 | 0.0748** | 780 | 218 | 0.133 |
| 6313-6482 | 0.1673** | 690 | 240 | 0.563 |
P < 0.01.
Figure 2.—

Differentiation among populations. Multilocus estimates of pairwise differentiation (FST/1 − FST) are plotted against geographic distances. The regression is y = 0.0332x + 0.0003.
Genetic differentiation and clinal variation:
To investigate whether particular alleles were correlated with altitude or with the B chromosome cline, the frequencies of 183 alleles were compared against altitude and the mean number of B's per plant by means of Spearman's coefficient of rank correlation. None of the alleles exhibited a systematic pattern of variation across the altitudinal cline and no association was found between allele frequencies and the mean number of B's per plant (P > 0.05 for the 183 comparisons).
The positive correlation between the mean number of B's per plant and altitude was originally reported by Rosato et al. (1998) for a sample of 16 populations using simple linear correlations. To verify this correlation for the subset of populations included in this study and to test the sensitivity of the Mantel test to detect such association, the Mantel matrix permutation test was performed for the 7 populations studied here, considering pairwise differences in cultivar altitude and in the mean number of B's per plant (ΔB). Again, a positive correlation was found between both variables (rs = 0.484, P = 0.037, 5000 permutations) (Figure 3A), supporting the use of the Mantel test for further comparisons.
Figure 3.—

Genetic differentiation and clinal variation. Plots of (A) pairwise ΔB against altitudinal distances between populations, (B) pairwise ΔB against geographic distances between populations, (C) pairwise FST against altitudinal distances between populations, and (D) pairwise ΔB against pairwise FST.
Despite being distributed across the altitudinal cline, the seven populations analyzed here are not distributed on a linear transect; thus, differences in altitude may not always reflect differences in linear geographic distances. However, there seems to be an overall accordance between these measures, since a positive correlation was found between both distances (rs = 0.493, P = 0.031, 5000 permutations). Given the positive correlations between ΔB and pairwise altitudinal distances, and pairwise geographic and altitudinal distances, the correlation between ΔB and geographic distance was also tested, showing no significant association (rs = −0.019, P = 0.445, 5000 permutations) (Figure 3B).
As mentioned earlier, if the altitudinal cline of B chromosomes is due to demographic processes, then a positive correlation is also expected between differences in cultivar altitude and the levels of differentiation assessed with the neutral markers. No evidence for association between pairwise FST and altitudinal distance was found (rs = 0.325, P = 0.147, 5000 permutations) (Figure 3C).
An additional prediction of the demographic hypothesis is that differences in the mean number of B's per plant should also be positively correlated with population differentiation at neutral markers. No significant association was found between ΔB and pairwise FST (rs = −0.091, P = 0.653, 5000 permutations) (Figure 3D), indicating that between-population differences in the mean number of B's per plant are independent of their neutral genetic background. A compelling example is given by the pairwise comparison involving populations 6476 and 6480, which in spite of showing a rather low FST value (FST = 0.064) it also exhibits one of the highest differences in the mean number of B's per plant (ΔB = 1.341).
DISCUSSION
The results presented here support the adaptive significance of the altitudinal cline of B chromosomes originally described by Rosato et al. (1998). The contrasting pattern of altitudinal divergence between the mean number of B's per plant and the genetic differentiation at SSR loci indicates that demographic processes cannot account for the observed levels of divergence in the mean number of B's (Figure 3).
No evidence of clinal variation was found at the 18 SSR loci included in this study, either on a global or on an individual basis, as demonstrated by the lack of correlation between allele frequencies and altitude. Moreover, if the altitudinal cline of B chromosomes had been the product of secondary contact, the effects of admixture would have been readily detectable. Admixture is expected to produce a bimodal distribution of pairwise FST values and linkage disequilibrium among loci that have different allele frequencies in the separate source populations (Storz and Dubach 2004). However, none of these phenomena was apparent.
The positive association between geographic and genetic distances (Figure 2) suggests that the seven populations distributed along the cline are at migration-drift equilibrium (Hutchinson and Templeton 1999). This implies that there are no significant barriers to dispersal and that the clinal distribution of B chromosomes persists, even in the presence of gene flow.
Tests of natural selection along environmental clines often rely on the comparison of the population structures of the characters on which selection may act and on that of neutral molecular markers (Gockel et al. 2001). Here, comparison of the levels of neutral genetic differentiation with both the mean number of B's per plant and pairwise altitudinal distances explicitly incorporates the relevant environmental variable into the analysis. Previous authors have used partial matrix correspondence tests to determine whether the association between morphological and environmental variables remained significant after controlling for neutral genetic divergence within the context of isolation by distance (Gubitz et al. 2000; Storz 2002). Partial correlation analyses were not required in the present study, given that neither ΔB nor pairwise altitudinal distances showed a significant association with pairwise FST.
It could be argued that between-population differences in the mean number of B's per plant do not provide an accurate estimate of B chromosome differentiation, since it may not be clear whether this measure represents genetic or phenotypic variation. To be meaningful, pairwise differences in the mean number of B's should be intimately related to the genetic variation underlying this character. The genetic basis of the mechanisms involved in the transmission of maize B's seems to support this assumption. Three main accumulation mechanisms determine the number of B's: (i) directed nondisjunction in the second pollen grain mitosis (Roman 1947; Carlson and Chou 1981), (ii) preferential fertilization by male gametes carrying B's (Roman 1948), and (iii) reduction of meiotic loss (Carlson and Roseman 1992). Recent studies on the dynamics of B transmission have demonstrated that a single autosomal gene controls preferential fertilization when B's are transmitted on the male side (Chiavarino et al. 1998, 2001), whereas on the female side a low transmission allele from a different autosomal locus seems to be implicated in promoting meiotic loss (González-Sánchez et al. 2003).
How could selection act on the maintenance of the cline?
Some effects of the B chromosomes appear to be attributable to the products of their genes, as is the case with genes controlling resistance to rust in the B chromosomes of Avena sativa (Dherawattana and Sadanaga 1973) and with the genes conferring resistance to antibiotics in the B chromosomes of the fungus Nectria haematococca (Miao et al. 1991). Maize B chromosomes are highly repetitive in nature and share the vast majority of their sequences with the regular complement (Peacock et al. 1981; Cheng and Lin 2003, 2004). Transcription of B-specific sequences was recently reported for the first time by Lamb et al. (2007). Portions of a retroposon-derived element were found to be differentially expressed in B-carrying individuals and the absence of polyadenylation suggested they are not translated into proteins. How the expression of these elements relates to the B life cycle is still unclear; however, their involvement in the altitudinal gradient described here cannot be discarded.
If not a consequence of their gene products, the selective advantages, or disadvantages, of B's may stem from their physical presence in the nucleus. There is ample evidence that B chromosomes can affect a multitude of cellular and physiological processes in both plants and animals (for a review, see Camacho et al. 2000; Jenkins and Jones 2004). Among the most remarkable, B chromosomes have been shown to affect mean cell chiasma frequency and the distribution of chiasmata within the A complement (Jones 1995; Colombo and Confalonieri 2004; Riera et al. 2004). Similar effects have been reported for maize (Hanson 1969), where the late replicating heterochromatin of the B's has been proposed to extend the first meiotic prophase therefore lengthening the time during which recombination takes place (Rhoades and Dempsey 1972). In addition to an increase in recombination, several cytological abnormalities have been demonstrated to derive from the presence of B's in maize plants. Micronucleus formation and A chromosome instability were found to be significantly increased in the tapetal cells of B-carrying individuals (Chiavarino et al. 2000), and reduced pollen viability was encountered in female high B-transmission lines, particularly in those plants having B's (González-Sánchez et al. 2004).
The selective basis of the cline does not necessarily imply that selection is acting directly on maize B chromosomes, because the positive association between B's and altitude could also be a by-product of selection at other traits. Earlier studies have shown that the occurrence of two or more B's induces loss of chromosomal segments from A chromosomes containing large blocks of heterochromatin (knobs) (Rhoades and Dempsey 1973). Given that maize knobs replicate very late relative to euchromatin (Pryor et al. 1980; Buckler et al. 1999) and that this may impact the rate of cell division and plant development, it has been suggested that large amounts of heterochromatin are not favored under the harsher climates and shorter growing seasons of high-altitude elevations (Reeves et al. 1998; Buckler et al. 1999). In agreement with this proposal, a negative correlation between altitude and knobs was found in the populations analyzed by Rosato et al. (1998), as well as a negative correlation between knobs and the mean number of B's per plant. In this context, the increase in the number of B's as a function of altitudinal distances may be interpreted as a consequence of their effects on the amount of heterochromatin in the autosomes. Although maize B's are also heterochromatic, and they have been shown to lengthen replication time, “preference” for some type of heterochromatin over the other may still exist, as it has been reported that the different classes of heterochromatin have their own distinctive impact on the duration of the cell cycle (Pryor et al. 1980).
An alternative explanation to the adaptive basis of the cline may reside in the genes controlling B transmission. As previously mentioned, two autosomal loci seem to govern these mechanisms (Chiavarino et al. 2001; González-Sánchez et al. 2003). If linkage disequilibrium exists between those alleles favoring B transmission and the alleles of some other locus subjected to selection in response to altitude, then the observed cline could be produced as a result of hitchhiking.
Implications for B chromosome evolution in maize:
Regardless of the identity and the nature of the selective forces acting on the maintenance of the cline, the implications of our findings in terms of B chromosome evolutionary dynamics still need to be considered.
The two most widely accepted models of B-chromosome evolution are the heterotic model (White 1973) and the parasitic or selfish model (Jones 1985). The heterotic model assumes an equilibrium balance between the positive fitness effects of B chromosomes (which show no accumulation mechanisms) when they occur in low numbers and their negative effects when they occur in high numbers. For the parasitic–selfish model, the equilibrium is the result of B chromosome accumulation and of its detrimental effects on the fitness of B-chromosome carriers (Camacho et al. 2000). A nonequilibrium model of long-term evolution has also been proposed by Camacho et al. (1997) on the basis of the behavior of B chromosomes in the grasshopper Eyprepocnemis plorans. According to this model, B's are neutralized by the A genome through the evolution of genes that suppress drive. Such neutralized B chromosomes are destined to disappear after a long period of stochastic fluctuation, in the course of which the B's can mutate into a new parasitic variant with high drive that restarts the cycle again.
Overall, the three models can be envisaged as particular instances of genomic conflict between A and B chromosomes, each model involving different strategies of attack and defense within a host–parasite relationship (Frank 2000). However, which one of them best explains the B-chromosome polymorphism described here is not easy to determine. The evidence provided in this study indicates that selective forces are acting on the maintenance of the altitudinal cline in the mean number of B's. However, clinal variation may arise either as a consequence of a beneficial effect of B's at higher elevations or as a result of B's being negatively selected at lower elevations, but increasingly tolerated with altitude. Strict compliance to the heterotic model would imply assuming the first case scenario and the absence of accumulation mechanisms, a criterion which is clearly not met by maize B's. Conversely, the second-case scenario is easily reconciled with a parasitic model in which the antagonistic forces leading to the equilibrium state in each population (i.e., accumulation and fitness reduction) change in magnitude as altitude increases. Although our observations can indeed be interpreted under the assumptions of the parasitic model, this model does not fully incorporate the complexities of the interaction between the A and B chromosomes studied here. The nonequilibrium model of long-term evolution, in which B influences may shift from parasitic to neutral and even beneficial, provides a more suitable framework for the interpretation of our results. High variation for B transmission rate (KB) has been reported among individuals from population 6313 on both the male (KB = 0.98–0.17) and the female (KB = 0.58–0.31) side, and the genetic component of this variation has been demonstrated (Rosato et al. 1996; Chiavarino et al. 2001; González-Sánchez et al. 2003). Although the transmission rate of B's has not been determined for the remaining populations of this study, these results indicate that a polymorphic system already exists in the A complement to counteract B accumulation mechanisms and thus promote B neutralization. In its final stage, the model suggests the possibility that neutralization can be overcome by the regeneration of new B variants that recuperate drive. We have no evidence for the generation of new B variants in maize landraces, and no differences in the size or morphology of B's were detected among the populations of the cline.
A coevolution model of B attack and A defense has already been proposed in maize to explain the distinctive B-transmission dynamics in males and females (González-Sánchez et al. 2003). From these results it was concluded that two autosomal loci are primarily responsible for B transmission, with a male dominant high-transmission allele that tends to increase the number of B's over Mendelian expectations and a female dominant low-transmission allele that tends to decrease it. In this connection, the estimation of the allelic frequencies of the genes controlling B transmission in the populations of the cline would provide further insights to determine whether B chromosomes are being positively selected (e.g., a higher frequency of high-transmission alleles at higher elevations) or simply tolerated at higher altitudes.
In conclusion, the altitudinal cline described here and the selective forces acting on its maintenance constitute a new level of complexity in the host–parasite relationship between the A and B chromosomes of maize. Future work is still needed to clarify whether B chromosomes are a direct target of selection or if they are genetically correlated with another trait under direct selection.
Acknowledgments
We thank the late C. Naranjo, J. Cámara Hernández, and A. M. Miante Alzogaray for providing us the plant material and for carrying out the taxonomic classification of the landraces included in this study. We are also indebted to the anonymous reviewers who have greatly improved this manuscript. Several grants from the Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 02296), the University of Buenos Aires (EX-317), and the Agencia Nacional de Promoción Científica y Tecnológica (BID 1201 OC-AR PICT 04443) are gratefully acknowledged.
References
- Bennett, M. D., 1987. Variation in genomic form in plants and its ecological implications. New Phytol. 106: 177–200. [Google Scholar]
- Beukeboom, L. W., 1994. Bewildering Bs: an impression of the 1st B-chromosome conference. Heredity 73: 328–336. [Google Scholar]
- Bianchi, A., M. V. Ghatnekar and A. Ghidoni, 1963. Knobs in italian maize. Chromosoma 14: 601–617. [Google Scholar]
- Blackwood, M., 1956. The inheritance of B chromosomes in Zea mays. Heredity 10: 353–366. [Google Scholar]
- Bretting, P. K., and M. M. Goodman, 1989. Karyotipic variation in Mesoamerican races of maize and its systematic signficance. Econ. Bot. 43: 107–124. [Google Scholar]
- Buckler, IV, E. S., T. L. Phelps-Durr, C. S. K. Buckler, R. K. Dawe, J. F. Doebley et al., 1999. Meiotic drive of chromosomal knobs reshaped the maize genome. Genetics 153: 415–426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camacho, J. P. M., M. W. Shaw, M. D. López–León, M. C. Pardo and J. Cabrero, 1997. Population dynamics of a selfish B chromosome neutralized by the standard genome in the grasshopper Eyprepocnemis plorans. Am. Nat. 149: 1030–1050. [DOI] [PubMed] [Google Scholar]
- Camacho, J. P. M., T. F. Sharbel and L. W. Beukeboom, 2000. B-chromosome evolution. Philos. Trans. R. Soc. Lond. 355: 163–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlson, W. R., 1986. The B chromosome of maize. Crit. Rev. Plant Sci. 3: 201–226. [Google Scholar]
- Carlson, W. R., and T. S. Chou, 1981. B chromosome nondisjunction in corn: control by factors near the centromere. Genetics 97: 379–389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlson, W. R., and R. Roseman, 1992. A new property of the maize B chromosome. Genetics 131: 211–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavalli-Sforza, L. L., 1966. Population structure and human evolution. Proc. R. Soc. Lond. B 164: 362–379. [DOI] [PubMed] [Google Scholar]
- Cheng, Y. M., and B. Y. Lin, 2003. Cloning and characterization of maize B chromosome sequences derived from microdissection. Genetics 164: 299–310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng, Y. M., and B. Y. Lin, 2004. Molecular organization of large fragments in the maize B chromosome: indication of a novel repeat. Genetics 166: 1947–1961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiavarino, A. M., M. Rosato, C. A. Naranjo, J. Camara Hernandez and L. Poggio, 1995. B chromosome polymorphism in N. Argentine populations of maize. Maize Genet. Coop. News Lett. 69: 94. [Google Scholar]
- Chiavarino, A. M., M. Rosato, P. Rosi, L. Poggio and C. A. Naranjo, 1998. Localization of the genes controlling B chromosome transmission rate in maize (Zea mays ssp. mays, Poaceae). Am. J. Bot. 85: 1581–1585. [PubMed] [Google Scholar]
- Chiavarino, A. M., M. Rosato, S. Manzanero, G. Jiménez, M. González-Sánchez et al., 2000. Chromosome nondisjunction and instabilities in tapetal cells are affected by B chromosomes in maize. Genetics 155: 889–897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiavarino, A. M., M. González Sánchez, L. Poggio, J. Puertas, M. Rosato et al., 2001. Is maize B chromosome preferential fertilization controlled by a single gene? Heredity 86: 743–748. [DOI] [PubMed] [Google Scholar]
- Colombo, P., and V. A. Confalonieri, 2004. Cytogeography and the evolutionary significance of B-chromosomes in relation to inverted rearrangements in a grasshopper species. Cytogenet. Genome Res. 106: 351–358. [DOI] [PubMed] [Google Scholar]
- Confalonieri, V., A. Sequeira, L. Todaro and J. C. Vilardi, 1998. Mitochondrial DNA and phylogeography of the grasshopper Trimerotropis pallidipennis in relation with clinal distribution of chromosome polymorphisms. Heredity 81: 444–452. [Google Scholar]
- Dellaporta, I. K., J. Wood and J. B. Hicks, 1983. A plant DNA minipreparation: version II. Plant Mol. Biol. Rep. 1: 19–21. [Google Scholar]
- Dherawattana, A., and K. Sadanaga, 1973. Cytogenetics of a crown, rust-resistant hexaploid oat with 42 + 2 fragment chromosomes. Crop Sci. 13: 591–594. [Google Scholar]
- Frank, S. A., 2000. Polymorphism of attack and defense. Trends Ecol. Evol. (Amst.) 15: 167–171. [DOI] [PubMed] [Google Scholar]
- Gaggiotti, O. E., O. Lange, K. Rassmann and C. Gliddon, 1999. A comparison of two indirect methods for estimating average levels of gene flow using microsatellite data. Mol. Ecol. 8: 1513–1520. [DOI] [PubMed] [Google Scholar]
- Gockel, J., W. J. Kennington, A. Hoffmann, D. B. Goldstein and L. Partridge, 2001. Nonclinality of molecular variation implicates selection in maintaining a morphological cline of Drosophila melanogaster. Genetics 158: 319–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- González-Sánchez, M., E. González-González, F. Molina, A. M. Chiavarino, M. Rosato et al., 2003. One gene determines maize B chromosome accumulation by preferential fertilization; another gene(s) determines their meiotic loss. Heredity 90: 122–129. [DOI] [PubMed] [Google Scholar]
- González-Sánchez, M., M. Rosato, M. Chiavarino and M. J. Puertas, 2004. Chromosome instabilities and programmed cell death in tapetal cells of maize with B chromosomes and effects on pollen viability. Genetics 166: 999–1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goudet, J., 2001. FSTAT, a program to estimate and test gene diversities and fixation indices,version 2.9.3. (http://www.unil.ch/popgen/softwares/fstat.htm)
- Gubitz, T., R. S. Thorpe and A. Malhotra, 2000. Phylogeography and natural selection in the Tenerife gecko Tarentola delalandii: testing historical and adaptive hypotheses. Mol. Ecol. 9: 1213–1221. [DOI] [PubMed] [Google Scholar]
- Hanson, G. P., 1969. B-chromosome-stimulated crossing over in maize. Genetics 63: 601–609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardy, O. J., and X. Vekemans, 2002. SPAGeDi: a versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2: 618–620. [Google Scholar]
- Hardy, O. J., N. Charbonnel, H. Fréville and M. Heuertz, 2003. Microsatellite allele sizes: a simple test to assess their significance on genetic differentiation. Genetics 163: 1467–1482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchinson, D. W., and A. R. Templeton, 1999. Correlation of pairwise genetic and geographic distance measures: Inferring the relative influences of gene flow and drift on the distribution of genetic variability. Evolution 53: 1898–1914. [DOI] [PubMed] [Google Scholar]
- Jenkins, G., and R. N. Jones, 2004. B chromosomes in hybrids of temperate cereals and grasses. Cytogenet. Genome Res. 106: 314–319. [DOI] [PubMed] [Google Scholar]
- Jones, R. N., 1985. Are B chromosomes selfish?, pp. 397–425 in The Evolution of Genome Size, edited by T. Cavalier-Smith. John Wiley & Sons, New York.
- Jones, R. N., 1995. Tansley Review No. 85. B chromosome in plants. New Phytol. 131: 411–434. [DOI] [PubMed] [Google Scholar]
- Jones, R.N., and A. Houben, 2003. B chromosomes in plants: escapees from the A chromosome genome? Trends Plant Sci. 8: 417–423. [DOI] [PubMed] [Google Scholar]
- Jones, R. N., and H. Rees, 1982. B Chromosomes. Academic Press, New York.
- Kimura, M., and J. F. Crow, 1964. The number of alleles that can be maintained in a finite population. Genetics 49: 725–738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura, M., and T. Ohta, 1978. Stepwise mutation model and distribution of allelic frequencies in finite populations. Proc. Natl. Acad. Sci. USA 75: 2868–2872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamb, J. C., N. C. Riddle, Y. M. Cheng, J. Theuri and J. A. Birchler, 2007. Localization and transcription of a retrotransposon-derived element on the maize B chromosome. Chromosome Res. 15: 383–398. [DOI] [PubMed] [Google Scholar]
- Lewontin, R. C., and J. Krakauer, 1973. Distribution of gene frequency as a test of the theory of the selective neutrality of polymorphisms. Genetics 74: 175–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lia, V. V., M. Bracco, A. M. Gottlieb, L. Poggio and V. A. Confalonieri, 2007. Complex mutational patterns and size homoplasy at maize microsatellite loci. Theor. Appl. Genet. (in press). [DOI] [PubMed]
- Longley, A. E., 1938. Chromosomes of maize from North American Indians. J. Agric. Res. 56: 177–195. [Google Scholar]
- Mantel, N., 1967. The detection of disease clustering and a generalized regression approach. Cancer Res. 27: 209–220. [PubMed] [Google Scholar]
- McClintock, B., T. A. Kato and A. Blumenschein, 1981. Chromosome Constitution of the Races of Maize. Colegio de Postgraduados, Chapingo, México.
- Miao, V. P., S. F. Convert and H. D. VanEtten, 1991. A fungal gene for antibiotic resistance on a dispensable (B) chromosome. Science 254: 1773–1776. [DOI] [PubMed] [Google Scholar]
- Naranjo, C. A., A. M. Chiavarino, M. Rosato, E. Quintela Fernández and L. Poggio, 1995. Tamaño del genoma y polimorfismo para cromosomas B en razas nativas argentinas y bolivianas de maiz., pp. 969–980. Abstracts of the III Latin American and XVI Andean Zone of Maize Researchers Meeting, Bolivia.
- Peacock, W. J., E. S. Dennis, M. M. Rhoades and A. J. Pryor, 1981. Highly repeated DNA sequence limited to knob heterochromatin in maize. Proc. Natl. Acad. Sci. USA 78: 4490–4494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perfectti, F., M. P. Ita, C. G. de la Vega, J. Gosalvez and J. P. M. Camacho, 2004. Spatio-temporal dynamics of a neutralized B chromosome in the grasshopper Eyprepocnemis plorans. Cytogenet. Genome Res. 106: 376–385. [DOI] [PubMed] [Google Scholar]
- Poggio, L., M. Rosato, A. M. Chiavarino and C. A. Naranjo, 1998. Genome size and environmental correlations in maize (Zea mays ssp. mays, Poaceae). Ann. Bot. 82: 107–115. [Google Scholar]
- Porter, H. L., and A. L. Rayburn, 1990. B-chromosome and C-band heterochromatin variation in Arizona maize populations adapted to different altitudes. Genome 33: 659–662. [Google Scholar]
- Pryor, A., K. Faulkner, M. M. Rhoades and W. J. Peacock, 1980. Asynchronous replication of heterochromatin of maize. Proc. Natl. Acad. Sci. USA 77: 6705–6709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raymond, M., and F. Rousset, 1995. GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J. Hered. 86: 248–249. [Google Scholar]
- Reeves, G., D. Francis, M. S. Davies, H. J. Rogers and T. R. Hodkinson, 1998. Genome size is negatively correlated with altitude natural populations of Dactylis glomerata. Ann. Bot. 82: 99–105. [Google Scholar]
- Rhoades, M. M., and E. Dempsey, 1972. On the mechanism of chromatin loss induced by the B chromosome of maize. Genetics 71: 73–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhoades, M. M., and E. Dempsey, 1973. Chromatin elimination induced by the B chromosome of maize. J. Hered. 64: 13–18. [Google Scholar]
- Rice, W., 1989. Analysing tables of statistical tests. Evolution 43: 223–225. [DOI] [PubMed] [Google Scholar]
- Riera, L., E. Petitpierre, C. Juan, J. Cabrero and J. P. M. Camacho, 2004. Evolutionary dynamics of a B chromosome invasion in island populations of the grasshopper Eyprepocnemis plorans. J. Evol. Biol. 17: 716–719. [DOI] [PubMed] [Google Scholar]
- Roman, H., 1947. Mitotic nondisjunction in the case of interchanges involving the B type chromosome in maize. Genetics 32: 391–409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roman, H., 1948. Directed fertilization in maize. Proc. Natl. Acad. Sci. USA 34: 36–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosato, M., A. M. Chiavarino, M. J. Puertas, C. A. Naranjo and L. Poggio, 1996. Genetic control of the B chromosome transmission rate in Zea mays (Poaceae). Am. J. Bot. 83: 1107–1112. [Google Scholar]
- Rosato, M., A. M. Chiavarino, C. A. Naranjo, J. Cámara Hernández and L. Poggio, 1998. Genome size and numerical polymorphism for the B chromosome in races of maize (Zea mays ssp. mays, Poaceae). Am. J. Bot. 85: 168–174. [PubMed] [Google Scholar]
- Schlötterer, C., and T. Wiehe, 1999. Microsatellites a neutral marker to infer selective sweeps, pp. 238–248 in Microsatellites - Evolution and Applications, edited by D. Goldstein and C. Schlötterer. Oxford University Press, London.
- Slatkin, M., 1995. A measure of population subdivision based on microsatellite allele frequencies. Genetics 139: 457–462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storz, J. F., 2002. Contrasting patterns of divergence in quantitative traits and neutral DNA markers: analysis of clinal variation. Mol. Ecol. 11: 2537–2551. [DOI] [PubMed] [Google Scholar]
- Storz, J. F., and J. M. Dubach, 2004. Natural selection drives altitudinal divergence at the albumin locus in deer mice, Peromyscus maniculatus. Evolution 58: 1342–1352. [DOI] [PubMed] [Google Scholar]
- Vigouroux, Y., J. S. Jaqueth, Y. Matsuoka, O. S. Smith, W. D. Beavis et al., 2002. Rate and pattern of mutation at microsatellite loci in maize. Mol. Biol. Evol. 19: 1251–1260. [DOI] [PubMed] [Google Scholar]
- Weir, B., and C. C. Cockerham, 1984. Estimating F-statistics for the analysis of population structure. Evolution 38: 1358–1370. [DOI] [PubMed] [Google Scholar]
- White, M. J. D., 1973. Animal Cytology and Evolution. Oxford University Press, London.
- Wright, S., 1978. Evolution and the Genetics of Populations, Volume 4: Variability Within and Among Natural Populations. University of Chicago Press, Chicago.