

Abstract
The contractile responses to capsaicin and anandamide, exogenous and endogenous agonists for transient receptor potential vanilloid receptor 1 (TRPV1), respectively, were investigated in muscle strips isolated from the rat urinary bladder. Capsaicin and anandamide produced concentration-dependent contractions of the muscle strips. The contractile response induced by capsaicin disappeared within approximately 20 min. In contrast, anandamide produced contractile responses lasting at least for 30 min. Capsaicin produced additive contractile responses in anandamide-treated muscle strips. The contractile response to anandamide was attenuated, but not abolished in strips desensitized by capsaicin. The response to capsaicin was abolished in the presence of a TRPV1 antagonist, N-(4-Tertiarybutylphenyl)-4-(3-chlorphyridin-2-yl)tetrahydropyrazine-1(2H)-carbox-amid e (BCTC), but not altered in the presence of either tetrodotoxin, atropine or indomethacin. In the presence of SR140333, a tachykinin NK1 receptor antagonist or SR48968, an NK2 receptor antagonist, the response to capsaicin was attenuated. The response to anandamide was partially attenuated in the presence of ONO8130, a prostanoid EP1 receptor antagonist, URB597, a fatty-acid amide hydrolase inhibitor, BCTC, SR140333 or SR48968, and almost completely abolished by indomethacin. Neither tetrodotoxin, atropine, a cannabinoid CB1 receptor antagonist, AM251, nor a cannabinoid CB2 receptor antagonist, AM630, had any effect on the response to anandamide. These results indicate that capsaicin produces muscle contractions by stimulating the TRPV1 receptor, followed by release of neuropeptides that can activate tachykinin NK1 and/or NK2 receptors in the bladder and that the contractile response to anandamide is mediated at least in part by activation of prostanoid EP1 receptors due to production of prostaglandins in addition to TRPV1 receptor activation.
Keywords: TRPV1, prostanoid EP1 receptor, capsaicin, anandamide, rat, urinary bladder
1. Introduction
The transient receptor potential vanilloid receptor 1 (TRPV1) is a nonselective cationic channel that responds to various noxious stimuli such as heat above 43°C, acid less than pH 6 and naturally occurring vanilloid capsaicin, one of the primary ingredients of hot chili peppers (Caterina et al., 1997). Studies with transgenic mice lacking functional TRPV1 receptors have provided evidence for a role of this receptor in the perception of thermal and inflammatory pain (Caterina et al., 2000; Davis et al., 2000). In the lower urinary tract, TRPV1 expression has been documented not only in nerve fibers within the bladder muscular, submucosal and mucosal layers, but also in urothelial cells (Birder et al., 2001), and animal studies have shown that TRPV1 has an important role in bladder pathophysiology in the context of pain (Vizzard, 2000) as well as bladder overactivity accompanying cystitis (Sculptoreanu et al., 2005) or spinal cord injury (de Groat, 1997).
The fatty acid amide anandamide was isolated from porcine brain and characterized as an endocannabinoid with an ability to bind cannabinoid receptors (Devane et al., 1992). In addition, since anandamide is structurally related to capsaicin, it is also known as an agonist for TRPV1 (Smart et al., 2000). So far, in the lower urinary tract, only anandamide has been thoroughly studied as an endovanilloid. Dinis et al (2004) revealed that the anandamide content increased in rat cystitis bladders in proportion to the development of hyperactivity. Furthermore, a high concentration of exogenous anandamide application or the blockade of endogenous anandamide degradation in naive bladders increased the frequency of bladder reflex contractions in a capsazepine dependent manner. Thus, anandamide seems to contribute to development of bladder overactivity via TRPV1 activation (Avelino et al., 2006).
However, it is not known whether capsaicin and anandamide can modulate detrusor smooth muscle activity in a similar manner via TRPV1 receptors. In this study, we therefore compared the contractile effect of capsaicin and anandamide in muscle strips isolated from the rat urinary bladder to clarify the differential mechanisms for the capsaicin or anandamide-mediated control of detrusor muscle function.
2. Materials and Methods
2.1. Drugs
Capsaicin, carbachol, tetrodotoxin, atropine sulfate, indomethacin were purchased from Sigma (St. Louis, MO, USA). Anandamide, N-(Piperidin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide (AM251) and 6-Iodo-2-methyl-1-[2-(4-morpholinyl)ethyl]-1H-indol-3-yl (4-methoxyphenyl)methanone (AM630) were purchased from Tocris (Ellisville, MO, USA).
N-(4-Tertiarybutylphenyl)-4-(3-chlorphyridin-2-yl)tetrahydropyrazine-1(2H)-carbox-amide (BCTC) was purchased from BIOMOL (Butler Pike, PA, USA). Cyclohexyl carbamic acid 3-carbamoyl-biphenyl-3-yl ester (URB597) was purchased from Cayman Chemical(Ann Arbor, MI, USA).
(S)1-(2-[3-(3,4-dichlorophenyl)-1-(3-isopropoxyphenylacetyl)piperidin-3-yl]ethyl)-4-phenyl-1-azoniabicyclo[2.2.2]octane chloride (SR140333), (S)-N-methyl-N[4-(4-acetylamino-4-phenyl piperidino)-2-(3,4-dichlorophenyl) butyl]benzamide (SR48968), (S)-(-)-N-(alpha-ethylbenzyl)-3-hydroxy-2-phenylquinoline-4-carboxamide (SB223412), and 4-{[(6-{isobutyl[(4-methyl-1,3-thiazol-2-yl)sulfonyl]amino}-2,3-dihydro-1H-inden-5-yl)oxy]methyl}benzoic acid (ONO8130) were prepared by Astellas Pharma INC. (Tsukuba, Japan). Capsaicin, indomethacin, BCTC, SR140333, SR48968, SB223412, URB597 and ONO8130 were dissolved in ethanol and serially diluted to desired concentrations. AM251 and AM630 were dissolved in dimethylsulfoxide and serially diluted to desired concentrations. Other drugs were dissolved in distilled water. The reported concentrations are described as the final bath concentrations. The final bath concentration of ethanol or dimethylsulfoxide never exceeded 0.1%.
2.2 Tissue preparation
Male Sprague-Dawley rats weighing 230–315 g were purchased from Hilltop Animal Care (Pittsburgh, PA, USA). Care and handling of animals were in accordance with institutional guidelines and approved by the University of Pittsburgh Institutional Animal Care and Use Committee. Rats were anesthetized with halothane (Halocarbon Laboratories, NJ, USA) and the urinary bladder was quickly removed. The detrusor muscle was excised into strips approximately 10 mm long and 2 mm wide. The strips were vertically suspended in a 30 ml organ bath containing Krebs-Henseleit solution (composition in mM: NaCl 118.0, CaCl2 2.5, KH2PO4 1.2, KCl 4.7, MgSO4 1.2, NaHCO3 25.0, glucose 10.0) maintained at 37°C and gassed with a mixture of 95% O2 and 5% CO2 to maintain pH 7.4. Strips were attached to isometric force displacement transducers through a carrier amplifier (Myobath system, World Precision Instruments, FL, USA), and contractile tones were recorded using a data acquisition system equipped with an analog-to-digital converter (Power Lab, AD Instruments, CO, USA). Baseline tension was set at 1.0 g and the strips were allowed to equilibrate for approximately 60 min before exposure to compounds.
2.3. Measurement of the effects of various substances
In all experiments, carbachol (10−6M) was initially added, and after the contractile response had reached plateau, the strips were washed and allowed to equilibrate for further 30 min. Contractile tone was measured following cumulative application of capsaicin (10−8-10−6M) and anandamide (10−6-10−4M). The contact times were about 1 min for capsaicin (10−8-10−6M) and anandamide (10−6M), and 2–5 min for anandamide (3×10−6-10−4M). The subsequent doses of capsaicin and anandamide were applied when the response had reached steady plateau approximately. In other experiments, after the strips were exposed to capsaicin (10−6M) or anandamide (10−4M) for 30 min, a contractile response to anandamide (10−4M) or capsaicin (10−6M) was respectively examined.
In a separate series of experiments, concentration-response curves were constructed with capsaicin (10−8-10−6M) and anandamide (10−6-10−4M) in the presence of one of the following compounds: tetrodotoxin (10−6M), atropine (3×10−6M), indomethacin (10−5M) (Craib et al., 2001), capsaicin (10−5M) (Harrison et al., 2003), SR140333, SR48968, SB223412 (tachykinin NK1, NK2 and NK3 receptor antagonists, respectively) (10−7-10−6M) (Harrison et al., 2003) (Trevisani et al., 2005) (Sarau et al., 1997), AM251, AM630 (cannabinoid CB1 and CB2 receptor antagonists, respectively) (10−6M) (White et al., 2001), BCTC (a TRPV1 antagonist) (10−6M) (Behrendt et al., 2004), URB597 (a fatty-acid amide hydrolase (FAAH) inhibitor) (10−6M) (Ho et al., 2007) and ONO8130 (a prostanoid EP1 receptor antagonist) (10−6M). These substances were added 15 min prior to the application of capsaicin or anandamide and their concentrations that show the maximal and specific effects were chosen based on the results in previous studies and our preliminary experiments.
2.4. Analysis of result
All data values are expressed as mean ± S.E.M.. Contractile responses are expressed as a percentage (%) of the response to carbachol (10−6M). Statistical significance was determined with Student's t-test or ANOVA followed by Dunnett's multiple test. P-values less than 0.05 were considered to be significant. All data analyses were performed using the SAS statistical software (SAS Institute, Cary, NC, USA).
3. Results
3.1. The comparison of the contractile responses to capsaicin and anandamide in muscle strips isolated from the rat urinary bladder
Capsaicin produced a rapidly developing phasic contractions within 1 min in a concentration-dependent manner (10−8-10−6M) in muscle strips. In contrast, anandamide (10−6-10−4M) concentration-dependently produced a slowly developing tonic contractions which peaked in 2–3 min after the application (Fig. 1 and Fig. 2A). The peak responses of capsaicin (10−6M) and anandamide (10−4M) were comparable (capsaicin: 45.8±7.2 % of 10−6M carbachol, anandamide: 47.2±6.4 % of 10−6 M carbachol, respectively), although the concentration-dependent curve of anandamide did not reach the maximum responses. The contractile response induced by capsaicin (10−6M) gradually disappeared within approximately 20 min. In contrast, the contractile response induced by anandamide (10−4) was maintained for at least 30 min (Fig. 1 and 2B).
Fig. 1.

Representative tracings showing the contractitle responses to capsaicin and anandamide in muscle strips isolated from the rat urinary bladder.
Fig. 2.

The comparison of the contractiles response to capsaicin and anandamide in muscle strips isolated from the rat urinary bladder A) Concentration response curves of capsaicin and anandamide. B) Time couses of disappearance of contractile responses to capsaicin and anandamide. Results are expressed as a percentage of the maximal response induced by 10−6M carbachol (CCh). Each point represents the mean ± S.E.M. of six to seven experiments.
After the cumulative responses of capsaicin (10−8-10−6), a second application of capsaicin (10−6M) failed to produce the contractile responses (Fig. 3). In these acutely desensitized strips by capsaicin, the contractile response to successive application of anandamide (10−4M) was attenuated, but not abolished (39.8±11.2% inhibition) (Fig. 3 and Fig. 4). Similar results were obtained when a higher concentration of capsaicin (10−5M) was used (Fig. 6C). In addition, capsaicin (10−6M) produced additive contractile responses in anandamide-treated muscle strips (29.9±3.3 % of 10−6M carbachol, n=5, data not shown).
Fig. 3.

Representative tracings showing the cross reaction between capsaicin and anandamide in muscle strips isolated from the rat urinary bladder.
Fig. 4.
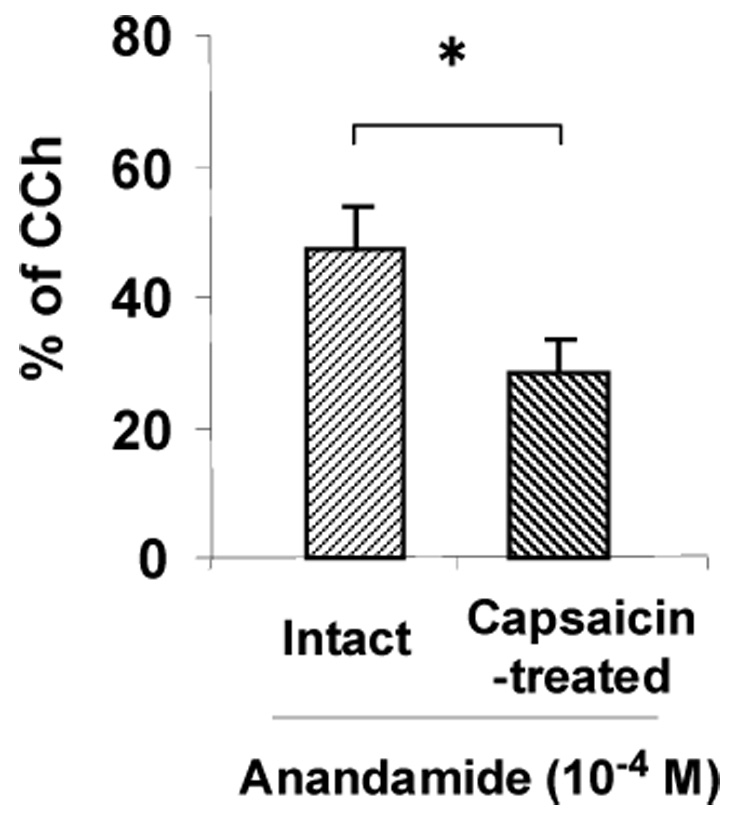
The contractile response to anandamide in capsaicin-desensitized strips. Results are expressed as a percentage of the maximal response induced by 10–6M carbachol (CCh). Each bar represents the mean ± S.E.M. of six to seven experiments. * P<0.05: significant difference to the intact group (Student’s t-test).
Fig. 6.

The effects of various drugs on contractile responses to anandamide in muscle strips isolated from the rat urinary bladder. A) Tetrodotoxin and atropine. B) AM251, a cannabinoid CB1 receptor antagonist and AM630, a cannabinoid CB2 receptor antagonist. C) Capsaicin-desensitization and BCTC. D) SR140333 and SR48968. E) Indomethacin, URB597, a FAAH inhibitor and ONO8130, a prostanoid EP1 receptor antagonist. Results are expressed as a percentage of the maximal response induced by 10−6M carbachol (CCh). Each bar reprsents the mean ± S.E.M. of four to seven experiments. * P<0.05, ** P<0.01: significant difference to the intact group (Dunnett’s multiple comparison test).
3.2. The effects of various drugs on contractile responses to capsaicin in muscle strips isolated from the rat urinary bladder
The contractile response to capsaicin (10−8-10−6M) was not altered in the presence of either tetrodotoxin (10−6M), atropine (3×10−6M) or indomethacin (10−5M), whereas BCTC (10−6M) completely suppressed it (94.2–100%) (Fig. 5A). In the presence of SR140333 (10−6M), a tachykinin NK1 receptor antagonist, or SR48968 (10−6–M), an NK2 receptor antagonist, the response to capsaicin (3×10−8-3×10−6M) was attenuated (80.7–98.5% and 54.9–97.4% inhibition, respectively) (Figs. 5B and 5C). In addition, SB223412 (10−6M), a tachykinin NK3 receptor antagonist, had no effect on the response to capsaicin (data not shown).
Fig. 5.

The effects of various drugs on contractile responses to capsaicin in muscle strips isolated from the rat urinary bladder. A) Tetrodotoxin, atropine, indomethacin and BCTC. B) SR140333, a tachykinin NK1 receptor antagonist. C) SR48968, a tachykinin NK2 receptor antagonist. Results are expressed as a percentage of the maximal response induced by 10−6M carbachol (CCh). Each bar represents the mean ± S.E.M. of four to seven experiments. * P<0.05, ** P<0.01: significant difference to the intact group (Dunnett’s multiple comparison test).
3.3. The effects of various drugs on contractile responses to anandamide in muscle strips isolated from the rat urinary bladder
Neither tetrodotoxin (10−6M), atropine (3×10−6M), AM251 (10−6M), a cannabinoid CB1 receptor antagonist, nor AM630 (10−6M), a cannabinoid CB2 receptor antagonist, altered the response to anandamide (Fig. 6A, B). In acutely desensitized strips by a high concentration of capsaicin (10−5), the contractile responses to anandamide (10−5 and 3×10−5M) were attenuated (92.3±1.5% and 68.5±9.4% inhibition, respectively) (Fig. 6C). In addition, anandamide (3×10−6 and 10−5M) -induced bladder contractions were decreased (98.2±6.1% and 67.4±10.4% inhibition, respectively) after BCTC (10−6M) application (Fig. 6C). In the presence of SR140333 (10−6M) or SR48968 (10−6M), the responses to anandamide (10−4M) were attenuated (58.2±11.1% and 55.2±9.8% inhibition, respectively) (Figs. 6D). The responses to anandamide (10−5 and 3×10−5M) were partially attenuated (94.3±3.6% and 75.5±11.7% inhibition, respectively) in the presence of ONO8130, a prostanoid EP1 receptor antagonist, and the responses to 10−4M of anandamide was also attenuated (48.0±6.6% inhibition) in the presence of URB597, a FAAH inhibitor. Indomethacin (10−5M), a cyclooxygenase inhibitor, almost completely abolished (79.7–100%) the anandamide (10−5-10−4M) -induced contraction (Fig. 6E).
4. Discussion
In the present study, the contractile responses to capsaicin and anandamide, exogenous and endogenous agonists for TRPV1, respectively, were investigated in muscle strips isolated from the rat urinary bladder.
Capsaicin produced rapidly developing phasic contractions in muscle strips and the contractile response gradually disappeared. In contrast, anandamide produced slowly developing and tonic long-lasting contractions, and anandamide was less efficacious than capsaicin in developing the contraction. Thus, the present study showed some differences between capsaicin and anandamide in contractile responses. The previous studies have demonstrated the contractile response to capsaicin (Santicioli et al., 1986) and anandamide (Harrison et al., 2003) in rat urinary bladder strips, and the contractile concentration ranges of these drugs in the present study were similar to those observed in previous studies. However, there are no studies that compared the time-course of contractile response between capsaicin and anandamide in muscle strips isolated from the rat urinary bladder. It is well known that the exposure of capsaicin evokes desensitization of TRPV1 receptors, while anandamide lacks a desensitizing effect on TRPV1 probably due to different allosteric modification of the TRPV1 receptor (Nagy et al., 2004). Therefore, in regard to the time course of contractile response through TRPV1, the different characteristic in desensitization mechanisms between capsaicin and anandamide might account for our results.
As indicated by previous investigations (Maggi et al., 1985, Santicioliet al., 1986), the contractile response to capsaicin was not altered in the presence of either tetrodotoxin or atropine. It has been documented that capsaicin-induced tetrodotoxin-resistant contractions may be due to the release of neurotransmitters from sensory nerve endings, followed by the stimulation of both tachykinin NK1 and NK2 receptors in the smooth muscle of the rat urinary bladder, and that the co-administration of tachykinin NK1 and NK2 receptor antagonists seem necessary to suppress capsaicin-induced contractions of rat bladders (Maggi et al., 1991). In the present study, BCTC that was sufficient to block TRPV1 (Valenzato et al., 2003) completely suppressed the contraction induced by capsaicin, indicating that this response was clearly mediated by TRPV1. In addition, in the presence of SR140333 or SR48968, which was sufficient to block tachykinin NK1 or NK2 receptor (Capasso et al., 2002), respectively, the response to capsaicin was attenuated while SB223412, a selective tachykinin NK3 receptor antagonist (Sarau et al., 1997; Zaratin et al., 2000), had no effect on the response to capsaicin. Thus, it seems likely that tachykinin NK1 and NK2, but not NK3 receptor, are involved in the capsaicin-induced contraction of the rat urinary bladder.
Anandamide is an endocannabinoid and it can bind to and activate cannabinoid receptors (Devane et al., 1992). However, in the present study, AM251 or AM630, which was sufficient to block cannabinoid CB1 or CB2 receptor (White et al., 2001), respectively, had no effect on the anandamide-induced contraction, indicating no involvement of cannabinoid CB1 or CB2 receptors. The contractile response to anandamide was not altered in the presence of either tetrodotoxin or atropine, but almost completely abolished by indomethacin in the rat urinary bladder. In terms of the contractile response to anandamide, the study using guinea-pig distal colon has reported that anandamide produced a sustained contraction in the presence of indomethacin and this response was abolished tetrodotoxin or atropine (Kojima et al., 2002). These results suggest the mechanism by which anandamide induces contractions could be different among tissues and/or animal species. The present study further revealed that the response to anandamide was partially attenuated in the presence of ONO8130, a prostanoid EP1 receptor antagonist that was sufficient to block prostanoid EP1 receptors in our preliminary study (data not shown). It has been reported that anandamide can be metabolized by cyclooxygenase-2 to prostaglandin E2 ethanolamine, which is able to contract the guinea pig trachea through prostanoid EP1 receptors (Ross et al., 2002). Alternatively, anandamide is degraded by FAAH to arachidonic acid, which can be a substrate for cyclooxygenases to yield prostaglandins (Burstein et al., 2000; Wahn et al., 2005). In the present study, URB597 that was sufficient to inhibit FAAH (Ho et al., 2007) attenuated the contraction induced by anandamide, indicating that this response was mediated by the production of prostaglandins. Thus, it is assumed that the contractile response to anandamide might be partly mediated by activation of prostanoid EP1 receptors due to the production of prostaglandins in the rat urinary bladder, and this mechanism should be taken into consideration especially when applying a high concentration of anandamide.
The present study also demonstrated that the contractile response to anandamide was significantly attenuated, but not abolished, in capsaicin-desensitized or BCTC-pretreated strips. On the other hand, in previous reports using bronchial rings or vagus nerves isolated from guinea-pigs, the contraction or depolarization responses induced by anandamide completely disappeared after capsaicin pre-treatment (Tucker et al., 2001, Kagaya et al., 2002). Thus, our findings suggest that contractile response to anandamide could be attributable not only to the stimulation to TRPV1, but also to other mechanisms in the rat urinary bladder. It has been demonstrated that prostanoid EP1 receptors in the peripheral afferent nerve and/or urothelium might facilitate afferent nerve activity (Ikeda et al., 2006). Furthermore, in the presence of SR140333 or SR48968, the contractile responses to anandamide were attenuated to the same level as seen in capsaicin-pretreated strips. Thus, it seems reasonable to assume that anandamide and its metabolites prostaglandins could stimulate C-fiber afferents directly by TRPV1 and/or indirectly by prostanoid EP1 receptors, then lead to the release of neuropeptides which can induce the contraction though both tachykinin NK1 and NK2 receptors in the rat urinary bladder.
In conclusion, capsaicin produces muscle contractions by stimulating the TRPV1 receptor, followed by release of neuropeptides that can activate tachykinin NK1 and/or NK2 receptors in the urinary bladder. In addition, the contractile response to anandamide is mediated at least in part by activation of prostanoid EP1 receptors due to production of prostaglandins in addition to TRPV1 receptor activation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Avelino A, Cruz F. TRPV1 (vanilloid receptor) in the urinary tract: expression, function and clinical applications. Naunyn-Schmiedebergs Arch. Pharmacol. 2006;373:287–299. doi: 10.1007/s00210-006-0073-2. [DOI] [PubMed] [Google Scholar]
- Behrendt HJ, Germann T, Gillen C, Hatt H, Jostock R. Characterization of the mouse cold-menthol receptor TRPM8 and vanilloid receptor type-1 VR1 using a fluorometric imaging plate reader (FLIPR) assay. Br. J. Pharmacol. 2004;141:737–745. doi: 10.1038/sj.bjp.0705652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birder LA, Nakamura Y, Kiss S, Nealen ML, Barrick S, Kanai AJ, Wang E, Ruiz G, De Groat WC, Apodaca G, Watkins S, Caterina MJ. Altered urinary bladder function in mice lacking the vanilloid receptor TRPV1. Nat. Neurosci. 2002;5:856–860. doi: 10.1038/nn902. [DOI] [PubMed] [Google Scholar]
- Burstein SH, Rossetti RG, Yagen B, Zurier RB. Oxidative metabolism of anandamide. Prostaglandins Other Lipid Mediat. 2000;61:29–41. doi: 10.1016/s0090-6980(00)00053-8. [DOI] [PubMed] [Google Scholar]
- Capasso R, Tavares IA. Effect of the flavonoid galangin on urinary bladder rat contractility in-vitro. J. Pharm. Pharmacol. 2002;54:1147–1150. doi: 10.1211/002235702320266334. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, Petersen-Zeitz KR, Koltzenburg M, Basbaum AI, Julius D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science. 2000;288:306–313. doi: 10.1126/science.288.5464.306. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816–824. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- Craib SJ, Ellington HC, Pertwee RG, Ross RA. A possible role of lipoxygenase in the activation of vanilloid receptors by anandamide in the guinea-pig bronchus. Br. J. Pharmacol. 2001;134:30–37. doi: 10.1038/sj.bjp.0704223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Groat WC. A neurologic basis for the overactive bladder. Urology. 1997;50:36–52. doi: 10.1016/s0090-4295(97)00587-6. [DOI] [PubMed] [Google Scholar]
- Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, Gibson D, Mandelbaum A, Etinger A, Mechoulam R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science. 1992;258:1946–1949. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- Dinis P, Charrua A, Avelino A, Yaqoob M, Bevan S, Nagy I, Cruz F. Anandamide-evoked activation of vanilloid receptor 1 contributes to the development of bladder hyperreflexia and nociceptive transmission to spinal dorsal horn neurons in cystitis. J. Neurosci. 2004;24:11253–11263. doi: 10.1523/JNEUROSCI.2657-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison S, De Petrocellis L, Trevisani M, Benvenuti F, Bifulco M, Geppetti P, Di Marzo V. Capsaicin-like effects of N-arachidonoyl-dopamine in the isolated guinea pig bronchi and urinary bladder. Eur. J. Pharmacol. 2003;475:107–114. doi: 10.1016/s0014-2999(03)02114-9. [DOI] [PubMed] [Google Scholar]
- Ho WS, Randall MD. Endothelium-dependent metabolism by endocannabinoid hydrolases and cyclooxygenases limits vasorelaxation to anandamide and 2-arachidonoylglycerol. Br. J. Pharmacol. 2007;150:641–651. doi: 10.1038/sj.bjp.0707141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeda M, Kawatani M, Maruyama T, Ishihama H. Prostaglandin facilitates afferent nerve activity via EP1 receptors during urinary bladder inflammation in rats. Biomed. Res. 2006;27:49–54. doi: 10.2220/biomedres.27.49. [DOI] [PubMed] [Google Scholar]
- Kagaya M, Lamb J, Robbins J, Page CP, Spina D. Characterization of the anandamide induced depolarization of guinea-pig isolated vagus nerve. Br. J. Pharmacol. 2002;137:39–48. doi: 10.1038/sj.bjp.0704840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kojima S, Sugiura T, Waku K, Kamikawa Y. Contractile response to a cannabimimetic eicosanoid, 2-arachidonoylglycerol, of longitudinal smooth muscle from the guinea-pig distal colon in vitro. Eur. J. Pharmacol. 2002;444:203–207. doi: 10.1016/s0014-2999(02)01650-3. [DOI] [PubMed] [Google Scholar]
- Maggi CA, Patacchini R, Santicioli P, Giuliani S. Tachykinin antagonists and capsaicin-induced contraction of the rat isolated urinary bladder: evidence for tachykinin-mediated cotransmission. Br. J. Pharmacol. 1991;103:1535–1541. doi: 10.1111/j.1476-5381.1991.tb09823.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maggi CA, Santicioli P, Meli A. Evidence for the involvement of endogenous substance P in the motor effects of capsaicin on the rat urinary bladder. J. Pharm. Pharmacol. 1985;37:203–204. doi: 10.1111/j.2042-7158.1985.tb05042.x. [DOI] [PubMed] [Google Scholar]
- Nagy I, Santha P, Jancso G, Urban L. The role of the vanilloid (capsaicin) receptor (TRPV1) in physiology and pathology. Eur. J. Pharmacol. 2004;500:351–369. doi: 10.1016/j.ejphar.2004.07.037. [DOI] [PubMed] [Google Scholar]
- Ross RA. Anandamide and vanilloid TRPV1 receptors. Br. J. Pharmacol. 2003;140:790–801. doi: 10.1038/sj.bjp.0705467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santicioli P, Maggi CA, Meli A. Functional evidence for the existence of a capsaicin-sensitive innervation in the rat urinary bladder. J. Pharm. Pharmacol. 1986;38:446–451. doi: 10.1111/j.2042-7158.1986.tb04608.x. [DOI] [PubMed] [Google Scholar]
- Sarau HM, Griswold DE, Potts W, Foley JJ, Schmidt DB, Webb EF, Martin LD, Brawner ME, Elshourbagy NA, Medhurst AD, Giardina GA, Hay DW. Nonpeptide tachykinin receptor antagonists: I. Pharmacological and pharmacokinetic characterization of SB 223412, a novel, potent and selective neurokinin-3 receptor antagonist. J. Pharmacol. Exp. Ther. 1997;281:1303–1311. [PubMed] [Google Scholar]
- Sculptoreanu A, de Groat WC, Buffington CA, Birder LA. Abnormal excitability in capsaicin-responsive DRG neurons from cats with feline interstitial cystitis. Exp. Neurol. 2005;193:437–443. doi: 10.1016/j.expneurol.2005.01.011. [DOI] [PubMed] [Google Scholar]
- Smart D, Gunthorpe MJ, Jerman JC, Nasir S, Gray J, Muir AI, Chambers JK, Randall AD, Davis JB. The endogenous lipid anandamide is a full agonist at the human vanilloid receptor (hVR1) Br. J. Pharmacol. 2000;129:227–230. doi: 10.1038/sj.bjp.0703050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trevisani M, Patacchini R, Nicoletti P, Gatti R, Gazzieri D, Lissi N, Zagli G, Creminon C, Geppetti P, Harrison S. Hydrogen sulfide causes vanilloid receptor 1-mediated neurogenic inflammation in the airways. Br. J. Pharmacol. 2005;145:1123–1131. doi: 10.1038/sj.bjp.0706277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker RC, Kagaya M, Page CP, Spina D. The endogenous cannabinoid agonist, anandamide stimulates sensory nerves in guinea-pig airways. Br. J. Pharmacol. 2001;132:1127–1135. doi: 10.1038/sj.bjp.0703906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valenzano KJ, Grant ER, Wu G, Hachicha M, Schmid L, Tafesse L, Sun Q, Rotshteyn Y, Francis J, Limberis J, Malik S, Whittemore ER, Hodges D. N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl)tetrahydropyrazine-1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic properties: I. in vitro characterization and pharmacokinetic properties. J. Pharmacol. Exp. Ther. 2003;306:377–386. doi: 10.1124/jpet.102.045674. [DOI] [PubMed] [Google Scholar]
- Vizzard MA. Alterations in spinal cord Fos protein expression induced by bladder stimulation following cystitis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000;278:R1027–R1039. doi: 10.1152/ajpregu.2000.278.4.R1027. [DOI] [PubMed] [Google Scholar]
- Wahn H, Wolf J, Kram F, Frantz S, Wagner JA. The endocannabinoid arachidonyl ethanolamide (anandamide) increases pulmonary arterial pressure via cyclooxygenase-2 products in isolated rabbit lungs. Am. J. Physiol. Heart Circ. Physiol. 2005;289:H2491–H2496. doi: 10.1152/ajpheart.00718.2005. [DOI] [PubMed] [Google Scholar]
- White R, Ho WS, Bottrill FE, Ford WR, Hiley CR. Mechanisms of anandamide-induced vasorelaxation in rat isolated coronary arteries. Br. J. Pharmacol. 2001;134:921–929. doi: 10.1038/sj.bjp.0704333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaratin P, Angelici O, Clarke GD, Schmid G, Raiteri M, Carita F, Bonanno G. NK3 receptor blockade prevents hyperalgesia and the associated spinal cord substance P release in monoarthritic rats. Neuropharmacology. 2000;39:141–149. doi: 10.1016/s0028-3908(99)00087-8. [DOI] [PubMed] [Google Scholar]