

Abstract
Ustilago maydis, a fungal pathogen of maize, alternates between budding and filamentous growth in response to mating and other environmental signals. Defects in components of the cAMP signaling pathway affect this morphological transition and reveal an association of budding growth with elevated cAMP levels and filamentous growth with low cAMP levels. We have identified two genes, adr1 and uka1, encoding catalytic subunits of cAMP-dependent protein kinase (PKA). Disruption of adr1 resulted in a constitutively filamentous growth phenotype similar to that of mutants deficient in adenylyl cyclase. Importantly, adr1 is required for pathogenicity and is responsible for the majority of PKA activity in fungal cells. In contrast, uka1 has little influence on pathogenicity, and deletion of the uka1 gene does not affect cell morphology. These results provide compelling evidence that regulated PKA activity is crucial during infectious development of U. maydis.
Pathogenesis, dimorphic growth, and sexual development are intricately interconnected in the corn smut fungus Ustilago maydis, and the perception of signals from the host plant is likely to play an important role in these processes (1, 2). Mating of haploid U. maydis cells, which are nonpathogenic and yeast-like, leads to the formation of infectious, dikaryotic hyphae. Dikaryons can be established in culture, but the resulting hyphae need the host environment to sustain mycelial proliferation. Infection of maize plants results in tumor induction and the eventual formation of masses of diploid teliospores.
The establishment of dikaryotic hyphae in U. maydis is controlled by two unlinked mating-type loci, a and b. The a locus encodes pheromones and pheromone receptors and mediates cell recognition and fusion (3). The multiallelic b locus encodes two homeodomain proteins bE and bW, which can associate to form heterodimers only if they are derived from different b alleles (4). The bE–bW heterodimers are believed to act as transcriptional regulators essential for orchestrating invasive, filamentous growth (2, 4).
Additional factors, besides mating, can affect the cellular morphology of U. maydis. Specifically, cells respond to high glucose concentrations with yeast-like, sporidial growth, whereas a limited supply of nutrients, exposure to air, or growth at low pH triggers filamentous growth for haploid cells (5–7). The cAMP pathway regulates at least one aspect of the transition between budding and filamentous growth because defects in the uac1 gene, encoding adenylyl cyclase, bypass the requirement for mating in the formation of a filamentous cell type (6).
cAMP controls the activity of cAMP-dependent protein kinase (PKA) by influencing the association of regulatory (R) and catalytic (C) subunits. Hence, the finding that uac1 mutants of U. maydis are filamentous indicated that low cAMP and low PKA activity trigger filamentous growth. However, haploid uac1 mutants were not pathogenic by themselves, a result anticipated given the essential role of the bE–bW heterodimer in pathogenicity. Interestingly, co-infection of maize seedlings with two adenylyl cyclase-deficient strains harboring compatible a and b mating-type alleles did not result in disease symptoms. This indicates that uac1 function is required for establishment of the infectious dikaryon (mating), for subsequent disease development, or for both processes (6).
Exogenous cAMP or mutations in the gene encoding the regulatory subunit of cAMP-dependent protein kinase (ubc1) restore budding growth to filamentous adenylyl cyclase mutants (6). However, wild-type budding was not restored by ubc1 suppression of the adenylyl cyclase defect; instead, a “multiple-budding” phenotype occurred, probably as a result of defects in bud site selection and cytokinesis (6). In addition, mating compatible ubc1 mutants were unable to form dikaryotic mycelium in culture, indicating that constitutively elevated PKA activity interferes with filament formation. However, co-injection of such strains into maize seedlings results in hyphal growth in planta, although tumor formation does not occur (8).
In this study, we address the role of two genes encoding catalytic subunits of PKA (adr1 and uka1) in the pathogenesis and filamentous growth of U. maydis. Surprisingly, uka1 mutants had no obvious defects in mating, morphogenesis, or virulence. In contrast, adr1 encodes the major catalytic subunit of PKA in U. maydis, and regulated Adr1 kinase activity is essential for the transition from budding to filamentous growth and for corn smut disease. These results further emphasize the emerging theme that the cAMP signal transduction pathway plays a crucial role in the virulence of plant pathogenic fungi (9).
MATERIALS AND METHODS
Strains and Growth Conditions.
The U. maydis mutants used in this study are listed in Table 1. Cell culture, mating tests, and plant inoculations were performed as previously described (10). Escherichia coli strains DH5α and DH10b (Bethesda Research Laboratories) were employed for plasmid construction, and strain BL21∷DE3 (Novagen) was used for protein expression.
Table 1.
U. maydis mutants
| Strain | Genotype (resistance) | Reference |
|---|---|---|
| d132-9 | a1/a2 b1∷hyg/b2 (Hygr) | 10 |
| C002P#17 | a1b1 uac1-2 (Phlr) | 6 |
| 521d | a1b1 ubc1-2 (Hygr) | 6 |
| 001uac−#18 | a2b2 uac1-2 (Phlr) | This work |
| 0505 | a2b2 Δubc1-4 (Phlr) | This work |
| 001-13 | a2b2 Δuka1-1 (Hygr) | This work |
| 001-44 | a2b2 Δuka1-1 (Hygr) | This work |
| 002-6 | a1b1 Δuka1-1 (Hygr) | This work |
| 001-12 | a2b2 adr1-1 (Phlr) | This work |
| 002-10 | a1b1 adr1-1 (Phlr) | This work |
| 001-44#5 | a2b2 Δuka1-1 adr1-1 (Hygr Phlr) | This work |
| 002-6#4 | a1b1 Δuka1-1 adr1-1 (Hygr Phlr) | This work |
| 002#5 | a1b1 adr1-1 ubc1-2 (Hygr Phlr) | This work |
| 001#9 | a2b2 Δuka1-1 Δubc1-3 (Hygr Phlr) | This work |
| 002#86 | a1b1 Δuka1-1 Δubc1-3 (Hygr Phlr) | This work |
| UD12 | a1/a2 b1/b2 Δubc1-3/ubc1 (Phlr) | This work |
| UD1201 | a1/a2 b1/b2 Δubc1-3/ubc1-2 (Hygr Phlr) | This work |
| UD1208 | a1/a2 b1/b2 ubc1-2/ubc1-2 (Hygr) | This work |
| d132#8 | a1/a2 b1/b2 adr1-1/adr1 (Phlr) | This work |
| d132#7 | a1/a2 b1/b2 adr1-1/adr1-1 (Phlr) | This work |
Isolation of the adr1 and uka1 Genes.
Standard procedures were followed for DNA manipulations and sequence analysis (11). The degenerate primers PKA5 5′-GTATCGAT(A/C/T)TA(C/T)(A/C)GIGA(C/T)(C/T)TIAA(A/G)CC-3′ and PKA3 5′-CACCGCGGIGCIA(G/A)(G/A)TA(T/C)TCI(T/G)GIGTICC-3′ (I = inosine; ClaI and SacII sites are underlined) were designed based on conserved amino acid residues in subdomains VI and VIII of the catalytic domain of protein kinases (12). A U. maydis λ ZAP cDNA library (6) was converted to a phagemid library and used as a template with primers PKA5 and PKA3 in PCR reactions (35 cycles; 1 min, 94°C; 1 min, 55°C; 1 min, 72°C). Two classes of PCR clones were identified. One class was identical to the U. maydis adr1 gene, encoding a homolog of PKA catalytic subunits (13). Therefore, primers 5′-CGGGATCCTATGTCTGCTATTCCAC-3′ and 5′-CGGGATCCTCAGAAATCCGGGAAAAG-3′ (BamHI sites underlined) were designed to amplify the adr1 gene from genomic U. maydis DNA. The resulting 1.2-kb PCR product was cloned in pBluescript II KS (Stratagene). The second class identified a separate PKA catalytic subunit gene designated uka1. Cosmids containing the uka1 gene were isolated from a U. maydis genomic library (14). DNA sequencing was carried out using AmpliTaq DyeDeoxy Terminator cycle sequencing technology (Applied Biosystems) and an Applied Biosystems automated DNA sequencer. DNA sequence database searches were carried out with the BLAST algorithm (15).
Disruption and Deletion Alleles of uka1, adr1, and ubc1.
The Δuka1-1 deletion allele was constructed by replacing two contiguous 230- and 870-bp XmnI fragments, encompassing 72% of the uka1 coding sequence and 250 bp of sequence upstream of the ATG, with a 2.5-kb hygromycin resistance cassette. The adr1-1 disruption allele was made by inserting a 1.9-kb phleomycin resistance cassette at the unique SnaBI site located at codon 171 of the adr1 coding sequence. The Δubc1-3 deletion allele was constructed by replacing a 530-bp ApaI–SphI fragment, encoding both cAMP binding sites, with a 1.9-kb phleomycin resistance cassette. The Δubc1-4 allele was derived from Δubc1-3 by the removal of codons 40–460 from the ubc1 gene by deletion of two contiguous XhoI fragments of 0.5 and 0.75 kb. The ubc1-2 disruption allele, in which a 3-kb hygromycin resistance cassette is inserted at a unique BglII site between the cAMP binding sites, has been described previously (6).
adr1-1 and Δuka1-1 single mutants were constructed in the compatible wild-type haploid strains 518 (a2 b2) and 521 (a1 b1) (10). One or both copies of the wild-type adr1 gene were also replaced in one step with the adr1-1 allele in the pathogenic diploid U. maydis strain d132 (a1/a2 b1/b2) to yield strains d132#8 and d132#7, respectively. This diploid was also used to construct mutants containing deletions or disruption alleles for one or both copies of the ubc1 gene. The Δuka1-1 strains 001-13 and 002-6 were transformed with the construct for Δubc1-3 to yield Δuka1-1 Δubc1-3 double mutants. The adr1-1 Δuka1-1 double mutants were constructed by transforming the Δuka1-1 strains 001-44 and 002-6 (Table 1) with the adr1-1 disruption construct. To generate adr1-1 ubc1-2 double mutants, ubc1-2 strain 521d (6) was transformed with the construct for adr1-1. U. maydis transformations were performed with linear fragments carrying the disruption or deletion alleles as described previously (16), and gene replacements were verified by hybridization. Gene names in U. maydis are represented in lower case italics (e.g., adr1) and mutated alleles are given an allele designation (e.g., adr1-1). A Δ symbol is used here in front of the names of alleles containing deletions. The names of proteins are not italicized (e.g., Adr1).
Overexpression and Purification of Ubc1.
A gene encoding full-length Ubc1 with a C-terminal His-tag fusion (Ubc1∷H6) was constructed by PCR using primers UBC54, 5′-CCGAATTCGCTACCATCTAGCTACAC-3′ and UBC34, 5′-CCGAATTCGGCGCGCTCCATGC-3′ (EcoRI sites are underlined). The resulting 1.6-kb PCR product was inserted at the EcoRI site of pET21b (Novagen) and transformed into BL21∷DE3. For expression of Ubc1∷H6, 200 ml of TYP (16 g/liter tryptone/16 g/liter yeast extract/5 g/liter NaCl/2.5 g/liter K2HPO4, pH 7.0) containing 200 μg/ml ampicillin, 2 mM MgCl2, and 0.2% glucose were inoculated with 400 μl of a washed overnight preculture of BL21∷DE3 (pET21b∷Ubc1). The culture was incubated for 4 h at 37°C to OD600 = 1.4, transferred to 28°C, and incubated for 30 min. Isopropyl β-d-thiogalactoside was then added to a final concentration of 0.25 mM, and incubation was continued for 3 h at 28°C. Cells were harvested, washed once with 40 mM Tris, pH 7.4, and cell pellets corresponding to 50 ml induced culture were stored at −70°C. Cell pellets were thawed on ice, resuspended in 5 ml of binding buffer BB (25 mM Tris, pH 7.9/0.5 M NaCl/5 mM imidazole/0.1% Triton X-100) and passed twice through a French press at 16,000 psi (1 psi = 6.89 kPa) internal pressure. Cell lysates were centrifuged at 20,000 × g for 20 min at 4°C, and the supernatant was passed through a 0.45-μm filter and loaded onto a 1-ml Ni-NTA column (Qiagen). The column was washed with 15 ml of BB, 9 ml of BB containing 50 mM imidazole, 9 ml of BB containing 100 mM imidazole and then eluted with BB containing 200 mM imidazole. The eluted Ubc1∷H6 protein was >95% pure, and the yield of purified Ubc1∷H6 was typically 1.5 mg per 50 ml of induced culture. The eluate was dialyzed against BB and stored at 4°C.
Enrichment and Assay of PKA Activity.
U. maydis strains 521d (ubc1-2), 002#5 (ubc1-2 adr1-1), and 002#86 (Δubc1-3 Δuka1-1), were grown in 200 ml of complete medium to mid-log phase (OD600 = 0.6–0.7); cells were harvested by centrifugation and washed once with UAB (20 mM Tris, pH 7.9/5 mM imidazole/150 mM NaCl), and cell pellets were frozen in liquid nitrogen and stored at −70°C. Cell pellets were thawed on ice, resuspended in 3 ml of UABC (UAB containing 0.1% Triton X-100, 1 mM phenylmethylsulfonyl fluoride, and 1 mM NaF), and passed twice through a French press at 18,000 psi internal pressure. Cell lysates were cleared by centrifugation at 40,000 × g for 20 min at 4°C. Cleared protein extracts (10–15 mg) were passed over Ubc1∷H6 affinity columns, which were prepared by fixing 0.5 mg of His-tagged recombinant Ubc1 protein on 0.5 ml of Ni-NTA agarose. Columns were washed twice with 10 ml of UABC, and bound PKA catalytic subunits were eluted with 1 mM cAMP in UABC (lacking Triton X-100). Five 1-ml elution fractions were collected, and 10-μl samples were assayed for transfer of 32PO4 from ATP to the PKA substrate Kemptide (LRRASLG) using a PKA assay kit (Pierce). Affinity purification on Ubc1 columns increased specific PKA activity of ubc1 extracts approximately 1000-fold. The cpm values obtained from the five cAMP elution fractions of the ubc1 uka1 and ubc1 adr1 double mutants were added and divided by the sum of the cpm values from the five cAMP elution fractions of the ubc1 mutant extracts. These relative values for PKA activity were corrected for the amount of protein extract loaded on the columns.
RESULTS AND DISCUSSION
Identification of PKA Catalytic Subunit Genes.
Two genes encoding homologs of PKA catalytic subunits were identified by PCR (see Materials and Methods). The first gene, adr1, had been isolated previously based on its ability to confer resistance to the fungicide vinclozolin (13), and the second gene, uka1 (Ustilago kinase A), was a novel gene. Sequence analysis revealed two in-frame ATG codons in the uka1 ORF that could potentially initiate translation of polypeptides of either 372 or 398 amino acids. The sequence context of the ATG for the predicted 372-amino acid polypeptide best matched the consensus sequence for translation initiation in filamentous fungi (CA(C/A)(A/C)TGNC; ref. 17). Both Adr1 and Uka1 have all of the primary amino acid sequence features of the protein kinase domain (12) including amino acids that are involved in the interaction with the regulatory subunit of PKA and a stretch of amino acid residues known to be required for high affinity binding of PKI, a peptide inhibitor of PKA (Fig. 1; refs. 18, 19).
Figure 1.

Amino acid sequence alignments of the two U. maydis PKA catalytic subunits Adr1 and Uka1 with fungal and mouse catalytic subunits of PKA. Amino acid identity (black boxes) and similarity (gray boxes) are shown within the protein kinase domain (C-terminal 80%). The 11 subdomains of the protein kinase catalytic domain are indicated with roman numerals, and the twelve most highly conserved residues are highlighted with filled circles (12). Filled diamonds indicate the three residues shown to be required for the association between catalytic and regulatory subunits of PKA (18). The autophosphorylation site is indicated with a filled square. The thick horizontal line shows residues implicated in binding of the PKI substrate (19). Be, B. emersonii (20); Mg, M. grisea (21); Sc, S. cerevisiae (22); Sp, S. pombe (23); Mm, Mus musculus (24). Sequences were aligned with the clustal w (1.60) program (25).
Pairwise alignments of the carboxyl-terminal regions encompassing the catalytic kinase core revealed that Adr1 and Uka1 are more closely related to PKA homologs from other fungi than they are to each other (Fig. 1). That is, Uka1 and Adr1 share only 51% sequence identity in this region. In contrast, Adr1 shares 68% sequence identity with CPKA from the rice blast fungus Magnaporthe grisea and 60–63% with PKA catalytic subunits from Schizosaccharomyces pombe, the aquatic fungus Blastocladiella emersonii, and Saccharomyces cerevisiae (Fig. 1). Uka1 has 56% amino acid sequence identity with the B. emersonii PKA catalytic subunit.
Adr1 Is the Principal Catalytic Subunit of PKA.
PKA assays were carried out to demonstrate that adr1 encodes a catalytic subunit of PKA and to evaluate the relative contributions of the uka1 and the adr1 genes to total cellular PKA activity. Catalytic subunits of PKA were enriched by affinity chromatography using columns containing recombinant Ubc1 protein (i.e., the PKA regulatory subunit). To ensure that all catalytic subunits were dissociated from the regulatory subunit and therefore capable of binding to the Ubc1 affinity column, protein extracts were prepared from strains carrying a mutation in the ubc1 gene. Specifically, PKA assays were performed on affinity-purified protein extracts from a ubc1 single mutant and from ubc1 adr1 and ubc1 uka1 double mutants. Fig. 2 shows the relative values for PKA activity in these strains. As expected from the genetic analysis of adr1 and uka1 mutants (see below), PKA activity (relative to the ubc1 mutant) was reduced approximately 10-fold in the adr1 ubc1 mutant (0.11 ± 0.07) and not significantly different for the uka1 ubc1 mutant (0.85 ± 0.31). Thus, Adr1 seems to be the major catalytic subunit of PKA in U. maydis.
Figure 2.

Contribution of adr1 and uka1 encoded enzymes to cellular PKA activity. Protein extracts from the ubc1 single mutant 521d (ubc1-2), the ubc1 uka1 double mutant 002#5 (ubc1-2 adr1-1), and the ubc1 adr1 double mutant 002#86 (Δubc1-3 Δuka1-1) were enriched for PKA activity by affinity chromatography on columns containing recombinant U. maydis regulatory subunit of PKA (Ubc1). The data shown represent averages from three independent experiments. PKA activities are presented as relative values for the ubc1 adr1 and ubc1 uka1 double mutants compared with the activity for the ubc1 mutant.
The uka1 and adr1 Genes Have Different Roles in Morphogenesis.
Mutants bearing disruptions or deletions of adr1 and uka1 genes were constructed in the two mating-compatible haploid strains 518 (a2 b2) and 521 (a1 b1) to explore the function of the genes in morphogenesis. Strains defective in uka1 formed smooth, yeast-like colonies comprising budding cells that were indistinguishable from wild-type cells (Fig. 3). In contrast, disruption of the adr1 gene resulted in a dramatic change in colony morphology in that adr1 mutants were covered with aerial hyphae, much like the uac1 mutants with a defect in adenylyl cyclase (6). In addition, adr1-1 cells formed filaments and cell aggregates in liquid medium, a morphological phenotype previously observed for uac1 mutants. Moreover, short single cells were also apparent in liquid cultures of adr1-1 strains (like uac1-1 mutants), especially when these cultures reached stationary phase.
Figure 3.
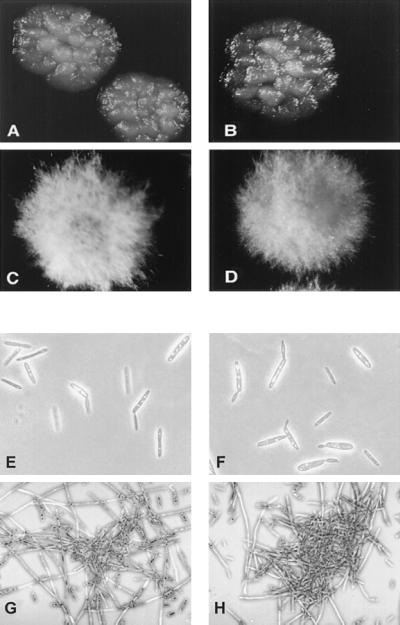
Colony and cell morphology of uka1 and adr1 mutants. Strains were grown on solid mating medium (A–D) or in liquid medium (E–H) (10, 26). The Δuka1-1 (A), adr1-1 (C), and uac1-2 (D) mutants were all derived from the wild-type strain 521 (B).
It should be noted that disruption of the adr1 gene in ubc1 mutants resulted in the same filamentous phenotype as displayed by adr1 mutants (data not shown), demonstrating that adr1 is epistatic to ubc1. In contrast, uka1 ubc1 double mutants display the multiple budding phenotype of ubc1 mutants (data not shown). This finding is consistent with the indication from the PKA assay that uka1 makes a relatively small contribution to total activity (Fig. 2). Double mutants defective for uka1 and adr1 were viable, suggesting that PKA function is dispensable in U. maydis for growth in culture or that U. maydis has additional genes encoding PKA catalytic subunits. There is preliminary evidence for a third catalytic subunit of PKA in U. maydis (G. Yang and J.W.K., unpublished data), and multiple genes encoding catalytic subunits of PKA are known in other fungi, such as S. cerevisiae (22). Surprisingly, the strains carrying mutations in both of the catalytic subunit genes were noticably less mycelial than the adr1 single mutants (data not shown). The reason for this unexpected phenotype is not clear. It also remains to be determined whether the enzymes encoded by uka1 and adr1 have different substrate specificities in addition to their different contributions to total PKA activity.
The adr1 Gene Is Required for Virulence.
Mixtures of compatible wild-type and mutant strains were injected into maize seedlings to evaluate the virulence of PKA catalytic subunit mutants (Table 2). Plants inoculated with two compatible adr1 mutants did not develop tumors and were essentially symptomless. Furthermore, adr1-1 disruption strains, in combination with compatible wild-type strains, induced tumors at a reduced level (tumors on 12% of the plants) when compared with infections caused by wild-type strains (tumors on 87% of the plants). This result suggests that adr1 mutants are impaired for mating because the defect in adr1 should have been complemented upon fusion of mutant and wild-type cells. In contrast to the findings for adr1, Δuka1 mutants were not affected in virulence (Table 2).
Table 2.
Virulence of mutants defective in genes encoding subunits of PKA
| Strains inoculated | No. of plants inoculated | No. of plants with tumors | % of plants with tumors |
|---|---|---|---|
| 518 × 521 | 15 | 13 | 87 |
| Δuka1-1 × uka1 | 46 | 42 | 91 |
| Δuka1-1 × Δuka1-1 | 40 | 37 | 93 |
| adr1-1 × adr1 | 65 | 8 | 12 |
| adr1-1 × adr1-1 | 51 | 0 | 0 |
| adr1-1 Δuka1-1 × adr1 uka1 | 95 | 2 | 2 |
| adr1-1 Δuka1-1 × adr1-1 Δuka1-1 | 95 | 0 | 0 |
| d132 | 63 | 32 | 51 |
| adr1-1/adr1 | 167 | 82 | 49 |
| adr1-1/adr1-1 | 179 | 0 | 0 |
| Δubc1-3/ubc1 | 22 | 12 | 55 |
| ubc1-2/ubc1-2 | 20 | 0 | 0 |
| Δubc1-3/ubc1-1 | 83 | 0 | 0 |
The data for each combination of mutant or diploid strains are pooled from two to three inoculation experiments. The data for heterozygous crosses are pooled from experiments in which the phenotype of the relevant mutations was tested in each of the two strain backgrounds (i.e., 518 and 521) when paired with a compatible wild-type strain. At least two independent transformants (Table 1) were used for each combination of mutant haploid strains or mutant diploid strains, except for the Δubc1-3/ubc1 diploid strain, where data are presented for only one transformant (UD12). Note that diploid strains such as d132 are known to be less virulent than mixtures of compatible haploid strains (10).
It should be pointed out that previous studies have not addressed the role of the cAMP pathway in a pathogenic cell type, as the reported pathogenicity data were based on co-inoculations of compatible U. maydis strains carrying mutations in the uac1 and/or the ubc1 gene (6, 8, 14). To investigate whether the lack of virulence observed in co-inoculations of adr1 mutants is caused solely by a mating defect or whether adr1 function is also required after cell fusion for pathogenic growth of the dikaryon, one or both copies of the adr1 gene were disrupted in the diploid U. maydis strain d132 (a1/a2 b1/b2). Diploid strains heterozygous at the b mating-type locus are filamentous on mating medium and pathogenic on corn plants; such strains obviate the requirement of cell fusion and dikaryon formation for virulence (10, 26). Diploid strains in which both adr1 genes were disrupted failed to induce tumors, whereas diploid strains heterozygous for the adr1 disruption were indistinguishable from d132 (Table 2). Not surprisingly, diploids lacking ubc1 function were not capable of inducing tumors in maize plants (Table 2) like the dikaryons lacking the regulatory subunit of PKA (8). We conclude that both the regulatory (Ubc1) and one catalytic (Adr1) subunit of PKA are required for pathogenic development in an infectious cell type. These results exclude mating defects in strains with a perturbed cAMP signaling pathway as the sole reason for their abolished or attenuated virulence.
Plant inoculations with mixtures of Δuka1-1 adr1-1 strains and compatible wild-type strains resulted in a reduced level of tumor formation (2%), compared with the corresponding heterozygous crosses of adr1 single mutants (12%; Table 2). Combined with the influence of uka1 on the filamentous growth of adr1 mutants, these results suggest that uka1 plays a minor role in PKA-mediated morphogenesis and virulence, as phenotypes of uka1 mutants become apparent only if the cAMP pathway is perturbed by inactivation of ubc1 or adr1.
Perturbations in cAMP signaling influence morphogenetic differentiation and pathogenic growth in other fungal phytopathogens (9). For example, mutations in the CPKA gene of M. grisea, encoding a catalytic subunit of PKA, were shown to delay appressorial differentiation and to attenuate appressorial penetration and, therefore, virulence (21, 27). In addition, disruption of cAMP signal transduction plays an important role in hypovirus-mediated attenuation of fungal virulence in the chestnut blight fungus Cryphonectria parasitica (28).
PKA and Mating in U. maydis.
The ability of adr1-1 mutant strains to mate and establish dikaryotic filaments could not be assessed with a standard plate mating assay because these strains form mycelial colonies as haploids. In contrast, the yeast-like growth of uka1 mutants allowed us to demonstrate that compatible Δuka1 strains behaved identically to wild-type strains in their ability to form aerial hyphae in a mating test (Fig. 4). Yeast-like growth also occurs for ubc1 mutants, and we have shown that mating-compatible ubc1 mutants fail to form aerial mycelium in the mating assay (6). Mutants defective in both uka1 and ubc1 were constructed to more closely examine whether deletion of the uka1 gene could suppress the defect of ubc1 mutants in formation of mating hyphae. Interestingly, weak mycelial growth was evident for mixtures of compatible uka1 ubc1 double mutants (Fig. 4), although the development of aerial hyphae was consistently reduced for these mutants compared with a wild-type mating response. This result is consistent with a contribution of Uka1 to total PKA activity. In contrast to partial suppression of the mating defect, deletion of the uka1 gene could not overcome the defect in tumor induction of ubc1 mutants (data not shown).
Figure 4.
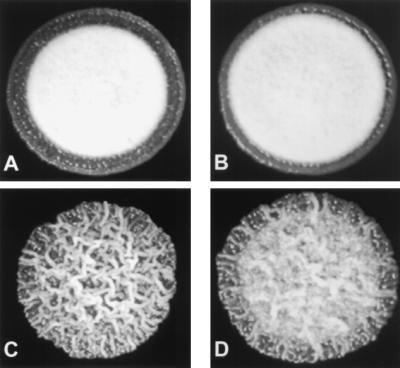
Mating reaction of Δuka1 Δubc1 double mutants. The following strains were co-spotted onto mating medium and incubated for 2 days: (A) wild-type strains, 518 (a2 b2) × 521 (a1 b1); (B) Δuka1 single mutants, 001#44(a2 b2 Δuka1-1) × 002#6 (a1 b1 Δuka1-1); (C) ubc1 single mutants, 0505 (a2 b2 Δubc1-4) × 521d (a1 b1 ubc1-2); (D) uka1 ubc1 double mutants, 001#9 (a2 b2 Δuka1-1 Δubc1-3) × 002#86 (a1 b1 Δuka1-1 Δubc1-3). The white mycelium indicative of the formation of dikaryotic hyphae is absent for the ubc1 single mutants; however, a weak mating reaction is observed with the Δuka1 Δubc1 double mutants.
The influence of uka1 on the mating reaction but not the virulence of ubc1 mutants also prompted a more detailed examination of the ability of ubc1 mutants to form aerial hyphae. Previous studies have shown that mating-type compatible ubc1-1 mutants are greatly attenuated for the formation of aerial hyphae in a mating test (6). Yet, if only one mating partner carries a mutation in the ubc1 gene, mating filaments are formed with the same efficiency as in a wild-type mating reaction, suggesting that mutations in ubc1 mainly affect post-cell fusion events. To directly assess the role of ubc1 in an infectious cell type, one or both copies of the ubc1 gene were replaced with deletion or disruption alleles in the pathogenic diploid strain d132. Fig. 5 demonstrates that the diploid strain UD1208, in which both copies of ubc1 were disrupted, is deficient in the formation of aerial mycelium compared with the wild-type diploid d132 and diploid UD12, the precursor of UD1208 in which only one copy of the ubc1 gene was mutated. The non-mycelial colonies of strain UD1208 are similar in appearance to those of strain d132-9; this diploid cannot form aerial hyphae because of a deletion at the b1 locus (a1/a2 b1∷hyg/b2). This experiment demonstrates that constitutively derepressed PKA activity interferes with mycelial growth in culture at an as yet unidentified step after cell fusion; certainly, this activity is able to override the b-derived input signal for filamentous growth, at least under the conditions of the classical mating assay.
Figure 5.

A functional ubc1 gene is required after cell fusion for mycelial growth. Strains were e1ither spotted alone (diploids) or co-spotted (haploids) on mating medium and incubated for 2 days at room temperature. The relevant genotypes of the strains are indicated in the margins.
The fission yeast S. pombe represents a useful fungal paradigm for the interconnection of mating and the cAMP signal transduction pathway. In this ascomycete, mutations that lead to low intracellular cAMP levels (e.g., cyr1−, encoding adenylyl cyclase; gpa2−, encoding a Gα subunit of heterotrimeric G protein) or abolish PKA activity (pka1−) bypass the requirement for nitrogen starvation for mating and sporulation and thus result in derepression of sexual development (23, 29). Similar connections between mating and cAMP may exist in U. maydis (Fig. 6). Our findings reveal that mutants defective in the cAMP pathway are attenuated for the mating reaction and for virulence. Further evidence for a connection comes from the analysis of the gpa3 gene that encodes a Gα subunit of heterotrimeric G proteins in U. maydis. This gene is required for pheromone signal transduction and for virulence; in addition, gpa3 mutants display an elongated cellular morphology similar to the partially filamentous phenotype of uac1 and adr1 mutants (30). The finding that addition of cAMP to the gpa3 mutant reverts the elongated morphology to wild-type cell shape (31) suggests that pheromone and cAMP signal transduction might share the same Gα subunit, Gpa3. Fig. 6 illustrates additional levels at which the mating (i.e., pheromone) and the cAMP signaling pathways might be interconnected. We note that direct interactions between pathways downstream of Gpa3 remain to be demonstrated at the molecular level, but it is interesting to speculate that the two pathways might converge on the same cellular targets.
Figure 6.

Two signal transduction pathways for mating, morphogenesis, and virulence in U. maydis. The pheromone response pathway on the left shows pheromone (Mfa) binding to a receptor (Pra), activation of a postulated MAP kinase module via a heterotrimeric G protein, and activation of the transcription factor Prf1. This factor increases transcription of the b genes encoding homeodomain transcription factors (bW1, bE1, bW2, bE2; ref. 34). The cAMP signal transduction pathway is shown on the right. The major PKA catalytic subunit Adr1 may phosphorylate potential target proteins or a proposed cAMP responsive transcription factor to influence target gene transcription. Dashed arrows indicate potential interconnections of the two pathways as described in Results and Discussion.
In S. pombe, the HMG domain transcription factor Ste11 is essential for sexual differentiation and pheromone response and has been shown to be negatively regulated by cAMP at the transcriptional level (32, 33). Prf1, the HMG domain transcription factor responsible for pheromone-stimulated induction of a and b mating-type genes in U. maydis, has recently been identified (34). It will therefore be interesting to see whether perturbed PKA activity in adr1 and ubc1 mutants affects the pheromone response in U. maydis. For example, it might be possible that Adr1 regulates Prf1 expression or activity.
The environmental signals feeding into the cAMP pathway and thereby modulating mating, morphogenesis, and pathogenic growth of U. maydis remain to be identified. Since pathogenic fungi have to switch from saprophytic to pathogenic growth once they colonize a host, it may not be surprising to find that signals derived from the host environment influence the cAMP pathway. In the case of U. maydis, these signals may include nutritional cues.
Acknowledgments
We thank Dr. Luc Giasson for providing primers PKA5 and PKA3 and Dave Laidlaw for technical help. This work was supported by a grant from the Medical Research Council of Canada (to J.W.K.) and by a fellowship from the Swiss National Fund (to F.D.).
ABBREVIATION
- PKA
cAMP-dependent protein kinase
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. AF025290).
References
- 1.Banuett F. Annu Rev Genet. 1995;29:179–208. doi: 10.1146/annurev.ge.29.120195.001143. [DOI] [PubMed] [Google Scholar]
- 2.Kronstad J W, Staben C. Annu Rev Genet. 1997;31:245–276. doi: 10.1146/annurev.genet.31.1.245. [DOI] [PubMed] [Google Scholar]
- 3.Bölker M, Urban M, Kahmann R. Cell. 1992;68:441–450. doi: 10.1016/0092-8674(92)90182-c. [DOI] [PubMed] [Google Scholar]
- 4.Kahmann R, Bölker M. Cell. 1996;85:145–148. doi: 10.1016/s0092-8674(00)81091-0. [DOI] [PubMed] [Google Scholar]
- 5.Kernkamp M F. Phytopathology. 1941;32:554–567. [Google Scholar]
- 6.Gold S, Duncan G, Barrett K, Kronstad J. Genes Dev. 1994;8:2805–2816. doi: 10.1101/gad.8.23.2805. [DOI] [PubMed] [Google Scholar]
- 7.Ruiz-Herrera J, Leon C G, Guevara-Olvera L, Carabez-Trejo A. Microbiology. 1995;141:695–703. [Google Scholar]
- 8.Gold S E, Brogdon S M, Mayorga M E, Kronstad J W. Plant Cell. 1997;9:1585–1594. doi: 10.1105/tpc.9.9.1585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kronstad J W. Trends Plant Sci. 1997;2:193–199. [Google Scholar]
- 10.Kronstad J W, Leong S A. Proc Natl Acad Sci USA. 1989;86:978–982. doi: 10.1073/pnas.86.3.978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 12.Hanks S K, Quinn A M, Hunter T. Science. 1988;241:42–52. doi: 10.1126/science.3291115. [DOI] [PubMed] [Google Scholar]
- 13.Orth A B, Rzhetskaya M, Pell E J, Tien M. Appl Environ Microbiol. 1995;61:2341–2345. doi: 10.1128/aem.61.6.2341-2345.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Barrett K J, Gold S E, Kronstad J W. Mol Plant-Microbe Interact. 1993;6:274–283. doi: 10.1094/mpmi-6-274. [DOI] [PubMed] [Google Scholar]
- 15.Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- 16.Wang J, Holden D W, Leong S A. Proc Natl Acad Sci USA. 1988;85:865–869. doi: 10.1073/pnas.85.3.865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ballance D J. In: Molecular Industrial Mycology: Systems and Applications for Filamentous Fungi. Leong S A, Berka R M, editors. New York: Dekker; 1991. pp. 1–29. [Google Scholar]
- 18.Knighton D R, Zheng J, Ten Eyck L F, Xuong N, Taylor S S, Sowadski J M. Science. 1991;253:414–420. doi: 10.1126/science.1862343. [DOI] [PubMed] [Google Scholar]
- 19.Orellana S A, McKnight G S. Proc Natl Acad Sci USA. 1992;89:4726–4730. doi: 10.1073/pnas.89.10.4726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.de Oliveira J C, Borges A C, do Valle Marques M, Gomes S L. Eur J Biochem. 1994;219:555–562. doi: 10.1111/j.1432-1033.1994.tb19971.x. [DOI] [PubMed] [Google Scholar]
- 21.Mitchell T K, Dean R A. Plant Cell. 1995;7:1869–1878. doi: 10.1105/tpc.7.11.1869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Toda T, Cameron S, Sass P, Zoller M, Wigler M. Cell. 1987;50:277–287. doi: 10.1016/0092-8674(87)90223-6. [DOI] [PubMed] [Google Scholar]
- 23.Maeda T, Watanabe Y, Kunitomo H, Yamamoto M. J Biol Chem. 1994;269:9632–9637. [PubMed] [Google Scholar]
- 24.Uhler M D, Charmichael D F, Lee D C, Chrivia J C, Krebs E G, McKnight G S. Proc Natl Acad Sci USA. 1986;83:1300–1304. doi: 10.1073/pnas.83.5.1300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Thompson J D, Higgins D G, Gibson T J. Nucleic Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Holliday R. In: Handbook of Genetics. King R C, editor. Vol. 1. New York: Plenum; 1974. pp. 575–595. [Google Scholar]
- 27.Xu J R, Urban M, Sweigard J A, Hamer J E. Mol Plant-Microbe Interact. 1997;10:187–194. [Google Scholar]
- 28.Nuss D L. Plant Cell. 1996;8:1845–1853. doi: 10.1105/tpc.8.10.1845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Isshiki T, Mochizuki N, Maeda T, Yamamoto M. Genes Dev. 1992;6:2455–2462. doi: 10.1101/gad.6.12b.2455. [DOI] [PubMed] [Google Scholar]
- 30.Regenfelder E, Spellig T, Hartmann A, Lauenstein S, Bölker M, Kahmann R. EMBO J. 1997;16:1934–1942. doi: 10.1093/emboj/16.8.1934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kahmann R, Basse C. Trends Plant Sci. 1997;2:366–367. [Google Scholar]
- 32.Aono T, Yanai H, Miki F, Davey J, Shimoda C. Yeast. 1994;10:757–770. doi: 10.1002/yea.320100607. [DOI] [PubMed] [Google Scholar]
- 33.Sugimoto A, Iino Y, Maeda T, Watanabe Y, Yamamoto M. Genes Dev. 1991;5:1990–1999. doi: 10.1101/gad.5.11.1990. [DOI] [PubMed] [Google Scholar]
- 34.Hartmann H A, Kahmann R, Bölker M. EMBO J. 1996;15:1632–1641. [PMC free article] [PubMed] [Google Scholar]