

Abstract
We have demonstrated chromosomal instability in the clonal descendants of hemopoietic stem cells after irradiating murine bone marrow with α-particles. However, because cells that are irradiated by α-particles are defined by a Poisson distribution of individual particle traversals, there is an inevitable proportion of unirradiated cells in the surviving population. The calculated expected proportions of irradiated and nonirradiated cells indicate that the number of clonogenic cells transmitting chromosomal instability is greater than the number expected to be hit and survive. To investigate further this discrepancy, we studied the effects of interposing a grid between the cells and the α-particle source so that the surviving population consists predominantly of untraversed stem cells. Comparison with the same irradiation conditions without the grid reveals that the same level of instability is induced. The data confirm that α-particles induce chromosomal instability but instability is demonstrated in the progeny of nonirradiated stem cells and must be due to unexpected interactions between irradiated and nonirradiated cells. This untargeted effect has important implications for mechanistic studies of radiation action and for assessment of radiation risk.
Environmental exposures to the α-particles produced by radon and its decay products are responsible for approximately 50% of the average annual effective dose from natural sources of ionizing radiation worldwide (1). In some situations artificial radionuclides, such as plutonium 239 or Americium 241 associated with the nuclear industry, make some small additional contribution. An important feature of α-irradiation is that, no matter how low the total dose to the whole body, a substantial dose of radiation (∼0.5 Gy) is delivered to an individual cell if it is traversed by a single α-particle (2) and it is conventionally assumed that cells that are not traversed are unaffected by the radiation. We are able to irradiate cells with α-particles in the laboratory, and because our conditions of dose, particle fluence and linear energy transfer are precisely defined (3, 4), we can calculate the mean number of α-particles per target cell (5). By using a clonogenic assay for hemopoietic stem cells (6), operationally defined as colony-forming unit type A (CFU-A), we are able to investigate the effects of environmentally relevant doses of α-particles. Previously, we have demonstrated that doses corresponding to a mean of approximately one α-particle per cell result in chromosomal instability in the descendants of hemopoietic stem cells (7, 8). Inevitably at these low doses, some cells, by chance, will not have been irradiated (4) and the data are consistent with instability being expresssed in the progeny of more clonogenic cells than were traversed by an α-particle. We have investigated further this apparent disrepancy and have demonstrated by direct experimental investigation that, after α-particle irradiation, chromosomal instability is demonstrated in the descendants of unirradiated stem cells.
MATERIALS AND METHODS
Bone marrow cells from male mice were irradiated with α-particles by using a versatile source containing a 20-mm-diameter disc of plutonium 238 as described (3). Immediately after irradiation the cells were washed and resuspended, and the CFU-A assay (6) was used to obtain clones of cells derived from members of the hemopoietic stem cell compartment. Cells were plated in 45-mm Petri dishes containing 2 ml of modified α Eagle’s medium supplemented with 25% pretested horse serum, 0.3% low-melting-point agarose, antibiotics, and sources of colony-stimulating activities as described (6). Coded cytogenetic preparations were obtained by a method for karyotyping hemopoietic colonies (7, 8). Briefly, metaphases in developing (day 7–9) colonies containing approximately 104 cells, that is some 13 cell divisions from initiation of clonal proliferation, were arrested by adding Colcemid at 0.02 μg/ml to the dishes. Individual colonies were transferred in 10-μl droplets of 0.5% KCl onto poly-(l-lysine)-coated microscope slides, and hypotonic treatment of the cells was achieved by inverting each slide to prevent attachment and to allow the cells to swell in a hanging droplet. After 25 min in a humidified incubator at 37°C, the slide was turned upright, and cells were allowed to attach to the coated surface of the slide. Fixed cells were spread on slides, air-dried, aged for 1 week at room temperature, and stained with Giemsa, and aberrations were classified as described (7, 8).
To obtain a situation where most clonogenic cells were unirradiated but in the vicinity of cells that were traversed by α-particles at the time of irradiation, bone marrow cells from male mice were irradiated with α-particles with and without a grid interposed between source and sample. In the latter case, 1-Gy absorbed dose was delivered to the unshielded cells only. The grid is composed of interwoven brass wires 0.34 mm in diameter with a rectangular pitch of 1.2 mm and was placed immediately below, but not touching, the dish base. The source was used at its standard distance with a reducing aperture of 4.5 mm in diameter and a source to sample distance of 68.5 mm giving an absorbed dose rate of 0.021 Gy/s and linear energy transfer of 121 keV/μm at the sample position. The sample was composed of 5 μl of a single cell suspension (2 × 108 cells per ml) sandwiched between the dish base and a 28.5-mm-diameter CR39 plastic disk (4). Subsequent track-etching of the CR39 plastic confirmed full transmission of the sample by the α-particle beam. Transmission measurements from the etched CR39 plastic immediately above the cell layer and from measurements with travelling microscope on the grid itself agree with each other to within 3%, showing that penumbra effects of the grid were negligible. Confocal microscopic observations of cells in the hostaphan dishes confirmed negligible cell movement (<2% of cells) during the 47.6 s required to deliver 1 Gy of α-particles.
Immediately after irradiation the cells were washed and resuspended, and the CFU-A assay was used to obtain clones of cells derived from members of the hemopoietic stem cell compartment (6). Some of these were used to determine clonogenic survival and others were used for cytogenetic analysis to demonstrate chromosomal instability by the presence of nonclonal aberrations.
RESULTS
Theoretical Considerations.
The clonogenic cells detected by the CFU-A assay have diameters in the range of 7.0–7.8 μm with an average diameter of 7.2 μm (4, 9). Using Poisson statistics, the probability P(n) of cells being traversed by n = 0, 1, 2, 3, etc., α-particles can be calculated from the particle fluence (5). From these values and the coefficient of the exponential survival curve, the proportions of cells in the surviving population that are irradiated Ps(>0) and not irradiated Ps(=0) can be calculated as
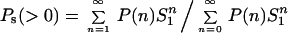 |
and
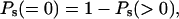 |
respectively, where S1, is the probability of a cell surviving the traversal of a single α-particle, evaluated from the slope of the survival curve as described (4).
If it is argued that the maximum expected proportion of stem cells exhibiting instability is equal to the proportion of surviving stem cells traversed by one or more α-particles, then the proportion of colonies actually showing instability not only fail to reflect the expected dose–response relationship but are well in excess of the expected values (Table 1). All the expected values fall outside the 60% binomial confidence limits of the experimental data and outside the 95% binomial confidence limits for the combined data. The probabilities of obtaining the observed results if the expected values represent the true values have been calculated by using an exact binomial test (10) and there is significant discrepancy.
Table 1.
Significant discrepancy in the observed and expected frequencies of haemopoietic colonies exhibiting chromosomal instability defined by the incidence of nonclonal cytogenetic aberrations (7, 8)
| α-Particle irradiation dose, Gy | Colonies exhibiting instability, n/n
|
Exact binomial test probability | |
|---|---|---|---|
| Observed | Expected Ps(>0) | ||
| 0.25 | 2/5 = 0.400* | 0.055 | 2.7 × 10−3 |
| 0.5 | 6/12 = 0.500* | 0.105 | 7.1 × 10−4 |
| 0.5 | 16/26 = 0.615† | 0.105 | 4.1 × 10−10 |
| 0.5 combined data | 22/38 = 0.579 | 0.105 | 1.2 × 10−12 |
| 1.0 | 4/10 = 0.400* | 0.200 | 0.12 |
| 1.0 | 41/64 = 0.640‡ | 0.200 | 1.5 × 10−13 |
| 1.0 combined data | 45/74 = 0.608 | 0.200 | 1.2 × 10−13 |
Experimental Studies.
To address experimentally the discrepancy in our observed and expected values, we manipulated the experimental conditions by interposing a shielding grid between the source of α-particles and the dishes in which the cells are irradiated (Fig. 1) and used a dose of 1 Gy to the exposed areas so that the majority of CFU-A in these areas are inactivated (4). In the absence of the grid, 20% of surviving CFU-A would be expected to have seen one or more α-particles; interposing the grid reduces the proportion of traversed clonogenic cells to a maximum of 3%. Bone marrow cells were irradiated under these conditions, and the percent transmission through the grid was measured as 0.51% by direct microscopic measurements of the grid dimensions and confirmed by track-etching the CR39 disc that lies above the cells. Survival data were obtained for CFU-A with and without the grid (Fig. 2) and linear quadratic and linear survival curves were fitted to the data by the method of maximum likelihood. The fraction (f) of the population exposed through the grid was calculated on the assumption of a constant proportion (1 − f) of shielded cells. A Poisson distribution of surviving cells was confirmed by likelihood-ratio dispersion tests and a comparison of the fits of the two models revealed no significant evidence for a quadratic component in the survival curves. From four experiments, the fraction f was calculated from the survival data to be 0.568 ± 0.056 and this does not differ significantly from the measured 0.51. Hence, the reduction in the number of exposed clonogenic cells shown in Fig. 2 is consistent with the reduction in exposed area due to the grid and consistent with the negligible movement of cells confirmed by confocal microscopy. These data demonstrate that the expected number of clonogenic cells were traversed by α-particles and that there is no involvement of indirect effects in cell killing.
Figure 1.

Schematic diagram of the α-particle irradiator indicating the shielding effect of interposing the grid between source and cells.
Figure 2.

Reduction in clonogenic survival of CFU-A is consistent with the reduction in the exposed area due to the grid. The mouse bone marrow cells were irradiated with α-particles either with or without an interposed grid between the source and the cells as shown in Fig. 1. The specified doses refer to the unshielded areas.
CFU-A-derived colonies from the two irradiation conditions were analyzed for chromosomal instability by determining the incidence of nonclonal cytogenetic aberrations (7). As expected there is a background incidence of chromosome breakage with a mean of 0.07 aberration per cell (Table 2), but it is is evident that breakage is very significantly increased in the colonies derived from clonogenic cells exposed to α-particles under both irradiation conditions (0.21 and 0.21 aberration per cell with and without the grid, respectively). Within each of the three treatment groups, the individual distributions were tested for homogeneity by using Pearson χ2 tests. Satisfactory homogeneity allowed comparison between treatment groups to be performed on the overall distributions of aberrations between metaphases, again using Pearson χ2 tests. Significant instability is expressed after 1 Gy of α-particles (Table 2, P = 1.5 × 10−6 compared with controls) and also after irradiation with the grid (P = 3.4 × 10−7), there being no significant difference between treatments with and without the grid (P = 0.63). Clonal aberrations were not seen in any of the colonies, but because most α-particle-induced aberrations are complex and unlikely to be transmissible over many cell divisions, this is not unexpected given the number of colonies studied. Consistent with previous observations, frequency of induction of instability is considerably greater than the frequency of induction of mutations at specific loci and is characterized by a high ratio of chromatid-type aberrations to chromosome-type aberrations (Table 2).
Table 2.
Significant discrepancy in the observed and expected frequencies of haemopoietic colonies exhibiting chromosomal instability with and without the grid interposed between the α-particle source and the cells (in the latter case, 1 Gy of absorbed dose is delivered to the unshielded cells only)
| α-Particle irradiation dose, Gy | No. colonies with aberrant cells/total no. | No. aberrant cells/total no. | No. chromatid breaks | No. chromatid exchanges and minutes | No. chromosome fragments | No. ring chromosomes | No. translocations | Mean no. aberrations per cell |
|---|---|---|---|---|---|---|---|---|
| Control | 22 /56 | 36 /662 | 36 | 0 | 12 | 0 | 0 | 0.07 |
| 1 Gy | 41 /64 | 137 /1009 | 191 | 4 | 24 | 3 | 0 | 0.22 |
| 1 Gy with grid | 41 /63 | 115 /871 | 128 | 15 | 29 | 5 | 6 | 0.21 |
P = 1.5 × 10−6 (control vs. 1 Gy); P = 3.4 × 10−7 (control vs. 1 Gy + grid); P = 0.63 (1 Gy vs. 1 Gy + grid).
DISCUSSION
Our experimental studies have confirmed the implications from our theoretical considerations that, after α-particle irradiation, chromosomal instability is demonstrated in the descendants of unirradiated stem cells. This has been demonstrated by the interposing of the grid between the α-particle source producing the expected reduction in the number of exposed clonogenic cells yet not reducing the number of colonies expressing chromosomal instability. Interestingly, there have been other reports indicative of biological effects not being restricted to those nuclei actually traversed by an α-particle (11–13) but detected in “extranuclear” targets. In our study, because our experimental conditions of dose, particle fluence, and linear energy transfer are precisely defined, our data prove unequivocally that more cells (nuclei plus cytoplasm and not just nuclei) than would be expected to be traversed by an α-particle are affected. Indeed, it is evident that biological effects are demonstrated in the progeny of an essentially unirradiated population of clonogenic cells.
There is growing evidence to support the view that radiation can induce genomic instability in a number of cell types that may result in mutational effects arising in the progeny of the irradiated cells. These effects include increased frequency of specific gene mutations (14–16) and chromosome aberrations (17–26), and delayed reproductive death or “lethal mutations” (27, 28) may also reflect induced genomic instability. The mechanisms underlying these induced instabilities are not understood, and our finding of instability in the progeny of unirradiated cells adds further complexity. Previously, in cultures where we have demonstrated induced genomic instability, we have also demonstrated increases in intracellular oxidants, oxidative DNA base damage, vulnerability to free radical-mediated membrane damage, and levels of O-tetradecanoylphorbol 13-acetate-linked superoxide production (27). These findings are consistent with aberrant oxy-radical metabolism contributing, at least in part, to the expression of instability in hemopoietic cells, and the biochemical basis of these changes and the factors involved in the transfer of the instability phenotype to nonirradiated cells should be investigated. Whatever the details of the mechanism, the present study demonstrates that the α-particle-induced instability in primary hemopoietic cells must be attributed to unexpected interactions between the irradiated and nonirradiated cells.
The health effects of environmental exposures to α-emitting radionuclides are not well established. Estimation of the consequences for humans is greatly complicated by the nonuniformity of the distribution of the radionuclides in the tissues, the short ranges of α-particles, and the consequent uncertainty about the dose to any target cell. However, α-emitting radionuclides in certain environments may be responsible for a significant proportion of the dose to specific organs of the body, particularly the lung but also the bone marrow (28–30). The potential mechanisms for α-radiation-induced malignant transformation have usually considered only lesions in “hit” cells. Cells actually hit by, and surviving, the traversal of an α-particle are a minority population in any tissue exposed to environmental levels of α-emitting radionuclides. However, our data clearly demonstrate the potential for interaction between irradiated and nonirradiated cells in the production of genetic damage. That the progeny of unirradiated stem cells can demonstrate an unstable phenotype raises important questions concerning the mechanisms underlying α-particle carcinogenesis and has significant implications for risk estimation based on the conventionally considered target cells.
ABBREVIATION
- CFU-A
colony-forming unit type A
References
- 1.United Nations Scientific Committee on the Effects of Atomic Radiation. Report to the General Assembly. New York: United Nations; 1993. [Google Scholar]
- 2.Goodhead D T. Adv Radiat Biol. 1992;16:7–44. [Google Scholar]
- 3.Goodhead D T, Bance D A, Stretch A, Wilkinson R E. Int J Radiat Biol. 1991;59:195–210. doi: 10.1080/09553009114550181. [DOI] [PubMed] [Google Scholar]
- 4.Lorimore S A, Goodhead D T, Wright E G. Int J Radiat Biol. 1993;63:655–660. doi: 10.1080/09553009314450851. [DOI] [PubMed] [Google Scholar]
- 5.Goodhead D T, Munson R J, Thacker J T, Cox R. Int J Radiat Biol. 1980;37:135–167. doi: 10.1080/09553008014550201. [DOI] [PubMed] [Google Scholar]
- 6.Lorimore S A, Pragnell I B, Eckman L N, Wright E G. Leuk Res. 1990;14:481–489. doi: 10.1016/0145-2126(90)90036-9. [DOI] [PubMed] [Google Scholar]
- 7.Kadhim M A, Macdonald D A, Goodhead D T, Lorimore S A, Marsden S J, Wright E G. Nature (London) 1992;355:738–740. doi: 10.1038/355738a0. [DOI] [PubMed] [Google Scholar]
- 8.Watson G E, Lorimore S A, Clutton S M, Kadhim M A, Wright E G. Int J Radiat Biol. 1997;71:497–503. doi: 10.1080/095530097143824. [DOI] [PubMed] [Google Scholar]
- 9.Visser J W M, Van Bekkum D W. Exp Hematol. 1990;18:248–256. [PubMed] [Google Scholar]
- 10.Conover W J. Practical Non-parametric Statistics. 3rd Ed. New York: Wiley; 1980. [Google Scholar]
- 11.Nagasawa H, Little J B. Cancer Res. 1992;52:6394–6396. [PubMed] [Google Scholar]
- 12.Deshpande A, Goodwin E H, Bailey S M, Marrone B L, Lehnert B E. Radiat Res. 1996;145:260–267. [PubMed] [Google Scholar]
- 13.Hickman A W, Jaramillo R J, Lechner J F, Johnson N F. Cancer Res. 1994;54:5797–5800. [PubMed] [Google Scholar]
- 14.Little J B, Gorgojo L, Vetros H. Int J Radiat Oncol, Biol Phys. 1990;19:1425–1429. doi: 10.1016/0360-3016(90)90354-m. [DOI] [PubMed] [Google Scholar]
- 15.Chang W P, Little J B. Mutat Res. 1992;270:191–199. doi: 10.1016/0027-5107(92)90130-t. [DOI] [PubMed] [Google Scholar]
- 16.Harper K, Lorimore S A, Wright E G. Exp Hematol. 1997;25:263–269. [PubMed] [Google Scholar]
- 17.Pampfer S, Streffer C. Int J Radiat Biol. 1989;55:85–92. doi: 10.1080/09553008914550091. [DOI] [PubMed] [Google Scholar]
- 18.Martins M B, Sabatier L, Ricoul M, Pinton A, Dutrillaux B. Mutat Res. 1993;285:229–237. doi: 10.1016/0027-5107(93)90111-r. [DOI] [PubMed] [Google Scholar]
- 19.Holmberg K, Falt S, Johansson A, Lambert B. Mutat Res. 1993;286:321–330. doi: 10.1016/0027-5107(93)90197-n. [DOI] [PubMed] [Google Scholar]
- 20.Marder B A, Morgan W F. Mol Cell Biol. 1993;13:6667–6677. doi: 10.1128/mcb.13.11.6667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kadhim M A, Walker C A, Plumb M A, Wright E G. Int J Radiat Biol. 1996;69:167–174. doi: 10.1080/095530096145995. [DOI] [PubMed] [Google Scholar]
- 22.Durante M, Grossi G F, Yang T C. Adv Space Res. 1996;18:99–108. doi: 10.1016/0273-1177(95)00796-h. [DOI] [PubMed] [Google Scholar]
- 23.Ponnaiya B, Cornforth M N, Ullrich R A. Radiat Res. 1997;147:121–125. [PubMed] [Google Scholar]
- 24.Ponnaiya B, Cornforth M N, Ullrich R A. Radiat Res. 1997;147:288–294. [PubMed] [Google Scholar]
- 25.Seymour C B, Mothersill C, Alper T. Radiat Biol. 1986;50:167–179. doi: 10.1080/09553008614550541. [DOI] [PubMed] [Google Scholar]
- 26.Gorgojo L, Little J B. Int J Radiat Biol. 1989;55:619–630. doi: 10.1080/09553008914550661. [DOI] [PubMed] [Google Scholar]
- 27.Clutton S M, Townsend K M S, Walker C, Ansell J D, Wright E G. Carcinogenesis. 1996;17:1633–1639. doi: 10.1093/carcin/17.8.1633. [DOI] [PubMed] [Google Scholar]
- 28.Committee on Medical Aspects of Radiation in the Environment (1984) The Incidence of Cancer and Leukaemia in Young People in the Vicinity of the Sellafield Site, West Cumbria, chairman, Bridges, B. A. (Her Majesty’s Stationery Office, London), 4th Report.
- 29.Dionian J, Muirhead C R, Wan S L, Wrixon A D. Report NRPB-R196. Didcot, U.K.: National Radiological Protection Board; 1986. [Google Scholar]
- 30.Richardson R B, Eatough J P, Henshaw D L. Br J Radiol. 1991;64:608–624. doi: 10.1259/0007-1285-64-763-608. [DOI] [PubMed] [Google Scholar]