

Abstract
In the normal eye, the complement system is continuously activated at low levels and both membrane-bound and soluble intraocular complement regulatory proteins tightly regulate this spontaneous complement activation. This allows protection against pathogens without causing any damage to self-tissue and vision loss. The complement system and complement regulatory proteins control the intraocular inflammation in autoimmune uveitis and play an important role in the development of corneal inflammation, age-related macular degeneration and diabetic retinopathy. The evidence derived from both animal models and patient studies support the concept that complement inhibition is a relevant therapeutic target in the treatment of various ocular diseases. Currently, several clinical trials using complement inhibitors are going on. It is possible that, in the near future, complement inhibitors might be used as therapeutic agents in eye clinics.
Keywords: Complement, Eye, corneal inflammation, Uveitis, Age related macular degeneration, Diabetic retinopathy
1. Introduction
The complement system is a key component of innate immunity. It consists of a large group of plasma and membrane bound proteins that play a central role in the defense against infection and in the modulation of immune and inflammatory responses (Atkinson and Frank, 1998; Ross, 1986; Muller-Eberhard, 1988; Frank and Fries, 1991). The complement system can be activated via three distinct pathways namely, the classical, the alternative and the lectin pathways and complement activation triggers a sequence of biological reactions (Atkinson and Frank, 1998; Ross, 1986; Muller-Eberhard, 1988; Frank and Fries, 1991; Reid, et al., 1986; Morgan and Harris, 1999; Thiel, et al., 1997). The classical pathway can be activated by immune complexes or by substances such as C-reactive protein, and the complement components involved include C1, C2, C4 and C3 (Muller-Eberhard, 1988; Frank and Fries, 1991). The alternative pathway provides a rapid, antibody-independent route of complement activation and amplification. The alternative pathway directly activates C3 when it interacts with certain activating surfaces (e.g. zymosan, lipopolysaccharides) and involves C3, Factor B, Factor D and properdin (Reid, et al., 1986; Morgan and Harris, 1999). The activation of the lectin pathway is also independent of immune complex generation and can be achieved by interaction of certain serum lectins, such as mannose binding lectin (MBL), with mannose and N-acetyl glucosamine residues present in abundance in bacterial cell walls (Thiel, et al., 1997).
Activated complement is a double-edged sword that not only helps defend the host against pathogens, but also has the potential to inflict damage to self-tissues (Liszewski, et al., 1996; Atkinson, 1987). Thus, it is critical for the body to maintain a balance between complement activation and complement inhibition (Atkinson, 1987). To protect the host from destructive effects of complement-mediated damage, complement activation is tightly regulated by the complement regulatory proteins - CRegs (Morgan and Harris, 1999). CRegs can be categorized into two classes-membrane bound and soluble proteins. Decay Accelerating Factor (DAF, CD55), Membrane Cofactor Protein (MCP, CD46), Complement Receptor 1 (CR1, CD35), and Membrane Inhibitor of Reactive Lysis (MIRL, CD59) are important membrane bound CRegs. DAF regulates the activation of C3 and C5 by preventing the formation of C3 and C5 convertases and by accelerating the decay of these convertases (Lublin, et al., 1987; Nicholson-Weller 1992). MCP down regulates the complement cascade by acting as a cofactor for factor I-mediated cleavage of C3b (Liszewski, et al., 1991, 1992, 1996). CR1 has both DAF and MCP functions (Krych-Goldberg and Atkinson, 2001). CD59 regulates the assembly and function of membrane attack complex - MAC (Holguin, et al., 1992; Watts, et al., 1990). Crry (5I2 antigen) is a membrane bound CReg that is present in rodents only and has both decay accelerating and cofactor activities. It controls complement activation at the critical step of C3 convertase formation (Kim et al., 1995a; Takizawa, et al., 1994; Molina, et al., 1992). Tissue distribution studies have shown that both Crry and CD59 are widely distributed proteins in rodents (Li, et al., 1993; Powell, et al., 1997; Qian, et al., 2000). DAF and MCP have also been identified in rodents (Spicer, et al., 1995; Miwa, et al., 1998). C1 inhibitor (C1INH), C4 binding protein (C4bp), complement factor H (CFH), and complement factor I (CFI), are some of the important soluble CRegs. C1INH regulates C1 whereas C4bp catalyses the cleavage of C4b by CFI. CFH acts as a cofactor for CFI-mediated cleavage of C3b and also has decay accelerating activity against the alternative pathway C3 convertase, C3bBb. Soluble forms of MCP, DAF, CR1, CD59 and Crry have been reported to be present in various biological fluids. These soluble forms may be the products resulting from proteolysis, alternative splicing of mRNA, or post-translational modification (Wheeler et al., 2002., Nonaka 1995., Nickells et al., 1994).
2. Complement and ocular protection
Several reports in the literature have suggested the presence of functionally active classical and alternative complement pathways in the cornea, aqueous humor, tears and retina (Mondino and Brady., 1981; Mondino et al., 1983; Bora et al., 1993; Willcox et al., 1997). Various proteins which regulate the activation of the complement system, such as C1 inhibitor, DAF, MCP, CD59, factor I and factor H, have been reported to be present in various ocular tissues, tears, aqueous and vitreous humor (Willcox et al., 1997; Bora et al., 1993). We have reported that a functionally active complement system is present in normal rodent eye (Sohn et al., 2000a). It was further demonstrated in this report that the complement system is continuously active at a low level in the normal rodent eye, and intraocular CRegs present in the intraocular fluid as well as on the cell membrane tightly regulate this spontaneous complement activation (Sohn et al., 2000a, 2000b). Control of complement activation at the level of C3 convertase was sufficient to prevent complement-mediated intraocular inflammation (Sohn et al., 2000a).
3. Complement and ocular diseases
The presence and activation of complement has been suggested to play a crucial role in the pathogenesis of a large number of diseases, including ocular diseases (Thurman et al., 2006).
3.1 Complement and corneal disease
The ocular surface that consists of the cornea and the conjunctiva, is constantly exposed to the external environment, therefore is in contact with a variety of pathogenic microorganisms and inflammatory antigens. The complement system plays an important role in protection of the cornea from these insults. Components of both the classical and alternative pathways of complement (C1, C4, C2, C3, C5, C6, C7, properdin and factor B) are present in normal cornea. It has been suggested that the cornea has the capability to activate the complement cascades in response to bacterial infection (Mondino and Sumner, 1990; Mondino et al., 1996). Cleveland and associates demonstrated that depletion of C3 by cobra venom factor (CVF) renders the normally resistant DBA/2 mice susceptible to corneal infection with Pseudomonas aeruginosa (Cleveland et al 1983; Hazlett et al., 1984). However C5 deficient mice are still resistant to corneal P. aeruginosa infection indicating that the complete lytic pathway of complement is not essential for host resistance towards this gram-negative bacterium. These findings suggest that the functions associated with C3 such as opsonization and regulation of phagocytosis, may be critical in protection of the cornea from bacterial infection (Cleveland et al 1983; Hazlett et al., 1984).
Although the complement system is critical for the protection of the cornea from infection, spontaneous complement activation can cause damage to the corneal tissue after the infection is cleared. To protect from this complement-mediated damage, the cornea expresses membrane bound CRegs such as MCP, DAF, Crry and CD59 (Bora et al., 1993; Bardenstein et al., 1994; Sohn et al., 2000a). These CRegs are heavily expressed in the corneal epithelium at the limbus, as well as in the central cornea. High expression of CRegs is crucial for the protection of cornea because the cornea is constantly being challenged by a variety of substances, including infectious organisms that produce phospholipase and other enzymes, which can remove CRegs from ocular cell surface (Cocuzzi et al., 2000). This bacterially induced loss of CRegs on the cornea could lead to the damage of ocular tissue by autologous complement activation during the course of complement attack on pathogens.
3.2 Complement and autoimmune uveitis
Uveitis is broadly defined as inflammation of the uvea (comprising choroids, iris and ciliary body), and is responsible for almost 3% of blindness in the United States. Each year, 17.6% of active uveitis patients experience a transient or permanent loss of vision. The study of uveitis is complicated by the fact that it encompasses a wide range of underlying etiologies. It may be idiopathic, associated with systemic diseases, or resulting from a variety of infectious agents. Anatomically, uveitis is classified as anterior (iritis, iridocyclitis), intermediate, posterior (vitritis, retinitis, choroiditis) or pan. Anterior uveitis (AU) is the most common form of uveitis and accounts for approximately 75% of cases. The most common form of anterior uveitis is of unknown (i.e. idiopathic) etiology (Bora et al., 2007a). In a non-referral clinic, 52% of patients may present with idiopathic AU.
Complement activation products such as C3b and C4b have been demonstrated to be present in the eyes of patients with AU (Mondino et al., 1984; 1986). Recently, we have shown that the presence and activation of complement is central to the development of experimental autoimmune AU (EAAU) (Jha et al., 2006). EAAU is an autoimmune disease of the eye, which serves as an animal model of idiopathic human AU (Broekhuyse, et al., 1991; Bora et al., 1995; Bora et al., 1997; Simpson et al., 1997; Kim et al., 1995b; Woon 1998; Bora et al., 2004). EAAU is induced in Lewis rats by an antigen specific CD4+ T cell response to an antigen derived from the iris and ciliary body (Bora et al., 1995, 1997). We demonstrated that the presence and activation of complement is critical for the development of EAAU induced by either active immunizations or the transfer of in vitro primed antigen-specific CD4+ T cells (Jha et al., 2006a). These results suggested that complement plays an important role in the induction of antigen specific T-cell responses in AU. A central role of complement in the immunopathogenesis of EAAU was further supported by several observations, such as decreased production of IFN-γ, IL-10, IP-10, ICAM-1 and LECAM-1 in complement-depleted animals during the course of EAAU. Levels of iC3b, a cleavage product of C3, increased within the eye during the peak of EAAU. Since activation of the complement system is necessary for the generation of C3 split products (Thomilson 1993), their increased levels within the eye of rats with EAAU provided indisputable evidence of local complement activation during autoimmune intraocular inflammation. The number of CR3 expressing cells also increased during the peak of the disease. The systemic injection of anti-CR3, which specifically blocks the interaction between iC3b and its receptor CR3, had a significant protective effect on EAAU thus suggesting that the interaction of iC3b with CR3 was involved in the generation and progression of uveitis in EAAU (Jha et al., 2006a).
The eye is under threat from homologous complement attack during EAAU, and unregulated complement activation can be detrimental to ocular structures that are vital for vision. Recently, we have demonstrated that various ocular tissues up-regulate Crry and CD59 during EAAU, to avoid self-injury from complement-mediated tissue damage and down-regulate intraocular inflammation (Jha et al., 2006b). It was observed that the suppression of function and/or expression of these CRegs by neutralizing monoclonal antibodies, anti-sense oligonucleotides, or small interfering RNA (siRNA) resulted in more severe disease with early onset and delayed resolution. The eyes from theses animals stained very strongly for C3, C3 activation products, and MAC compared with control animals (Jha et al., 2006b). There was no apparent histologic damage to ocular tissue after the inflammation cleared in these animals. These results demonstrated that the suppression of Crry and CD59 leads to increased intraocular complement activation as evidenced by the increased deposition of C3 activation products and MAC. Taken together, our data demonstrated that the direct interference with the function and/or cell surface expression of Crry and CD59 in vivo resulted in a significant reduction in Crry and CD59, increased complement activation and the exacerbation of EAAU (Jha et al., 2006b).
Complement system has also been reported to play an important role in the immunopathogenesis of experimental autoimmune uveoretinitis (EAU). EAU is an animal model of posterior uveitis (Schalken et al., 1988; Fox et al., 1987). In this model, complement depletion by cobra venom factor has been shown to decrease the inflammatory response (Marak et al., 1979). A recent report demonstrated that C3-deficient (C3-/-) mice had reduced severity and incidence of EAU (Read et al., 2006). In the same study it was demonstrated that the local expression of Crry resulted in decreased incidence and severity of EAU.
Taken together these studies indicate that complement plays an important role in the pathogenesis of autoimmune uveitis. However, the role of specific complement pathway(s) in the pathogenesis of EAAU or EAU remains to be established.
3.3 Complement and age-related macular degeneration
There is a substantial body of evidence implicating complement in age-related macular degeneration (AMD) both in humans and in experimental animals (Sivaprasad and Chong, 2006). AMD is the major cause of irreversible central vision loss in the elderly world wide (Tezel et al., 2004). Two major clinical phenotypes of AMD are recognized - non-exudative (dry type), and exudative (wet type).
Complement C3adesArg concentration has been found to increase in the plasma of patients with AMD compared to the age-matched controls (Sivaprasad et al., 2007). The presence of C3adesArg is an indicator of complement activation. One of the early signs of nonexudative AMD is the accumulation of abnormal extracellular deposits, called drusen, adjacent to the basal surface of the retinal-pigmented epithelium (RPE) (Johnson et al., 2000, 2001). Drusens are located between the basal lamina of RPE and the inner layer of Bruch’s membrane. Various studies in the literature have indicated a potential role for complement in drusen formation in the non-exudative form of AMD in humans (Johnson et al., 2000, 2001; Mullins 2001). Complement components, complement activation products (C3a, C5a, MAC) and complement regulatory proteins (CD46, Vitronectin) have been localized in drusens in patients with AMD (Mullins et al., 2000; Johnson et al., 2001; Anderson et al., 2002). One study reported that antibody-dependent complement activation may not be responsible for drusen formation (Johnson et al., 2001) and suggested that complement activation via the alternative pathway may be involved in this process. In the same study many complement regulators including - vitronectin, CR1 and MCP were detected in drusens. Also factor H and FHL-1 protein has been shown to be present in macular region in the eyes of patients with early AMD (Skerka et al., 2007).
Recently, variation in the factor H gene (HF1) that encodes a key regulatory protein of the alternative pathway has been associated with significantly increased attributable risk (50%) of AMD in humans (Rivera et al., 2005; Klein et al., 2005; Edwards et al., 2005; Haines et al., 2005; Hageman et al., 2005). The human gene locus for CFH encodes two proteins- CFH (composed of 20 SCR domains) and factor H-like protein (FHL-1; composed of 7 N-terminal domains). Both CFH and FHL-1 regulate the complement system by acting as a cofactor for factor I mediated inactivation of C3b and have decay accelerating activities. Both CFH and FHL-1 are present in vitreous fluid of eye and expressed by RPE cells (Skerka et al., 2007). A tyrosine to histidine mutation at codon 402 (Y402H) within CFH is strongly associated with AMD pathology (Rivera et al., 2005; Klein et al., 2005; Edwards et al., 2005; Haines et al., 2005; Hageman et al., 2005) and has been suggested to be a global marker for AMD (Kaur et al., 2006). The tyrosine residue at position (CFHYY402) is considered as protective variant whereas the histidine residue at this position (CFHYH402 and CFHHH402) a risk variant for AMD. Recently FHL-1 402H risk variant protein has also been reported to be associated with pathology of AMD (Skerka et al., 2007). Due to the coding polymorphism at position 402, risk variants of both CFH and FHL-1 have reduced cell-binding activity that leads to reduced complement regulation at the cell surface (Skerka et al., 2007). Also, the risk variants of these two regulators have decreased cofactor activity and are less efficient in degrading C3 (Skerka et al., 2007). Defective function of CFH and FHL-1 due to the polymorphism leads to unregulated complement activation at the cell surface. This results in induction of the inflammatory cascade leading to the tissue damage and ultimately drusen formation in AMD (Skerka et al., 2007). Recent findings have demonstrated that polymorphisms in two other complement proteins, factor B (BF) and C2, are also associated with AMD (Gold et al, 2006). The results from these studies suggest that a defect in a component of the alternative pathway provides protection whereas a defect in a regulator of alternative pathway like CFH increases the risk of AMD (Gold et al, 2006).
Choroidal neovascularization (CNV), the hallmark lesion of wet AMD, causes the sudden loss of central vision and thus, has a profound impact on the quality of life. One of the ways to induce CNV in animals is to rupture Bruch’s membrane with laser photocoagulation (Ryan, 1982; Miller et al., 1990; Kimura et al., 1995). We have used the murine model of laser-induced CNV to explore the role of complement in choroidal angiogenesis (Bora et al., 2005). The requirement of complement in the development of CNV was demonstrated by the inability to induce CNV in C3-/- mice after laser treatment. Furthermore, in vivo complement depletion by CVF and inhibition of C6 with anti-C6 polyclonal antibody, markedly reduced the incidence and development of CNV in C57BL/6 mice. There was an increased deposition of MAC in the CNV complex of complement sufficient animals. The absence of MAC in the laser spots also coincided with low/basal levels of various growth factors such as VEGF, TGF-β2 and β-FGF (Bora et al., 2005).
In a separate study, we confirmed the role of MAC in CNV by using C5-deficient mice (C5-/-) (Bora et al., 2006). These mice are unable to form MAC and laser-induced CNV was significantly inhibited (Bora et al., 2006). In the same study, siRNA and mice deficient in key components of the complement cascade were used to define the relative contributions of the classical, lectin, and alternative pathways in laser-induced CNV in C57BL/6 mice. The absence of C1q as a result of C1q siRNA treatment had no effect on laser-induced CNV. Furthermore, mice with targeted deletion of C4 gene also developed CNV following laser. Thus, by using C4-/- mice and inhibiting C1q by siRNA, involvement of the classical and lectin pathways in the development of CNV was ruled out. Inhibition of Factor B using siRNA targeted against factor B, completely inhibited CNV. Factor B is a key component of the alternative pathway. Thus the inhibition of CNV in factor B siRNA treated mice implicated the alternative pathway as a key player in the development of laser-induced CNV in mouse. The role of the alternative pathway in laser-induced CNV was further supported by the observation that expression of factor B was up-regulated, while expression of factor H was down-regulated during the development of CNV in complement-sufficient animals.
Recently, we explored the expression and role of the CReg CD59a in laser-induced CNV (Bora et al., 2007b). CD59a is the primary regulator of MAC in mice (Baalasubramanian et al., 2004). In this study we observed that the levels of CD59a were down-regulated during laser-induced CNV, indicating that this may facilitate complement activation leading to increased MAC deposition and release of angiogenic factors, thus inducing choroidal angiogenesis. Furthermore, deficiency of the CD59a gene exacerbated laser-induced CNV by allowing unregulated MAC deposition. Interestingly, inhibition of MAC formation following intra-peritoneal injection of recombinant soluble CD59a inhibited the development of choroidal angiogenesis in the mouse model of laser-induced CNV.
Taken together, our studies demonstrate that the formation of MAC as a result of complement activation via the alternative pathway in the posterior segment of the eye is crucial for the release of angiogenic growth factors that drive the development of laser-induced CNV in mice. The alternative pathway activation during CNV is due to increased production of a key component, factor B, as well as reduced expression of regulatory proteins, factor H and CD59. Specific blockade of the complement pathway using recombinant CRegs such as CFH and CD59 may represent a novel therapeutic strategy for AMD in future. We have summarized our results in Figure 1.
Figure 1. Complement activation in the development of CNV.
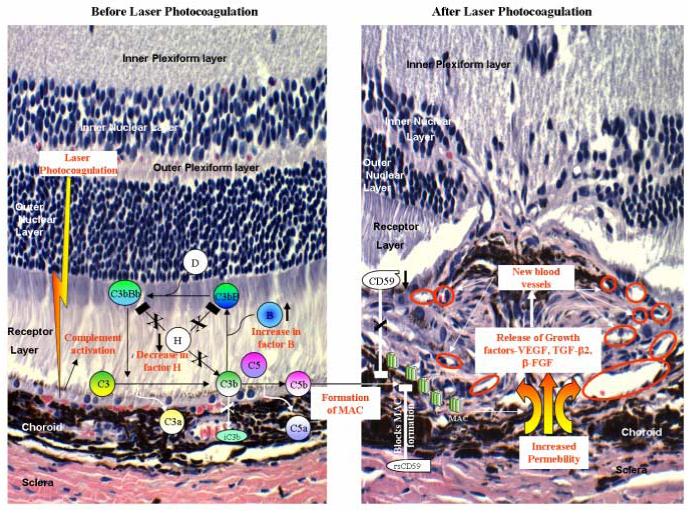
Laser photocoagulation results in up-regulation of factor B (↑) expression and decrease in (↓) factor H levels. This causes the activation and amplification of alternative pathway leading to increased deposition of MAC on the cell surface. After laser treatment the expression of CD59 is also down-regulated (↓) causing further loss of regulation (shown with X) and increased formation of MAC. Deposition of MAC results in release of growth factors such as VEGF, TGF-β2 and β-FGF, which leads to the formation of new blood vessels (shown within red circles). Administration of recombinant soluble CD59 (rs CD59) blocks (□) the formation of MAC and this results in inhibition of choroidal angiogenesis
Recently, one study demonstrated the importance of the anaphylotoxins C3a and C5a that are also found in drusens (Nozaki et al., 2006). Mice deficient in C3aR and C5aR, the receptors for these inflammatory peptides, developed significantly reduced CNV after laser treatment. The reduction in CNV was attributed to reduced leukocyte recruitment and VEGF expression due to the absence of C3aR and C5aR (Nozaki et al., 2006).
3.4 Complement and diabetic retinopathy
Diabetic retinopathy is one of the leading causes of vision loss in middle-aged individuals (Moss et al., 1998). It is characterized by the loss of pericytes, hypertrophy of basement membrane, endothelial leakage, neovascularization and fibrovascular proliferation. Usually, loss of vision is caused by proliferative diabetic retinopathy (caused by proliferation of retinal vessels) and diabetic macular edema (caused by increase in permeability of vessels in retina; Aiello, 2005). It is now well established that the immune system is involved in the pathogenesis of diabetic retinopathy (Kaštelan et al., 2007). Several studies have shown that activation of the complement system plays an important role in the pathogenesis of diabetic retinopathy. Gerl et al., (2002) reported the evidence for extensive complement activation in the choriocapillaries of eyes with diabetic retinopathy. Using immunohistochemistry, these investigators found dense staining for MAC and C3d, implicating complement activation in eyes with diabetic retinopathy. However, in the same study no positive staining was observed for C1q, C4 or MBL eliminating the involvement of classical and lectin pathways of complement activation (Gerl et al., 2002). Similar results were also reported by Zhang et al. (2002), who have shown that, although MAC staining co-localized with C3 in the retinal sections of the eyes obtained from diabetic patients; these sections did not stain for C1q and C4. Therefore the observed deposition of MAC may be the result of complement activation via the alternative pathway. Retinal samples from diabetic donors showed a significant reduction in CD55 and CD59, indicating the loss of regulatory mechanism may be the cause of increased complement activation (Zhang et al., 2002). Similarly, increased complement activation and decrease in CD59 and CD55 levels were also observed in rats with streptozotocin-induced diabetes (Zhang et al., 2002). In a separate study, the involvement of alternative pathway was confirmed by detection of factor B in the vitreous fluid of patients with proliferative diabetic retinopathy (Garcia-Ramirez et al., 2007). In addition to factor B, other complement proteins like C3, C4b and C9 were also detected at higher levels in vitreous fluid from patients with proliferative diabetic retinopathy compared to those from non-diabetic patients. These results indicate that, although alternative pathway is important during early phase of diabetic retinopathy, classical pathway may also be involved during the later stages of the disease. Activation of complement system has been implicated in other retinal diseases, including vitreoretinopathy (Grisanti et al., 1991) and glaucoma (Ahmed et al., 2004).
In conclusion, complement activation and subsequent generation of inflammatory molecules contributes to the pathology of ocular diseases such as uveitis, age-related macular degeneration, corneal diseases and diabetic retinopathy. Therefore, continued efforts are necessary to fully understand the role of complement in the pathogenesis of these ocular diseases so that efficient and safe therapies for future use could be developed.
Acknowledgements
This work was supported in part by NIH grants EY13335, EY014623, EY016205, and by an unrestricted grant from Research to Prevent Blindness and the Pat & Willard Walker Eye Research Center, Harvey & Bernice Jones Eye Institute, University of Arkansas for Medical Sciences.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Ahmed F, Brown KM, Stephan DA, Morrison JC, Johnson EC, Tomarev SI. Microarray analysis of changes in mRNA levels in the rat retina after experimental elevation of intraocular pressure. Invest. Ophthalmol. Vis. Sci. 2004;45:1247–1258. doi: 10.1167/iovs.03-1123. [DOI] [PubMed] [Google Scholar]
- Aiello LP. Angiogenic pathways in diabetic retinopathy. N. Engl. J. Med. 2005;353:839–841. doi: 10.1056/NEJMe058142. [DOI] [PubMed] [Google Scholar]
- Anderson DH, Mullins RF, Hageman GS, Johnson LV. A role for local inflammation in the formation of drusen in the aging eye. Am. J. Ophthalmol. 2002;134:411–431. doi: 10.1016/s0002-9394(02)01624-0. [DOI] [PubMed] [Google Scholar]
- Atkinson JP, Frank MM. Complement. In: Parker CW, editor. Clinical Immunology. WB Saunders; Philadelphia: 1980. pp. 219–271. [Google Scholar]
- Atkinson JP, Farries T. Separation of self from non-self in the complement system. Immunol. Today. 1987;8:212–215. doi: 10.1016/0167-5699(87)90167-8. [DOI] [PubMed] [Google Scholar]
- Baalasubramanian S, Harris CL, Donev RM, Mizuno M, Omidvar N, Song WC, Morgan BP. CD59a is the primary regulator of membrane attack complex assembly in the mouse. J. Immunol. 2004;173:3684–3692. doi: 10.4049/jimmunol.173.6.3684. [DOI] [PubMed] [Google Scholar]
- Bardenstein DS, Dietz Y, Lass JH, Medof ME. Localization of the complement membrane attack complex inhibitor (CD59) in human conjunctiva and lacrimal gland. Curr. Eye Res. 1994;13:851–855. doi: 10.3109/02713689409015085. [DOI] [PubMed] [Google Scholar]
- Bora NS, Gobleman CL, Atkinson JP, Pepose JS, Kaplan HJ. Differential expression of the complement regulatory proteins in the human eye. Invest. Ophthalmol. Vis. Sci. 1993;34:3579–3584. [PubMed] [Google Scholar]
- Bora NS, Kaplan HJ. Intraocular diseases - anterior uveitis. Chem. Immunol. Allergy. 2007;92:213–220. doi: 10.1159/000099272. [DOI] [PubMed] [Google Scholar]
- Bora NS, Kim MC, Kabeer NH, Simpson SC, Tandhasetti MT, Cirrito TP, Kaplan AD, Kaplan HJ. Experimental autoimmune anterior uveitis. Induction with melanin-associated antigen from the iris and ciliary body. Invest. Ophthalmol. Vis. Sci. 1995;36:1056–1066. [PubMed] [Google Scholar]
- Bora NS, Woon MD, Tandhasetti MT, Cirrito TP, Kaplan HJ. Induction of experimental autoimmune anterior uveitis by a self-antigen: melanin complex without adjuvant. Invest. Ophthalmol. Vis. Sci. 1997;38:2171–2175. [PubMed] [Google Scholar]
- Bora NS, Sohn JH, Kang SG, Cruz JM, Nishihori H, Suk HJ, Wang Y, Kaplan HJ, Bora PS. Type I collagen is the autoantigen in experimental autoimmune anterior uveitis. J. Immunol. 2004;172:7086–7094. doi: 10.4049/jimmunol.172.11.7086. [DOI] [PubMed] [Google Scholar]
- Bora PS, Sohn JH, Cruz JM, Jha P, Nishihori H, Wang Y, Kaliappan S, Kaplan HJ, Bora NS. Role of complement and complement membrane attack complex in laser-induced choroidal neovascularization. J. Immunol. 2005;174:491–497. doi: 10.4049/jimmunol.174.1.491. [DOI] [PubMed] [Google Scholar]
- Bora NS, Kaliappan S, Jha P, Xu Q, Sohn JH, Dhaulakhandi DB, Kaplan HJ, Bora PS. Complement activation via alternative pathway is critical in the development of laser-induced choroidal neovascularization: role of factor B and factor H. J. Immunol. 2006;177:1872–1878. doi: 10.4049/jimmunol.177.3.1872. [DOI] [PubMed] [Google Scholar]
- Bora NS, Kaliappan S, Jha P, Xu Q, Sivasankar B, Harris CL, Morgan BP, Bora PS. CD59, a complement regulatory protein, controls choroidal neovascularization in a mouse model of wet-type age-related macular degeneration. J. Immunol. 2007;178:1783–1790. doi: 10.4049/jimmunol.178.3.1783. [DOI] [PubMed] [Google Scholar]
- Broekhuyse RM, Kuhlman ED, Winkens HJ, Van Vugt AHM. Experimental autoimmune anterior uveitis (EAAU), a new form of experimental uveitis: I: Induction by a detergent-insoluble, intrinsic protein fraction of the retina pigment epithelium Exp. Eye Res. 1991;52:465–474. doi: 10.1016/0014-4835(91)90044-f. [DOI] [PubMed] [Google Scholar]
- Cleveland RP, Hazlett LD, Leon MA, Berk RS. Role of complement in murine corneal infection caused by Pseudomonas aeruginosa. Invest. Ophthalmol. Vis. Sci. 1983;24:237–242. [PubMed] [Google Scholar]
- Cocuzzi E, Guidubaldi J, Bardenstein DS, Chen R, Jacobs MR, Medof EM. Release of complement regulatory proteins from ocular surface cells in infections. Curr. Eye Res. 2000;21:856–866. doi: 10.1076/ceyr.21.5.856.5539. [DOI] [PubMed] [Google Scholar]
- Edwards AO, Ritter R, 3rd, Abel KJ, Manning A, Panhuysen C, Farrer LA. Complement factor H polymorphism and age-related macular degeneration. Science. 2005;308:421–424. doi: 10.1126/science.1110189. [DOI] [PubMed] [Google Scholar]
- Fox GM, Kuwabara T, Wiggert B, Redmond TM, Hess HH, Chader GJ, Gery I. Experimental autoimmune uveoretinitis (EAU) induced by retinal interphotoreceptor retinoid-binding protein (IRBP): differences between EAU induced by IRBP and by S-antigen. Clin. Immunol. Immunopathol. 1987;43:256–264. doi: 10.1016/0090-1229(87)90133-4. [DOI] [PubMed] [Google Scholar]
- Frank MM, Fries LF. The role of complement in inflammation and phagocytosis. Immunol. Today. 1991;12:322–326. doi: 10.1016/0167-5699(91)90009-I. [DOI] [PubMed] [Google Scholar]
- Garcia-Ramirez M, Canals F, Hernandez C, Colome N, Ferrer C, Carrasco E, Garcia-Arumi J, Simo R. Proteomic analysis of human vitreous fluid by fluorescence-based difference gel electrophoresis (DIGE): a new strategy for identifying potential candidates in the pathogenesis of proliferative diabetic retinopathy. Diabetologia. 2007;50:1294–1303. doi: 10.1007/s00125-007-0627-y. [DOI] [PubMed] [Google Scholar]
- Gerl VB, Bohl J, Pitz S, Stoffelns B, Pfeiffer N, Bhakdi S. Extensive deposits of complement C3d and C5b-9 in the choriocapillaris of eyes of patients with diabetic retinopathy. Invest. Ophthalmol. Vis. Sci. 2002;43:1104–1108. [PubMed] [Google Scholar]
- Gold B, Merriam JE, Zernant J, Hancox LS, Taiber AJ, Gehrs K, Cramer K, Neel J, Bergeron J, Barile GR, Smith RT, AMD Genetics Clinical Study Group. Hageman GS, Dean M, Allikmets R. Variation in factor B (BF) and complement component 2 (C2) genes is associated with age-related macular degeneration. Nat. Genet. 2006;38:458–462. doi: 10.1038/ng1750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grisanti S, Wiedemann P, Weller M, Heimann K, Zilles K. The significance of complement in proliferative vitreoretinopathy. Invest. Ophthalmol. Vis. Sci. 1991;32:2711–2717. [PubMed] [Google Scholar]
- Hageman GS, Anderson DH, Johnson LV, Hancox LS, Taiber AJ, Hardisty LI, Hageman JL, Stockman HA, Borchardt JD, Gehrs KM, Smith RJ, Silvestri G, Russell SR, Klaver CC, Barbazetto I, Chang S, Yannuzzi LA, Barile GR, Merriam JC, Smith RT, Olsh AK, Bergeron J, Zernant J, Merriam JE, Gold B, Dean M, Allikmets R. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad, Sci. U S A. 2005;102:7227–7232. doi: 10.1073/pnas.0501536102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haines JL, Hauser MA, Schmidt S, Scott WK, Olson LM, Gallins P, Spencer KL, Kwan SY, Noureddine M, Gilbert JR, Schnetz-Boutaud N, Agarwal A, Postel EA, Pericak-Vance MA. Complement factor H variant increases the risk of age-related macular degeneration. Science. 2005;308:419–421. doi: 10.1126/science.1110359. [DOI] [PubMed] [Google Scholar]
- Hazlett LD, Berk RS. Effect of C3 depletion on experimental Pseudomonas aeruginosa ocular infection: histopathological analysis. Infect Immun. 1984;43:783–790. doi: 10.1128/iai.43.3.783-790.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holguin MH, Parker CJ. Membrane inhibitor of reactive lysis. Curr. Topics Microbiol. Immunol. 1992;178:61–85. doi: 10.1007/978-3-642-77014-2_5. [DOI] [PubMed] [Google Scholar]
- Jha P, Sohn JH, Xu Q, Wang Y, Kaplan HJ, Bora PS, Bora NS. Suppression of complement regulatory proteins (CRPs) exacerbates experimental autoimmune anterior uveitis (EAAU) J. Immunol. 2006;176:7221–7231. doi: 10.4049/jimmunol.176.12.7221. [DOI] [PubMed] [Google Scholar]
- Jha P, Sohn JH, Qin X, Nishihori H, Wang Y, Nishihori S, Manickam B, Kaplan HJ, Bora PS, Bora NS. Complement system plays a critical role in the development of experimental autoimmune anterior uveitis (EAAU) Invest. Ophthalmol. Vis. Sci. 2006;47:1030–1038. doi: 10.1167/iovs.05-1062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson LV, Ozaki S, Staples MK, Erickson PA, Anderson DH. A potential role for immune complex pathogenesis in drusen formation. Exp. Eye Res. 2000;70:441–449. doi: 10.1006/exer.1999.0798. [DOI] [PubMed] [Google Scholar]
- Johnson LV, Leitner WP, Staples MK, Anderson DH. Complement activation and inflammatory processes in Drusen formation and age related macular degeneration. Exp. Eye Res. 2001;73:887–896. doi: 10.1006/exer.2001.1094. [DOI] [PubMed] [Google Scholar]
- Kaštelan S, Zjačić-Rotkvić V, Kaštelan Ž. Could diabetic retinopathy be an autoimmune disease? Medical Hypotheses. 2007;68:1016–1018. doi: 10.1016/j.mehy.2006.05.073. [DOI] [PubMed] [Google Scholar]
- Kaur I, Hussain A, Hussain N, Das T, Pathangay A, Mathai A, Hussain A, Nutheti R, Nirmalan PK, Chakrabarti S. Analysis of CFH, TLR4, and APOE polymorphism in India suggests the Tyr402His variant of CFH to be a global marker for age-related macular degeneration. Invest. Ophthalmol. Vis. Sci. 2006;47:3729–3735. doi: 10.1167/iovs.05-1430. [DOI] [PubMed] [Google Scholar]
- Kim YU, Kinoshita T, Molina H, Hourcade D, Seya T, Wagner LM, Holers VM. Mouse complement regulatory protein Crry/p65 uses the specific mechanisms of both human decay-accelerating factor and membrane cofactor protein. J. Exp. Med. 1995;181:151–159. doi: 10.1084/jem.181.1.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim MC, Kabeer NH, Tandhasetti MT, Kaplan HJ, Bora NS. Immunohistochemical studies in melanin associated antigen (MAA) induced experimental autoimmune anterior uveitis (EAAU) Curr. Eye Res. 1995;14:703–710. doi: 10.3109/02713689508998498. [DOI] [PubMed] [Google Scholar]
- Kimura H, Sakamoto T, Hinton DR, Spee C, Ogura Y, Ryan SJ. A new model of subretinal neovascularization in the rabbit. Invest. Ophthalmol. Vis. Sci. 1995;36:2110–2119. [PubMed] [Google Scholar]
- Klein RJ, Zeiss C, Chew EY, Tsai JY, Sackler RS, Haynes C, Henning AK, SanGiovanni JP, Mane SM, Mayne ST, Bracken MB, Ferris FL, Ott J, Barnstable C, Hoh J. Complement factor H polymorphism in age-related macular degeneration. Science. 2005;308(5720):385–389. doi: 10.1126/science.1109557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krych-Goldberg M, Atkinson JP. Structure-function relationships of complement receptor type 1. Immunol. Rev. 2001;180:112–122. doi: 10.1034/j.1600-065x.2001.1800110.x. [DOI] [PubMed] [Google Scholar]
- Li B, Sallee C, Dehoff M, Foley S, Molina H, Holers VM. Mouse Crry/p65: characterization of monoclonal antibodies and the tissue distribution of a functional homologue of human MCP and DAF. J. Immunol. 1993;151:4295–4305. [PubMed] [Google Scholar]
- Liszewski MK, Farries TC, Lublin DM, Rooney IA, Atkinson JP. Control of the complement system. Adv. Immunol. 1996;61:201–283. doi: 10.1016/s0065-2776(08)60868-8. [DOI] [PubMed] [Google Scholar]
- Liszewski MK, Post TW, Atkinson JP. Membrane cofactor protein (MCP or CD46): Newest member of the regulators of complement activation gene cluster. Ann. Rev. Immunol. 1991;9:431–455. doi: 10.1146/annurev.iy.09.040191.002243. [DOI] [PubMed] [Google Scholar]
- Liszewski MK, Atkinson JP. Membrane cofactor protein. Curr. Topics Microbiol. Immunol. 1992;178:45–60. doi: 10.1007/978-3-642-77014-2_4. [DOI] [PubMed] [Google Scholar]
- Lublin DM, Atkinson JP. Decay-accelerating factor and membrane cofactor protein. Curr. Topics Microbiol. Immunol. 1987;153:123–145. doi: 10.1007/978-3-642-74977-3_7. [DOI] [PubMed] [Google Scholar]
- Marak GE, Wacker WB, Rao NA, Jack R, Ward PA. Effects of complement depletion on experimental allergic uveitis. Ophthalmic. Res. 1979;11:97–107. [Google Scholar]
- Miller H, Miller B, Ishibashi T, Ryan SJ. Pathogenesis of laser-induced choroidal subretinal neovascularization. Invest. Ophthalmol. Vis. Sci. 1990;31:899–908. [PubMed] [Google Scholar]
- Miwa T, Nonaka M, Okada N, Wakana S, Shiroishi T, Okada H. Molecular cloning of rat and mouse membrane cofactor protein (MCP, CD46): preferential expression in testis and close linkage between the mouse Mcp and Cr2 genes on distal chromosome 1. Immunogenetics. 1998;48:363–371. doi: 10.1007/s002510050447. [DOI] [PubMed] [Google Scholar]
- Molina H, Wong W, Kinoshita T, Brenner C, Foley S, Holers VM. Distinct receptor and regulatory properties of recombinant mouse complement receptor 1 (CR1) and Crry, the two genetic homologues of human CR1. J. Exp. Med. 1992;175:121–129. doi: 10.1084/jem.175.1.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mondino BJ, Brady KJ. Distribution of hemolytic complement in the normal cornea. Arch. Ophthalmol. 1981;99:1430–1433. doi: 10.1001/archopht.1981.03930020304022. [DOI] [PubMed] [Google Scholar]
- Mondino BJ, Rao H. Complement levels in normal and inflamed aqueous humor. Invest. Ophthalmol. Vis. Sci. 1983;24:380–384. [PubMed] [Google Scholar]
- Mondino BJ, Sumner HL. Generation of complement-derived anaphylatoxins in normal human donor corneas. Invest. Ophthalmol. Vis. Sci. 1990;31:1945–1949. [PubMed] [Google Scholar]
- Mondino BJ, Chou HJ, Sumner HL. Generation of complement membrane attack complex in normal human corneas. Invest. Ophthalmol. Vis. Sci. 1996;37:1576–1581. [PubMed] [Google Scholar]
- Mondino BJ, Glovsky MM, Chekiere L. Activated complement in inflamed aqueous humor. Invest. Ophthalmol. Vis. Sci. 1984;25:871–873. [PubMed] [Google Scholar]
- Mondino BJ, Sumner H. Anaphylatoxin levels in human aqueous humor. Invest. Ophthalmol. Vis. Sci. 1986;27:1288–1292. [PubMed] [Google Scholar]
- Morgan BP, Harris CL. The complement system. In: Morgan BP, Harris CL, editors. Complement regulatory proteins. Academic Press; San Diego: 1999. pp. 1–13. [Google Scholar]
- Moss S, Klein R, Klein B. The 14-year incidence of visual loss in a diabetic population. Ophthalmology. 1998;105:998–1003. doi: 10.1016/S0161-6420(98)96025-0. [DOI] [PubMed] [Google Scholar]
- Muller-Eberhard HJ. Molecular organization and function of the complement system. Annu. Rev. Biochem. 1988;57:321–347. doi: 10.1146/annurev.bi.57.070188.001541. [DOI] [PubMed] [Google Scholar]
- Mullins RF, Aptsiauri N, Hageman GS. Structure and composition of drusen associated with glomerulonephritis: implications for the role of complement activation in drusen biogenesis. Eye. 2001;15:390–395. doi: 10.1038/eye.2001.142. [DOI] [PubMed] [Google Scholar]
- Mullins RF, Russell SR, Anderson DH, Hageman GS. Drusen associated with aging and age-related macular degeneration contain proteins common to extracellular deposits associated with atherosclerosis, elastosis, amyloidosis, and dense deposit disease. FASEB J. 2000;14:835–846. [PubMed] [Google Scholar]
- Nicholson-Weller A. Decay accelerating factor (CD55) Curr. Topics Microbiol. Immunol. 1992;178:7–30. doi: 10.1007/978-3-642-77014-2_2. [DOI] [PubMed] [Google Scholar]
- Nickells MW, Alvarez JI, Lublin DM, Atkinson JP. Characterization of DAF-2, a high molecular weight form of decay-accelerating factor (DAF; CD55), as a covalently cross-linked dimer of DAF-1. J. Immunol. 1994;152:676–685. [PubMed] [Google Scholar]
- Nonaka M, Miwa T, Okada N, Nonaka M, Okada H. Multiple isoforms of guinea pig decay-accelerating factor (DAF) generated by alternative splicing. J. Immunol. 1995;155:3037–3048. [PubMed] [Google Scholar]
- Nozaki M, Raisler BJ, Sakurai E, Sarma JV, Barnum SR, Lambris JD, Chen Y, Zhang K, Ambati BK, Baffi JZ, Ambati J. Drusen complement components C3a and C5a promote choroidal neovascularization. Proc. Natl. Acad. Sci. U S A. 2006;103:2328–2333. doi: 10.1073/pnas.0408835103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell MB, Marchbank KJ, Rushmere NK, van den Berg CW, Morgan BP. Molecular cloning, chromosomal localization, expression, and functional characterization of the mouse analogue of human CD59. J. Immunol. 1997;158:1692–1702. [PubMed] [Google Scholar]
- Qian YM, Qin X, Miwa T, Sun X, Halperin JA, Song WC. Identification and functional characterization of a new gene encoding the mouse terminal complement inhibitor CD59. J. Immunol. 2000;165:2528–2534. doi: 10.4049/jimmunol.165.5.2528. [DOI] [PubMed] [Google Scholar]
- Read RW, Szalai AJ, Vogt SD, McGwin G, Barnum SR. Genetic deficiency of C3 as well as CNS-targeted expression of the complement inhibitor sCrry ameliorates experimental autoimmune uveoretinitis. Exp. Eye Res. 2006;82:389–394. doi: 10.1016/j.exer.2005.07.011. [DOI] [PubMed] [Google Scholar]
- Reid KBM. Activation and control of the complement system. Essays Biochem. 1986;22:27–68. [PubMed] [Google Scholar]
- Rivera A, Fisher SA, Fritsche LG, Keilhauer CN, Lichtner P, Meitinger T, Weber BH. Hypothetical LOC387715 is a second major susceptibility gene for age-related macular degeneration, contributing independently of complement factor H to disease risk. Hum. Mol. Genet. 2005;14:3227–3236. doi: 10.1093/hmg/ddi353. [DOI] [PubMed] [Google Scholar]
- Ross GD, editor. Immunobiology of the complement system. Academic Press Inc; Florida: 1986. [Google Scholar]
- Ryan SJ. Subretinal neovascularization. Natural history of an experimental model. Arch. Ophthalmol. 1982;100:1804–1809. doi: 10.1001/archopht.1982.01030040784015. [DOI] [PubMed] [Google Scholar]
- Schalken JJ, Winkens HJ, van Vugt AH, Bovee-Geurts PH, de Grip WJ, Broekhuyse RM. Rhodopsin-induced experimental autoimmune uveoretinitis: dose-dependent clinicopathological features. Exp. Eye Res. 1988;47:135–145. doi: 10.1016/0014-4835(88)90030-9. [DOI] [PubMed] [Google Scholar]
- Simpson SC, Kaplan HJ, Bora NS. Uveitogenic proteins isolated from bovine iris and ciliary body. Eye. 1997;11:206–208. doi: 10.1038/eye.1997.52. [DOI] [PubMed] [Google Scholar]
- Sivaprasad S, Chong NV. The complement system and age-related macular degeneration. Eye. 2006;20:867–872. doi: 10.1038/sj.eye.6702176. [DOI] [PubMed] [Google Scholar]
- Sivaprasad S, Adewoyin T, Bailey TA, Dandekar SS, Jenkins S, Webster AR, Chong NV. Estimation of systemic complement C3 activity in age-related macular degeneration. Arch. Ophthalmol. 2007;125:515–519. doi: 10.1001/archopht.125.4.515. [DOI] [PubMed] [Google Scholar]
- Skerka C, Lauer N, Weinberger AA, Keilhauer CN, Suhnel J, Smith R, Schlotzer-Schrehardt U, Fritsche L, Heinen S, Hartmann A, Weber BH, Zipfel PF. Defective complement control of Factor H (Y402H) and FHL-1 in age-related macular degeneration. Mol. Immunol. 2007;44:3398–3406. doi: 10.1016/j.molimm.2007.02.012. [DOI] [PubMed] [Google Scholar]
- Sohn JH, Kaplan HJ, Suk HJ, Bora PS, Bora NS. Chronic low level complement activation within the eye is controlled by intraocular complement regulatory proteins. Invest. Ophthalmol. Vis. Sci. 2000;41:3492–3502. [PMC free article] [PubMed] [Google Scholar]
- Sohn JH, Kaplan HJ, Suk HJ, Bora PS, Bora NS. Complement regulatory activity of normal human intraocular fluid is mediated by MCP, DAF, and CD59. Invest. Ophthalmol. Vis. Sci. 2000;41:4195–4202. [PMC free article] [PubMed] [Google Scholar]
- Spicer AP, Seldin MF, Gendler SJ. Molecular cloning and chromosomal localization of the mouse decay-accelerating factor genes. Duplicated genes encode glycosylphosphatidylinositol-anchored and transmembrane forms. J. Immunol. 1995;155:3079–3091. [PubMed] [Google Scholar]
- Takizawa H, Okada N, Okada H. Complement inhibitor of rat cell membrane resembling mouse Crry/p65. J. Immunol. 1994;152:3032–3038. [PubMed] [Google Scholar]
- Tezel TH, Bora NS, Kaplan HJ. Pathogenesis of age-related macular degeneration. Trends Mol. Med. 2004;10:417–420. doi: 10.1016/j.molmed.2004.07.004. [DOI] [PubMed] [Google Scholar]
- Thiel S, Vorup-Jensen T, Stover CM, Schwaeble W, Laursen SB, Poulsen K, Willis AC, Eggleton P, Hansen S, Holmskov U, Reid KB, Jensenius JC. A second serine protease associated with mannan-binding lectin that activates complement. Nature. 1997;386:506–510. doi: 10.1038/386506a0. [DOI] [PubMed] [Google Scholar]
- Thurman JM, Holers VM. The central role of the alternative complement pathway in human disease. J. Immunol. 2006;176:1305–1310. doi: 10.4049/jimmunol.176.3.1305. [DOI] [PubMed] [Google Scholar]
- Watts MJ, Dankert JR, Morgan BP. Isolation and characterization of a membrane-attack-complex inhibiting protein present in human serum and other biological fluids. Biochem. J. 1990;265:471–477. doi: 10.1042/bj2650471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler SF, Rudd PM, Davis SJ, Dwek RA, Harvey DJ. Comparison of the N-linked glycans from soluble and GPI-anchored CD59 expressed in CHO cells. Glycobiology. 2002;12:261–271. doi: 10.1093/glycob/12.4.261. [DOI] [PubMed] [Google Scholar]
- Willcox MD, Morris CA, Thakur A, Sack RA, Wickson J, Boey W. Complement and complement regulatory proteins in human tears. Invest. Ophthalmol. Vis. Sci. 1997;38:1–8. [PubMed] [Google Scholar]
- Woon MD, Kaplan HJ, Bora NS. Kinetics of cytokine production in experimental autoimmune anterior uveitis (EAAU) Curr. Eye. Res. 1998;17:955–961. doi: 10.1076/ceyr.17.10.955.5246. [DOI] [PubMed] [Google Scholar]
- Zhang J, Gerhardinger C, Lorenzi M. Early complement activation and decreased levels of glycosylphosphatidylinositol-anchored complement inhibitors in human and experimental diabetic retinopathy. Diabetes. 2002;51:3499–3504. doi: 10.2337/diabetes.51.12.3499. [DOI] [PubMed] [Google Scholar]