

Abstract
NPC1L1 (Niemann–Pick C1-like 1), the pharmacological target of the cholesterol-uptake inhibitor ezetimibe, is a transporter localized on the brush border of enterocytes. Although this protein plays a key role in intestinal uptake of sterols, multiple molecular events that underlie intestinal cholesterol absorption have not been fully characterized. Two proteins that might be involved in this process are NPC1 and NPC2 (Niemann–Pick disease type C proteins 1 and 2), which function in the endosomal/lysosomal cholesterol egress pathway and whose deficiency results in NPC (Niemann–Pick type C) disease. The involvement of these proteins in intestinal cholesterol absorption was examined in mutant mice lacking either NPC1 or NPC2. Our data indicate that deficiencies in either protein do not have an effect on cholesterol uptake or absorption. This contrasts with recent results obtained for the fruitfly Drosophila melanogaster, which indicate that a deficiency of NPC1 (dNPC1a being its Drosophila homologue) leads to activation of an NPC1L1 (Drosophila homologue dNPC1b)-independent cholesterol uptake pathway, underscoring fundamental differences in mammalian and non-mammalian cholesterol metabolism.
Keywords: cholesterol transport, ezetimibe, lysosomal protein, Niemann–Pick type C disease (NPC disease), Niemann–Pick C1-like 1 (NPC1L1)
Abbreviations: dNPC1a and dNPC1b, putative Drosophila orthologues of mammalian NPC1 (Niemann–Pick disease type C protein 1) and NPC1L1 (Niemann–Pick C1-like 1) respectively; LDL, low-density lipoprotein; NPC, Niemann–Pick type C; NPC1L1, Niemann–Pick C1-like; NPC1 and NPC2, Niemann–Pick disease type C proteins 1 and 2; SSD, sterol-sensing domain
INTRODUCTION
Intestinal cholesterol absorption is an important biological process that entails multiple steps [1]. A central player in this process is NPC1L1 (Niemann–Pick C1-like 1) [2], which is an integral membrane protein localized on the brush border of absorptive enterocytes [2,3]. NPC1L1-deficient mice exhibited a markedly lower fractional cholesterol absorption compared with wild-type mice [2], and NPC1L1 is the target of ezetimibe, an azetidinone drug that lowers cholesterol levels by inhibiting its intestinal uptake [4,5]. These data demonstrate that NPC1L1 is intimately linked to transport of cholesterol across enterocyte membranes.
Two proteins that are critical for transport of cholesterol out of the endosomal/lysosomal system in various cell types are NPC1 and NPC2 (Niemann–Pick disease type C proteins 1 and 2). Genetic and phenotypic studies of mutant mice lacking either one or both proteins suggest that NPC1 and NPC2 participate in different steps of the same pathway, and neither can compensate for deficiency of the other [6]. Deficiency of either protein underlies NPC (Niemann–Pick type C) disease, a neurovisceral disorder characterized at the cellular level by lysosomal accumulation of cholesterol and other lipids.
NPC1, a lysosomal membrane glycoprotein [7], shares approx. 50% sequence identity with NPC1L1 [2]. In both proteins, five putative transmembrane helices constitute SSDs (sterol-sensing domains) similar to those of other integral membrane proteins involved in regulation of cholesterol [7]. Like NPC1L1, NPC1 is intimately involved in transmembrane cholesterol transport.
The other protein associated with NPC disease, NPC2, encoded by the gene NPC2, is a soluble lysosomal glycoprotein [8]. Recent in vitro studies showed that NPC2 can bind not only cholesterol but also other sterols [9] and it transfers cholesterol to phospholipid membranes [10,11]. One possible scenario is that NPC2 transfers cholesterol from intraluminal membranes or aggregates to NPC1, either directly or via the limiting lysosomal membrane, and NPC1 transports it out of the endosomal/lysosomal system.
NPC2 is also found extracellularly in fluids such as epididymal fluid [12], breast milk [13] and bile [14], where its function is unclear. Milk and bile, as well as secretion by intestinal cells can deliver NPC2 to the lumen of the intestine. Given the presumed functional interaction between NPC2 and NPC1, combined with the similarity between sequences and postulated molecular activities of NPC1 and NPC1L1, it is plausible that NPC2 might function together with NPC1L1 to transport cholesterol from intraluminal cholesterol–bile salt micelles to NPC1L1 and/or enterocyte brush-border membranes (Scheme 1, steps 2 and 3). If so, deficiencies in NPC2 should inhibit intestinal uptake of cholesterol.
Scheme 1. Postulated pathways for intestinal sterol absorption.

NPC1L1, a transporter located on the brush border of enterocytes, is required for uptake of cholesterol in the intestine. In the intestinal lumen, sterols might be presented to NPC1L1 in the form of mixed micelles (MM) (1) or NPC2, a lysosomal cholesterol-binding protein, might deliver the sterols to NPC1L1 (2). Once taken up in enterocytes via NPC1L1 (3), cholesterol is transported to the endoplasmic reticulum (ER) by pathways that might bypass (4) or involve (5) the endosomal (LE)/lysosomal (LYS) system. In the latter case, NPC1, a lysosomal cholesterol transporter, and NPC2 would be required for egress cholesterol from the endosomal/lysosomal system (6). Once in the endoplasmic reticulum (ER), cholesterol is esterified (CE) and packaged into chylomicron particles (CHY) that are exported in the lymph (7). TGN, trans-Golgi network.
It is also possible that NPC2 and NPC1 might be important in enterocytes for steps in cholesterol absorption that are downstream of its transport across the enterocyte brush border. Within enterocytes, cholesterol is transported into the lumen of the endoplasmic reticulum, esterified, packaged into chylomicrons, secreted across the basolateral membrane in the lymphatic system and eventually reaches the bloodstream and liver (Scheme 1, step 7). It is not known how cholesterol reaches the lumen of the endoplasmic reticulum in enterocytes, but it is possible that this process involves transport through the endosomal/lysosomal system (Scheme 1, steps 5 and 6), analogously to the transport of LDL (low-density lipoprotein)-derived cholesterol to the endoplasmic reticulum in various cell types. If so, then deficiencies in NPC2 or NPC1 should affect transport of cholesterol from enterocytes to the bloodstream and liver.
Intriguing observations were recently reported regarding cholesterol uptake in mutants of the fruitfly Drosophila melanogaster with defects in dNPC1a and dNPC1b, the putative orthologues of mammalian NPC1 and NPC1L1 respectively [15,16]. Like NPC1L1-deficient mice, dNPC1b-deficient fruitflies showed a decrease in cholesterol uptake. dNPC1a-deficient fruitflies showed a significant increase in cholesterol uptake either in wild-type or in dNPC1b-deficient backgrounds, suggesting activation of a dNPC1b-independent cholesterol uptake pathway. Given the biomedical importance of cholesterol uptake in humans, it is of considerable interest to decipher whether a similar NPC1-regulated cholesterol uptake pathway exists in mammals, as well as to elucidate other steps in enterocyte cholesterol trafficking.
To test these possibilities, we analysed fractional cholesterol absorption, sensitivity to ezetimibe treatment and acute intestinal uptake and absorption of cholesterol in wild-type, NPC1-deficient (NPC1−) and NPC2-hypomorph (NPC2−) mice.
MATERIALS AND METHODS
Chemicals
Cholesterol was purchased from Sigma–Aldrich (St Louis, MO, U.S.A.). [4-14C]Cholesterol (53 mCi/mmol) and [9,10-3H (n)]-trioleoylglycerol (66.8 Ci/mmol) were purchased from PerkinElmer Life and Analytical Sciences (Waltham, MA, U.S.A.). [5,6-3H]Sitostanol (50 Ci/mmol) was purchased from American Radiolabeled Chemicals (St Louis, MO, U.S.A.).
Animals and diets
All experiments and procedures involving live animals were conducted in compliance with animal protocols approved by the Robert Wood Johnson Medical School Institutional Animal Care and Use Committee. All mice had BALB/c genetic backgrounds; npcnih (NPC1−) [17] and NPC2 hypomorph (NPC2−) mice were described previously [6]. Mice were fed a standard chow diet (Picolab rodent diet; Labdiet Co., St Louis, MO, U.S.A.). During experiments, animals were housed in individual cages. Animals, consisting of males and females, were analysed at typically 5–7 weeks of age. At this age, the mice were presymptomatic in terms of exhibiting the marked weight loss and ataxia that are later manifestations of mutations in Npc1 or Npc2.
Measurement of fractional cholesterol absorption by the faecal dual-isotope method
Mice were orally gavaged with 2 μCi of [5,6-3H]sitostanol (17 ng) and 1 μCi of [4-14C]cholesterol (with or without additional unlabelled cholesterol, corresponding to either 0.11 mg or 0.01 mg) in 0.1 ml of corn oil. In the experiments involving ezetimibe, mice were gavaged with 0.1 ml of corn oil alone or containing 10 mg of ezetimibe/kg of body weight 30 min prior to the gavage with radiolabelled sterols and then gavaged with vehicle alone or ezetimibe after 24 h.
Faeces were collected daily for 2 days and analysed as a pool. The samples were dried at 60 °C for 48 h and then powdered. Two 0.1 g aliquots from each sample were extracted with 1.0 ml of chloroform/methanol (2:1, v/v) at 50 °C for 3 min and the extracts were dried [18]. The extracts were saponified by refluxing at 60 °C for 1 h in 0.5 ml of methanolic 1 M NaOH, following which the sterols were extracted in 0.5 ml of hexane and transferred to liquid-scintillation vials. After evaporation of hexane, samples were suspended in 2.0 ml of Ecoscint A (National Diagnostics USA, Atlanta, GA, U.S.A.) and counted for radioactivity in a Beckman LS 6500 liquid-scintillation counter. Data from the two aliquots were similar in all cases and were averaged. Statistical analyses were performed using Prism, version 5 (GraphPad Software, San Diego, CA, U.S.A.). Results were calculated as follows:
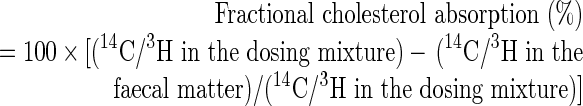 |
Measurement of acute cholesterol uptake and absorption
Mice were orally gavaged with a mixture containing 1 μCi of [4-14C]cholesterol (0.11 mg) and 1 μCi of [9,10-3H (n)]trioleoylglycerol (26 ng) in 0.1 ml of corn oil [2,19]. After 2 h, mice were deeply anaesthetized by intraperitoneal injection of pentobarbital sodium/Phenytoin (Euthasol; Delmarva Laboratories, Midlothian, VA, U.S.A.), killed by cardiac puncture and exsanguination, and the intestine, serum and liver were collected. The liver and intestine, which were pre-cleaned using 0.9% NaCl, were cut into small pieces.
The liver and intestine samples were mixed with chloroform/methanol (2:1, v/v) in a 1:20 (w/v) ratio [20]. The mixtures were agitated on a rotator for 2 h at room temperature (22 °C), and then centrifuged at 3500 g for 20 min. Supernatants were washed with 0.2 vol. of 0.9% NaCl by vortex-mixing for 30 s, followed by centrifugation at 1500 g for 10 min. The upper aqueous phases were removed by aspiration. The top surfaces of the organic phases were then washed twice by addition and removal of methanol/water (1:1, v/v) without mixing. The organic phases were air-dried and counted in 4.0 ml of Ecoscint A. Serum (0.2 ml) was counted directly in 4.0 ml of Ecoscint A and then scaled for each animal, assuming total blood volume is 4.0% of body weight.
RESULTS
Fractional cholesterol absorption in wild-type and NPC− mice
To investigate the role of NPC1 and NPC2 in intestinal cholesterol uptake (Scheme 1, step 3), the fractional absorption of [4-14C]cholesterol was measured in wild-type, NPC1− and NPC2− mice using [5,6-3H]sitostanol as a non-absorbed sterol control. All three groups of animals exhibited similar fractional [4-14C]cholesterol absorption with the 0.01 mg dose (59, 59 and 54% for wild-type, NPC1− and NPC2−respectively) (Figure 1, white bars). The mutant animals exhibited a minor decrease in fractional [4-14C]cholesterol absorption with the 0.11 mg dose (54, 45 and 47% for wild-type, NPC1− and NPC2− respectively) (Figure 1, hatched bars) that did not reach statistical significance (P>0.2; analysis of variance).
Figure 1. Fractional cholesterol absorption in wild-type and NPC− mutant mice.

Fractional absorption of orally gavaged [4-14C]cholesterol was measured using the fecal dual-isotope technique in wild-type (WT), NPC1− (NPC1) and NPC2− (NPC2) mice, either untreated or treated orally with 10 mg of ezetimibe/kg of body weight (+EZ). The absorption was measured at two different cholesterol levels in the dosing mixture: 0.01 mg cholesterol (white bars); 0.11 mg of cholesterol (hatched bars). Results are means±S.E.M. for each genotype set, which typically included three or four animals. ANOVA did not reveal any significant genotype-specific effect on fractional cholesterol absorption (P>0.2), whereas ezetimibe treatment elicited a strong decrease (P<0.0001 for all genotypes).
Sensitivity to ezetimibe
To ensure that the observed fractional cholesterol absorption in the mutant mice is indeed taking place via the NPC1L1-mediated pathway, we tested sensitivity to ezetimibe treatment. When gavaged with 0.11 mg of cholesterol, the wild-type, NPC1− and NPC2− mice absorbed 78, 71 and 69% less cholesterol respectively in the presence of ezetimibe compared with absorption in the absence of ezetimibe (ezetimibe effect; P<0.0001 for all genotypes). Treatment of mice with ezetimibe had more dramatic effects on the wild-type and mutant mice at the lower cholesterol dose (Figure 1).
Acute cholesterol uptake and absorption in wild-type and NPC− mice
We tested the possibility that later steps in intestinal cholesterol absorption (Scheme 1, beyond step 3) are affected by alterations in NPC1 or NPC2 function by determining acute uptake of [4-14C]cholesterol and [9,10-3H (n)]trioleoylglycerol following gavage. Trioleoylglycerol (triolein) was used as a control to verify the functionality of the chylomicron assembly process in enterocytes. Acute uptake was measured as the total labelled lipids present within the intestine, liver and serum. In these experiments, labelled lipids present in the liver and serum together reflect absorption out of the intestine. The average acute uptake of [4-14C]cholesterol relative to the gavaged dose was ∼20% in all three groups 2 h after gavage (Figure 2A). At this time, 90% of the [4-14C]-cholesterol taken up was still associated with the intestine, with 10% being absorbed out of the intestine into the liver and serum for all three genotypes (Figures 2A and 2B, crossed-hatched bars). The total uptake of [9,10-3H (n)]trioleoylglycerol and its distribution in the intestine, liver and serum of the NPC mice were equivalent to that seen in the wild-type mice (Figures 2A and 2B, hatched bars). Similar measurements were also performed 4 h after gavage to allow for greater uptake of [4-14C]cholesterol by the intestine (∼30%) and to allow more time for transport of cholesterol out of the intestine. In this case, the liver and serum absorbed ∼20% of the [4-14C]cholesterol taken up, with all three genotypes exhibiting similar distributions (results not shown). These results indicate that neither NPC1 nor NPC2, nor by implication the lysosomal cholesterol transport system, are essential for the subsequent steps involved in cholesterol transport within enterocytes and chylomicron secretion.
Figure 2. Acute absorption of radiolabelled lipids in the liver and serum of wild-type and NPC− mutant mice.

A dose containing [4-14C]cholesterol (crossed-hatched bars) and [9,10-3H (n)]trioleoylglycerol (‘Triolein’, hatched bars) was orally gavaged into wild-type (WT), NPC1-deficient (NPC1) and NPC2-deficient (NPC2) mice. (A) After 2 h, lipids were measured in the intestine, liver and serum (results are means±S.E.M.). (B) Absorbed lipids were calculated as the percentage absorption out of the intestine (liver+serum) relative to total uptake (intestine+liver+serum). Error bars represent S.E.M. for each genotype set, which typically included four or five animals. Analysis of variance did not reveal significant differences among the genotypes (P>0.1).
DISCUSSION
NPC2 is not required for NPC1L1-mediated cholesterol uptake
In a previous study, NPC1L1− mice took up only ∼16% of the orally gavaged cholesterol, representing a reduction of ∼69% compared with wild-type mice [2]. Unlike NPC1L1− mice, NPC2− mice showed fractional cholesterol absorption similar to that in wild-type mice, suggesting that NPC2 does not participate in the NPC1L1-mediated uptake pathway. However, to rule out the possibility that the uptake observed in NPC2− mice was a consequence of a compensatory pathway that might counterbalance a decrease in the NPC1L1-mediated pathway, we examined the sensitivity of cholesterol uptake to ezetimibe, a drug that lowers intestinal cholesterol uptake by inhibiting NPC1L1. NPC2− mice exhibited the same sensitivity as wild-type mice to orally administered ezetimibe. Thus the fractional absorption observed in NPC2− mice proceeds through the NPC1L1-mediated pathway.
The data indicate that, unlike NPC1-mediated cholesterol transport in lysosomes, NPC1L1-mediated cholesterol uptake in the intestine does not require NPC2. As NPC2 does not display significant sequence similarity to any other human expressed sequence tag in the annotated human genome, there are no candidate NPC2-like partners for NPC1L1. These mechanistic differences in NPC1 and NPC1L1 function presumably reflect either differences in the presentation of hydrophobic ligands in the lysosomal and intestinal lumen (e.g. the presence of bile salts in the intestine), intrinsic differences in the membranes in which the proteins and/or sterol ligands reside, or fundamental mechanistic differences in the proteins themselves.
NPC1 deficiency does not impair cholesterol uptake
Fractional cholesterol absorption in NPC1− mice was similar to that in wild-type mice at the 0.01 mg cholesterol dose and was slightly decreased at the 0.11 mg dose (a decrease of 17%). A previous study of NPC1− mice reported a decrease of about 40% in net cholesterol absorption – a combined effect of total food intake per kg of body weight per day and fractional cholesterol absorption in the intestine [21]. The observed decrease in net absorption of cholesterol in NPC1− mice can be accounted for by the increase in biliary cholesterol in the intestine [22] and the decreased food intake. Thus our data are in agreement with those obtained in the previous study, and both indicate that NPC1 deficiency does not significantly affect intestinal cholesterol uptake.
The lack of an effect on cholesterol uptake in NPC1− mice is supported by our observation that ezetimibe treatment led to a decrease in fractional cholesterol absorption in these mice similar to that seen in wild-type mice. Furthermore, it has been recently reported that the total hepatic content of cholesterol in NPC1− mice is inversely proportional to the dose of ezetimibe administered [23], suggesting that, in NPC1− mice, intestinal cholesterol uptake occurs via an NPC1L1-mediated pathway and that there is no compensatory up-regulation of an NPC1L1-independent pathway. This is consistent with our data, which indicate that a block in the endosomal/lysosomal cholesterol pathway that involves NPC1 and NPC2 does not lead to a significant change in intestinal cholesterol uptake. It is also noteworthy that, in spite of similarities in sequences and presumed molecular activities of NPC1 and NPC1L1, deficiency in NPC1 does not appear to alter the functional activity of NPC1L1, implying their independent roles and regulation.
Cholesterol metabolism differs in mice and fruitflies
A deficiency of NPC1L1 (or dNPC1b) has similar consequences in mice [2] and fruitflies [15]. In both organisms, deficiency of NPC1L1 results in decreased cholesterol uptake (Table 1). However, deficiency of NPC1 (or dNPC1a) has different consequences in mice and fruitflies (Table 1). We found that deficiency of NPC1 alone causes little change in cholesterol uptake in mice, but in combination with pharmacological inhibition of NPC1L1 leads to a decrease in intestinal cholesterol uptake. In contrast, in fruitflies, a deficiency of dNPC1a alone, or in combination with the dNPC1b deficiency, led to a significant increase in the efficiency of cholesterol uptake above the respective background uptake levels [15] (see Table 1), suggesting activation of a dNPC1b-independent cholesterol uptake pathway in dNPC1a− fruitflies.
Table 1. Comparison of cholesterol metabolism in mice and fruitflies.
Data sources were as follows: NPC1L1− mice [2,19]; NPC1− mice: the present study and [24–26]; dNPC1b− and dNPC1a− fruitflies: [15]. Abbreviation: N.D., not determined.
| Functional protein | Cholesterol uptake | Cholesterol biosynthesis | |||
|---|---|---|---|---|---|
| NPC1L1 (dNPC1b) | NPC1 (dNPC1a) | Mouse | Fruitfly | Mouse | Fruifly |
| + | + | Normal | Normal | Normal | None |
| − | + | ↓↓↓ | ↓↓↓ | ↑↑ | None |
| + | − | Normal | ↑↑ | ↑↑ | None |
| − | − | ↓↓↓* | ↓ | N.D. | None |
* Pharmacological inhibition of NPC1L1 using ezetimibe.
The NPC1L1 (dNPC1b)-independent compensatory pathway postulated for fruit flies does not appear to exist in mice. The dNPC1b-independent pathway has perhaps evolved to maintain cholesterol homoeostasis in insects which, unlike mammals, lack the ability for de novo cholesterol biosynthesis. In fact, the rates of cholesterol biosynthesis have been found to be higher in most organs in NPC1− mice [24–26]. Such fundamental differences in cholesterol metabolism need to be considered when drawing parallels between insect and mammalian cholesterol uptake pathways. More generally, these observations illustrate that deficiencies in orthologous proteins can produce different phenotypes in different experimental systems as a consequence of tissue- or organism-specific compensatory pathways.
Acknowledgments
We thank Dr Margaret van Heek and Dr Harry Davis (Schering-Plough Corporation, Kenilworth, NJ, U.S.A.) and Dr G. S. Tint (Medical Service, Veterans Affairs Medical Center, East Orange, NJ, U.S.A.) for invaluable help in designing the gavage experiments, Dr John Dietschy and Dr Stephen Turley (University of Texas Southwestern Medical School, Dallas, TX, U.S.A.) and Dr Dirk F. Moore (Department of Biostatistics, School of Public Health–UMDNJ, Piscataway, NJ, U.S.A.) for helpful discussions, and Mukarram El-Banna, Ling-ling Wang and Amy Huang (UMDNJ–RWJMS) for help with the gavage procedure. This research was supported by the Ara Parseghian Medical Research Foundation (Tucson, AZ, U.S.A.). A. M. S. is an Investigator of the Howard Hughes Medical Institute.
References
- 1.Wang D. Q. Regulation of intestinal cholesterol absorption. Annu. Rev. Physiol. 2007;69:221–248. doi: 10.1146/annurev.physiol.69.031905.160725. [DOI] [PubMed] [Google Scholar]
- 2.Altmann S. W., Davis H. R., Jr, Zhu L. J., Yao X., Hoos L. M., Tetzloff G., Iyer S. P., Maguire M., Golovko A., Zeng M., et al. Niemann–Pick C1 like 1 protein is critical for intestinal cholesterol absorption. Science. 2004;303:1201–1204. doi: 10.1126/science.1093131. [DOI] [PubMed] [Google Scholar]
- 3.Iyer S. P. N., Yao X., Crona J. H., Hoos L. M., Tetzloff G., Davis J. H. R., Graziano M. P., Altmann S. W. Characterization of the putative native and recombinant rat sterol transporter Niemann–Pick C1 like 1 (NPC1L1) protein. Biochim. Biophys. Acta. 2005;1722:282–292. doi: 10.1016/j.bbagen.2004.12.021. [DOI] [PubMed] [Google Scholar]
- 4.Van Heek M., France C. F., Compton D. S., McLeod R. L., Yumibe N. P., Alton K. B., Sybertz E. J., Davis H. R., Jr In vivo metabolism-based discovery of a potent cholesterol absorption inhibitor, SCH58235, in the rat and rhesus monkey through the identification of the active metabolites of SCH48461. J. Pharmacol. Exp. Ther. 1997;283:157–163. [PubMed] [Google Scholar]
- 5.Garcia-Calvo M., Lisnock J., Bull H. G., Hawes B. E., Burnett D. A., Braun M. P., Crona J. H., Davis H. R., Jr, Dean D. C., Detmers P. A., et al. The target of ezetimibe is Niemann–Pick C1-like 1 (NPC1L1) Proc. Natl. Acad. Sci. U.S.A. 2005;102:8132–8137. doi: 10.1073/pnas.0500269102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sleat D. E., Wiseman J. A., El-Banna M., Price S. M., Verot L., Shen M. M., Tint G. S., Vanier M. T., Walkley S. U., Lobel P. Genetic evidence for nonredundant functional cooperativity between NPC1 and NPC2 in lipid transport. Proc. Natl. Acad. Sci. U.S.A. 2004;101:5886–5891. doi: 10.1073/pnas.0308456101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Carstea E. D., Morris J. A., Coleman K. G., Loftus S. K., Zhang D., Cummings C., Gu J., Rosenfeld M. A., Pavan W. J., Krizman D. B., et al. Niemann–Pick C1 disease gene: homology to mediators of cholesterol homeostasis. Science. 1997;277:228–231. doi: 10.1126/science.277.5323.228. [DOI] [PubMed] [Google Scholar]
- 8.Naureckiene S., Sleat D. E., Lackland H., Fensom A., Vanier M. T., Wattiaux R., Jadot M., Lobel P. Identification of HE1 as the second gene of Niemann–Pick C disease. Science. 2000;290:2298–2301. doi: 10.1126/science.290.5500.2298. [DOI] [PubMed] [Google Scholar]
- 9.Liou H. L., Dixit S. S., Xu S., Tint G. S., Stock A. M., Lobel P. NPC2, the protein deficient in Niemann–Pick C2 disease, consists of multiple glycoforms that bind a variety of sterols. J. Biol. Chem. 2006;281:36710–36723. doi: 10.1074/jbc.M608743200. [DOI] [PubMed] [Google Scholar]
- 10.Baker C. S., Magargee S. F., Hammerstedt R. H. Cholesterol transfer proteins from ram cauda epidydimal and seminal plasma. Biol. Reprod. Suppl. 1993;48:86. [Google Scholar]
- 11.Cheruku S. R., Xu Z., Dutia R., Lobel P., Storch J. Mechanism of cholesterol transfer from the Niemann–Pick type C2 protein to model membranes supports a role in lysosomal cholesterol transport. J. Biol. Chem. 2006;281:31594–31604. doi: 10.1074/jbc.M602765200. [DOI] [PubMed] [Google Scholar]
- 12.Kirchhoff C., Osterhoff C., Young L. Molecular cloning and characterization of HE1, a major secretory protein of the human epididymis. Biol. Reprod. 1996;54:847–856. doi: 10.1095/biolreprod54.4.847. [DOI] [PubMed] [Google Scholar]
- 13.Larsen L. B., Ravn P., Boisen A., Berglund L., Petersen T. E. Primary structure of EPV20, a secretory glycoprotein containing a previously uncharacterized type of domain. Eur. J. Biochem. 1997;243:437–441. doi: 10.1111/j.1432-1033.1997.0437a.x. [DOI] [PubMed] [Google Scholar]
- 14.Klein A., Amigo L., Retamal M. J., Morales M. G., Miquel J. F., Rigotti A., Zanlungo S. NPC2 is expressed in human and murine liver and secreted into bile: potential implications for body cholesterol homeostasis. Hepatology. 2006;43:126–133. doi: 10.1002/hep.20985. [DOI] [PubMed] [Google Scholar]
- 15.Voght S. P., Fluegel M. L., Andrews L. A., Pallanck L. J. Drosophila NPC1b promotes an early step in sterol absorption from the midgut epithelium. Cell Metab. 2007;5:195–205. doi: 10.1016/j.cmet.2007.01.011. [DOI] [PubMed] [Google Scholar]
- 16.Ioannou Y. A. Niemann–Pick C proteins in sterol transport and absorption: flies in the ointment. Dev. Cell. 2007;12:481–483. doi: 10.1016/j.devcel.2007.03.017. [DOI] [PubMed] [Google Scholar]
- 17.Loftus S. K., Morris J. A., Carstea E. D., Gu J. Z., Cummings C., Brown A., Ellison J., Ohno K., Rosenfeld M. A., Tagle D. A., et al. Murine model of Niemann–Pick C disease: mutation in a cholesterol homeostasis gene. Science. 1997;277:232–235. doi: 10.1126/science.277.5323.232. [DOI] [PubMed] [Google Scholar]
- 18.Wang D. Q. H., Carey M. C. Measurement of intestinal cholesterol absorption by plasma and fecal dual-isotope ratio, mass balance, and lymph fistula methods in the mouse: an analysis of direct versus indirect methodologies. J. Lipid Res. 2003;44:1042–1059. doi: 10.1194/jlr.D200041-JLR200. [DOI] [PubMed] [Google Scholar]
- 19.Davis H. R., Jr, Zhu L. J., Hoos L. M., Tetzloff G., Maguire M., Liu J., Yao X., Iyer S. P., Lam M. H., Lund E. G., et al. Niemann-Pick C1 like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 2004;279:33586–33592. doi: 10.1074/jbc.M405817200. [DOI] [PubMed] [Google Scholar]
- 20.Folch J., Lees M., Stanley G. H. S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957;226:497–509. [PubMed] [Google Scholar]
- 21.Xie C., Turley S. D., Dietschy J. M. Cholesterol accumulation in tissues of the Niemann–Pick type C mouse is determined by the rate of lipoprotein-cholesterol uptake through the coated-pit pathway in each organ. Proc. Natl. Acad. Sci. U.S.A. 1999;96:11992–11997. doi: 10.1073/pnas.96.21.11992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Amigo L., Mendoza H., Castro J., Quinones V., Miquel J. F., Zanlungo S. Relevance of Niemann–Pick type C1 protein expression in controlling plasma cholesterol and biliary lipid secretion in mice. Hepatology. 2002;36:819–828. doi: 10.1053/jhep.2002.35617. [DOI] [PubMed] [Google Scholar]
- 23.Beltroy E. P., Liu B., Dietschy J. M., Turley S. D. Lysosomal unesterified cholesterol content correlates with liver cell death in murine Niemann–Pick type C disease. J. Lipid Res. 2007;48:869–881. doi: 10.1194/jlr.M600488-JLR200. [DOI] [PubMed] [Google Scholar]
- 24.Xie C., Turley S. D., Pentchev P. G., Dietschy J. M. Cholesterol balance and metabolism in mice with loss of function of Niemann–Pick C protein. Am. J. Physiol. Endocrinol. Metab. 1999;276:E336–E344. doi: 10.1152/ajpendo.1999.276.2.E336. [DOI] [PubMed] [Google Scholar]
- 25.Pentchev P. G., Boothe A. D., Kruth H. S., Weintroub H., Stivers J., Brady R. O. A genetic storage disorder in BALB/C mice with a metabolic block in esterification of exogenous cholesterol. J. Biol. Chem. 1984;259:5784–5791. [PubMed] [Google Scholar]
- 26.Liscum L., Ruggiero R. M., Faust J. R. The intracellular transport of low density lipoprotein-derived cholesterol is defective in Niemann–Pick type C fibroblasts. J. Cell Biol. 1989;108:1625–1636. doi: 10.1083/jcb.108.5.1625. [DOI] [PMC free article] [PubMed] [Google Scholar]