

Despite the recent evidence of population-level handedness in nonhuman primates, continued debate persists over the analysis and interpretation of findings on handedness, particularly in great apes (Hopkins, 1999; McGrew and Marchant, 1997). Palmer (2002) recently questioned the analysis and interpretation of findings on hand preference in nonhuman primates, and in particular, results suggesting population-level right-handedness in chimpanzees. Specifically, Palmer (2002) reanalyzed hand-preference data for bimanual feeding in chimpanzees reported by Hopkins (1994) and argued, based on his analysis using funnel-graphs, that there were unusual patterns in the data that raised questions on the validity of the results for these data. With respect to the findings reported by Hopkins (1994), Palmer (2002) argued that a fundamental problem with the data was in the relationship between the number of individual data points collected for each subject and the distribution of hand preferences in the sample. Palmer (2002) quartiled the sample (N = 140) on the basis of the total number of observations used to derive the individual handedness scores (see his Table 1), and showed that as the number of observation increased, the number of ambiguously handed subjects increased. According to Palmer, this pattern of results should not have been evident under basic statistical assumptions of increasing effects with increasing sample size. In short, the data regressed toward a mean of zero, or no hand preference, with increasing sample size. Palmer (2002) also created funnel graphs of individual variation in percent right-hand use as a function of sample size, and argued that the patterns did not conform to patterns that would normally reflect nonsampling error in the data of Hopkins (1994). Lastly, Palmer (2002) applied funnel-graph analysis to 14 selective studies on hand preference in chimpanzees, and argued that most studies found no evidence of population-level handedness.
TABLE 1. Distribution of hand preference as a function of sex and rearing history.
| Hand preference | |||
|---|---|---|---|
| Left | Ambiguous | Right | |
| Sex | |||
| Males | 12 | 13 | 34 |
| Females | 17 | 19 | 37 |
| Rearing history | |||
| Mother | 14 | 19 | 38 |
| Nursery | 15 | 13 | 33 |
With respect to the data of Hopkins (1994), Palmer (2002) fairly and representatively indicated the inherent problems with the variation in percent right-hand use as a function of sample size. Our main response to this criticism is that the bulk of the youngest subjects make up the group comprising the largest N and most ambiguously handed subjects. In other words, sample size is confounded by subject age, and it is clearly legitimate to assume that there may be age differences in the expression and magnitude of hand preference in chimpanzees. If the youngest subjects are removed from the analysis, one-sample t-tests still reveal population-level right-handedness for bimanual feeding. The effect is not terribly robust, and this is largely due to the fact that there are so many ambiguously handed subjects. The fact that there were so many ambiguously handed subjects was clearly noted as a limitation of this measure in Hopkins (1994). Further, in Hopkins (1994), it is emphasized that measures involving coordinated actions of the hands may better elicit population-level hand preferences in chimpanzees than the bimanual feeding measure employed in that study.
Although we recognize the limitations of the findings by Hopkins (1994), we do not believe that all studies of chimpanzee handedness are subject to the same criticism, and in this paper we report on evidence that contradicts the conclusions of Palmer (2002).
Specifically, we developed a measure of hand preference for coordinated bimanual actions, referred to as the TUBE task (Hopkins, 1995; Hopkins et al., 2001). This measure of hand use also elicits a population-level hand preference in captive chimpanzees, but unlike the feeding task used by Hopkins (1994), the majority (90%) of chimpanzees show a significant hand preference (either left or right). Palmer (2002, personal communication) has been equally critical of these data, in terms of the application of funnel plots to the distribution of hand preference in relation to sample size.
To address this issue of whether individual differences in sample size influence the distribution of hand preference in chimpanzees, we conducted an experiment in which we assessed hand preference for the TUBE task. This experiment differed from previous studies in that the exact same number of observations was collected in each subject. If the degree of hand preference found in chimpanzees is determined by sampling biases associated with N, then no population-level hand preferences should be found for this measure when sample sizes are identical for all subjects. In contrast, if the reported effects of population-level right handedness are consistent and not influenced by sample size, then population-level right-handedness should be found for this measure of hand use in this sample of chimpanzees.
Method
Subjects
Subjects were 132 chimpanzees housed at the Yerkes Regional Primate Research Center (YRPRC) of Emory University. There were 73 females and 59 males, and subjects ranged in age from 4–47 years (mean, 15.51 years; SD, 10.02). Within the female sample, there were 49 mother- and 24 nursery-reared subjects. For the male sample, there were 22 mother- and 37 nursery-reared subjects. Mother-reared chimpanzees were those reared by their biological, conspecific mother for more than 30 days of life. Nursery-reared subjects were those which were brought to the YRPRC nursery before 31 days of life. The standard protocol for nursery-rearing chimpanzees was described in detail elsewhere (Bard, 1996). All chimpanzees lived in social groups ranging from 2–16 individuals. Cage sizes varied according to group size, but all cages met minimum standards as required by federal regulations.
Procedure
The procedure for the TUBE task was described in detail elsewhere (Hopkins, 1995; Hopkins et al., 2001). Briefly, peanut butter is smeared on the inside edge of poly-vinyl-chloride (PVC) tubes approximately 15 cm in length and 2.5 cm in diameter. Peanut butter is smeared on both ends of the PVC pipe, and is placed far enough down the tube that the chimpanzees cannot lick the contents completely off with their mouths, but rather must use their fingers to remove the substrate. The PVC tubes were handed to the subjects in their home cages, and focal sampling was used to collect individual data from each subject. The hand used by the experimenters to give the PVC to the subjects was randomized across subjects and tests. During each test, each time the subject used its finger to extract the peanut butter, the hand used was recorded as either right or left by the experimenter. Each subject was tested on four occasions separated by at least 10 min, but no longer than 1 day. During each test, the first 20 hand-use responses were recorded for a total of exactly 80 observations per subject. A 5–10-min interval separated each test session, during which time the PVC pipes were retrieved from the chimpanzees, cleaned, and refilled with peanut butter.
Data analysis
Hand preferences were characterized in two ways. First, a handedness index (HI) was calculated for the overall frequency following the formula HI = (#R − #L)/(80). Positive HI values reflected right-hand biases, and negative HI values reflected left-hand biases. The absolute value of the handedness score reflected the magnitude of hand preference. Second, based on the total left and right hand bouts and frequencies, z-scores were used to evaluate whether the hand preferences of individual subjects deviated significantly from chance. This is the procedure most frequently used in the nonhuman primate literature (see Hopkins, 1999). Subjects with z-scores greater than 1.95 or less than −1.95 were classified as right- and left-handed, respectively. All other subjects were classified as ambiguously handed. For all analyses, alpha was set at P < 0.05.
Results
Descriptive statistics
Based on the individual z-scores, there were 29 left-handed, 32 ambiguously handed, and 71 right-handed subjects, a distribution that differs significantly from chance as revealed by chi-square goodness-of-fit test: χ2(2, N = 132) = 24.96, P < 0.001. The number of right-handed chimpanzees was significantly higher than the number of left-handed (χ2(1, N = 100) = 17.64, P < 0.001) and ambiguously-handed (χ2(1, N = 103) = 14.77, P < 0.001) subjects. Log linear analysis of the effects of rearing history and sex on hand preference failed to reveal any significant interactions. The distribution of hand preference as a function of sex and rearing history is shown in Table 1. Using HI scores rather than hand preference classifications, the analyses revealed similar findings. One-sample t-tests revealed that the mean HI score (mean = 0.223) was significantly different from zero and skewed to the right (t(131) = 4.52, P < 0.001). A factorial analysis of variance, with sex and rearing history serving as between-group variables, failed to reveal any significant main effects of interactions. A Pearson product moment correlation between age and HI scores was not statistically significant (r = 0.08, df = 130, n.s.).
Consistency within and between studies
To evaluate the consistency in hand use within individuals and between tests, several additional analyses were performed. Accordingly, handedness indices were calculated based on the 20 responses recorded during each of the four handedness tests (referred to as HI1, HI2, HI3, and HI4, respectively). Pearson product moment correlation analysis indicated significant positive correlations between handedness index scores for each of the test sessions (see Table 2). HI scores were also compared using a repeated-measures analysis of variance, and this failed to reveal a statistically significant effect of test session on handedness. However, one-sample t-tests revealed significant population-level right handedness for HI1 (t(131) = 3.47, P < 0.001), HI2 (t(131) = 3.34, P < 0.001), HI3 (t(131) = 3.97, P < 0.001), and HI4 (t(131) = 4.34, P < 0.001). These results are significant even after adjusting alpha to guard against type I error due to the number of analyses performed on the data. The mean handedness scores for each test session can be seen in Figure 1. Lastly, the overall handedness scores for this study were correlated with the individual handedness scores from Hopkins et al. (2001), in order to evaluate consistency between studies. A total of 98 chimpanzees was used in both studies, and a significant positive correlation was found between the handedness scores for the two studies (r = 0.67, df = 96, P < 0.001).
TABLE 2. Intercorrelations in hand use between tests.
P < 0.001, N = 132.
Fig. 1.
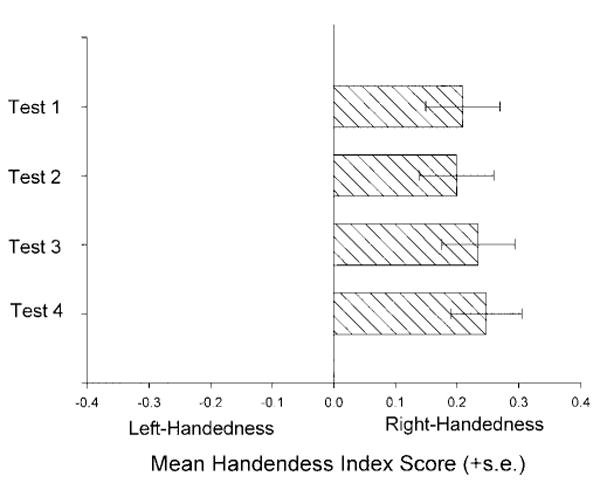
Mean handedness index scores for each of four test sessions.
Discussion
The results of this experiment are straightforward. In a sample of 132 chimpanzees, population-level right-handedness was found for a task measuring coordinated bimanual actions. Neither sex nor rearing history significantly influenced hand preference. These results cannot be attributed to variation in individual sample size, and therefore do not support the contention by Palmer (2002) that previously published hand-preference data in chimpanzees are flawed because unequal numbers of observations were made in all subjects. The application of funnel graphs to hand-preference data seems appropriate, but only when there are unequal sample sizes in the number of observations used to evaluate individual hand preferences. With equal sample sizes, funnel graphs are not necessary, and investigators should be encouraged to use either approach, when necessary.
The evidence of population-level right-handedness in this study for the TUBE task is consistent with results from two previous studies (Hopkins, 1995; Hopkins et al., 2001), despite the fact that the methods of data collection and test procedures were different. In Hopkins (1995), chimpanzees were tested twice, and hand-use data were collected until the subjects dropped the PVC pipe or lost interest in the food. In Hopkins et al. (2001), chimpanzees were tested on four occasions, and on each of two occasions, we intentionally tried to bias hand use by making subjects take the tube with each hand equally often. In Hopkins et al. (2001), bouts of hand use were also scored in addition to frequency of hand use, with handedness scores being nearly identical for two measures. In the present study, subjects were tested four times, but only the first 20 responses were recorded during each test, and the total frequency of hand use was recorded. For all three studies, population-level right-handedness was found. Moreover, variation in test-retest correlations was not different for tests administered 10 min apart compared to 1 year apart or even 6 years apart (for 6-year correlation coefficients, see Hopkins et al., 2001). Collectively, these results indicate that chimpanzees housed at the YRPRC are consistently right-handed for a task measuring coordinated bimanual actions.
Acknowledgments
We appreciate the assistance of Michael J. Wesley and Autumn Hostetter in data collection.
Grant sponsor: NIH; Grant numbers: RR-00165, NS-29574, NS-36605, HD-38051.
Literature Cited
- Bard KA. Responsive care: behavioral intervention for nursery-reared chimpanzees. Ridgefield, CT: Jane Goodall Institute; 1996. [Google Scholar]
- Hopkins WD. Hand preferences for bimanual feeding in 140 captive chimpanzees (Pan troglodytes): rearing and ontogenetic factors. Dev Psychobiol. 1994;27:395–407. doi: 10.1002/dev.420270607. [DOI] [PubMed] [Google Scholar]
- Hopkins WD. Hand preferences for a coordinated bimanual task in 110 chimpanzees (Pan troglodytes): cross-sectional analysis. J Comp Psychol. 1995;105:178–190. doi: 10.1037/0735-7036.109.3.291. [DOI] [PubMed] [Google Scholar]
- Hopkins WD. On the other hand: statistical issues in the assessment and interpretation of hand preference data in nonhuman primates. Int J Primatol. 1999;20:851–866. [Google Scholar]
- Hopkins WD, Fernanadez-Carriba S, Wesley MJ, Hostetter A, Pilcher D, Poss S. The use of bouts and frequencies in the evaluation of hand preferences for a coordinated bimanual task in chimpanzees (Pan troglodytes): an empirical study comparing two different indices of laterality. J Comp Psychol. 2001;115:294–299. doi: 10.1037//0735-7036.115.3.294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGrew WC, Marchant LF. On the other hand: current issues in and meta-analysis of the behavioral laterality of hand function in nonhuman primates. Yrbk Phys Anthropol. 1997;40:201–232. [Google Scholar]
- Palmer AR. Chimpanzee right-handedness reconsidered: evaluating the evidence with funnel plots. Am J Phys Anthropol. 2002;118:191–199. doi: 10.1002/ajpa.10063. [DOI] [PubMed] [Google Scholar]