

Abstract
The coat protein (CP) of alfalfa mosaic virus was used as a carrier molecule to express antigenic peptides from rabies virus and HIV. The antigens were separately cloned into the reading frame of alfalfa mosaic virus CP and placed under the control of the subgenomic promoter of tobacco mosaic virus CP in the 30BRz vector. The in vitro transcripts of recombinant virus with sequences encoding the antigenic peptides were synthesized from DNA constructs and used to inoculate tobacco plants. The plant-produced protein (virus particles) was purified and used for immunization of mice. Both antigens elicited specific virus-neutralizing antibodies in immunized mice.
Recent studies have shown that plants and plant viruses can be effective tools for antigen production and delivery (1–4). Plants offer several advantages for the production of vaccine antigens and other therapeutic proteins, including lack of contamination with animal pathogens, relative ease of genetic manipulation, eukaryotic protein modification machinery, and economical production. Like liposomes and microcapsules, plant cells and plant viruses are expected to provide natural protection for the passage of antigens through the gastrointestinal tract.
A variety of important protein antigens have been expressed in transgenic plants, including hepatitis B surface antigen (1), Escherichia coli heat-labile enterotoxin (2), rabies virus glycoprotein (3), and Norwalk virus capsid protein (4). However, the level of antigenic protein produced by transgenic plants is relatively low, suggesting the need for new approaches to express foreign proteins in plants.
One approach is to engineer virus coat proteins (CPs) to function as carrier molecules for fused antigenic peptides. Such carrier proteins may have the potential to self-assemble and form recombinant virus particles displaying the desired antigenic epitopes on their surfaces. A commonly used carrier molecule is the CP of viruses that infect bacteria (5–7), animals (8–10), and plants (11–16). The CP of tobacco mosaic virus (TMV) was among the first plant virus proteins to be used as a carrier molecule for antigenic epitopes from other sources (11). Fitchen et al. (14) and McLain et al. (15) showed that antigens produced in plants as a result of infection with engineered TMV and cowpea mosaic viruses can elicit specific antibodies when injected into mice. A limitation of this approach is the failure of virus assembly when even moderately sized peptides are incorporated into the CP. Only peptides less than 25 amino acids in length have been successfully introduced into the TMV CP. This is a significant limitation for the production of various molecules of biomedical importance using plant viruses. For example, a chimeric peptide antigen that successfully protects mice against rabies virus and is therefore suitable for the use in a plant based vaccine has been described (17). However, this peptide is 38 amino acids long, far in excess of capacity of TMV CP. We have overcome this size limitation by using the CP of alfalfa mosaic virus (AIMV), which can accommodate larger peptide sequences. The CP of AIMV forms particles of different sizes (20–60 nm) and shapes (spherical, ellipsoid, and bacilliform), depending on the length of the encapsidated RNA. The ability to form particles of varying sizes and shapes reflects the flexibility of the AIMV CP in protein–protein interactions. The N terminus of the AIMV CP is located on the surface of the virus particles and does not appear to interfere with virus assembly (18). We have shown previously that the AIMV CP with an additional 37-amino acid peptide at its N terminus forms particles in vitro and retains biological activity (19). These characteristics make the AIMV CP an excellent candidate as a carrier molecule for the expression of foreign antigens.
In the present study we have engineered two constructs containing the antigenic peptide from rabies virus and from HIV-1, respectively, fused them with the AIMV CP and cloned them into the 30BRz vector for expression in virus-infected plants. The plants were inoculated with chimeric TMV transcripts, synthesized from engineered 30BRz containing nucleotide sequences that encode for recombinant AIMV CP. Virus particles formed from the recombinant AIMV CP were purified from the infected plant tissue and used to immunize mice resulting in a antigen specific humoral immune response, with virus-neutralizing antibodies.
MATERIALS AND METHODS
DNA Constructs.
All cloning and cell transformations were performed according to Sambrook et al. (20). E. coli DH5α competent cells (Life Technologies, Gaithersburg, MD) were used for transformation. To clone the V3 loop of HIV-1, a plasmid DNA containing sequences of the HIV-1 MN isolate (21) was used as a template for the PCR, performed using 5′-AGATCTCGAGATGAGTTCATCTGTAGAAATTAATTGTACA-3′ as the first-strand and 5′-CGGCTCGAGCTACTAATGTTACAATG-3′ as the second-strand primers. Antigenic peptide (Drg24) containing B-cell epitope from rabies glycoprotein (G5-24) and a T cell epitope from rabies nucleoprotein (31D) was engineered using overlapping oligonucleotides based on the published amino acid sequences (refs. 17 and 22; Fig. 1). First- and second-strand primers, respectively, were 5′-GCGCTCGAGATGTCCGCCGTCTACACCCGAATTATGATGAACGGAGGACGACTTAAGCGACCACCAGACCAGCTTG-3′ and 5′-GCGCTCGAGTCCTCTTCCACCACAAGGTGCTCATTTTCGTCGGATCGGAAGTCGTGAAGGTTCACAAGCTGGTCTGGTGGTCGCTTAAGTCGTCC-3′. The TMV-based DNA plasmid vector 30BRz (Fig. 1) was used for the expression of chimeric genes. 30BRz (a gift of William Dawson, Florida State University, Gainsville) contains the TMV genome and is similar to the TMV vector TB2 (23), except that 30BRz contains multiple cloning sites for subcloning genes under the control of the TMV CP subgenomic promoter as well as a ribozyme for self-cleavage of in vitro RNA transcripts (W. Dawson, personal communication).
Figure 1.
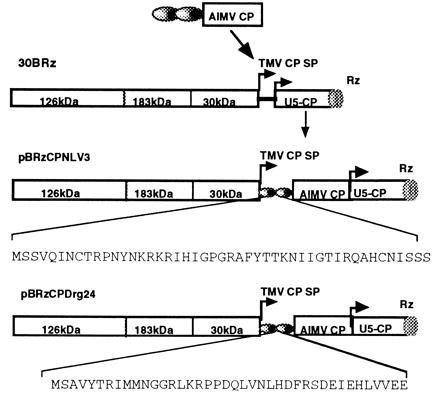
Schematic representation of the genome of 30BRz (derivative of TMV) and the cloning strategy: the 126- and 183-kDa proteins are required for the TMV replication, 30-kDa protein is the viral movement protein, and CP is viral CP. The arrow under TMV CP SP indicates the subgenomic promoter of TMV CP. The two ellipsoids indicates peptides fused to the AIMV CP. Rz, ribozyme for self-cleavage. pBRzCPMNV3 and pBRzCPDrg24 are viruses engineered to express antigenic peptides representing V3 loop of HIV-1 named MNV3 or chimeric epitope of rabies virus named Drg24, respectively. The amino acid sequence of inserted region of each peptide is shown under the linear map of plasmid.
In Vitro Transcription.
In vitro transcripts of recombinant TMV were synthesized using T7 RNA polymerase (Promega) and CsCl-purified plasmid DNA, according to the manufacturer’s guidelines. Transcripts were capped using the RNA cap structure analog m7G(5)ppp(5)G (Biolabs, Northbrook, IL).
Western Blot Analysis.
Recombinant proteins produced in virus-infected plants were analyzed by Western blot (24). Proteins from purified virus particles were separated electrophoretically on SDS/polyacrylamide gels and electroblotted onto a nylon membrane overnight at 33 mA. After blocking with milk (Kirkegaard & Perry), proteins were allowed to react with appropriate antibodies and detected using a Vectastain ABC kit (Vector Laboratories). The recombinant proteins were detected using antibodies specific for: the AIMV CP (25); the linear epitope (G5-24) of rabies glycoprotein (22); and the V3 loop of HIV-1 (National Institutes of Health AIDS Research and Reference Reagent Program).
Plant Infection and Virus Isolation.
Tobacco leaves were inoculated with in vitro transcription products of recombinant TMV strains. The transcription products of recombinant virus were diluted 1:1 in 30 mM sodium phosphate, pH 7.2, and applied to expanding tobacco leaves after abrading the leaf surface with carborundum (320 grit; Fisher). Inoculation was effected by gentle rubbing to spread the inoculum and further abrade the leaf surface. The recombinant virus was isolated 12–14 days after the inoculum was applied, as described (14). Briefly, leaf tissue was ground and the sap separated from cell debris by centrifugation. Virus particles were selectively precipitated using 5% polyethylene glycol.
Electron Microscopy.
Crude extracts of infected plant tissue were adsorbed on carbon-coated grids and stained in 2% uranyl acetate. Grids were examined using a electron microscopy (magnification, ×30,000).
Mice.
Eight-week-old female Swiss–Webster mice were used in these experiments (Taconic Farms). Mice were housed in a temperature- and light-cycle-controlled room at the Thomas Jefferson University Animal Facility.
Viruses and Cell Culture.
The baby hampster kidney cell line used for rabies virus neutralization assay was maintained as described (26). The human T-lymphotropic virus, type I/MT2 cell lines (obtained from the National Institutes of Health AIDS Research and Reference Reagent Program) were used as target cells in HIV-1 neutralization assay. These cells were maintained in RPMI medium 1640 supplemented with 10% fetal bovine serum, penicillin-streptomycin, and pyruvate. The cell-free HIV-1/MN isolate was propagated in human T-lymphotropic virus, type I/MT2 cells as described (21).
Immunization of Mice with Recombinant Plant Virus.
Mice (five in each group) were immunized intraperitoneally with seven doses (10 μg per injection) of recombinant AIMV virus at 2-week intervals, and serum samples were collected 12 days after the third and seventh immunizations (27). For each engineered construct four groups of mice were tested: two groups immunized with recombinant virus with or without adjuvant, and two control groups immunized in the presence or absence of adjuvant with a mixture of wild-type AIMV and TMV prepared from 30BRz-infected plants (30B/AIMV). In the group receiving adjuvant, mice were injected with 10 μg of recombinant virus in complete Freund’s adjuvant (CFA), followed 14 days later with a second dose of antigen in incomplete Freund’s adjuvant; all subsequent inoculations were performed without adjuvant.
ELISA.
Mouse sera were analyzed for the presence of antigen-specific antibody by direct ELISA as described (27), using 96-well plates coated with 100 μl per well of inactivated ERA strain rabies virus or with a synthetic peptide (5 μg/ml) derived from the sequences of the V3 loop of HIV-1 overnight at 24°C. Peroxidase-conjugated goat anti-mouse IgG (γ-chain-specific) was used as secondary antibody.
Neutralization Assay.
Sera from mice inoculated with recombinant AIMV particles consisting of CP Drg24 fusion (CPDrg24) and control virus (30B/AIMV) were heat-inactivated at 56°C for 30 min and incubated with CVS-11 strain rabies virus. Pretreated virus was used for the infection of baby hamster kidney indicator cells (26). The neutralizing activity of rabies virus specific serum antibodies was determined using a rapid fluorescent focus-forming inhibition test as described (26). Neutralizing antibodies to HIV-1/MN were measured in a syncytia-forming assay as described (21, 28). Cell-free virus (100 TCID50) was preincubated with different dilutions of heat-inactivated preimmune or immune sera for 1 hr at 37°C. Following incubation, the pretreated virus was used to infect human T-lymphotropic virus, type I/MT2 cells. Syncytia formation was evaluated 5 days after inoculation of human T-lymphotropic virus, type I/MT2 cells by phase contrast microscopy (28).
RESULTS
Gene Engineering and Plasmid Constructs.
We have engineered recombinant genes encoding peptides that represent important antigenic domains of HIV-1 and of rabies virus as fusion proteins with the AIMV CP. The starting plasmid pSPΔAUG (29) contains an AIMV CP modified so that the AUG translation initiation codon is replaced by TCG to create an XhoI (CTCGAG) site for cloning and an RNA molecule defective in translation. To create a fusion protein consisting of a full-length AIMV CP and the V3 loop of the HIV-1 MN isolate (MNV3), a 141-bp PCR fragment (coding for 47 amino acids) containing the V3 loop and XhoI sites at each end was digested with XhoI and ligated into pSPΔAUG linearized by XhoI to engineer CPMNV3. The resulting plasmid is termed pSPCPMNV3. Translation of the recombinant protein initiates from the AUG codon created at the 5′ end of the chimeric gene, which is upstream of nucleotide sequences encoding the HIV V3 loop. The recombinant plasmid also contains linking 5′ (37 nucleotides upstream from the wild-type AIMV CP translation start codon) and 3′ (192 nucleotides following the AIMV CP stop codon and containing the AIMV origin of assembly) noncoding regions of the AIMV CP. An EcoRI–SmaI fragment of pSPCPMNV3 consisting of the HIV-1 V3 loop, the AIMV CP, and the 5′ and 3′ noncoding regions of the AIMV CP was blunt-ended and ligated into the XhoI site of 30BRz. Fig. 1 shows a linear map of the resulting plasmid, pBRzCPMNV3.
We engineered a linear epitope, G5-24, of rabies virus glycoprotein as a chimera (Drg24) with an epitope from the rabies virus nucleoprotein (31D) and fused it with the AIMV CP. The chimeric epitope, Drg24, was synthesized by PCR using oligonucleotides containing 18 complementary nucleotides between the first- and second-strands that can anneal and initiate the PCR reaction. The 120-bp (coding for 40 amino acids) PCR product was digested with XhoI and cloned into pSPΔAUG to create pSPCPDrg24. The full-length fusion protein with the 5′ and 3′ noncoding regions of the AIMV CP was excised from pSPCPDrg24 by digestion with EcoRI and SmaI, and ligated into 30BRz, linearized and blunt-ended as above, to create pBRzCPDrg24 (Fig. 1).
Expression of Chimeric AIMV CP in Recombinant Virus-Infected Plants.
Infectious, in vitro produced RNA transcripts of chimeric TMV (pBRzCPMNV3, pBRzCPDrg24, or 30BRz) were inoculated on expanding leaves of Nicotiana benthamiana, and recombinant protein expression was assessed by Western blot analysis of virus purified from locally and systemically infected leaves. Monoclonal antibodies specific for the AIMV CP (Fig. 2 A and B) detected proteins of 28.9 kDa, indicating production of full-length fusion proteins in the virus-infected plants. Virus prepared from both locally and systemically infected tissue contained the transgene epitope Drg24 (Fig. 2A, lanes 2 and 3) or MNV3 (Fig. 2B, lanes 1 and 2) with an expected size of 28.9 kDa. A portion of these samples were used to determine the identity of the antigenic epitopes in the fusion proteins. Mouse mAbs specific for the G5-24 epitope of rabies virus glycoprotein detected protein (CPDrg24) of the correct size (28.9 kDa) in samples from locally and systemically infected tissue (Fig. 2C, lanes 2 and 3). Antibodies specific for the V3 loop of HIV-1 detected CPMNV3 (Fig. 2D, lanes 2 and 3). The control protein (wild-type AIMV CP) also had the correct size of 24 kDa (Fig. 2A, lane 1). Virus samples from plants inoculated with unmodified vector (Fig. 2B, lane 3) or AIMV (Fig. 2 C and D, lanes 4) were used as a negative control. These findings demonstrates that the recombinant viruses are stable and retain the fusion protein during systemic movement throughout the plant tissue.
Figure 2.
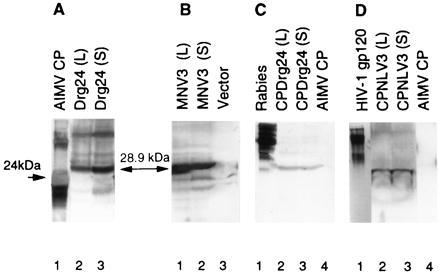
Western blot showing accumulation of recombinant AIMV CP fused to MNV3 of HIV-1 or Drg24 of rabies virus in locally (L) or systemically (S) infected tobacco leaves. The tobacco leaves were inoculated with transcripts of recombinant virus. Proteins were separated by electrophoresis through a SDS/13% polyacrylamide gel and bound with mAbs for the AIMV CP (A, lanes 2 and 3; B, lanes 1 and 2), for the linear epitope (G5-24) of rabies virus glycoprotein (C, lanes 2 and 3), or for the V3 loop of the HIV-1 (D, lanes 2 and 3). Wild-type AIMV CP (24 kDa) bound only with antibodies against AIMV CP (A, lane 1, positive control) and did not bind with antibodies against fusion peptides (C and D, lanes 4; negative control). Each protein is indicated above the lane.
Particle Assembly.
The recombinant viruses containing the AIMV CP fused to antigenic epitopes have two subgenomic promoters. One promoter produces a subgenomic RNA encoding the TMV CP to encapsidate the viral vector. The second promoter produces subgenomic RNA consisting of the coding region for the recombinant AIMV CP gene and also contains the AIMV origin of assembly in the 3′ noncoding region. Consequently, the AIMV CP produced in recombinant virus-infected tissue is expected to encapsidate this subgenomic RNA into spherical or ellipsoid particles. The tissue infected with transcripts produced from constructs pBRzCPMNV3 or pBRzCPDrg24 should then contain two types of particles: (i) rod-shaped TMV particles consisting of recombinant RNA encapsidated with wild-type TMV CP; and (ii) spherical particles consisting of recombinant AIMV CP and subgenomic RNA. Indeed, electron microscopy revealed spherical particles representing recombinant AIMV and rod-shaped particles representing TMV assembled upon infection with transcripts from both pBRzCPMNV3 and pBRzCPDrg24 (Fig. 3).
Figure 3.
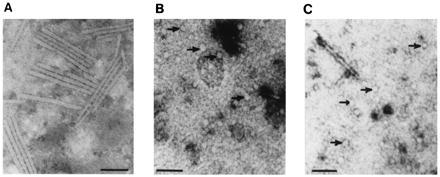
Electron micrographs of particles from tobacco plants, infected with 30BRz (A), pBRzCPMNV3 (B), and pBRzCPDrg24 (C). The spherical recombinant AIMV particles carrying antigens are indicated with arrows. The particles were negatively stained using 2% uranyl acetate. The bars indicate 100 nm. The amount of road shaped particles in chimeric virus infected plants were less compared with that of control.
ELISA and Neutralization Assay.
Sera from mice immunized with CPDrg24 or CPMNV3 were assessed for the presence of antibodies specific for ERA strain rabies virus or for the synthetic peptide derived from the V3 loop of HIV-1 (see Materials and Methods). Low levels of serum antibodies specific for both HIV and rabies virus antigens were detectable after the third inoculation (data not shown). Fourteen days after the last immunization with CPDrg24, high titers of rabies-specific antibodies were detected by ELISA in the sera of mice inoculated with or without CFA, although titers were somewhat higher when CFA was used (Fig. 4 a and b). Thus, plant-produced rabies virus antigen can induce an immune response in mice. Moreover, sera from mice immunized with CPDrg24 neutralized CVS-11 strain rabies virus in vitro (Fig. 4c). At a serum dilution of 1:160, virus neutralization was, on average, 90%. Sera from eight out of 10 mice neutralized the virus while preimmune and control sera had no virus-neutralizing activity (Fig. 4c). When mice were immunized with CPMNV3 serum antibodies specific for the V3 loop of the HIV-1 MN isolate were detected in both ELISA and neutralization assays as shown in Fig. 5. It should be noted that the data in Fig. 5 a and b represent average values from all mice including one of five mice inoculated in the presence of CFA and two of five not given the adjuvant that had failed to respond. While preimmune sera had low neutralizing activity (≈25%) at dilutions 1:128 and 1:256, sera from experimental mice inoculated with CPMNV3 demonstrated, on average, ≈80% and 76% neutralizing activity at the same dilutions, respectively (Fig. 5c).
Figure 4.
Serum antibody response of mice immunized intraperitoneally with CPDrg24 measured by ELISA (a and b) on a plates coated with inactivated rabies virus (ERA) and neutralizing activity of these antibodies. Each graph represents the average of the values calculated for individual mice. 30B/AIMV indicates the sera from the mice immunized with control (wild-type virus lacking insert). (a) The data when the antigen was administered without CFA; whereas b represents the data when CFA was incorporated. The curves reflect average ELISA data at different dilutions of sera. Graph c shows neutralization of the CVS-11 strain rabies virus by sera from mice immunized with recombinant CPDrg24, 30B/AIMV, or preimmune. The curves represent neutralizing titers of sera after seven immunizations. (c) Average data with SD from eight mice that had neutralizing serum antibodies.
Figure 5.
Serum antibody response of mice immunized with CPMNV3 and neutralizing activity of these antibodies. Serum antibody response was measured by ELISA on a plates coated with synthetic peptide resembling the V3 loop of HIV-1. (a) Serum antibody response when the antigen was administered without CFA; whereas b represents when CFA was added. Also shown are the ELISA data for preimmune and mice immunized with control virus. (c) Neutralization of HIV-1/MN isolate by sera from mice immunized with CPMNV3. Data points in c are averages obtained using preimmune and sera after the last (seventh) inoculation of antigen.
DISCUSSION
TMV has been successfully used as a vector for the expression of foreign proteins in plants (23, 24). Its wide host range permits the use of a variety of plant species as production and delivery systems. In addition, expression of the recombinant protein is controlled by the viral replication machinery and does not interfere with plant nuclear transcription. For vaccine development, expression of antigenic peptides in the context of the viral CP is desired because of the high levels of accumulation of this protein, its capacity to self-assemble into particles, and the potential to display antigenic epitopes on the surfaces of virus particles. While these features make TMV an attractive vector for the expression of foreign proteins, the use of the CP of TMV as a carrier molecule is limited due to the failure of self-assembly when peptides greater than 24–25 amino acids are inserted. On the other hand, as we have confirmed here, the CP of AIMV can self-assemble while carrying significantly larger peptides. In this study we have demonstrated that TMV can be used as a vector to express recombinant AIMV CP containing antigenic peptides from rabies virus and HIV. Plants infected with these constructs produce immunogenic particles both at the site of inoculation and systemically. Moreover, the recombinant TMV remains infectious and retains the foreign sequences during passage from plant to plant (data not shown).
The rabies virus glycoprotein has been identified as a major antigen that induces protective immunity against rabies virus (22). Using synthetic peptides, Dietzschold et al. (17) showed that mice can be effectively immunized against rabies virus when a chimeric polypeptide representing linear epitope of rabies glycoprotein and dominant epitope of nucleoprotein is used as antigen. Indeed, immunization of mice with recombinant virus particles containing the chimeric Drg24 stimulated the production of rabies virus-specific antibodies with capacity to neutralize CVS-11 strain rabies virus. Immunization of mice with recombinant antigen containing the V3 loop of HIV-1, which is known to elicit virus-neutralizing antibodies (30), also resulted in the production of specific neutralizing serum antibodies. For example sera from mice immunized with CPMNV3 resulted in up to 80% neutralization of an HIV-1/MN isolate.
Our data describe the capacity of recombinant AIMV CP to function as a carrier molecule with an inherent capacity to assemble into virus particles. In addition to facilitating antigen purification this particulate nature may enhance the immunogenicity of the construct. In terms of vaccine development, it may be expected that expression of antigenic determinants in the form of a virus particles may prove to be more immunogenic than other vehicles of antigen production and delivery. The results that we have obtained with recombinant particles containing rabies Drg24 and HIV MNV3 support the notion that plants and plant viruses may provide a safe and efficient production system for vaccines and other molecules of biomedical importance. With the potential to use edible plant tissue to economically produce large quantities of vaccines, such as CPDrg24 or CPMNV3, the system described here may be ideally suited for use in stimulating mucosal immunity.
Acknowledgments
We thank Dr. W. Dowson and Dr. S. Shivprasad for the gift of 30BRz and for the help in infection of plants, M. Bennett for the help with HIV neutralization assay, and Dr. B. Dietzschold and Dr. P. McGarvey for valuable discussions and for critical reading of this manuscript. This work was funded by a grant from the Commonwealth of Pennsylvania.
ABBREVIATIONS
- TMV
tobacco mosaic virus
- CP
coat protein
- AIMV
alfalfa mosaic virus
- CFA
complete Freund’s adjuvant
References
- 1.Mason H S, Lam D M-K, Arntzen C J. Proc Natl Acad Sci USA. 1992;89:11745–11749. doi: 10.1073/pnas.89.24.11745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Haq T A, Mason H, Clements J D, Arntzen C J. Science. 1995;268:714–716. doi: 10.1126/science.7732379. [DOI] [PubMed] [Google Scholar]
- 3.McGarvey P B, Hammond J, Dienelt M M, Hooper D C, Fu Z F, Dietzschold B, Koprowski H, Michaels F H. Bio/Technology. 1995;13:1484–1487. doi: 10.1038/nbt1295-1484. [DOI] [PubMed] [Google Scholar]
- 4.Mason H S, Ball J M, Shi J J, Jiang X, Estes M K, Arntzen C J. Proc Natl Acad Sci USA. 1996;93:5335–5340. doi: 10.1073/pnas.93.11.5335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Smith G P. Science. 1985;228:1315–1317. doi: 10.1126/science.4001944. [DOI] [PubMed] [Google Scholar]
- 6.Greenwood J, Willis A E, Perham R N. J Mol Biol. 1991;220:821–827. doi: 10.1016/0022-2836(91)90354-9. [DOI] [PubMed] [Google Scholar]
- 7.Mastico R A, Talbot S J, Stockley P G. J Gen Virol. 1993;74:541–548. doi: 10.1099/0022-1317-74-4-541. [DOI] [PubMed] [Google Scholar]
- 8.Burke K L, Dunn G, Ferguson M, Minor P D, Almond J W. Nature (London) 1988;332:81–82. doi: 10.1038/332081a0. [DOI] [PubMed] [Google Scholar]
- 9.Dedieu J F, Ronco J, van der Werf S, Hogle J M, Henin Y, Girard M. J Virol. 1992;66:3161–3167. doi: 10.1128/jvi.66.5.3161-3167.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Arnold G F, Resnick D A, Li Y, Zhang A, Smith A D, Geisler S C, Jacobo-Molina A, Lee W, Webster R G, Arnold E. Virology. 1994;198:703–708. doi: 10.1006/viro.1994.1082. [DOI] [PubMed] [Google Scholar]
- 11.Hamamoto H, Sugiyama Y, Nakagawa N, Hashida E, Matsunaga Y, Takemoto S, Watanabe Y, Okada Y. Bio/Technology. 1993;11:930–932. doi: 10.1038/nbt0893-930. [DOI] [PubMed] [Google Scholar]
- 12.Usha R, Rohll J B, Spall V E, Shanks M, Maule A J, Johnson J E, Lomonosoff G P. Virology. 1993;197:366–374. doi: 10.1006/viro.1993.1598. [DOI] [PubMed] [Google Scholar]
- 13.Porta C, Spall V E, Loveland J, Johnson J E, Barker P J, Lomonossoff G P. Virology. 1994;202:949–955. doi: 10.1006/viro.1994.1417. [DOI] [PubMed] [Google Scholar]
- 14.Fitchen J, Beachy R N, Hein M B. Vaccine. 1995;13:1051–1057. doi: 10.1016/0264-410x(95)00075-c. [DOI] [PubMed] [Google Scholar]
- 15.McLain L, Porta C, Lomonossoff G P, Durrani Z, Dimmock N J. AIDS Res Hum Retroviruses. 1995;11:327–34. doi: 10.1089/aid.1995.11.327. [DOI] [PubMed] [Google Scholar]
- 16.Turpen T H, Reini S J, Charoenvit Y, Hoffman S L, Fallarme V, Grill L K. Bio/Technology. 1995;13:53–57. doi: 10.1038/nbt0195-53. [DOI] [PubMed] [Google Scholar]
- 17.Dietzschold B, Gore M, Marchadier D, Niu H-S, Bunschoten H M, Otvos L, Wunner W H, Ertl H C J, Osterhaus A D M E, Koprowski H. J Virol. 1990;64:3804–3809. doi: 10.1128/jvi.64.8.3804-3809.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bol J F, Van Vloten-Doting L, Jaspars E M J. Virology. 1971;46:73–85. doi: 10.1016/0042-6822(71)90007-9. [DOI] [PubMed] [Google Scholar]
- 19.Yusibov V M, Kumar A, North A, Johnson J E, Loesch-Fries L S. J Gen Virol. 1995;77:567–573. doi: 10.1099/0022-1317-77-4-567. [DOI] [PubMed] [Google Scholar]
- 20.Sambrook S, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 21.Wang B, Ugen K E, Srikantan V, Agadjanyan M G, Dang K, Rafaeli Y, Sato A, Boyer J, Williams W V, Winer D B. Proc Natl Acad Sci USA. 1993;90:4156–4160. doi: 10.1073/pnas.90.9.4156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Dietzschold B, Ertl H C J. Crit Rev Immunol. 1991;10:427–439. [PubMed] [Google Scholar]
- 23.Donson J, Kearney C M, Hilf M E, Dawson W O. Proc Natl Acad Sci USA. 1991;88:7204–8. doi: 10.1073/pnas.88.16.7204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yusibov V M, Loesch-Fries L S. Proc Natl Acad Sci USA. 1995;92:8980–8984. doi: 10.1073/pnas.92.19.8980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Loesch-Fries L S, Hall T C. J Gen Virol. 1980;47:323–332. [Google Scholar]
- 26.Wiktor T J, Macfarlan R I, Foggin C M, Koprowski H. Dev Biol Stand. 1984;57:199–211. [PubMed] [Google Scholar]
- 27.Hooper D C, Pierard I, Modelska A, Otvos L, Fu Z F, Koprowski H, Dietzschold B. Proc Natl Acad Sci USA. 1994;91:10908–10912. doi: 10.1073/pnas.91.23.10908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nara P. In: Techniques in HIV Research. Aldovini A, Walker B D, editors. New York: Stockton; 1989. pp. 77–86. [Google Scholar]
- 29.Yusibov V M, Loesch-Fries L S. Virology. 1995;208:405–407. doi: 10.1006/viro.1995.1168. [DOI] [PubMed] [Google Scholar]
- 30.Rencher S D, Slobod K S, Dawson D H, Lockey T D, Hurwitz J L. AIDS Res Hum Retroviruses. 1995;11:1131–1133. doi: 10.1089/aid.1995.11.1131. [DOI] [PubMed] [Google Scholar]