

Abstract
Tumor necrosis factor (TNF) α has been shown to be a major therapeutic target in rheumatoid arthritis with the success of anti-TNFα antibody clinical trials. Although signaling pathways leading to TNFα expression have been studied in some detail, there is evidence for considerable differences between individual cell types. This prompted us to investigate the intracellular signaling pathways that result in increased TNFα synthesis from macrophages in the diseased synovial joint tissue. Using an adenoviral system in vitro we report the successful delivery of genes to more than 95% of normal human macrophages. This permitted us to show, by using adenoviral transfer of IκBα, the natural inhibitor of NF-κB, that induction of TNFα in normal human macrophages by lipopolysaccharide, but not by some other stimuli, was inhibited by 80%. Furthermore the spontaneous production of TNFα from human rheumatoid joint cell cultures was inhibited by 75%, indicating that the NF-κB pathway is an essential step for TNFα synthesis in synovial macrophages and demonstrating that NF-κB should be an effective therapeutic target in this disease.
There is increasing evidence that chronic inflammatory diseases such as rheumatoid arthritis (RA) are caused by prolonged production of proinflammatory cytokines, especially tumor necrosis factor (TNF) α. These concepts were first demonstrated by using dissociated rheumatoid joint cell cultures (1–3), then substantiated in animal models of arthritis (4, 5), and formally proven by the success of clinical trials of anti-TNFα antibody (6–9) and TNF receptor-Ig fusion proteins (10). The definition of TNFα as a therapeutic target in RA, and subsequently Crohn’s disease (11), has led to a search for the intracellular signaling pathways that drive TNFα production in these diseases, the blockage of which might mimic the clinical benefit of anti-TNFα therapy. In RA joints, the cells producing the majority of TNFα are macrophages (12), and hence there is considerable interest in understanding the regulation of TNFα production in human cells of this lineage.
The expression of the TNFα gene is under complex control, with the p38 mitogen-activated protein kinase pathway controlling translation possibly by actions on the 3′ untranslated region (13), and with the 5′ untranslated region/promoter containing binding sites for multiple transcription factors, including NF-κB, AP-1, NF-interleukin (IL) 6, and NF-AT (14–16). It is not known which of these transcription factors is essential or rate limiting for TNFα expression. In the murine RAW 264 cell line and bone marrow-derived macrophages, NF-κB was shown to be essential for TNFα production (14, 17), whereas in human lymphoid cell lines NF-AT rather than NF-κB was required for TNFα production (15, 16). No data has yet been reported on the TNFα regulation in normal primary human macrophages or cells from RA synovium, the most relevant models for TNFα production in RA.
There are a number of obstacles in studying the biochemical signaling pathways that are important in regulating TNFα production in macrophages. One of the most problematic has been the difficulty in transfecting DNA into macrophages (18), possibly because this cell lineage is specialized for phagocytosis and consequently may degrade DNA. To enable the analysis of such pathways in macrophages we sought a technique to permit efficient gene transfer into this cell lineage. Adenoviral gene transfer has many desirable features (19). Adenoviruses infect a wide spectrum of nondividing cells, in particular respiratory and gut epithelium (19), the natural targets for infection, but also endothelium and fibroblasts. Hemopoietic cells are not natural targets for adenoviral infection, although there are some reports of 10–20% infection of activated T cells (20). Adenoviral infection of fresh monocytes has not been reported, but up to 50% of monocytes cultured with growth factors can be infected (20, 21). In this study we have used adenoviral vectors to investigate signaling pathways in macrophages and human disease tissue. We report that by optimizing conditions very high adenovirus infection efficiencies can be achieved in human macrophages with >95% cells expressing β-galactosidase (β-gal), for example. By using an adenoviral vector encoding IκBα (AdvIκBα), we were able to obtain high levels of expression of this protein with concomitant inhibition of NF-κB. This resulted in suppression of lipopolysaccharide (LPS)-induced TNFα mRNA and protein in macrophages. There was also suppression of the spontaneous TNFα production in RA synovial cells. These data have major implications for NF-κB as a potential therapeutic target in RA.
MATERIALS AND METHODS
Cells.
Mononuclear cells were isolated from single donor plateletphoresis residues obtained from the North London Blood Transfusion Centre (United Kingdom) by Ficoll-Hypaque centrifugation preceding monocyte separation in a Beckman GEL elutriator. Monocyte purity was assessed by flow cytometry using directly congregated anti-CD45 and anti-CD14 antibodies and was routinely greater than 90%. Cells were cultured in RPMI medium 1640 with 25 mM Hepes and 2 mM l-glutamine supplemented with 5% heat-inactivated fetal calf serum and 100 units/ml of penicillin/streptomycin at 37°C. Murine RAW 246.7 cells were maintained in the same supplemented medium.
Monocytes were untreated or treated with granulocyte/macrophage colony-stimulating factor (GM-CSF) (5,000 units/ml), or M-CSF (100 ng/ml), generous gifts of Glenn Larsen, Genetics Institute, Boston), for 48 hr. Expression of integrins was assayed by indirect immunofluoresence and FACS analysis using mAbs to αVβ3. LM609, provided by IXSYS, (San Diego, CA), αVβ5 (undiluted active supernatant of P5H9-E11 kindly provided by William Smith and J. Gamble, Hanson Centre, Adelaide, Australia) or the negative control mAb OX14 (100 μg/ml). Cells were stimulated with 10 ng/ml LPS (Salmonella typhimurium, Sigma).
Synovium from rheumatoid patients undergoing joint replacement surgery was dissociated by cutting into small pieces, digested with collagenase and DNase (1). The total cell mixture, with T cells and macrophages as the most abundant cells, was cultured in 48-well plates at 1 × 106 cells/ml in the same medium containing 10% heat-inactivated fetal calf serum.
Adenoviral Vectors and their Propagation.
Recombinant, replication-deficient adenoviral vectors encoding the Escherichia coli β-gal (Advβgal) or having no insert (AdvO) were provided by A. Byrnes and M. Wood (University of Oxford, U.K.). An adenovirus encoding porcine IκBα with a cytomegalovirus promoter and a nuclear localization sequence (AdvIκBα) was as published (22). Viruses were propagated in the 293 human embryonic kidney cell line and purified by ultracentrifugation through two cesium chloride gradients (23). Titers of viral stocks were determined by plaque assay in 293 cells after exposure to virus for 2 hr in serum-free RPMI medium 1640. After cytokine treatment macrophages were exposed to virus for 2 hr in serum-free RPMI medium 1640 followed by washing and reculturing the cells in complete medium for 48–72 hr. Intracellularly expressed β-gal was assayed by FACS as previously described (24).
Western and Northern Blotting, Electrophoretic Mobility-Shift Assay, and ELISA Procedures.
Cells were scraped off the plates, and cytosolic and nuclear extracts were prepared as described by Whiteside et al. (25). Proteins were separated by SDS/PAGE on a 10% (wt/vol) polyacrylamide gel, ensuring equivalent amounts loaded per track, followed by electro-transfer onto nitrocellulose membranes. IκBα and NF-κB p50 and p65 were detected by using antibodies kindly provided by R. Hay (University of St. Andrews, U.K.), and a horseradish peroxidase-conjugated donkey anti-rabbit antibody (Amersham) and visualized by enhanced chemiluminescence (Amersham). Anti-p42/44 MAPK was obtained from Santa Cruz Biotechnology.
For electrophoretic mobility-shift assay, nuclear extracts were prepared and 20 μg of protein was analyzed for NF-κB or AP-1 activity as previously described (24). Northern blotting was performed as described (1), and ELISA for TNFα was as previously reported (26).
RESULTS
Efficient Gene Transfer into Macrophages with Adenovirus.
Like others (20, 21), we were unable to virally infect freshly prepared monocytes, as assessed by using an adenovirus encoding β-gal under the control of cytomegalovirus promoter (Advβgal), even at a multiplicity of infection (moi) of >1,000 (data not shown). αV integrins have been described to be cofactors for adenovirus infection (27). αVβ3 and αVβ5 expression was found to be very low on resting monocytes. Treatment with GM-CSF or M-CSF for 48 hr up-regulated the expression of αVβ3 and αVβ5, respectively (Fig. 1A). Previous treatment for 72 hr with GM-CSF or M-CSF resulted in 52% or >90% of the cells, respectively, expressing β-gal after infection with Advβgal at 100 moi (Fig. 1B). Thus subsequent studies concentrated on using M-CSF. Moi of 100 and 50 gave >90% infection, but at a moi of 25, infection was reduced to 20% (Fig. 1C). The capacity of adenovirus to infect monocytic cell lines then was investigated. Human U937 and THP-1 were totally refractory, even after treatment with GM-CSF or M-CSF, which did not up-regulate either of the αV integrins (data not shown). In contrast, the murine macrophage cell line RAW 264.7 was readily infected by adenovirus (>95% at moi of 100, >60% at moi of 10, results not shown), even without previous exposure to M-CSF.
Figure 1.

The effect of cytokine treatment on integrin expression and adenoviral infection of primary human monocytes. Monocytes were untreated or treated with GM-CSF and M-CSF for 48 hr followed by assay for (A) expression of integrins αVβ3 (broken line), αVβ5 (dotted line), or the negative control OX14 (solid line) using indirect immunofluoresence and FACS (FL1); or β-gal activity subsequent to infection with Advβgal (dotted line) at a moi of 100 (B) or various moi (C) assayed by FACS.
With successful gene transfer of more than 90%, and approaching 100% of the monocytes, it became possible to determine whether the adenoviral system could be used to analyze the intracellular signaling pathways controlling TNFα. For this purpose we used a replication-deficient adenoviral vector expressing porcine IκBα under the control of the cytomegalovirus promoter that binds human NF-κB (22). A nuclear localization sequence also had been engineered into the IκBα, as nuclear expression of this protein can prevent NF-κB binding to DNA. Infection of endothelial cells with this virus (AdvIκBα) resulted in specific inhibition of NF-κB and markedly reduced expression of proteins such as vascular cell adhesion molecule 1, and also reduced mRNA expression for cytokines such as IL-6 and IL-8, which depend on NF-κB (22).
Infection of RAW 264.7 cells and M-CSF-treated normal human macrophages with AdvIκBα resulted in the overexpression of IκBα (Fig. 2), whereas infection by control virus had no effect. The antibody used does not recognize murine IκBα, hence the endogenous protein was not detected in RAW 264.7 cells. Blots were reprobed for p42 mitogen-activated protein kinase, which showed no change in expression after adenovirus infection. In human macrophages nuclear IκBα was also detectable after infection with AdvIκBα (Fig. 2C).
Figure 2.

Expression of IκBα in AdvIκBα-infected macrophages and RAW 264.7 cells. RAW 264.7 cells (A) or M-CSF-treated human monocytes (B) were untreated or infected with control adenovirus (Adv0) or infected with the IκBα adenovirus (AdvIκBα) at the given moi. After 48 hr, cytosolic (A and B) and nuclear (C) extracts were prepared and assayed for IκBα expression by Western blotting. Blots were stripped and reprobed for p42/44MAPK as a control.
LPS induces degradation of IκBα (Fig. 3A). Human monocytes infected with AdvIκBα, but not control virus-infected cells, expressed high levels of IκBα even after LPS stimulation (Fig. 3A). In contrast, IκBβ was not affected. Translocation of the NF-κB p65 subunit into the nucleus after activation by LPS was nearly abolished by the excess IκBα expression, whereas NF-κB p50 nuclear localization was only partly diminished (Fig. 3A). Using the electrophoretic mobility-shift assay, nuclear NF-κB DNA binding activity was observed to be greatly reduced in AdvIκBα-infected cells. In contrast, AP-1 activity, which is constitutive in M-CSF-treated monocytes, was not significantly altered by LPS or IκBα overexpression (Fig. 3B).
Figure 3.
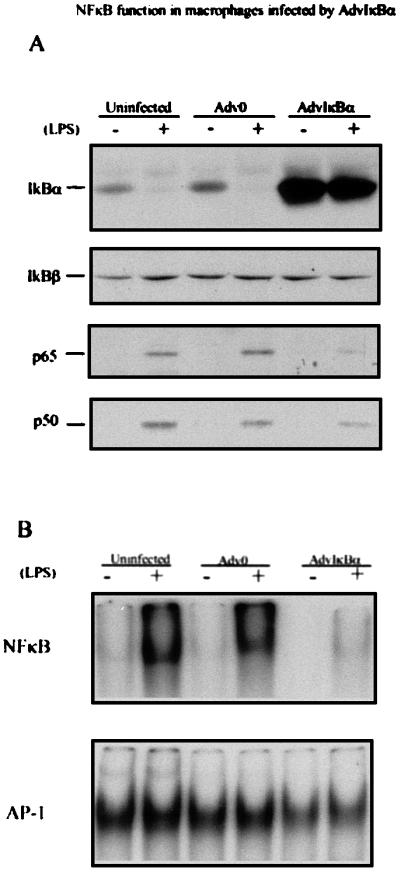
Inhibition of NF-κB function in AdvIκBα-infected macrophages. M-CSF-treated monocytes were infected with AdvO or AdvIκBα at a moi of 50. A third group was untreated. After an additional 3 days in M-CSF the cells were washed, and half of each group was treated (+) or otherwise (−) with 10 ng/ml LPS for 30 min. (A) The cells then were lysed, and cytosolic and nuclear extracts were prepared and analyzed for IκBα/β expression in the cytosol, or p65/p50 expression in the nucleus, by immunowestern blotting. Protein (150 μg and 50 μg) was analyzed for each cytosol or nuclear sample, respectively. (B) Nuclear extracts were prepared and 20 μg protein analyzed for NF-κB (Upper) or AP-1 (Lower) activity by electrophoretic mobility-shift assay.
Analysis of the Role of NF-κB in TNFα Production in Normal LPS-Stimulated Macrophages and in Human Rheumatoid Synovial Tissue.
Having demonstrated that infection of macrophages with the AdvIκBα virus essentially abolished NF-κB activity in the nucleus, this tool could be used to evaluate whether NF-κB is required for macrophage TNFα expression. Previous studies in other systems have given variable results, and the role of NF-κB in TNFα expression has never been tested in human primary macrophages, the most relevant cells in the context of chronic inflammatory disease. The effect of AdvIκBα on LPS-induced TNFα production from normal human macrophages revealed a dose-dependent inhibition, with >80% inhibition at a moi of 80 (Fig. 4A), whereas control adenovirus had no effect on TNFα expression. AdvIκBα-inhibited LPS induced TNFα mRNA expression, whereas there was no effect on a housekeeping gene, glyceraldehyde-3-phosphate dehydrogenase mRNA (Fig. 4B). The ability of anti-TNFα antibody to interfere with other proinflammatory cytokines constitutively produced in RA joint cell cultures was our first clue that TNFα was a therapeutic target in RA (1–3). The capacity of AdvIκBα to inhibit constitutive TNFα production in rheumatoid joint cell cultures, an in vitro model of RA thus was investigated. AdvIκBα reduced TNFα production, detected by ELISA, by 75 ± 15% at a moi of 40 in five different RA synovial joint cell cultures (Fig. 4C). This finding indicates that NF-κB activation is an important step in generation of TNFα in RA joints, and hence that inhibition of the NF-κB pathway would be an effective therapeutic target in RA.
Figure 4.

AdvIκBα infection inhibits TNFα induction in primary cells and rheumatoid joint cell cultures. Human monocytes cultured in the presence of M-CSF for 48 hr were untreated, infected with control adenovirus (Adv0), or infected with AdvIκBα at various moi. Two days after infection, cells were replated at 5 × 105 cells per well on a 96-well plate (A) or at 5–10 × 106 cells per 100 mm Petri dish (B) and stimulated with 10 ng/ml LPS for 2 hr (A) or 16 hr (B). (A) Supernatants were removed and analyzed for TNFα by ELISA. (B) Cells were harvested and mRNA extracted and subjected to Northern analysis with the indicated probes. A represents results from seven different blood donors. Data in A and B are expressed as a percentage of the TNFα production from LPS-activated uninfected cells. Using a paired comparisons Student’s t test, there was significant (P < 0.001) inhibition at 20:1, 40:1, and 80:1 of AdvIκB. (C) Cells from rheumatoid synovium were cultured in 48-well plates at 1 × 106 cells and infected with either AdvIκBα or AdvO at a moi of 40 to 1. The supernatants were taken for ELISA assay at 48 hr; pooled data from the five patients is shown (±SD). The null hypothesis that there was no change in TNF production whether cells were infected with AdvIκB or Adv0 could be rejected (P < 0.05) using a Wilcoxon’s signed rank test on paired differences compared with untreated cells on each patient.
Blockage of NFκB in Normal Human Macrophages Does Not Augment Apoptosis.
Inhibition of TNFα-induced NFκB has been reported to promote apoptosis in a variety of cell lines, e.g., HeLa, HT1080, Jurkat (28–30). It was thus conceivable that the mechanism of AdvIκBα-induced reduction of TNFα production may have been caused by apoptosis-mediated cell depletion. This is unlikely as there was no reduction in cell numbers in IκB virus-treated macrophages at the moi used, and no indication of cell death as measured by microscopy or the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide assay (data not shown). Secondly, using a DNA fragmentation assay (31), no induction of apoptosis was detectable in AdvIκBα-infected macrophages after treatment with exogenous TNFα (Fig. 5). In these macrophages a mixture of TNFα and cycloheximide induced apoptosis (Fig. 5). Failure to induce apoptosis in macrophages was not caused by the effectiveness of the method of inhibiting NF-κB activity, because TNFα treatment of AdvIκBα virus-infected Hela cells induced massive apoptosis (Fig. 5). More likely our results allude to the fact that NF-κB does not have a universal role in preventing apoptosis, and that this role depends on the cell type and/or various stages of cell differentiation and cell cycle. In support of this view, NF-κB has been demonstrated to promote apoptosis in human embryonic kidney 293 cell line (32).
Figure 5.

Lack of apoptosis with TNFα in IκBα-infected macrophages. Human monocytes treated with M-CSF for 48 hr (A) or Hela cells (B) were plated on 60-mm Petri dishes at a density of 2–3 × 106 cells per dish. They were left untreated or infected with control adenovirus or AdvIκBα at a moi of 40. Two days after infection, TNFα (20 ng/ml) and cycloheximide (2 μg/ml) were added as indicated. After 16 hr cells were stained for 30 min in 1.5 ml of a hypotonic fluorochrome solution (50 μg/ml of propidium iodide in 0.1% sodium citrate plus 0.1% Triton X-100), and the resulting propidium iodide-stained nuclei were analyzed by flow cytometry. The percentage of cells in each sector is indicated.
DISCUSSION
We have shown here that adenoviruses may be highly useful in delivering genes to normal human macrophages, permitting analysis of the signaling pathways involved in the production of cytokines by these cells, and evaluating the pathways involved in a complex disease such as RA. This technique permitted us to demonstrate that LPS-induced TNFα production was NF-κB dependent and set the stage for evaluating whether NF-κB is involved in the constitutive TNFα production occurring in RA joints. The subsequent demonstration of inhibition indicates that NF-κB is important for TNFα production in RA and is a therapeutic target in this disease. Although NF-κB has been shown to be activated in RA synovium previously by immunohistology (33), we have demonstrated a direct link between NF-κB activation and the production of TNFα in human RA tissue.
The most practical way of blocking NF-κB function remains to be ascertained. Several publications have described the effect of small molecules such as gliotoxin, free radical scavengers, and protease inhibitors as potential NF-κB inhibitors but it is unlikely that these will be sufficiently specific. Previously, antisense oligonucleotides to p65 have been used to block NF-κB activation and this was effective when administered locally in a murine model of colitis (34). The possible benefits or otherwise of inhibiting a single NF-κB subunit, versus the use of IκBα, which will inhibit the various preformed dimeric NF-κB factors, are unclear. The recently cloned IκB kinases (35–39), or other steps in the IκB degradation pathway, or the NF-κB nuclear localization pathway also would be appropriate drug targets (40, 41). Gene therapy may be a possibility for a disease where the diseased tissue is accessible, including RA, Crohn’s disease, or psoriasis (42, 43). These results also suggest that suppressing the activity of NF-κB may be a useful target for drugs aimed at reducing the inflammatory effects of excess LPS, such as in endotoxin-induced septic shock (44).
Anti-TNFα antibody treatment of RA joint cell cultures has widespread effects on the cytokine environment. There is diminished expression of a variety of proinflammatory cytokines, e.g., IL-1, GM-CSF, and IL-6 (3). This probably helps account for the efficacy of this antibody in vivo as does its down-regulation of adhesion molecules and chemokines leading to reduced leukocyte trafficking (45). Preliminary data on the expression of cytokines by AdvIκBα-infected primary macrophages and RA synovial cells reveals widespread inhibitory effects on IL-1, IL-6, and IL-8 (unpublished work). An unexpected result was the failure to observe any apoptosis induced by the AdvIκBα in human macrophages, although cell death was induced in Hela cells, as in previously published papers (28, 29, 46). To date most studies linking NF-κB to antiapoptotic effects have been performed in cell lines. However, normal human macrophages provide a primary nonproliferating model, and it is unclear whether the lack of apoptosis is caused by the nondividing nature of the cells or whether the role of NF-κB in protecting cells depends on cell type.
The technique of efficient gene transfer into normal human macrophages described here provides an approach for evaluating the relative importance of various biochemical pathways of signal transduction in the many diseases that involve excess activation of human macrophages, as well as determining the possible therapeutic effects of interrupting intracellular signaling pathways. A key question is the specificity of inhibition by AdvIκB. This also has been addressed by assessing effects of the virus on cytokines like IL-10 and IL-1 receptor antagonist, which are not NF-κB dependent. Furthermore, some stimuli that induce TNFα, e.g., zymosan, are not inhibited by AdvIκB (J.B., K.B., F.B., B.F., and M.F., unpublished work). Another benefit of efficient infection of normal cells is that it allows the regulation of the endogenous chromosomal genes to be studied, without the requirement for the coexpression of ectopically expressed reporter constructs or alternatively the clonal selection of stable cell lines to ensure that an homogenously modified cell population is being investigated. Such stable gene transfer of IκBα cDNA using a retrovirus into THP-1, a monocytic cell line, recently was described (47). This was found to partially inhibit LPS-induced NF-κB activation and markedly reduce IL-1β transcription, IL-6, and IL-8 secretion, results that are consistent with our study (47). However, this retroviral technique is not possible with most normal cells, because only dividing cells integrate retrovirus. Thus, at present the adenoviral method described here is currently the only one known to permit the dissection of signaling pathways in normal macrophages. This has permitted us to demonstrate the importance of NF-κB in the constitutive TNFα production in rheumatoid synovium, predicting that blocking NF-κB should be an effective therapeutic target for RA.
Acknowledgments
We thank Prof. R. N. Maini for his encouragement. This work was funded by the Arthritis and Rheumatism Council (United Kingdom), Wellcome Trust and the European Community Training and Mobility Research fellowship, and the Swedish Institute.
ABBREVIATIONS
- TNF
tumor necrosis factor
- LPS
lipopolysaccharide
- RA
rheumatoid arthritis
- AdvIκBα
adenoviral vector encoding IκBα
- GM-CSF
granulocyte/macrophage colony-stimulating factor
- β-gal
β-galactosidase
- moi
multiplicity of infection
- IL
interleukin
References
- 1.Buchan G, Barrett K, Turner M, Chantry D, Maini R N, Feldmann M. Clin Exp Immunol. 1988;73:449–455. [PMC free article] [PubMed] [Google Scholar]
- 2.Brennan F M, Chantry D, Jackson A, Maini R, Feldmann M. Lancet. 1989;2:244–247. doi: 10.1016/s0140-6736(89)90430-3. [DOI] [PubMed] [Google Scholar]
- 3.Feldmann M, Brennan F M, Maini R N. Annu Rev Immunol. 1996;14:397–440. doi: 10.1146/annurev.immunol.14.1.397. [DOI] [PubMed] [Google Scholar]
- 4.Thorbecke G J, Shah R, Leu C H, Kuruvilla A P, Hardison A M, Palladino M A. Proc Natl Acad Sci USA. 1992;89:7375–7379. doi: 10.1073/pnas.89.16.7375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Williams R O, Feldmann M, Maini R N. Proc Natl Acad Sci USA. 1992;89:9784–9788. doi: 10.1073/pnas.89.20.9784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Elliott M J, Maini R N, Feldmann M, Long-Fox A, Charles P, Katsikis P, Brennan F M, Walker J, Bijl H, Ghrayeb J, Woody J. Arthritis Rheum. 1993;36:1681–1690. doi: 10.1002/art.1780361206. [DOI] [PubMed] [Google Scholar]
- 7.Elliott M J, Maini R N, Feldmann M, Kalden J R, Antoni C, Smolen J S, Leeb B, Breedveld F C, Macfarlane J D, Bijl H, et al. Lancet. 1994;344:1105–1110. doi: 10.1016/s0140-6736(94)90628-9. [DOI] [PubMed] [Google Scholar]
- 8.Elliott M J, Maini R N, Feldmann M, Long-Fox A, Charles P, Bijl H, Woody J N. Lancet. 1994;344:1125–1127. doi: 10.1016/s0140-6736(94)90632-7. [DOI] [PubMed] [Google Scholar]
- 9.Rankin E C C, Choy E H S, Kassimos D, Kingsley G H, Sopwith S M, Isenberg D A, Panayi G S. Br J Rheumatol. 1995;34:334–342. doi: 10.1093/rheumatology/34.4.334. [DOI] [PubMed] [Google Scholar]
- 10.Moreland L W, Baumgartner S W, Schiff M H, Tindall E A, Fleischmann R M, Weaver A L, Ettlinger R E, Cohen S, Koopman W J, Mohler K, et al. N Engl J Med. 1997;337:141–147. doi: 10.1056/NEJM199707173370301. [DOI] [PubMed] [Google Scholar]
- 11.Van Dullemen H M, Van Deventer S J H, Hommes D W, Bijl H A, Jansen J, Tytgat G N J, Woody J. Gastroenterology. 1995;109:129–135. doi: 10.1016/0016-5085(95)90277-5. [DOI] [PubMed] [Google Scholar]
- 12.Chu C Q, Field M, Feldmann M, Maini R N. Arthritis Rheum. 1991;34:1125–1132. doi: 10.1002/art.1780340908. [DOI] [PubMed] [Google Scholar]
- 13.Lee J C, Laydon J T, McDonnell P C, Gallagher T F, Kumar S, Green D, McNulty D, Blumenthal M J, Heys J R, Landvatter S W, et al. Nature (London) 1994;372:739–746. doi: 10.1038/372739a0. [DOI] [PubMed] [Google Scholar]
- 14.Shakhov A N, Collart M A, Vassalli P, Nedospasov S A, Jongeneel C V. J Exp Med. 1990;171:35–47. doi: 10.1084/jem.171.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Goldfeld A E, Strominger J L, Doyle C. J Exp Med. 1991;174:73–81. doi: 10.1084/jem.174.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McCaffrey P G, Goldfeld A E, Rao A. J Biol Chem. 1994;269:30445–30450. [PubMed] [Google Scholar]
- 17.Han J, Beutler B. Eur Cytokine Netw. 1990;1:71–75. [PubMed] [Google Scholar]
- 18.Stacey K J, Ross I L, Hume D A. Immunol Cell Biol. 1993;71:75–85. doi: 10.1038/icb.1993.8. [DOI] [PubMed] [Google Scholar]
- 19.Shenk T. In: Fields Virology. 3rd Ed. Fields B N, Knipe D M, Howley P M, Chanock R M, Melnick J L, Monath T P, Roizman B, Straus S E, editors. Philadelphia: Lippincott–Raven; 1996. pp. 2111–2148. [Google Scholar]
- 20.Huang S, Endo R I, Nemerow G R. J Virol. 1995;69:2257–2263. doi: 10.1128/jvi.69.4.2257-2263.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Haddada H, Lopez M, Martinache C, Ragot T, Abina M A, Perricaudet M. Biochem Biophys Res Commun. 1993;195:1174–1183. doi: 10.1006/bbrc.1993.2168. [DOI] [PubMed] [Google Scholar]
- 22.Wrighton C J, Hofer-Warbinek R, Moll T, Eytner R, Bach F H, de Martin R. J Exp Med. 1996;183:1013–1022. doi: 10.1084/jem.183.3.1013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Graham F L, Prevec L. Mol Biotechnol. 1995;3:207–220. doi: 10.1007/BF02789331. [DOI] [PubMed] [Google Scholar]
- 24.Clarke C J, Taylor Fishwick D A, Hales A, Chernajovsky Y, Sugamura K, Feldmann M, Foxwell B M. Eur J Immunol. 1995;25:2961–2966. doi: 10.1002/eji.1830251037. [DOI] [PubMed] [Google Scholar]
- 25.Whiteside S T, Visvanathan K V, Goodbourn S. Nucleic Acids Res. 1992;20:1531–1538. doi: 10.1093/nar/20.7.1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Engelberts I, Moller A, Schoen G J M, van der Linden C J, Buurman W A. Lymphokine Cytokine Res. 1991;10:60–76. [PubMed] [Google Scholar]
- 27.Wickham T, Filardo E J, Cheresh O A, Nemerow G R. Cell. 1995;73:309–319. doi: 10.1016/0092-8674(93)90231-e. [DOI] [PubMed] [Google Scholar]
- 28.Beg A A, Baltimore D. Science. 1996;274:782–784. doi: 10.1126/science.274.5288.782. [DOI] [PubMed] [Google Scholar]
- 29.Wang C-Y, Mayo M W, Baldwin A S. Science. 1996;274:784–787. doi: 10.1126/science.274.5288.784. [DOI] [PubMed] [Google Scholar]
- 30.Van Antwerp D J, Martin S J, Kafri T, Green D R, Verma I M. Science. 1996;274:787–789. doi: 10.1126/science.274.5288.787. [DOI] [PubMed] [Google Scholar]
- 31.Nicoletti I, Migliorati G, Pagliacci M C, Grignani F, Riccardi C. J Immunol Methods. 1991;139:271–279. doi: 10.1016/0022-1759(91)90198-o. [DOI] [PubMed] [Google Scholar]
- 32.Grimm S, Bauer M K A, Baeuerle P A, Schulze-Osthoff K. J Cell Biol. 1996;134:13–23. doi: 10.1083/jcb.134.1.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Marok R, Winyard P G, Coumbe A, Kus M L, Gaffney K, Blades S, Mapp P I, Morris C J, Blake D R, Kaltschmidt C, Baeuerle P A. Arthritis Rheum. 1996;39:583–591. doi: 10.1002/art.1780390407. [DOI] [PubMed] [Google Scholar]
- 34.Neurath M F, Pettersson S, Meyer Zum Buschenfelde K-H, Strober W. Nat Med. 1996;2:998–1004. doi: 10.1038/nm0996-998. [DOI] [PubMed] [Google Scholar]
- 35.Mercurio F, Zhu H, Murray B W, Shevchenko A, Bennett B L, Li J W, Young D B, Barbosa M, Mann M. Science. 1997;278:860–866. doi: 10.1126/science.278.5339.860. [DOI] [PubMed] [Google Scholar]
- 36.Zandi E, Rothwarf D M, Delhase M, Hayakawa M, Karin M. Cell. 1997;91:243–252. doi: 10.1016/s0092-8674(00)80406-7. [DOI] [PubMed] [Google Scholar]
- 37.DiDonato J A, Hayakawa M, Rothwarf D M, Zandi E, Karin M. Nature (London) 1997;388:548–554. doi: 10.1038/41493. [DOI] [PubMed] [Google Scholar]
- 38.Regnier C H, Song H-Y, Gao X, Goeddel D V, Cao Z, Rothe M. Cell. 1997;90:373–383. doi: 10.1016/s0092-8674(00)80344-x. [DOI] [PubMed] [Google Scholar]
- 39.Woronicz J D, Gao X, Cao Z, Rothe M, Goeddel D V. Science. 1997;278:866–869. doi: 10.1126/science.278.5339.866. [DOI] [PubMed] [Google Scholar]
- 40.Palombella V, Rando O, Goldberg A, Maniatis T. Cell. 1994;78:773–785. doi: 10.1016/s0092-8674(94)90482-0. [DOI] [PubMed] [Google Scholar]
- 41.Baldwin A S. Annu Rev Immunol. 1996;14:649–683. doi: 10.1146/annurev.immunol.14.1.649. [DOI] [PubMed] [Google Scholar]
- 42.Chernajovsky Y, Feldmann M, Maini R N. Br Med Bull. 1995;51:503–516. doi: 10.1093/oxfordjournals.bmb.a072976. [DOI] [PubMed] [Google Scholar]
- 43.Evans C H, Robbins P D. Curr Opin Rheumatol. 1996;8:230–234. doi: 10.1097/00002281-199605000-00011. [DOI] [PubMed] [Google Scholar]
- 44.Glauser M P, Zanetti G, Baumgartner J-D, Cohen J. Lancet. 1991;338:732–739. doi: 10.1016/0140-6736(91)91452-z. [DOI] [PubMed] [Google Scholar]
- 45.Paleolog E M, Hunt M, Elliott M J, Feldmann M, Maini R N, Woody J N. Arthritis Rheum. 1996;39:1082–1091. doi: 10.1002/art.1780390703. [DOI] [PubMed] [Google Scholar]
- 46.Crowe P D, Walter B N, Mohler K M, Otten-Evans C, Black R A, Ware C F. J Exp Med. 1994;181:1205–1210. doi: 10.1084/jem.181.3.1205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Makarov S S, Johnston W N, Olsen J C, Watson J M, Mondal K, Rinehart C, Haskill J S. Gene Ther. 1997;4:846–852. doi: 10.1038/sj.gt.3300461. [DOI] [PubMed] [Google Scholar]