

Abstract
Two ADP receptors have been identified on human platelets: P2Y1 and P2Y12. The P2Y12 receptor blocker clopidogrel is widely used to reduce the risks in acute coronary syndromes, but, currently, there is no P2Y1 blocker in clinical use. Evidence for variable responses to clopidogrel has been described in several reports. The mechanistic explanation for this phenomenon is not fully understood. The aim of this study was to examine mechanisms responsible for variability of 2MeS-ADP, a stable ADP analogue, induced platelet reactivity in clopidogrel-treated patients. Platelet reactivity was assessed by flow cytometry measurements of P-selectin (CD62P) and activated GpIIb/IIIa complex (PAC-1). Residual 2MeS-ADP activation via the P2Y12 and P2Y1 receptors was determined by co-incubation with the selective antagonists AR-C69931 and MRS2179 in vitro. P2Y1 and P2Y12 receptor expression on both RNA and protein level were determined, as well as the P2Y12 H1 or H2 haplotypes. Our data suggest that the residual platelet activation of 2MeS-ADP after clopidogrel treatment is partly due to an inadequate antagonistic effect of clopidogrel on the P2Y12 receptor and partly due to activation of the P2Y1 receptor, which is unaffected by clopidogrel. Moreover, a correlation between increased P2Y12 protein expression on platelets and decreased response to clopidogrel was noticed, r2=0.43 (P<0.05). No correlation was found between P2Y12 mRNA levels and clopidogrel resistance, indicating post-transcriptional mechanisms. To achieve additional ADP inhibition in platelets, antagonists directed at the P2Y1 receptor could be more promising than the development of more potent P2Y12 receptor antagonists.
Key words: AR-C 69931, clopidogrel, MRS 2179, P2Y1, P2Y12, platelets
Introduction
ADP receptors play an important role in the activation of platelets by acting as a positive feedback mechanism. There is evidence of two different ADP receptors expressed on human platelets: the Gq-coupled P2Y1 and the Gi-coupled P2Y12 [1, 2]. The P2Y12 antagonist clopidogrel has been shown to be an important platelet inhibitor. The Clopidogrel versus Aspirin in Patients at Risk of Ischaemic Events (CAPRIE) and Clopidogrel in Unstable Angina to Prevent Recurrent Events (CURE) studies demonstrate that inhibition of the P2Y12 receptor is beneficial, both alone and as a complement to standard aspirin treatment [3, 4]. At present, the standard regimen after a non-ST elevation myocardial infarction or coronary stenting is a combined treatment with both clopidogrel and aspirin.
However, evidence for a variable response to clopidogrel has recently been studied [5–7], and clopidogrel resistance has been associated with an increase in cardiovascular events [8]. The mechanisms underlying this phenomenon have not been fully elucidated. An association of clopidogrel resistance with the activity of the hepatic cytochrome CYP3A4 has been suggested [9]. Medication with atorvastatin has also been proposed to influence the metabolism of clopidogrel, by acting as a competitive substrate for CYP3A4 [10]. However, later studies have not been able to confirm this relationship [11–13]. Other potential explanations may be variations in receptor expression and function or variation in intracellular signaling pathways. A single nucleotide polymorphism (SNP) in the P2Y12 receptor associated with increased function of the receptor and increased risk for peripheral arterial disease (PAD) has been described and characterized [14, 15]. Again, a more recent study has not been able to confirm this association [16]. It is also plausible that the second ADP receptor expressed on human platelets, P2Y1, may be involved in the insufficient inhibition of ADP-mediated platelet activation after clopidogrel treatment.
We recently developed a real-time quantitative polymerase chain reaction (PCR) method for studies of transcriptional regulation of specific mRNA transcript in human platelets [1]. It has been useful in clinical studies demonstrating a down-regulation of P2Y12 receptor mRNA in patients with systemic lupus erythematosus (SLE) compared with healthy controls [17].
The aim of the present study was to investigate whether residual 2MeS-ADP, a stable ADP analogue, induced platelet reactivity after clopidogrel treatment due to unblocked P2Y1 or to insufficient inhibition of P2Y12 receptor activity. We were also interested in evaluating if variations in residual platelet reactivity after clopidogrel treatment could be correlated with receptor properties: P2Y12 expression, on mRNA or protein level, or to the P2Y12 haplotype.
Material and methods
Selection of patients
Our study group consisted of 19 patients (six women and 13 men) recently treated by percutaneous coronary intervention (PCI). Blood samples were collected during a visit to an out patient clinic, at least 2 weeks after the intervention. All patients were on both aspirin and clopidogrel and had a median age of 66 years (range 49–84 years). Eight had a history of previous acute myocardial infarction, and four had documented diabetes mellitus. There were 17 on statins and 14 on either angiotensin-converting enzyme (ACE) inhibitors or angiotensin II receptor blockers.
The Ethics Committee of Lund University approved the project. All patients had given written consent prior to participation in the study.
Blood collection and handling
For flow cytometry, blood was collected in vacutainer 4.5 ml citrate tubes. All experimental procedures were carried out as fast as possible and always within 10 min of blood collection. For RNA and protein analysis, blood was drawn in six vacuette 6 ml EDTA tubes. Platelet rich plasma (PRP) was then prepared by centrifugation at 160 g for 20 min at 4°C. PRP was carefully collected without disturbance of the buffy coat and was centrifuged again at 160 g for 10 min at 4°C to remove contaminating leukocytes. The PRP was then transferred to a new tube, and the platelets were collected by centrifugation at 1000 g for 10 min at 4°C. The platelet pellet was immediately dissolved in 1 ml TRIzol and frozen at −80°C.
Flow cytometric assessment of platelet activation
Flow cytometric studies were performed on a Becton and Dickinson FacScan. The following antibodies were used: fluorescein isothiocyanate (FITC)-conjugated anti-GPIIb/IIa (PAC-1) antibody, phycoerythrin (PE)-conjugated anti-P-selectin (CD62P) and peridinin chlorophyll-a protein (PerCP)-conjugated anti-CD61. 2MeS-ADP (1 μM ) was used as a stable P2Y agonist. We used the same concentration, 10 μM, of both antagonists, AR-C 69931 and MRS 2179, based on a dose-finding study made prior to the clinical study. AR-C 69931 is more potent (IC50 around 0.4 nM) than MRS 2179 (reported IC50 in human platelets147 nM), but the fact that the P2Y12 receptor in the clopidogrel-treated patient is already partly blocked changes this relationship [18, 19]. To achieve roughly the same amount of inhibition a more potent P2Y12 antagonist is needed. Five microliters of whole blood was incubated in phosphate buffered saline (PBS) solution, pH 7.4, containing agonist, appropriate antagonist and saturating concentrations of antibodies in a total volume of 50 μl for 20 min in the dark at room temperature. The reaction was then stopped by the addition of 3 ml 1% paraformaldehyde solution at 4°C followed by 20 min incubation at 4°C in the dark. Three-color flow cytometry was then performed. Forward scatter and side scatter were used to gate the platelet population. A total of 10,000 gated events was collected. To further exclude other cell types, we included only CD61 positive cells in the analysis. P-selectin positive platelets were quantified by determining the percentage of cells with fluorescence greater than mouse IgG control. For anti-GPIIb/IIIa antibody, PAC-1, the mean fluorescence intensities (MFIs) were measured. Inhibition of platelet activation was expressed as the relative inhibition of platelet activation after addition of drugs compared with stimulated platelets.
RNA and protein extraction
Total cellular RNA and protein was extracted using TRIzol reagent (Gibco BRL, Life Technology) according to the supplier’s instructions. The RNA concentration was determined spectrophotometrically, with a ratio of optical density (OD) 260:280>1.6 being considered as pure. Samples were stored at −70°C until required. The quality of RNA and protein was fully satisfactory in only 14 subjects. Protein pellets were vacuum dried and dissolved in 1% sodium dodecyl sulfate solution. DC protein assay (BioRad Laboratories, USA) was used to determine protein concentrations. Protein samples were stored at −20°C until required.
Quantitative analysis of P2 receptors by real-time reverse transcription polymerase chain reaction
Total RNA was reverse-transcribed using Multiscribe RT Kit (Qiagen, CA, USA) following the manufacturer’s instructions. Real-time PCR on platelet cDNA was performed using the following primers: P2Y12 5′-CTATCGATCGCTACCAGAAGAC-3′, 5′-TCATGCCAGACTAGACCGAA-3′, GAPDH 5′-GTCCACTGGCGTCTTCACCA-3′, 5′-GTGGCAGTGATGGCATGGAC-3′. Relative gene expression levels were determined using LightCycler (Roche, Basle, Switzerland). As a template, 1 μl platelet cDNA was used in a total volume of 10 μl [3 mM MgCl2, 0.5 μM of each primer (described below) and 1×LightCycler DNA Master SYBR Green I mix (Roche)]. The following PCR amplification protocol was used: initial denaturation at 95°C for 600 s, followed by 45 PCR cycles at 95°C for 1 s, 55°C (P2Y12) or 58°C (GAPDH) respectively for 6 s, and 74°C for 23 s. Target gene amplification levels were quantified using a second-derivative maximum method available in the LightCycler software. P2Y12 receptor expression was determined using the housekeeping gene GAPDH as a reference. To validate specificity of amplification reactions, the LightCycler PCR products were sequenced using BigDye v.3.1 (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s instructions.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blotting
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting were performed as described elsewhere [20]. Briefly, 20 μg protein per lane was separated using 10% SDS-polyacrylamide gels and transferred onto Immobilon-P membranes (Millipore, Bedford, MA, USA). Sequential detection of P2Y1 and P2Y12 (at 1:200 dilution) and GAPDH (at 1:500 dilution) was performed according to the following procedure in the above-mentioned order. Membranes were incubated for 1 h at room temperature with the appropriate primary antibody, washed three times with TBS 0.1% Tween, incubated for 1 h with HRP-conjugated secondary antibodies, washed three times and visualized using the ECL western blotting system and Hyperfilm. After each round of immunodetection, the membranes were incubated for 30 min at 50°C in stripping buffer (100 mM 2-mercapto-ethanol, 2% SDS, 62.5 mM Tris-HCl, pH 6.7) to remove previously bound antibodies, washed twice in TBS 0.1 % Tween, blocked with 5% fat-free milk in TBS 0.1% Tween and used again for the next set of antibodies.
The signal intensity (integral volume) of the appropriately sized bands was scanned and analyzed using Quantity One software (BioRad). The signal intensities of the different P2 receptors were normalized to GAPDH to account for variations in the amounts of total protein present in each lane.
Determination of P2Y12 haplotype
DNA was isolated from whole blood using the TRIzol reagent according to the manufacturer’s instructions. A PCR using HotStarTaq (Qiagen) and the primers 5′-GAGCACTCAAGACTTTACTGACG-3′ and 5′-GAGAAGATCAGAAATGACTGTGTTC-3′ was used to amplify the entire second and parts of the third exon of the of the P2Y12 gene. The 2233 bp amplification product, containing two TaiI restriction sites, [at polymorphism i-T744C (1008 bp from the 5′ end of the amplification product)] and at a control TaiI site (at 1588 bp), was diluted three times with water, digested for 1 h with 2U TaiI at 65°C in a total volume of 30 μl and visualized by agarose gel electrophoresis. The P2Y12 H1 allele was identified by the presence of two bands (1588 bp- and 645 bp-long restriction fragments), and the H2 allele by three bands (1008 bp-, 580 bp- and 465 bp-long restriction fragments) [13].
Reagents, antibodies and drugs
Unless otherwise stated all reagents and drugs were purchased from Sigma Chemical Corp., St. Louis, MI, USA. PCR and RT-PCR consumables were purchased from Perkin-Elmer Applied Biosystems Inc., Foster City, CA, USA. PCR primers were from MWG Biotech AG, Ebersberg, Germany and TRIzol from Invitrogen, Carlsbad, CA, USA. Western blot reagents were from Amersham Pharmacia Biotech, UK, or Bio-Rad Laboratories, USA. Anti-P2Y1 and anti- P2Y12 antibodies were purchased from Alomone Labs, Israel, and the anti-GAPDH was from Chemicon, USA. Anti-CD61, anti-CD62P and anti-PAC1 antibodies were from BD, USA. MRS 2179 was from Tocris, and AR-C 69931 was a kind gift from Astra-Zeneca, Sweden. All drugs were dissolved in 0.9% saline solution.
Statistics and analysis
Calculations and statistics were performed using GraphPad Prism 4.00 software. Repeated-measures analysis of variance (ANOVA) with Bonferroni’s post-test was used to compare flow cytometry data. Values are presented as mean ± SEM. Comparison of quantitative real-time data was performed using the method described by Pfaffl [21]. The amplification efficiencies of each primer pair were tested prior to analysis.
Results
Flow cytometry assessment of platelet activation
After stimulation of platelets with 1 μM 2MeS-ADP, additional antagonism of the P2Y12 receptor with AR-C 69931 (10 μM), to platelets from patients already treated with clopidogrel and aspirin, inhibited the residual percentage of P-selectin positive platelets by 35.8±3.5% (mean ± SEM) (n=19) and the expression of activated GPIIb/IIIa by 43.5±3.4%. The blocking of the P2Y1 receptor with MRS 2179 (10 μM) resulted in a significantly greater inhibition of the percentage of P-selectin positive platelets by 67.3±4.4% and of activated GPIIb/IIIa to 54.2±2.6%. Combining the antagonists yielded an even greater significant inhibition: 78.4±3.8% for P-selectin and 72.2±2.5% for GPIIb/IIIa. Data are presented in Figure 1.
Figure 1.
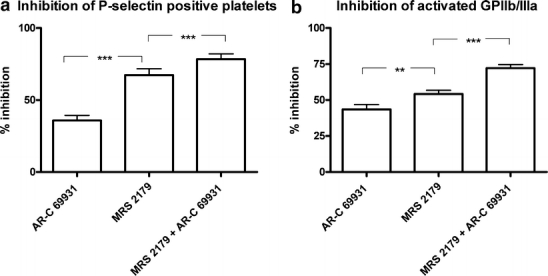
Graph showing the increased inhibition of residual ADP-induced P-selectin positive platelets (a) and expression of activated GPIIb/IIIa (b) after the further blocking of the P2Y12 receptor with AR-C 69931 (10 μM) and the P2Y1 receptor with MRS 2179 (10 μM) and the blocking of both receptors with a combination of both antagonists. These effects are seen in platelets already blocked by clopidogrel and aspirin. Blocking the P2Y12 receptor further with AR-C 69931 (10 μM) significantly inhibited the percentage of P-selectin positive platelets by 35.8±3.5% (mean ± SEM) (n =19) and the expression of activated GPIIb/IIIa by 43.5±3.4%. A significantly greater inhibition was seen when the P2Y1 antagonist MRS 2179 (10 μM) was used, where the inhibition of P-selectin increased to 67.3±4.4% and the inhibition of activated GPIIb/IIIa increased to 54.2±2.6%. Combining the antagonists yielded an even greater significant inhibition: 78.4±3.8% for P-selectin and 72.2±2.5% for activated GPIIb/IIIa. There were significance differences between the groups, P <0.01, in all cases (n =19). Repeated-measures ANOVA with Bonferroni’s post-test were used to compare data
Correlation of function with protein and RNA expression
Proteins were run on western blot, and bands from the P2Y12 receptor and GAPDH were detected. Signal intensities were quantified, and each P2Y12 reading was normalized to the GAPDH expression in the same sample to correlate with different loading amounts. The relative normalized expression of P2Y12 protein correlated with the percentage of cells expressing P-selectin after stimulation by 2MeS-ADP, with a linear regression of r2=0.43 (P <0.05) (n =14). No correlations were found between P2Y1 protein levels and P-selectin positive platelets (r2=0.002, P =0.88, n =14). Data are presented in Figure 2.
Figure 2.
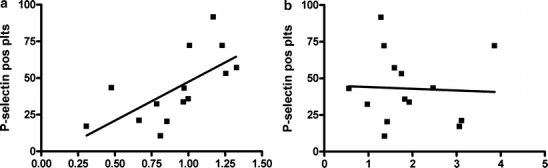
The relative expression and GAPDH-normalized expression of P2Y12 (a) and P2Y1 (b) protein correlated with the percentage of P-selectin positive platelets after stimulation with 1 μM of 2MeS-ADP. In (a) there is a significant correlation of P-selectin positive platelets with P2Y12 protein expression, with r2=0.43 (P <0.05) (n =14). In (b) there is no significant correlation of P-selectin positive platelets and P2Y1 protein expression (r2=0.002, P=0.88, n=14) (pos plts positive platelets)
P2Y12 and GAPDH mRNA expressions were quantified, and no correlation was found between the relative expression levels of the P2Y12 mRNA and the percentage of cells expressing P-selectin or activated GPIIb/IIIa after stimulation with 2MeS-ADP (r2=0.30, P=0.20 for P-selectin; r2=0.04, P=0.19 for GPIIb/IIIa) (data not shown).
P2Y12 haplotype and function
The haplotype was determined by PCR sequencing and restriction digestion. Thirteen patients were homozygous for the H1 haplotype (H1/H1) and six patients were heterozygous (H1/H2), whereas none of the patients was homozygous for the H2 haplotype. There was no difference in the percentage of platelets expressing P-selectin after 2MeS-ADP (1 μM) in the homozygous (H1/H1) group compared with the heterozygous (H1/H2) group (45.2±6.4% for H1/H1 vs 37.0±9.0% for H1/H2, P=0.47) (unpaired t-test).
Discussion
This study suggests that the residual 2MeS-ADP platelet reactivity after clopidogrel treatment is in part due to an inadequate antagonistic effect of clopidogrel on the P2Y12 receptor and partly due to the P2Y1 receptor, which is unaffected by clopidogrel. Moreover, a correlation between increased P2Y12 protein expression on platelets and decreased response to clopidogrel was noticed. No correlation was seen between platelet activation and P2Y12 mRNA expression or the P2Y12 H1/H2 haplotype.
Our cohort of patients was chosen to reflect the real clinical situation as much as possible, resulting in a heterogeneous group having other illnesses and taking several other medications apart from clopidogrel and aspirin. The cohort was too small for us to be able to do any subgroup analyses in terms of other medication, previous diseases or gender. Since all of the patients were on aspirin, the impact of reduced prostaglandin synthesis was present throughout. Despite the inhibition by both clopidogrel and aspirin, we were able to activate their platelets further with 2MeS-ADP. The remaining 2MeS-ADP activation was dependent on both insufficient P2Y12 blockade by clopidogrel and the unaffected ADP receptor P2Y1. Most likely, many different factors contribute to produce the observed clinical phenotype. One of the most important factors is still the second ADP receptor, P2Y1, that remains totally unaffected by clopidogrel. To achieve full ADP inhibition in the future, increasing the effect of clopidogrel or other P2Y12 blockers will not suffice. Thus, the clinical benefit of a P2Y1 blocker will be an interesting alternative.
Despite the use of both AR-C 69931 and MRS 2179, platelets were not completely inhibited after 2MeS-ADP stimulation. This was probably due to the very high concentration of agonist used to stimulate the system; 1 μM of 2MeS-ADP corresponds to a significantly higher dose of unmodified ADP. The platelet stimulation was also conducted in whole blood, where leukocytes and erythrocytes may also influence the degree of platelet stimulation.
Resistance to the COX-1 inhibitor acetylsalicylic acid (aspirin) has been studied and debated for many years. Small observational studies suggest that non-responders to aspirin are at greater risk of cardiovascular events than are responders [22–25]. Recently, resistance to clopidogrel has emerged as an interesting occurrence with a clear clinical relevance [6–8]. The fact that clopidogrel resistance has an impact on the cardiovascular outcome in patients with coronary artery disease merits further studies to elucidate the underlying mechanisms [8]. Theoretically, many factors may influence the clopidogrel response: bioavailability, genetic or transcriptional variations in the metabolizing enzymes or in the target receptor and interactions with other compounds. One early study demonstrated a correlation between clopidogrel resistance and atorvastatin medication, but this hypothesis was later dismissed by several other studies [11–13]. Currently, two studies have investigated possible mechanisms underlying clopidogrel resistance. One study was focused on the hepatic metabolism of the drug, and the other was focused on the bioavailability of clopidogrel [9, 26]. The bioavailability and the hepatic metabolism of the pro-drug clopidogrel do seem to be important for clopidogrel resistance [9, 26]. For the first time we find that increased platelet P2Y12 receptor protein expression is a component of clopidogrel resistance. Increased abundance of a receptor protein may thereby result in less inhibition by its antagonist. However, whether these patients are constituent high P2Y12 expressers that increase their risk of myocardial infarction is not known. Animal studies have shown a correlation between function and overexpression of P2Y1 protein, but we could not find this association in our clinical material [27].
Platelets are anuclear cells and are therefore unable to produce new nuclear-derived mRNA transcripts. However, it has been shown that platelets contain megakaryocyte-derived mRNA that can be activated upon platelet stimulation to produce, for example, IL1-β and PAI-1 [28, 29]. Thus, translational regulation can account for the variation in protein expression. Interestingly, there was no correlation between P2Y12 mRNA levels and variation of clopidogrel inhibition. These results suggest that post-transcriptional regulation of the P2Y12 receptor expression is responsible for the correlation between P2Y12 protein expression and the clopidogrel effect. Also, mRNA concentrations in platelets are low, and any mRNA correlation that might have existed in the megakaryocyte may have been lost in circulating platelets.
Haplotype studies have suggested that the P2Y12 H2 haplotype may have a higher mRNA expression than the H1 haplotype, and that this would explain the functional differences between the different genotypes [14]. In our small sample group we could not find any such correlation, and our results are in concordance with a larger P2Y12 haplotype study by Hetherington et al. [16]. It should be emphasized that our sample is small and it is therefore hard to draw any reliable conclusions between polymorphisms and function.
There are several P2Y12 antagonistic compounds in early and late clinical trials at the moment: prasugrel, cangrelor, AZD6140. It will be very interesting to see if these will encounter the same response variation problems as clopidogrel has. It might also be that different drugs will be better suited for different clinical phenotypes. With many different P2Y12 antagonists available in the clinic, a simple clinical test evaluating patient phenotype would be useful in choosing the optimal platelet inhibitor. This study also shows the relevance of designing drugs targeting the P2Y1 receptor.
Conclusion
Variation of clopidogrel inhibition is partly due to the unblocked P2Y1 receptor and partly due to a variable antagonistic effect of clopidogrel on the P2Y12 receptor. A correlation between increased P2Y12 protein expression on platelets and decreased response to clopidogrel was seen, indicating a novel explanation to the clopidogrel resistance phenomenon.
Acknowledgments
A special thanks to Goran Olivecrona for helpful linguistic guidance. The study was supported by the Swedish Heart and Lung Foundation, the Franke and Margareta Bergqvist Foundation, the Bergwall Foundation, the Zoegas Foundation and Swedish Medical Research Council Grant 13130.
References
- 1.Wang L, Ostberg O, Wihlborg A-K, Brogren H, Jern S, Erlinge D (2003) Quantification of ADP and ATP receptor expression in human platelets. J Thromb Haemost 1:328–334 [DOI] [PubMed]
- 2.Kunapuli SP, Dorsam RT, Kim S, Quinton TM (2003) Platelet purinergic receptors. Curr Opin Pharmacol 3:175–180 [DOI] [PubMed]
- 3.Yusuf S, Zhao F, Mehta SR, Chrolavicius S, Tognoni G, Fox KK (2001) Effects of clopidogrel in addition to aspirin in patients with acute coronary syndromes without ST-segment elevation. N Engl J Med 345:494–502 [DOI] [PubMed]
- 4.CAPRIE Steering Committee (1996) A randomised, blinded, trial of clopidogrel versus aspirin in patients at risk of ischaemic events (CAPRIE). Lancet 348:1329–1339 [DOI] [PubMed]
- 5.Gurbel PA, Bliden KP, Hiatt BL, O’Connor CM (2003) Clopidogrel for coronary stenting: response variability, drug resistance, and the effect of pretreatment platelet reactivity. Circulation 107:2908–2913 [DOI] [PubMed]
- 6.Jaremo P, Lindahl TL, Fransson SG, Richter A (2002) Individual variations of platelet inhibition after loading doses of clopidogrel. J Intern Med 252:233–238 [DOI] [PubMed]
- 7.Muller I, Besta F, Schulz C, Massberg S, Schonig A, Gawaz M (2003) Prevalence of clopidogrel non-responders among patients with stable angina pectoris scheduled for elective coronary stent placement. Thromb Haemost 89:783–787 [PubMed]
- 8.Matetzky S, Shenkman B, Guetta V, Shechter M, Bienart R, Goldenberg I, et al (2004) Clopidogrel resistance is associated with increased risk of recurrent atherothrombotic events in patients with acute myocardial infarction. Circulation 109:3171–3175 [DOI] [PubMed]
- 9.Lau WC, Gurbel PA, Watkins PB, Neer CJ, Hopp AS, Carville DG, et al (2004) Contribution of hepatic cytochrome P450 3A4 metabolic activity to the phenomenon of clopidogrel resistance. Circulation 109:166–171 [DOI] [PubMed]
- 10.Lau WC, Waskell LA, Watkins PB, Neer CJ, Horowitz K, Hopp AS, et al (2003) Atorvastatin reduces the ability of clopidogrel to inhibit platelet aggregation: a new drug–drug interaction. Circulation 107:32–37 [DOI] [PubMed]
- 11.Saw J, Steinhubl SR, Berger PB, Kereiakes DJ, Serebruany VL, Brennan D, et al (2003) Lack of adverse clopidogrel–atorvastatin clinical interaction from secondary analysis of a randomized, placebo-controlled clopidogrel trial. Circulation 108:921–924 [DOI] [PubMed]
- 12.Mitsios JV, Papathanasiou AI, Rodis FI, Elisaf M, Goudevenos JA, Tselepis AD (2004) Atorvastatin does not affect the antiplatelet potency of clopidogrel when it is administered concomitantly for 5 weeks in patients with acute coronary syndromes. Circulation 109:1335–1338 [DOI] [PubMed]
- 13.Wienbergen H, Gitt AK, Schiele R, Juenger C, Heer T, Meisenzahl C, et al (2003) Comparison of clinical benefits of clopidogrel therapy in patients with acute coronary syndromes taking atorvastatin versus other statin therapies. Am J Cardiol 92:285–288 [DOI] [PubMed]
- 14.Fontana P, Dupont A, Gandrille S, Bachelot-Loza C, Reny JL, Aiach M, et al (2003) Adenosine diphosphate-induced platelet aggregation is associated with P2Y12 gene sequence variations in healthy subjects. Circulation 108:989–995 [DOI] [PubMed]
- 15.Fontana P, Gaussem P, Aiach M, Fiessinger JN, Emmerich J, Reny JL (2003) P2Y12 H2 haplotype is associated with peripheral arterial disease: a case–control study. Circulation 108:2971–2973 [DOI] [PubMed]
- 16.Hetherington SL, Singh RK, Lodwick D, Thompson JR, Goodall AH, Samani NJ (2005) Dimorphism in the P2Y1 ADP receptor gene is associated with increased platelet activation response to ADP. Arterioscler Thromb Vasc Biol 25:252–257 [DOI] [PubMed]
- 17.Wang L, Erling P, Bengtsson AA, Truedsson L, Sturfelt G, Erlinge D (2004) Transcriptional down-regulation of the platelet ADP receptor P2Y(12) and clusterin in patients with systemic lupus erythematosus. J Thromb Haemost 2:1436–1442 [DOI] [PubMed]
- 18.Burnstock G, Williams M (2000) P2 purinergic receptors: modulation of cell function and therapeutic potential. J Pharmacol Exp Ther 295:862–869 [PubMed]
- 19.Pillarisetti K, Toomey J, Erhardt J (2003) ADP mediated platelet–leukocyte interactions require activation of both P2Y1 and P2Y12 receptors. In: Carrico C (ed) Mid-Atlantic Pharmacology Society meeting, 2003. The American Society for Pharmacology and Experimental Therapeutics, 2003; p 139
- 20.Wang L, Karlsson L, Moses S, Hultgardh-Nilsson A, Andersson M, Borna C, et al (2002) P2 receptor expression profiles in human vascular smooth muscle and endothelial cells. J Cardiovasc Pharmacol 40:841–853 [DOI] [PubMed]
- 21.Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:e45 [DOI] [PMC free article] [PubMed]
- 22.Chen WH, Lee PY, Ng W, Tse HF, Lau CP (2004) Aspirin resistance is associated with a high incidence of myonecrosis after non-urgent percutaneous coronary intervention despite clopidogrel pretreatment. J Am Coll Cardiol 43:1122–1126 [DOI] [PubMed]
- 23.Mueller MR, Salat A, Stangl P, Murabito M, Pulaki S, Boehm D, et al (1997) Variable platelet response to low-dose ASA and the risk of limb deterioration in patients submitted to peripheral arterial angioplasty. Thromb Haemost 78:1003–1007 [PubMed]
- 24.Grotemeyer KH, Scharafinski HW, Husstedt IW (1993) Two-year follow-up of aspirin responder and aspirin non-responder. A pilot-study including 180 post-stroke patients. Thromb Res 71:397–403 [DOI] [PubMed]
- 25.Gum PA, Kottke-Marchant K, Welsh PA, White J, Topol EJ (2003) A prospective, blinded determination of the natural history of aspirin resistance among stable patients with cardiovascular disease. J Am Coll Cardiol 41:961–965 [DOI] [PubMed]
- 26.Taubert D, Kastrati A, Harlfinger S, Gorchakova O, Lazar A, von Beckerath N, et al (2004) Pharmacokinetics of clopidogrel after administration of a high loading dose. Thromb Haemost 92:311–316 [DOI] [PubMed]
- 27.Hechler B, Zhang Y, Eckly A, Cazenave JP, Gachet C, Ravid K (2003) Lineage-specific overexpression of the P2Y1 receptor induces platelet hyper-reactivity in transgenic mice. J Thromb Haemost 1:155–163 [DOI] [PubMed]
- 28.Brogren H, Karlsson L, Andersson M, Wang L, Erlinge D, Jern S (2004) Platelets synthesize large amounts of active plasminogen activator inhibitor 1. Blood 104:3943–3948 [DOI] [PubMed]
- 29.Lindemann S, Tolley ND, Dixon DA, McIntyre TM, Prescott SM, Zimmerman GA, et al (2001) Activated platelets mediate inflammatory signaling by regulated interleukin 1beta synthesis. J Cell Biol 154:485–490 [DOI] [PMC free article] [PubMed]