

Abstract
It is well established that several cell surface receptors interact with each other to form dimers and oligomers, which are essential for their activation. Since little is known about the quaternary structure of P2Y receptors, in the present work, we investigated the expression of the G-protein-coupled P2Y4 subunit as monomeric or higher-order complex protein. We examined both endogenously expressed P2Y4 subtype with the aid of specific anti-P2Y4 antiserum, and heterologously transfected P2Y4-tagged receptors with the use of antitag antibodies. In both cases, we found the P2Y4 receptor displaying molecular masses corresponding to monomeric, dimeric and oligomeric structures. Experiments performed in the absence of reducing agents demonstrated that there is a strict correlation among the multiple protein bands and that the multimeric forms are at least partially assembled by disulphide bonds. The direct demonstration of P2Y4 homodimerisation comes instead from co–transfection and differential co–immunoprecipitation experiments, with the use of differently tagged P2Y4 receptors and antitag antibodies. The structural propensity of the P2Y4 protein to form homo-oligomers may open the possibility of a novel regulatory mechanism of physiopathological functions for this and additional P2Y receptors.
Key words: G-protein-coupled receptors, PC12 cells, purinergic receptor, receptor dimerisation, SH-SY5Y cells
Introduction
Molecular cloning and heterologous receptor expression studies have led to the identification and characterisation of eight different P2Y receptor subtypes (P2Y1,2,4,6,11–14) [1]. P2Y1,2,4,6,11 receptors display 25–52% amino acid identity and couple, via heterotrimeric G proteins of the Gq family, to activation of phospholipase C, generation of inositol phosphates, and mobilisation of intracellular Ca2+ stores [2, 3]. Only the P2Y11 receptor also couples to Gs proteins to activate adenylyl cyclase and promote cyclic AMP accumulation [4–6]. The recently identified P2Y12–14 receptors have high sequence identity with each other (40–48%), but share relatively little identity (22–25%) with the other P2Y receptors. Moreover, differently from P2Y1,2,4,6,11 receptors, they are associated with Gi proteins, to inhibit adenylyl cyclase [7, 8]. P2Y receptors can alternatively be classified into purinergic and pyrimidinergic subclasses. In particular, P2Y1,11,12,13,14 receptors are activated by adenine nucleotides [1]; P2Y2 and rat P2Y4 subunits display no selectivity between ATP and UTP [9]; human P2Y4 (hP2Y4) and P2Y6 subtypes are mainly pyrimidinergic [10], but with ATP behaving as a potent competitive antagonist at the hP2Y4 subunit [11].
One of the most broadly expressed P2Y receptor in various brain regions is P2Y4. Highest mRNA levels are present in the cerebellum, hypothalamus, medial frontal gyrus, parahippocampal gyrus, striatum and thalamus; lowest expression levels were detected in the amygdala and spinal cord [12]. Moreover, P2Y4 mRNA was found in the dorsal root, nodose and trigeminal ganglion [13]. In situ hybridisation studies confirmed a similar wide distribution, with highest incidence in the pineal gland and ventricular system [14]. At the protein level, the presence of the P2Y4 receptor was demonstrated by immunohistochemistry in sensory ganglia [13], the brainstem and ventral white matter of the spinal cord during rat brain development [15].
Although according to amino acid sequence the predicted molecular mass of the P2Y4 protein is 41 kDa [14], Western blot studies identified a protein corresponding to 88 kDa in rat cerebellar granule neurons [16, 17]. This same high molecular weight was demonstrated in glutamatergic neurons from rat hippocampus, in astrocytoma 1321N1 cells stably transfected with P2Y4 receptor [18] and in NGF-differentiated PC12 cells [19]. In rat oligodendrocyte precursors [20] and rat cortical astrocytes [21], the 88 kDa protein was associated with a lower molecular mass band, of about 45 kDa, while in gerbil brain, the P2Y4 receptor displayed three bands (respectively, 75, 55 and 36 kDa) [22]. While classical studies postulated that G-protein-coupled receptors (GPCRs) exist only as monomers, many recent findings instead demonstrated that many GPCRs form dimers and oligomers [23]. For example, it was recently demonstrated that P2Y1 receptor forms heterocomplexes with A1 proteins [24–26] and that P2Y4 coimmunoprecipitates with NMDA receptors [17].
Since only little is known specifically on the quaternary structure of P2Y receptors, the purpose of the present study was to analyse the potential expression of the P2Y4 subunit in monomeric and higher-order complex proteins. The attainment of this knowledge could be of great importance in understanding the regulation and function not only of the P2Y4 subtype but also of additional P2Y receptors.
Materials and methods
Cell line cultures
Rat phaeochromocytoma PC12nnr5 cells were cultured on collagen-coated dishes in RPMI 1640 (Gibco BRL, MI, Italy) medium supplemented with 10% heat-inactivated horse serum and 5% foetal calf serum (FCS) (Gibco BRL). Human neuroblastoma SH-SY5Y cells were grown in Dulbecco’s modified Eagle’s medium (DMEM)/F12 (Sigma-Aldrich, MI, Italy) supplemented with 10% FCS. All culture media were supplemented with glutamine (2 mM), penicillin (50 units/ml), and streptomycin (50 µg/ml), and all cell lines were grown at 37 °C in 5% CO2.
Dissociated primary cell cultures
Cerebellar granule neurons (CGN) from Wistar 8-day-old rat cerebellum were prepared as described [27] and seeded on poly-L-lysine-coated dishes, in Eagle’s basal medium (BME) (Gibco BRL), with 25 mM KCl, 2 mM glutamine, 0.1 mg/ml gentamycin, and 10% heat inactivated FCS. At 1 day in vitro, cultures were supplemented with 10 µM cytosine arabinoside and kept for 9 days, without replacing the culture medium.
Construction of epitope-tagged human P2Y4 receptor and transient transfection
The plasmid containing the cDNA encoding for human P2Y4 was a kind gift from Prof. T.K. Harden. cDNA was digested with EcoRI and XhoI restriction enzymes and ligated into a pCMV expression vector that incorporated a FLAG epitope tag (DYKDDDDK) at the 5′ terminus. Moreover, by the use of polymerase chain reaction (PCR) with Platinum Pfx polymerase (Invitrogen, Paisley, UK), human P2Y4 cDNA was amplified with oligonucleotides containing EcoRI and XhoI restriction sequences for the subcloning into a pCS2 plasmid containing myc epitope tags (EQKLISEEDL) at the 5′ terminus. All constructs were verified by sequencing analysis. SH-SY5Y cells in 35-mm plates were transiently transfected with epitope-tagged P2Y receptor cDNAs individually or in combination. Cells were incubated with 4 µg of total DNA and 4 µl of Lipofectamine™ 2000 (Invitrogen) in 2 ml of serum-free medium (Opti-MEM, Invitrogen) for 24h.
Cell protein extraction
In order to isolate total protein extracts, cells were harvested with ice-cold RIPA buffer [phosphate-buffered saline (PBS), 1% Nonidet P-40-NP-40-, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulphate (SDS)], whereas for immunoprecipitation experiments, cells were lysed in buffer G (1% Triton X-100, 10% glycerol, 150 mM NaCl, 1,5 mM MgCl2, 1 mM CaCl2, 1 mM EGTA). Both buffers were added with 1 mM phenylmethylsulphonyl fluoride (PMSF) and 10 µg/ml leupeptin. Cellular lysates were kept for 30 min on ice and then centrifuged for 10 min at 14,000 × g at 4°C. Supernatants were collected and assayed for protein quantification by the Bradford method [28].
Immunoprecipitation
Purified cell lysates (100 µg) were combined with 15 µl (packed gel) of either anti-c-Myc or anti-FLAG M2 affinity agarose (Sigma-Aldrich) and kept for 2 h at 4 °C on a rotator. The immunoadsorbents were recovered by centrifugation for 2 min at 3,000 × g and washed three times by resuspension centrifugation for 2 min at 3,000 × g in buffer G. The samples were eluted in 30 µl of sample buffer used for SDS-polyacrylamide gel electrophoresis (PAGE).
SDS-PAGE and Western blotting analysis
Analysis of protein components was performed on 10% polyacrylamide gels, as described (Laemmli, 1970), loading the same amount of protein for each sample. Gels were transferred onto nitrocellulose membranes (Amersham Biosciences, Cologno Monzese, Italy), and blots were probed overnight at 4 °C, with anti-P2Y4 antiserum (Alomone Labs, Jerusalem, Israel), (1:300), which specificity was directly tested by immunoreactions in the presence of the neutralising peptide (ratio 1:1 between peptide and antiserum) or anti c-myc (9E10) antibody (Chemicon International Inc., Temecula, CA, USA), (1:200). Subsequently, blots were incubated for 1 h with horseradish peroxidase-coupled anti-rabbit (1:5,000) or anti-mouse (1:2,500) antibodies (Cell Signaling Technology, Inc., Beverly, MA, USA), and visualised using electrogenerated chemiluminescence (ECL) (Amersham Biosciences). Image analysis and quantifications were performed by Kodak Image Station (KDS IS440CF 1.1) with 1-dimensional (1D) image analysis software.
Immunofluorescence analysis
SH-SY5Y cells were fixed in 4% paraformaldehyde for 15 min, washed three times with PBS, permeabilised with 0.2% Triton X-100 for 5 min, rinsed with PBS and incubated with PBS containing 0.1% bovine serum albumin (BSA) for 30 min. Anti-c-Myc antibody was added at a dilution of 1:200 in PBS/BSA for 1 h. Cells were washed and stained with Cy2-conjugated donkey anti-mouse IgG (1:100, Jackson Immunoresearch, West Grove, PA, USA) for 1 h, rinsed again and coverslipped with gel/mount− anti-fading medium (Biomeda, Foster City, CA, USA). Immunofluorescence was analysed by means of a confocal laser scanning microscope (CLSM) (LSM 510, Carl Zeiss, Arese, MI, Italy) equipped with an argon laser emitting at 488 nm, a helium/neon laser emitting at 543 nm and a helium/neon laser emitting at 633 nm.
Intracellular [Ca2+] measurements
Changes in intracellular calcium levels were investigated by a microfluorometry imaging technique. Briefly, cells were grown on glass coverslips to ∼80% confluence and transfected with pGreen Lanterni™-1 (Gibco BRL) together with wild-type P2Y4 or tagged P2Y4 (1:8 molar ratio). After 24 h, cells were loaded with 1 mM Fura-2/AM for 30 min, washed and placed in a recording chamber mounted on the stage of an upright microscope (Axiscop2 FS, Carl Zeiss, AG, Germany) equipped for infrared video microscopy and microfluorimetry (Till Photonics GmbH, Gräfelfing, Germany). The coverslip was continuously superfused at 2.5 ml/min with artificial cerebrospinal fluid (NaCl 126 mM, KCl 2.5 mM, NaH2PO4 1.2 mM, MgCl2 1.2 mM, CaCl2 2.4 mM, glucose 10 mM, NaHCO3 19 mM, pH 7.4, equilibrated with a mixture of O2 95% / CO2 5%). UTP (100 µM) was delivered to the cells by bath applications in the extracellular medium through a three-way tap syringe for 2 min. An area of interest containing green fluorescent protein (GFP) positive cells was chosen and excited for 10 ms alternatively at 340 and 380 nm wavelength for Fura-2 stimulation at 0.5 Hz. The emitted fluorescence was acquired and digitalised by a chargecoupled device (CCD) camera (Photonic Science, Millham, UK). For off line analysis of the images, four regions of interest (ROI) that included the fluorescence areas coming from the cell body of selected GFP-positive cells were measured. Each image was background subtracted, and a ratio of F340/F380 was calculated.
Statistical analysis
Statistical analysis was carried out using a two-way ANOVA, followed by a post-hoc (Newman-Keuls) test for multiple comparisons. A value of p < 0.05 was considered significant: *p < 0.05, **p < 0.01, ***p < 0.001.
Results
Different neuronal cell lines display two protein bands for P2Y4 receptor
One of the earliest evidence that a receptor might form dimers or multimers is the presence of multiple bands on Western blot analysis. Therefore, we performed Western blots of total protein extracted from rat phaeochromocytoma PC12 cell variant nnr5 and human neuroblastoma SH-SY5Y cell line, probed with anti-P2Y4 antiserum (Figure 1). In both cell lines, we found the presence of a low-molecular-mass band matching the predicted monomeric P2Y4 receptor (41 kDa) and a higher-order band with molecular mass of about 80 kDa. The occurrence of the two bands is obtained by standard reducing [50 mM dithiothreitol (DTT) and sample boiling] and denaturing (SDSPAGE) conditions. Moreover, both bands raised specific signals, as evidenced by the lack of immunoreaction after preadsorption of the P2Y4 antiserum with the antigen peptide. These data support the hypothesis that both rat and human P2Y4 receptors form stable dimers.
Figure 1.
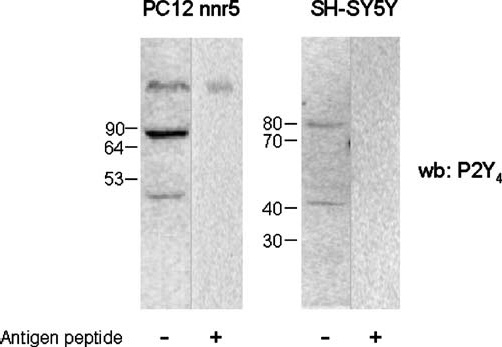
Western blotting detection of P2Y4 receptor in PC12nnr5 and SH-SY5Y cells. Total protein isolated from PC12nnr5 and SH-SY5Y were subjected to sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) on 10% polyacrylamide gel and transferred onto nitrocellulose membrane. Filters were probed with anti-P2Y4 antiserum in the absence (±) or presence (+) of the antigen peptide. Protein bands were detected with secondary anti-rabbit antiserum coupled to horseradish peroxidase by electrogenerated chemiluminescence (ECL)
Tagged P2Y4 receptor is functional and displays multiple protein bands
In order to confirm the existence of a high-molecular-mass complex of the P2Y4 receptor, bypassing the employment of the P2Y4 polyclonal antiserum (even though affinity purified on immobilised antigen), we constructed tagged P2Y4 receptors to be expressed in SH-SY5Y cells and identified with the aid of tag-specific antibodies. Therefore, after transient transfection of SH-SY5Y cells with Myc-P2Y4 cDNA, we first proved by immunofluorescence that the receptor was indeed expressed (we found expression in 25–30% of total cells, data not shown) and displayed a distinguishable cytoplasmic and membrane localisation (Figure 2a). We next evaluated whether the epitope-tagged subunits retained their functional integrity by intracellular Ca2+ mobilisation measurements (Figure 2b–e). The addition of 100 µM UTP gave a slight Ca2+ transient in control SH-SY5Y cells (probably due to naive P2Y4 and P2Y6 receptors) (Figure 2b) while it determined a marked increase in intracellular Ca2+ in cells expressing tagged P2Y4 receptors (Figure 2d, e), with kinetic properties similar to their P2Y4-transfected wild-type parent (Figure 2c, e). In addition, the immunoprecipitation of transfected Myc-P2Y4 with anti-c-Myc antiserum (developed in rabbit) and further detection with anti-c-Myc (9E10) antibody (developed in mouse) displayed three signals, which were instead completely absent in the mocktransfected sample (Figure 3). As for endogenously expressed P2Y4, we found a low-molecular-mass signal at approximately 49 kDa, corresponding to the monomeric Myc-P2Y4 receptor and a higher molecular band (about 100 kDa), which again resembles a potential dimeric form of the P2Y4 subunit. This band seems peculiar for the P2Y4 receptor since transfected Myc-P2Y1, -P2Y2, -P2Y6 and - P2Y11 immunoprecipitated by the same anti-c-Myc antibody used for Myc-P2Y4 did not show any comparable denaturing- and reducing-resistant signal (data not shown). Moreover, in the upper part of the gel, we also found a diffuse intense signal that we predict might correspond to oligomeric forms of the P2Y4 receptor.
Figure 2.
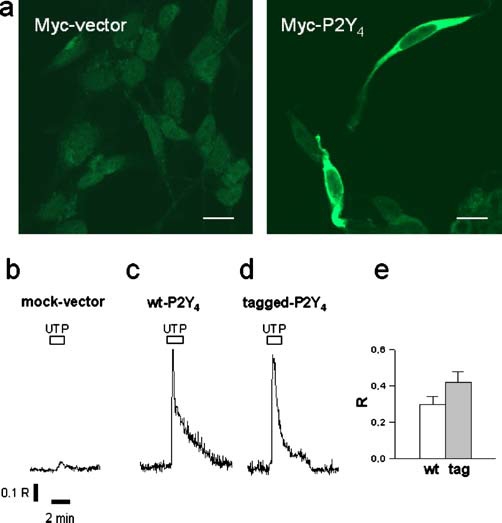
Expression and function of tagged P2Y4 receptors. SH-SY5Y cells transiently transfected with Myc-vector or Myc-P2Y4 were fixed, stained with anti-Myc (9E10) antibody and visualised by Cy2-conjugated anti-mouse antibody by means of a confocal laserscanning microscope. Scale bar, 10 µm (a). Cells transfected for 24 h with mock plasmid, wild-type P2Y4 or tagged P2Y4 in combination with green fluorescent protein (GFP)-expressing plasmid were loaded with Fura-2/AM and stimulated with 100 µM UTP for 2 min. Single traces show the time-course of ratio values (R, F340/F380) and are representative of intracellular Ca2+ mobilisation of each condition reported (b–d). The histogram (e) represents the mean amplitudes ± standard error of mean (SEM) of the peak of background-subtracted ratio values in respect to baseline of 18 GFP-positive wild-type transfected cells (wt) and 28 GFP-positive tag-transfected cells (tag)
Figure 3.
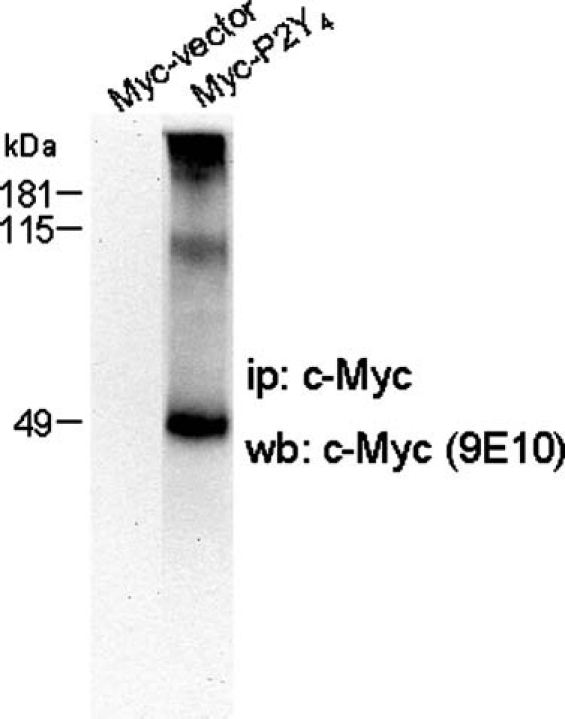
Western blotting pattern of Myc-P2Y4 receptor. SH-SY5Y cells transiently transfected with Myc-vector or Myc-P2Y4 were lysed and immunoprecipitated with anti-c-Myc-conjugated agarose. Immunoadsorbents were run on 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE), blotted and probed with anti-Myc (9E10) antibody. Protein bands were detected with secondary anti-mouse antiserum by electrogenerated chemiluminescence (ECL)
These findings show that the ability of endogenous P2Y4 to form high-order complexes is a property not only shown in different cell types in different species but also shared with recombinant Myc-P2Y4.
Reducing conditions alter the monomeric and multimeric ratio of P2Y4 receptor
We next analysed the effects of reducing agents on the multiple expression of P2Y4 protein (Figure 4). In the presence of 50 mM DTT and 5% β-mercaptoethanol (β-ME), 80% of the total P2Y4-positive signal is given in CGN by the 40 kDa monomeric band while the remaining 20% is constituted by the 80 kDa dimeric signal. This ratio is then inverted when protein separation is performed in the absence of reducing agents: the monomeric band is reduced to 25%, and the higher molecular mass band is increased to 70% of total protein (Figure 4a). A similar situation occurred for the transfected Myc-P2Y4 receptor. Under reducing conditions, the oligomer represented 35% of the total P2Y4 (instead of 55% under nonreducing conditions), the dimer was reduced to 5% (from 20%) and the monomer reached 65% (instead of 30%) (Figure 4b).
Figure 4.
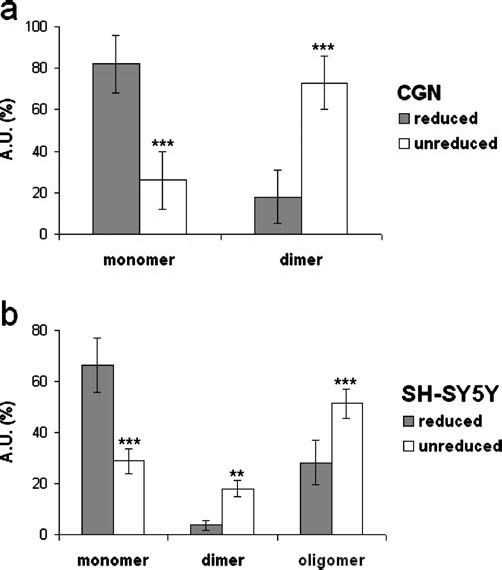
P2Y4 protein expression under reducing and nonreducing conditions. Total protein from cerebellar granule neurons (CGN) (a), and Myc-precipitated proteins recovered from Myc-P2Y4-transfected SH-SY5Y cells (b) were run on sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) in the absence or presence of 5% β-mercaptoethanol (β-ME) and 50 mM dithiothreitol (DTT). After immunoblotting with anti-P2Y4 or anti-c-Myc (9E10) antibodies, band intensity [expressed as arbitrary units (A.U.)] was analysed by Kodak 1.D Image Analysis software and reported as percent of total protein expression. Data represent means ± standard error of mean (S.E.M) from three (a) and five (b) independent experiments. Statistical analysis was performed using a two-way ANOVA followed by a post-hoc (Newman-Keuls) test for multiple comparisons (*p < 0.05, **p < 0.01, ***p < 0.001)
Myc-P2Y4 and FLAG-P2Y4 coexpresssed in SH-SY5Y cells associate to form homo-oligomeric complexes
To provide further direct evidence that P2Y4 subunits can form homo-oligomeric structures and to test their direct interaction and coassembly, both the Myc- and FLAG-tagged P2Y4 cDNAs were simultaneously cotransfected in SH-SY5Y cells. As shown in Figure 5, only under this condition, the immunoprecipitate of the FLAG-P2Y4 protein contained its cognate Myc-P2Y4 subunit, as detected by Western blot using the anti-Myc (9E10) antibody. The signal was absent after each single transfection, thus eliminating any possibility of cross-reaction of the antibodies. Moreover, the absence of signal after in vitro mixing of both the Myc- and FLAG-P2Y4 extracts, also ruled out the possibility that the interaction was caused by nonspecific receptor aggregation occurring in vitro during cell lysis and detergent protein extraction.
Figure 5.
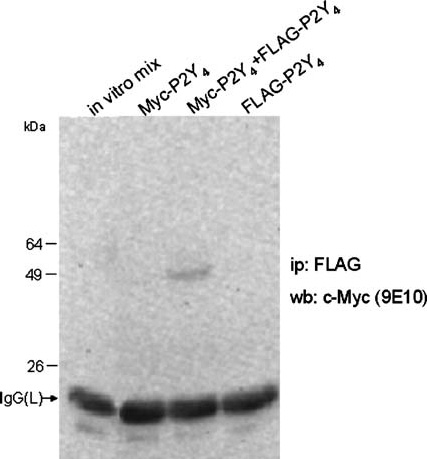
Homo-dimerisation of tagged-P2Y4 receptor. SH-SY5Y cell were transiently transfected with Myc-P2Y4, FLAG-P2Y4 or a combination of both tagged receptors. Solubilised proteins were immunoprecipitated with the anti-FLAG resin and detected by Western blot with the anti-Myc antibody. Protein bands were detected with secondary anti-mouse antiserum by electrogenerated chemiluminescence. The first lane (in vitro mix) derives from cells expressing individual tagged receptors that were mixed during lysis and then immunoprecipitated and detected at the above-mentioned conditions
Discussion
Extracellular nucleotides elicit very heterogeneous physiological effects, by activating not only ionotropic P2X receptors, but also G-protein-coupled P2Y metabotropic receptors, which are both widely distributed in the central nervous system (CNS) and peripheral nervous system (PNS), in neuronal and glial cells. P2Y receptors are generally present as 308–379 amino acid proteins, with a mass ranging from 41 to 53 kDa after glycosylation. The architecture of P2Y was studied by structural comparison based on sequence analysis, mutagenesis and homology modelling and established that they possess tertiary structures with seven transmembrane domains, extracellular N-terminus and intracellular C-terminus. Very little is known, of the possible quaternary structure of P2Y receptors. In this context, our work greatly improves the current knowledge about metabotropic receptors, demonstrating that at least the P2Y4 subunit can form higher-order complexes. These multimers appear stable, being to some extent resistant to denaturing and reducing conditions, thus indicating that they derive, at least in part, from covalent disulphide bonds occurring between the subunits. Moreover, both rat and human endogenous P2Y4 receptors, originating either from cell lines or primary neurons from both the PNS and CNS, appear as stable dimers, and this occurs also for the heterologous P2Y4 receptor transiently transfected in the neuroblastoma SH-SY5Y cell line. We would therefore consider this feature as an intrinsic property of the P2Y4 receptor, not requiring the transfection of auxiliary proteins in order to form oligomeric structures. Moreover, a particularity of the P2Y4 receptor is the absence of any potential N-glycosylation site. Since glycosylation is a post-translational modification common to many GPCRs involved in certain cases in the regulation of receptor surface expression and dimerisation [29], its potential participation in the control of P2Y4 oligomerisation can be likely ruled out.
Different authors often reported apparently conflicting molecular masses for the P2Y4 receptor [22, 30]. It was often postulated that the reason for this variability could rely on post-translational modifications, diverse among different tissues, such as glycosylation. With our work, we now demonstrate that different aggregation states could also account for the higher-order molecular masses reported for the P2Y4 receptor.
Totally new within the P2Y class, our results instead confirm what is already established for other GPCRs, which can directly associate as either homo- or hetero-oligomers, altering their respective pharmacology and sustaining, in this way, different functions from that evoked by the original receptors. For example, it is well documented that adenosine A1 and A2A receptors form homodimers [24, 31, 32], that these same subunits heterodimerise with dopamine D1 and D2 receptors [33–35] or with metabotropic glutamate receptors (mGluR) [36, 37]. Specifically within the purinergic class, co-localisation at the membrane level between the metabotropic P2Y4 and the ionotropic NMDAR1, and their functional interplay occurring during glucose starvation [17] was recently demonstrated in CGNs and human neuroblastoma cells. Moreover, the P2Y1 subunit is able to form heteromeric complexes with the adenosine A1 subtype [24–26], generating an adenosine receptor with P2Y-like agonistic pharmacology. As a consequence, adenosine 5′-O-(2-thiodiphosphate), a potent P2Y1 agonist, binds to the A1 pocket of the A1/P2Y1 complex, inhibiting adenylyl cyclase activity via Gi/o protein and, in this way, the hetero-oligomerisation between adenosine and P2Y receptors might be one of the mechanisms for the adenine nucleotide-mediated inhibition of neurotransmitter release [38]. Since these mechanisms further generate functional and structural heterogeneity among P2 receptors, they certainly contribute to the explanation of how different cells can evoke diverse responses in different tissues upon the same kind of agonist stimulation. For example, it appears from the literature that the ATP effect on the human P2Y4 receptor in transfected cells is variable. ATP has been actually described either as a partial agonist with antagonistic properties [39] or a full antagonist [40]. Whereas some of the discrepancies have been ascribed to the different receptor expression level in different transfected cell lines, in light of our results, we can now hypothesise that the heterogeneous behaviour exerted by ATP could also rely on the proportion of P2Y4 oligomerisation.
Under this perspective, it is therefore noteworthy to have established that the P2Y4 subtype can retain at least homo-oligomeric structures, both in native and transfected cell systems. The main physiological relevance of this investigation will come from studies regarding the exact sub-cellular localisation of the oligomeric forms of the receptor. This and analysis of the possibility that additional P2Y receptors might undergo homo- or hetero-multimeric assembly are therefore our next goals.
In conclusion, since the composition of the oligomers profoundly affects the biological response of P2 receptors and different subtype combinations thus yield different receptor characteristics, allowing diversity in agonists and antagonists selectivity, transmission signalling, channel and desensitisation properties, we do believe that the quaternary structure of the P2Y4, among other probable P2Y receptors, might in part contribute to explain how P2 metabotropic subtypes can trigger so many purinergic functions.
Acknowledgement
The research presented was supported by Progetto MIUR 2004 “Purinoceptors and Neuroprotection.” We thank Prof. T. K. Harden and Prof. M. P. Abbracchio for kindly providing the P2Y4 receptor cDNA.
References
- 1.Abbracchio MP, Boeynaems JM, Barnard EA, Boyer JL, Kennedy C, Miras-Portugal MT, King BF, Gachet C, Jacobson KA, Weisman GA, Burnstock G (2003) Characterization of the UDP-glucose receptor (re-named here the P2Y14 receptor) adds diversity to the P2Y receptor family. Trends Pharmacol Sci 24:52′5 [DOI] [PMC free article] [PubMed]
- 2.North RA, Barnard EA (1997) Nucleotide receptors. Curr Opin Neurobiol 7:346′57 [DOI] [PubMed]
- 3.King BF, Townsend-Nicholson A, Burnstock G (1998) Metabotropic receptors for ATP and UTP: exploring the correspondence between native and recombinant nucleotide receptors. Trends Pharmacol Sci 19:506′14 [DOI] [PubMed]
- 4.Communi D, Govaerts C, Parmentier M, Boeynaems JM (1997) Cloning of a human purinergic P2Y receptor coupled to phospholipase C and adenylyl cyclase. J Biol Chem 272:31969′1973 [DOI] [PubMed]
- 5.Communi D, Robaye B, Boeynaems JM (1999) Pharmacological characterization of the human P2Y11 receptor. Br J Pharmacol 128:1199′206 [DOI] [PMC free article] [PubMed]
- 6.Qi AD, Zambon AC, Insel PA, Nicholas RA (2001) An arginine/glutamine difference at the juxtaposition of transmembrane domain 6 and the third extracellular loop contributes to the markedly different nucleotide selectivities of human and canine P2Y11 receptors. Mol Pharmacol 60:1375′382 [DOI] [PubMed]
- 7.Communi D, Gonzalez NS, Detheux M, Brezillon S, Lannoy V, Parmentier M, Boeynaems JM (2001) Identification of a novel human ADP receptor coupled to G(i). J Biol Chem 276:41479′1485 [DOI] [PubMed]
- 8.Chambers JK, Macdonald LE, Sarau HM et al (2000) A G protein-coupled receptor for UDP-glucose. J Biol Chem 275:10767′0771 [DOI] [PubMed]
- 9.Lustig KD, Shiau AK, Brake AJ, Julius D (1993) Expression cloning of an ATP receptor from mouse neuroblastoma cells. Proc Natl Acad Sci USA 90:5113′117 [DOI] [PMC free article] [PubMed]
- 10.Nicholas RA, Watt WC, Lazarowski ER, Li Q, Harden K (1996) Uridine nucleotide selectivity of three phospholipase C-activating P2 receptors: identification of a UDP-selective, a UTP-selective, and an ATP- and UTP-specific receptor. Mol Pharmacol 50:224′29 [PubMed]
- 11.Herold CL, Qi AD, Harden TK, Nicholas RA (2004) Agonist versus antagonist action of ATP at the P2Y4 receptor is determined by the second extracellular loop. J Biol Chem 279:11456′1464 [DOI] [PMC free article] [PubMed]
- 12.Moore DJ, Chambers JK, Wahlin JP, Tan KB, Moore GB, Jenkins O, Emson PC, Murdock PR (2001) Expression pattern of human P2Y receptor subtypes: a quantitative reverse transcription-polymerase chain reaction study. Biochim Biophys Acta 1521:107′19 [DOI] [PubMed]
- 13.Ruan HZ, Burnstock G (2003) Localisation of P2Y1 and P2Y4 receptors in dorsal root, nodose and trigeminal ganglia of the rat. Histochem Cell Biol 120:415′26 [DOI] [PubMed]
- 14.Webb TE, Henderson DJ, Roberts JA, Barnard EA (1998) Molecular cloning and characterization of the rat P2Y4 receptor. J Neurochem 71:1348′357 [DOI] [PubMed]
- 15.Cheung KK, Ryten M, Burnstock G (2003) Abundant and dynamic expression of G protein-coupled P2Y receptors in mammalian development. Dev Dyn 228:254′66 [DOI] [PubMed]
- 16.Amadio S, D’Ambrosi N, Cavaliere F, Murra B, Sancesario G, Bernardi G, Burnstock G, Volonte C (2002) P2 receptor modulation and cytotoxic function in cultured CNS neurons. Neuropharmacology 42:489′01 [DOI] [PubMed]
- 17.Cavaliere F, Amadio S, Angelini DF, Sancesario G, Bernardi G, Volonte C (2004) Role of the metabotropic P2Y(4) receptor during hypoglycemia: cross talk with the ionotropic NMDAR1 receptor. Exp Cell Res 300:149′58 [DOI] [PubMed]
- 18.Rodrigues RJ, Almeida T, Richardson PJ, Oliveira CR, Cunha RA (2005) Dual presynaptic control by ATP of glutamate release via facilitatory P2X1, P2X2/3, and P2X3 and inhibitory P2Y1, P2Y2, and/or P2Y4 receptors in the rat hippocampus. J Neurosci 25:6286′295 [DOI] [PMC free article] [PubMed]
- 19.D’Ambrosi N, Murra B, Cavaliere F, Amadio S, Bernardi G, Burnstock G, Volonte C (2001) Interaction between ATP and nerve growth factor signalling in the survival and neuritic outgrowth from PC12 cells. Neuroscience 108:527′34 [DOI] [PubMed]
- 20.Agresti C, Meomartini ME, Amadio S, Ambrosini E, Serafini B, Franchini L, Volonte C, Aloisi F, Visentin S (2005) Metabotropic P2 receptor activation regulates oligodendrocyte progenitor migration and development. Glia 50:132′44 [DOI] [PubMed]
- 21.Fumagalli M, Brambilla R, D’Ambrosi N, Volonte C, Matteoli M, Verderio C, Abbracchio MP (2003) Nucleotide-mediated calcium signaling in rat cortical astrocytes: role of P2X and P2Y receptors. Glia 218:30 [DOI] [PubMed]
- 22.Sage CL, Marcus DC (2002) Immunolocalization of P2Y4 and P2Y2 purinergic receptors in strial marginal cells and vestibular dark cells. J Membr Biol 18:103′15 [DOI] [PubMed]
- 23.Bouvier M (2001) Oligomerization of G-protein-coupled transmitter receptors. Nat Rev Neurosci 2:274′86 [DOI] [PubMed]
- 24.Yoshioka K, Hosoda R, Kuroda Y, Nakata H (2002) Hetero-oligomerization of adenosine A1 receptors with P2Y1 receptors in rat brains. FEBS Lett 531:299′03 [DOI] [PubMed]
- 25.Yoshioka K, Saitoh O, Nakata H (2002) Agonist-promoted heteromeric oligomerization between adenosine A(1) and P2Y(1) receptors in living cells. FEBS Lett 523:147′51 [DOI] [PubMed]
- 26.Yoshioka K, Saitoh O, Nakata H (2001) Heteromeric association creates a P2Y-like adenosine receptor. Proc Natl Acad Sci USA 98:7617′622 [DOI] [PMC free article] [PubMed]
- 27.Levi G, Aloisi F, Ciotti MT, Thangnipon W, Kingsbury A, Balazs R (1989) Preparation of 98% pure cerebellar granule cell cultures. In: Shahar A, deVellis J, Vernadakis A, Haber B (eds) A dissection and tissue culture manual of the nervous system. Alan R. Liss, New York, pp 211′14
- 28.Bradford M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248′52 [DOI] [PubMed]
- 29.Xu J, He J, Castleberry AM, Balasubramanian S, Lau AG, Hall RA (2003) Heterodimerization of 2A- and 1-Adrenergic Receptors. J Biol Chem 278:10770′0777 [DOI] [PubMed]
- 30.Delbro DS, Nylund G, Nordgren S (2005) Demonstration of P 2 Y purinergic receptors in the HT human colon cancer cell line. Auton Autacoid Pharmacol 25:163′66 [DOI] [PubMed]
- 31.Ciruela F, Casado V, Mallol J, Canela EI, Lluis C, Franco R (1995) Immunological identification of A1 adenosine receptors in brain cortex. J Neurosci Res 42:818′28 [DOI] [PubMed]
- 32.Canals M, Burgueno J, Marcellino D, Cabello N, Canela EI, Mallol J, Agnati L, Ferre S, Bouvier M, Fuxe K, Ciruela F, Lluis C, Franco R (2004) Homodimerization of adenosine A2A receptors: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J Neurochem 88:726′34 [DOI] [PubMed]
- 33.Gines S, Hillion J, Torvinen M, Le Crom S, Casado V, Canela EI, Rondin S, Lew JY, Watson S, Zoli M, Agnati LF, Verniera P, Lluis C, Ferre S, Fuxe K, Franco R (2000) Dopamine D1 and adenosine A1 receptors form functionally interacting heteromeric complexes. Proc Natl Acad Sci USA 97:8606′611 [DOI] [PMC free article] [PubMed]
- 34.Canals M, Marcellino D, Fanelli F, Ciruela F, de Benedetti P, Goldberg SR, Neve K, Fuxe K, Agnati LF, Woods AS, Ferre S, Lluis C, Bouvier M, Franco R (2003) Adenosine A2A-dopamine D2 receptor-receptor heteromerization: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J Biol Chem 278:46741′6749 [DOI] [PubMed]
- 35.Hillion J, Canals M, Torvinen M, Casado V, Scott R, Terasmaa A, Hansson A, Watson S, Olah ME, Mallol J, Canela EI, Zoli M, Agnati LF, Ibanez CF, Lluis C, Franco R, Ferre S, Fuxe K (2002) Coaggregation, cointernalization, and codesensitization of adenosine A2A receptors and dopamine D2 receptors. J Biol Chem 277:18091′8097 [DOI] [PubMed]
- 36.Ciruela F, Escriche M, Burgueno J, Angulo E, Casado V, Soloviev MM, Canela EI, Mallol J, Chan WY, Lluis C, McIlhinney RA, Franco R (2001) Metabotropic glutamate 1alpha and adenosine A1 receptors assemble into functionally interacting complexes. J Biol Chem 276:18345′8351 [DOI] [PubMed]
- 37.Ferre S, Karcz-Kubicha M, Hope BT, Popoli P, Burgueno J, Gutierrez MA, Casado V, Fuxe K, Goldberg SR, Lluis C, Franco R, Ciruela F (2002) Synergistic interaction between adenosine A2A and glutamate mGlu5 receptors: implications for striatal neuronal function. Proc Natl Acad Sci USA 99:11940′1945 [DOI] [PMC free article] [PubMed]
- 38.Nakata H, Yoshioka K, Kamiya T, Tsuga H, Oyanagi K (2005) Functions of heteromeric association between adenosine and P2Y receptors. J Mol Neurosci 26:233′38 [DOI] [PubMed]
- 39.Communi D, Motte S, Boeynaems JM, Pirotton S (1996) Pharmacological characterization of the human P2Y4 receptor. Eur J Pharmacol 317:383′89 [DOI] [PubMed]
- 40.Kennedy C, Qi AD, Herold CL, Harden TK, Nicholas RA (2000) ATP, an agonist at the rat P2Y(4) receptor, is an antagonist at the human P2Y(4) receptor. Mol Pharmacol 57:926′31 [PubMed]