

Abstract
Recent evidence suggests a role for corticotropin-releasing factor (CRF) in the regulation of pair bonding in prairie voles. We have previously shown that monogamous and non-monogamous vole species have dramatically different distributions of CRF receptor type 1 (CRF1) and CRF receptor type 2 (CRF2) in the brain, and that CRF1 and CRF2 receptor densities in the nucleus accumbens (NAcc) are correlated with social organization. Monogamous prairie and pine voles have significantly lower levels of CRF receptor type 1 (CRF1), and significantly higher levels of type 2 (CRF2) binding, in NAcc than non-monogamous meadow and montane voles. Here, we report that microinjections of CRF directly into the NAcc accelerate partner preference formation in male prairie voles. Control injections of CSF into NAcc, and CRF into caudate-putamen, did not facilitate partner preference. Likewise, CRF injections into NAcc of non-monogamous meadow voles also did not facilitate partner preference. In prairie voles, this CRF-facilitation effect was blocked by co-injection of either CRF1 or CRF2 receptor antagonists into NAcc. Immunocytochemical staining for CRF and Urocortin-1 (Ucn-1), two endogenous ligands for CRF1 or CRF2 receptors in the brain, revealed that CRF, but not Ucn-1, immunoreactive fibers were present in NAcc. This supports the hypothesis that local CRF release into NAcc could activate CRF1 or CRF2 receptors in the region. Taken together, our results reveal a novel role for accumbal CRF systems in social behavior.
Keywords: nucleus accumbens, attachment, CRF1, CRF2, corticotropin-releasing factor, corticotrophin-releasing hormone, vole, neuropeptide receptors, pair bond, social behavior, monogamy, species differences
The corticotropin-releasing factor (CRF) system is involved in the neurobiology underlying stress and anxiety, but much less is known about its role in social behavior. Microtine rodents exhibit diverse social organizations and thus offer an excellent comparative approach in the study of the neurobiology of social behavior (Young and Wang, 2004). Prairie (Microtus ochrogaster) and pine voles (Microtus pinetorum) are monogamous; adult mates form long-lasting selective pair bonds in the field and the laboratory (Getz, Carter, and Gavish, 1981; Salo, Shapiro, and Dewsbury, 1993). In contrast, closely-related meadow (Microtus pennsylvanicus) and montane voles (Microtus montanus) are promiscuous and solitary (Gruder-Adams and Getz, 1985; Shapiro and Dewsbury, 1990). Past research has revealed that the brain distribution of neuropeptide receptors for oxytocin and vasopressin appears to be responsible for the species differences in social organization (Insel and Shapiro, 1992; Insel, Wang, and Ferris, 1994; Lim, Wang, Olazabal, Ren, Terwilliger, and Young, 2004b). However, more recent evidence suggests that another neuropeptide system, CRF, also appears to modulate pair bonding in prairie voles (DeVries, Guptaa, Cardillo, Cho, and Carter, 2002).
There are relatively few studies examining the role of stress hormones in social behavior. One study found that administration of exogenous corticosterone to male prairie voles facilitated pair bond formation with a novel female (DeVries, DeVries, Taymans, and Carter, 1996). A subsequent study found that CRF administered intracerebroventricularly (i.c.v.) facilitated partner preference in male prairie voles, even at extremely low doses that did not appear to affect locomotor activity or anxiety-like behavior (DeVries et al., 2002). Furthermore, partner preference was blocked by i.c.v. administration of alpha-helical CRF which non-selectively blocks CRF receptors in the brain (DeVries et al., 2002). These data suggest that CRF may play a role in pair bond formation via anxiety-independent mechanisms through the involvement of centrally acting brain receptors. However, i.c.v.-infused CRF could potentially be acting upon any number of brain regions to facilitate partner preference, and it is presently unknown which brain regions are specifically involved.
Since the CRF system has been implicated in the regulation of pair bond formation, we predicted that neural circuits for this system would differ between monogamous and non-monogamous species. We have previously shown that, although the distributions of CRF mRNA and peptide appear highly conserved among vole species, the distributions of CRF receptor types 1 and 2 (CRF1 and CRF2) dramatically differ throughout the brain in four vole species exhibiting varying social organizations (Lim, Nair, and Young, 2005; Lim, Tsivkovskaia, Bai, Young, and Ryabinin, 2006). Receptor binding appeared to correlate with monogamous social structure in several brain regions; however, only the nucleus accumbens (NAcc) consistently segregated with both monogamous vole species and both non-monogamous vole species. Monogamous prairie and pine voles have significantly lower levels of CRF receptor type 1 (CRF 1), and significantly higher levels of type 2 (CRF2) binding, in NAcc than non-monogamous meadow and montane voles (Lim et al., 2005).
Based upon our neuroanatomical studies demonstrating species differences in CRF1 and CRF2 densities in NAcc, we hypothesized that CRF action within NAcc, in particular, was critical for monogamous social behavior in prairie voles. First, we determined whether CRF injected directly into NAcc can facilitate partner preference formation after an abbreviated cohabitation time with the partner. Next, we performed the identical experiment in non-monogamous meadow voles. Then, we manipulated CRF1 and CRF2 in NAcc using pharmacological antagonists to determine their relative contributions to CRF-facilitated partner preference formation. Lastly, we show evidence for immunoreactive staining of two endogenous ligands for CRF1 and CRF2 receptors in the brain, CRF- and Urocortin-1 (Ucn-1), in NAcc in prairie voles. The results from these studies demonstrate for the first time that CRF, acting in the NAcc can promote social attachment, and furthermore, that both CRF1 and CRF2 receptors are involved in this process.
METHODS
Subjects
Animals were adult, sexually-naïve, male and female prairie (70–100 days of age) from a laboratory breeding colony at Florida State University that were originally derived from field-captured voles in Illinois, USA. Adult sexually-naïve meadow voles were from the laboratory breeding colony at Emory University. After weaning at 21 days of age, subjects were housed in same-sex sibling pairs or trios and water and Purina rabbit chow provided ad libitum. All cages were maintained on a 14:10 light:dark cycle with the temperature at 20° C. Data from 87 male prairie voles were included in the CRF pharmacology experiments, along with equal numbers of stimulus female prairie voles for the pair bonding assay. Data from 10 male meadow voles were used as well, along with equal numbers of stimulus female meadow voles. Eight prairie voles were used in the CRF immunocytochemistry studies (n = 4 for each sex).
CRF facilitation of partner preference
Adult male prairie voles (n = 31) were bilaterally cannulated into NAcc using stereotaxic methods as previously described (Aragona, Liu, Curtis, Stephan, and Wang, 2003a; Liu and Wang, 2003). Subjects were anesthetized with sodium pentobarbital (2.5 mg per 40 gm body weight), and 26 gauge bilateral guide cannulas (Plastics One, Roanoke, VA) aimed at the NAcc were implanted stereotaxically (Anterior 1.7 mm, Bilateral ±1 mm, Ventral −4.0 mm to bregma). Control injections (n = 6) were aimed at the caudate-putamen (Anterior 1.7 mm, Bilateral ±1 mm, Ventral −2.5 mm to bregma). After 3–5 days recovery, subjects received microinjections (200 nl per side) of either artificial CSF or drug dissolved in CSF. Injections were made with a 33 gauge needle that extended 1 mm below the guide cannula into the target area. The needle was connected to a Hamilton syringe (Hamilton, Reno, NV) via polyethylene-20 tubing, through which the solution was infused slowly by a pump (MasterFlex L/S standard drive, Model 7016-21) at a speed of 200 nl/min, per side. Human/rat CRF was obtained from Sigma (St. Louis, MO).
Animals were divided into one of four groups: CSF control (n = 7), 0.01 pg CRF into NAcc (n = 9), 0.1 pg CRF into NAcc (n = 15), and 0.1 pg CRF into caudate-putamen (n = 6). The caudate-putamen (CP) is a brain region just dorsal to the NAcc and also contains CRF2 receptors, thus serving as an anatomical control region for CRF effects. Each animal received bilateral injections of 200 nl volume prior to an abbreviated 6 hour cohabitation with a novel female. The concentration of 0.01 pg CRF in 200 nL is 10 nM, while the concentration of 0.1 pg CRF in 200 nL is 100 nM. The calculated Ki for CRF1 is 11 nM, while the calculated Ki for CRF2 is 25 nM relative to 125I-sauvagine (Primus, Yevich, Baltazar, and Gallager, 1997). Cohabitation for 6 hours without mating consistently does not induce partner preference, as demonstrated in previous studies (Aragona et al., 2003a; Aragona, Liu, Yu, Curtis, Detwiler, Insel, and Wang, 2006). Immediately following cohabitation, subjects were tested for partner preference.
Partner preference testing consisted of placing the male into a 3-chambered apparatus in which the female partner was tethered in one cage, and a novel female (“stranger”) of the same age and sociosexual experience was tethered in a second cage, as previously described (Carter, DeVries, and Getz, 1995). Each stimulus female was used in two separate partner preference tests, once as the partner and again as another subject’s stranger, and therefore each female had equivalent social and sexual exposure during the 20 hour cohabitation. Subjects were allowed to freely roam throughout the apparatus, and time spent in contact with the partner and stranger was quantified over the course of the 3 hour test.
Locomotor activity was measured during the partner preference test to determine whether chosen doses of CRF treatment affected general locomotor activity or anxiety-like behavior, as previously described (Hotta, Shibasaki, Arai, and Demura, 1999). The number of cage crossings through both tunnels of the partner preference apparatus was assessed by using infrared detectors. There are four infrared beams across the three cages of the partner preference apparatus, with two beams flanking each tunnel connecting two cages. The total number of photo beam breaks was totaled for each animal during the three-hour period. After behavioral testing, subjects were sacrificed and injection sites were verified histologically.
Adult male meadow voles (n=10) were also tested for CRF-facilitation of partner preference. Animals were cannulated bilaterally into NAcc as described above and randomly assigned to one of two groups: CSF control (n=4), or CRF 0.1 pg (n=6). Cohabitation and partner preference tests were performed exactly as described above for prairie voles. Adult male meadow voles from our colony typically do not form partner preferences when cohabitated with a female (Lim et al., 2004b).
CRF1- and CRF2-selective pharmacology and partner preference
Adult prairie voles were cannulated bilaterally into NAcc as described above and divided into one of three groups: 0.1 pg CRF (n = 10), 0.1 pg CRF plus 10 pg CRF1 antagonist (CP-154,526) (n = 25), and 0.1 pg CRF plus 10 pg CRF2 antagonist (anti-sauvagine-30) (n = 15). Anti-sauvagine-30 was obtained from Sigma (St. Louis, MO), and the CP-154,526 from Michael Owens, Ph.D. (Emory University, Atlanta, GA). The concentration of 10 pg CRF1 antagonist (CP-154,526) in 200 nL solution is 100 nM, while the concentration of 10 pg CRF2 antagonist (anti-sauvagine-30) in 200 nL solution is 10 nM. Each animal received bilateral injections of 200 nl volume directly into NAcc prior to an abbreviated 6 hour cohabitation with a novel female. Immediately following cohabitation, subjects were tested for partner preference as described above. After behavioral testing, subjects were sacrificed and injection sites were verified histologically. Animals whose cannulation sites were placed outside the NAcc were excluded from data analysis and not reflected in the total number of animals used.
Data Analysis
Data from the partner preference test for each experiment were analyzed using a 2 way ANOVA in which stimulus (partner or stranger) and treatment were factors. In addition, Student’s t-tests were used to compare time in contact with the partner and stranger within each treatment group. Bonferroni corrections for the level of significance were made for each experiment in order to minimize risk of type-I error risk due to multiple comparisons. Males were categorized as having developed a partner preference if they spent more than twice as much time in contact with the partner than the stranger.
Data collected for locomotor activity was totaled as the number of infrared beam breaks for each animal, and averaged within each treatment group. Results were analyzed using a one-way ANOVA with treatment as the independent factor.
CRF and Urocortin-1 Immunohistochemistry
Adult prairie voles were deeply anesthetized between the hours of 10:00 and 14:00 with isoflurane and perfused with saline followed by 2% paraformaldehyde in 10 mM (pH 7.4) phosphate-buffered saline (PBS). Dissected brains were postfixed overnight in the 2% paraformaldehyde/PBS solution and cryoprotected in 30% sucrose/PBS. Thirty μm thick free-floating coronal sections were sliced on a cryostat and processed for immunohistochemistry according standard protocols (Ryabinin, Criado, Henriksen, Bloom, and Wilson, 1997; Weitemier, Tsivkovskaia, and Ryabinin, 2005) with adjustments for vole tissue made in earlier experiments (Lim et al., 2006). Briefly, endogenous peroxidase activity was quenched by 15-minute incubation with 0.3% hydrogen peroxide. For the Urocortin-1-specific antibody, blocking was performed by a five hour incubation with 2% Bovine Serum Albumin, 0.1% heparin, 0.01% Triton X-100 in PBS. For the CRF-specific antibody, blocking was performed by a five hour incubation with 4.5% goat serum, 0.3% Triton X-100 in PBS. The primary rabbit antibodies recognizing Urocortin-1 (Sigma-Aldrich, St. Louis, MO) were used in dilution 1:5,000. The primary antibodies recognizing CRF (Peninsula Laboratories, San Carlos, CA) were used in dilution 1:15,000. Biotynylated anti-rabbit secondary antibodies (Vector Laboratory Inc., Burlingame, CA) were used to detect the primary antibodies. Detection of the secondary antibodies was made using Vectastain ABC kit (Vector), and enzymatic development was done with the Metal Enhanced DAB kit (Pierce, Rockford, IL, USA). The specificity of immunostaining was assessed by a complete lack of immunoreactivity in areas not known to express CRF or Urocortin-1. Futhermore, control preabsorption experiments for these antibodies were previously performed (Bachtell, Weitemier, Galvan-Rosas, Tsivkovskaia, Risinger, Phillips, Grahame, and Ryabinin, 2003).
Qualititative image analysis was performed using a system consisting of an Olympus BX40 microscope and a high-resolution digital videocamera (Olympus Qcolor3) interfaced to a Macintosh personal computer running OS-X. Images from a single section best matched across animals for each brain region were digitally collected at the same lighting intensity. Since no immunopositive staining was observed in cell bodies in NAcc, the number of immunoreactive neurons was not quantitatively counted.
RESULTS
Pharmacological manipulation of CRF receptors in NAcc
It had been previously demonstrated that infusion of CRF i.c.v. facilitates partner preference in prairie voles (DeVries et al., 2002). Based on our neuroanatomical data demonstrating species differences in CRF1 and CRF2 in NAcc, we hypothesized that the NAcc was the site of action for the CRF-facilitation of partner preference. The earlier study showed that dose-dependent i.c.v. administration of CRF could facilitate partner preferences in male prairie voles after an abbreviated cohabitation with a female (DeVries et al., 2002). Based on this study, we designed CRF doses for site-specific injections into NAcc. The doses that we used, 0.1 pg and 0.01 pg CRF dissolved in 200 nL isotonic solution (or 100 nM and 10 nM, respectively), were well below the minimum effective i.c.v doses of 0.1 ng and 1 ng CRF dissolved in 1 μL (or 210 nM and 2.1 μM, respectively) (DeVries et al., 2002). Although CRF has been shown to bind preferentially to CRF1, it also binds to CRF2 with substantial affinity (Ki equal to 11 and 25, respectively) (Primus et al., 1997).
Analysis of the overall data set using a 2-way ANOVA revealed a significant main effect of stimulus animal (F(1,66) = 6.77, p<0.05), but no other main effects or interactions were detected. To determine which groups preferentially spent time in contact with the partners over the strangers, Student’s t-tests were performed with Bonferroni corrections of the p-value. Control prairie voles with bilateral injections of artificial CSF into the NAcc septal pole, or 0.1 pg CRF into the caudate-putamen, did not spend significantly more time in contact with the partner than the stranger stimulus animal (p>0.3, Student’s t-test, Bonferroni level set at p<0.01) (Figure 1A). Prairie voles injected with the lower CRF dose, 0.01 pg, trended towards spending more time in contact with the partner than the stranger (p>0.08, Student’s t-test, Bonferroni level set at p<0.01). In contrast, prairie voles receiving bilateral injections of the 10-fold higher CRF dose, 0.1 pg into the NAcc septal pole, spent significantly more time in contact with the partner than the stranger (p<0.01, Student’s t-test, Bonferroni level set at p<0.01) (Figure 1A). Furthermore, while only 6 of the 13 control animals displayed a partner preference, defined as spending twice as much time in contact with the partner compared to the stranger, 12 of the 15 animals receiving 0.1 pg of CRF displayed a partner preference (Figure 1B). The 3 animals that did not display a partner preference had strong stranger preferences, which likely contributed to the lack of a main effect of treatment or an interaction effect in the 2-way ANOVA. Thus, while CRF infusions into the NAcc significantly increased the time in contact with the partner relative to the stranger, it did not result in an overall increase in contact time with the partner.
Figure 1.

Bilateral micro-injections of CRF into NAcc facilitate partner preference in male prairie voles. (A) Control animals that received artificial CSF into the NAcc, or 0.1 pg CRF into the caudate-putamen (CP), did not significantly prefer the partner over the stranger after 6 hours of cohabitation with the partner (p>0.3, Students t-test). Animals that received 0.01 pg CRF into NAcc (or 10 nM) trended towards significant partner preferences (p>0.08, Students t-test). Animals receiving a 10-fold higher dose, 0.1 pg CRF (or 100 nM), into the NAcc spent significantly more time in contact with the partner than the stranger (p<0.01, Student’s t-test, Bonferroni level set at p<0.01). (B) Less than half of the control animals displayed a partner preference, while 80% of the animals receiving 0.1 pg of CRF into the NAcc developed a partner preference. Partner preference is defined as spending twice as much time in contact with the partner than with the stranger.
Because of dramatic species differences in CRF1 and CRF2 densities in NAcc, we hypothesized that CRF action within NAcc would only facilitate partner preference formation in prairie voles, and not in non-monogamous meadow voles. Indeed, meadow voles injected with CSF, or the high dose 0.1 pg CRF, did not spend significantly more time in contact with the partner than the stranger (p>0.5, Student’s t-test) (Figure 2).
Figure 2.
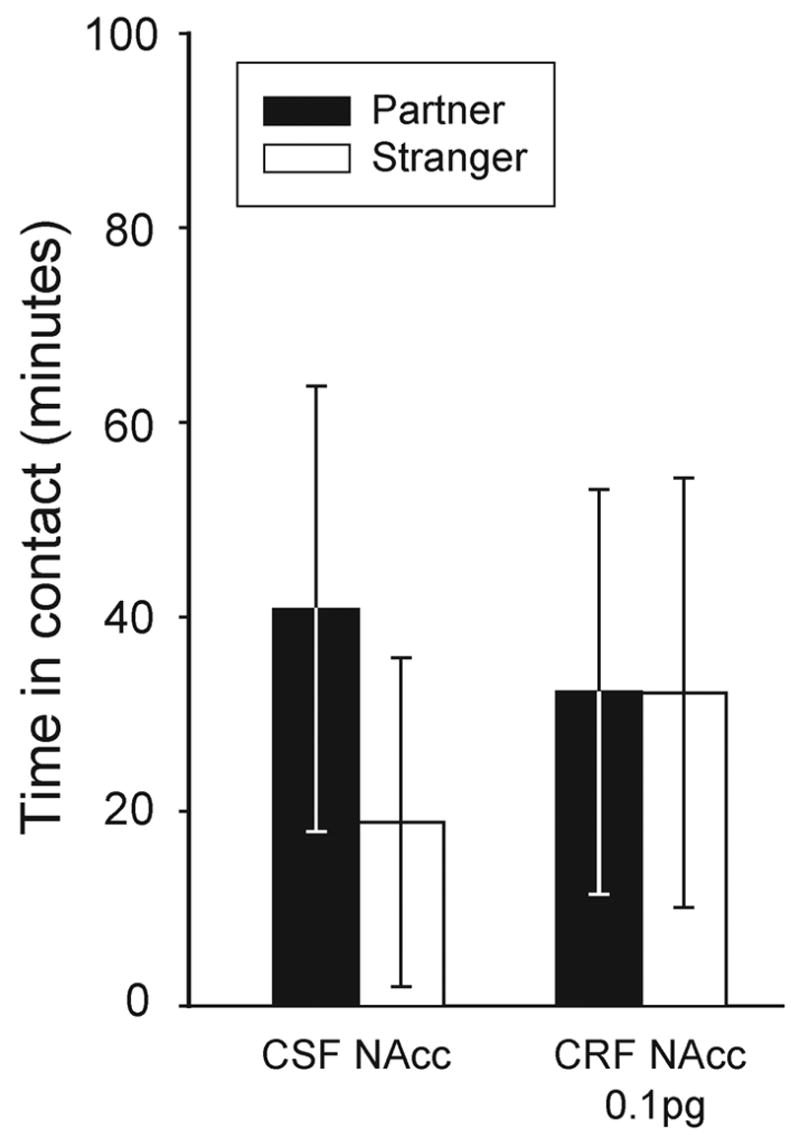
Bilateral micro-injections of CRF into NAcc fail to facilitate partner preference in non-monogamous male meadow voles. Adult meadow voles injected with artificial CSF, or 0.1 pg of CRF into NAcc, did not spend more time in contact with the partner than with the stranger (p>0.5, Student’s t-test).
Based on the species differences in CRF1 and CRF2 distribution in NAcc, we hypothesized that both CRF1 and CRF2 would modulate partner preference behavior, perhaps in opposite directions. Analysis of the overall data set using a 2-way ANOVA revealed a significant main effect of stimulus animal (F(1,94) = 7.52, p<0.05), but no other main effects or interactions were detected. Prairie voles injected with a cocktail of 0.1 pg of CRF, plus 10 pg selective CRF2 antagonist anti-sauvagine-30, did not spend significantly more time with the partner or the stranger (p>0.3, Student’s t-test, Bonferroni level set at p<0.016) (Figure 3A). Interestingly, prairie voles injected with a cocktail of 0.1 pg of CRF, plus 10 pg selective CRF1 antagonist CP-154,526-1, also showed a blockade of partner preference (p>0.5, Student’s t-test, Bonferroni level set at p<0.016) (Figure 3A). Control prairie voles injected with 0.1 pg CRF into the NAcc were tested simultaneously, and were found to replicate the original results of facilitation of partner preference (p<0.01, Student’s t-test, Bonferroni level set at p<0.016) (Figure 3A). In addition, while 8 out of the 10 CRF-treated prairie voles displayed a partner preference, only 11 of the 25 animals receiving CRF1 antagonist, and 6 out of 15 animals receiving CRF2 antagonist, displayed a partner preference (Figure 3B). Our results suggest that the activation of both CRF1 and CRF2 receptors in NAcc are necessary for CRF-induced facilitation of partner preference in prairie voles.
Figure 3.

Both CRF1 and CRF2 receptors in the NAcc are necessary for CRF-facilitated partner preference formation in prairie voles. (A) Animals injected with 0.1 pg CRF into NAcc spent more time in contact with the partner than the stranger (p<0.01, Student’s t-test, Bonferroni level set at p<0.016). This facilitation effect was blocked by the co-infusion of a selective CRF1 antagonist (CP-154,526) and a selective CRF2 antagonist (anti-sauvagine-30) (p>0.3, Student’s t-test). (B) 80% of the CRF-treated animals developed a partner preference in the absence of antagonist, while less than half of those received CRF1 or CRF2 antagonist developed a partner preference.
Locomotor activity did not significantly differ between treatment groups (F(1,80) = 1.37, p>0.05, one-way ANOVA), although locomotor activity tended to be lower in the animals receiving CRF into the NAcc. Results are shown in Table 1. A representative histology section showing the cannulation site for the NAcc is shown in Figure 4.
Table 1.
Locomotor activity represented by the total number of infrared beam breaks or cage crossings, averaged within each treatment group. There are no significant differences between treatment groups (F = 1.37, p>0.05, one-way ANOVA).
| Treatment | Beam breaks | Std Err |
|---|---|---|
| CSF in Nacc | 379 | ± 87 |
| 0.1 pg CRF in CP | 446 | ± 95 |
| 0.01 pg CRF in Nacc | 292 | ± 62 |
| 0.1 pg CRF in Nacc | 290 | ± 27 |
| CRF+R1 antag in Nacc | 469 | ± 73 |
| CRF+R2 antag in Nacc | 402 | ± 77 |
Figure 4.
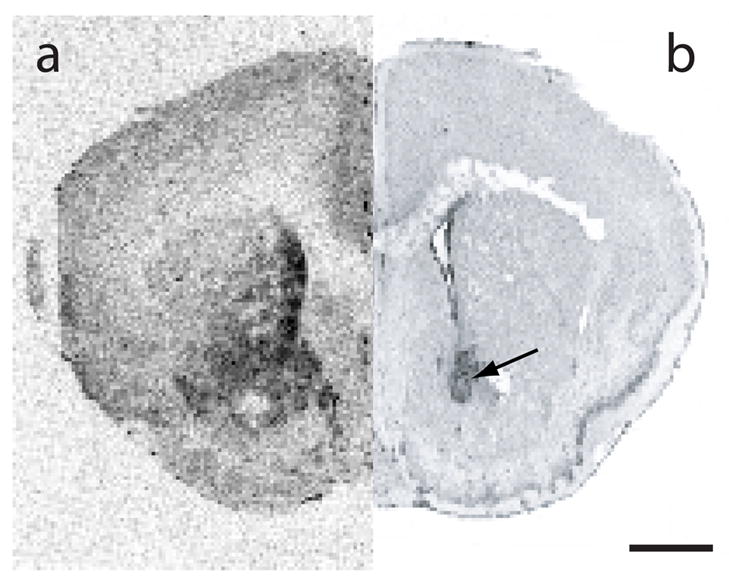
Histological verification of cannula placement. (a) Receptor autoradiogram on the left half depicts the location of CRF2 in prairie vole NAcc. (b) Representative photomicrograph of Nissl-stained brain section depicting cannula placement terminating within NAcc (see arrow). Scale bar = 1 mm.
CRF- and Ucn-1 immunoreactivity in NAcc
To show photomicrographs of which endogenous CRF receptor ligands are present in NAcc, we performed CRF- and Urocortin-1 (Ucn-1) immunoreactivity in adult prairie voles. Representative brain sections processed for CRF immunocytochemistry are shown in Figure 5. CRF-immunoreactive fibers were seen in NAcc in both sexes with no obvious differences in distribution or density of fibers (Figure 5b). Ucn-1 fibers were not detected in NAcc in either male or female prairie voles (Figure 5c). Thus, it is possible that CRF is one of the endogenous ligands which could physiologically bind CRF1 and CRF2 receptors in the vole NAcc to facilitate partner preference. Of note, CRF has been shown to bind to both CRF1 and CRF2, with approximately two- to ten-fold affinity higher affinity for CRF1 (Primus et al., 1997). However, we cannot rule out the possibility that other endogenous ligands such as Urocortin-2 or Urocortin-3 may also contribute in the neural control of pair bond formation.
Figure 5.

CRF and Ucn-1 immunoreactivity in prairie voles. (a) Rat atlas schematic showing the area of 10x magnification of NAcc (see rectangle) (Paxinos and Watson, 1998). (b) Prairie vole section showing CRF-immunoreactive fibers in NAcc (see arrows). (c) Prairie vole section stained for Ucn-1 immunnocytochemistry. Note the lack of Ucn-1 immunoreactive fibers in NAcc. ac = anterior commissure. Scale bar = 100 μm.
DISCUSSION
In the previous studies, we identified species differences in accumbal CRF1- and CRF2 expression that correlated with social organization across four vole species. Monogamous prairie and pine voles had higher levels of CRF2 in NAcc, and lower levels of CRF1 in NAcc, compared to non-monogamous meadow and montane vole species (Lim et al., 2005). Based on this data, we hypothesized that CRF action within NAcc was critical for monogamous social behavior in prairie voles. In the present study, we show for the first time that micro-injections of CRF directly into NAcc do in fact facilitate partner preference in male prairie voles. Two-way ANOVA analysis detected a main effect of stimulus animal, i.e. overall more time was spent in contact with the partner than the stranger, but no main effect of treatment or interaction was detected. Strong stranger preferences in 3 animals of the 0.1 pg CRF NAcc group inflated the variance, preventing detection of an interaction effect. However, separate comparisons of time spent with partner verses stranger revealed significant preferences for the partner in the 0.1 pg CRF NAcc group. This effect was replicated in the antagonist study, attesting to the robustness of the effect. This shift in partner preferences was not associated with a statistically significant increase in time spent with the partner or a decrease in time spent with stranger, but rather was the result of an overall increased preference for the partner relative to the stranger. In contrast, CRF has no effect on partner preference in non-monogamous meadow voles. Additionally, we show that this facilitation effect is modulated by action of CRF at both CRF1 and CRF2 receptors. Lastly, we show photomicrograph evidence that CRF-immunoreactive fibers are present within the prairie vole NAcc, indicating that CRF could be one of the endogenous ligands acting upon CRF1 and CRF2 receptors in NAcc during partner preference formation. Taken together, these data demonstrate a novel role for CRF systems acting within NAcc in social behavior.
Our data showing that CRF in NAcc facilitates partner preference supports our initial hypothesis that accumbal CRF systems are involved in pair bond formation in prairie voles. We had further hypothesized that CRF2 receptors, in particular, were critical given the abundance of CRF2 receptors in the two monogamous vole species but not the two non-monogamous vole species (Lim et al., 2005). The results from the meadow vole experiment support this hypothesis, because CRF infusion has no effect on partner preference in a species that effectively lacks CRF2 receptors in NAcc. In addition, we found that co-administration of a CRF2-selective antagonist blocks partner preference in prairie voles. These data reveal a potentially critical role for CRF2 receptors in pair bond formation.
However, we also found that co-administration of a CRF1-selective antagonist blocked partner preference in prairie voles. This result was more surprising, given that CRF1 receptors are expressed in NAcc in both non-monogamous and monogamous species. Taken together, these data underscore the importance of both receptor subtypes for the expression of partner preference, and point to the possibility that receptor specificity is a complex issue which may benefit from further exploration. It is possible that there may be dynamic interplay between the two receptor subtypes in NAcc during pair bonding behavior, and it might be interesting to further explore the cellular phenotypes of CRF1- and CRF2-expressing neurons, or to see if CRF1 and CRF2 receptors might even colocalize to the same neurons. It is also possible that other agents such as the CRF binding protein, which may act as a reservoir for endogenous CRF, may be involved (Jahn, Eckart, Brauns, Tezval, and Spiess, 2002).
The effective CRF doses for site-specific injections into NAcc that facilitated partner preference resulted in no significant effect of drug treatment on locomotor activity, which is commonly interpreted as anxiety-like behavior. DeVries and Carter (2002) found effective CRF i.c.v. doses for partner preference at 0.1 and 1.0 ng (210 nM and 2.1 μM, respectively ), and did not detect differences in locomotor activity between treatment groups (DeVries et al., 2002). Their doses were 1000 to 10,000 fold greater and at least twice to twenty-fold higher in concentration than the doses that we used site-specifically into NAcc (10 nM and 100 nM). In our study, though we did not detect significant differences in locomotor activity across treatment groups, there was, however, a slight trend toward fewer cage crossings in the animals receiving CRF alone into NAcc. While it is possible that the CRF may exert subtle effects on anxiety-like behavior and therefore locomotion, which could in turn affect partner preference formation, we believe the more plausible explanation is that decreased cage crossings in the CRF alone group is a byproduct of increased partner preference, i.e., time spent within the partner’s cage only. This supports the hypothesis that CRF may have a novel, separate role in regulating social behavior, possibly independent from HPA axis effects on anxiety.
We also show photomicrograph evidence of CRF-immunoreactivity in NAcc in the same region as CRF2 receptors in monogamous prairie voles. This suggests that CRF could be one of the endogenous ligands which acts upon CRF1 and CRF2 receptors in NAcc. Although CRF has been shown to bind preferentially to CRF1, it also binds to CRF2 with substantial affinity (Primus et al., 1997). Ucn-1-immunoreactive fibers were not seen in NAcc, but were observed in other regions of the brain such as the Edinger-Westphal nucleus (Lim et al., 2006). We were unable to map Urocortin-2 or Urocortin-3 fibers in the vole brain due to lack of specific immunostaining; however, it would be interesting to determine if these potential ligands, which also bind CRF2 receptors with high affinity, are also present in NAcc along with CRF2 receptors.
The ventral forebrain, and particularly the NAcc, has been repeatedly identified as the critical brain region for pair bond formation in prairie voles. Given the role of NAcc in the mesolimbic dopamine reward pathway, it has been hypothesized that natural reward and reinforcement mechanisms underlie pair bond formation, such that the partner is selectively associated with reward (Aragona et al., 2003a; Lim, Murphy, and Young, 2004a). We have previously shown through pharmacological and genetic manipulations that ventral forebrain vasopressin V1a receptors are necessary for male pair bond formation, even when over-expressed in non-monogamous vole species (Lim et al., 2004b; Lim and Young, 2004). Oxytocin receptors in the NAcc are necessary for partner preference in female prairie voles (Young, Lim, Gingrich, and Insel, 2001). Accumbal dopamine D1 and D2 receptors have also been shown to modulate partner preference formation and maintenance in both males and females, and in fact dopamine interacts with oxytocin during this behavioral process (Aragona et al., 2003a; Aragona et al., 2006; Aragona, Liu, Yu, Damron, Perlman, and Wang, 2003b; Liu and Wang, 2003). Thus, CRF receptor activation likely contributes to a larger circuit which converges in the NAcc to produce this complex social behavior. Consistent with this hypothesis, there is evidence that CRF receptors in the NAcc can modulate dopamine release into the striatum (Lu, Liu, Huang, and Zhang, 2003), and recent preliminary evidence suggesting that NAcc CRF receptor activation can stimulate bar pressing for natural reinforcement (Berridge, Pecine, and Schulkin, 2004). Another study has shown that CRF2 receptors in the ventral tegmental area, which sends dopaminergic projections to NAcc, can induce long-term potentiation, a physiologic correlate of behavioral learning and reward association (Ungless, Singh, Crowder, Yaka, Ron, and Bonci, 2003). Because partner preference is postulated to be a form of natural reward learning, CRF receptors in NAcc may play a similar role in underlying synaptic potentiation during pair bond formation in prairie voles.
Pair bond formation in nature is a complex cognitive process that requires the integration of many external stimuli and internal states. Pair bond formation results from the synthesis of several behavioral processes including social recognition, approach and social motivation, and involves learning and memory. Oxytocin and vasopressin are integrally involved in the neural processing of social stimuli and the formation of social memories (Bielsky, Hu, Szegda, Westphal, and Young, 2004; Ferguson, Young, Hearn, Matzuk, Insel, and Winslow, 2000). Dopamine may be involved in the heightened motivational state driving social interaction with one’s partner, and the reinforcement needed to establish a partner preference. CRF may provide a mechanism by which the internal stress state modulates partner preference. CRF signaling could also enable the long-term changes in neural plasticity during pair bond formation. Each neurotransmitter system plays a different but crucial role in the complex behavior of pair bonding, and blockade of any one system would disrupt the formation of a pair bond.
The role of CRF in the regulation of social behavior has been minimally studied, despite the abundance of studies relating CRF to stress and anxiety behaviors. There is strong evidence that CRF2 receptor activation functions to decrease anxiety and depression-like behaviors in mice (Bale, Contarino, Smith, Chan, Gold, Sawchenko, Koob, Vale, and Lee, 2000; Bale and Vale, 2003). Social behavior, stress, and anxiety are heavily interrelated, especially in behaviors involving social support or coping with social isolation. Prairie voles that have formed pair bonds show elevated plasma corticosterone levels during social separation from the partner, and reunion with the partner returns these levels back to baseline (Carter, DeVries, Taymans, Roberts, Williams, and Getz, 1997). Male prairie voles undergoing forced swimming, a psychological stressor, show a facilitation of pair bond formation after an abbreviated cohabitation with the partner (DeVries et al., 1996). Finally, pair-bonded males that are separated from their partners show more passive coping strategies in the forced swim test than their sibling-separated counterparts, and such behavioral changes are accompanied by an increase in CRF mRNA in the NAcc (Bosch, Nair, Neumann, and Young, 2005).
These data suggest that social and stress behaviors have a reciprocal relationship, and furthermore, the same molecules implicated in stress and anxiety also play an important role in social behavior. In fact, there is evidence that the “social” neuropeptides vasopressin and oxytocin may modulate stress and anxiety behavior (Bielsky et al., 2004; Landgraf, Gerstberger, Montkowski, Probst, Wotjak, Holsboer, and Engelmann, 1995; Liebsch, Wotjak, Landgraf, and Engelmann, 1996; Mantella, Vollmer, Li, and Amico, 2003; Ring, Malberg, Potestio, Ping, Boikess, Luo, Schechter, Rizzo, Rahman, and Rosenzweig-Lipson, 2006; Windle, Shanks, Lightman, and Ingram, 1997). Thus, the same molecules that modulate the internal stress state may contribute to the regulation of social behaviors such as pair bond formation, and that the molecules and circuits evolved for the purpose of one behavior may in fact dynamically control the other.
Acknowledgments
At Emory University, we would like to thank Dr. Michael J. Owens for graciously providing us with the CP-154,526 compound. We would also like to thank Lorra Miller and Meera Modi for their assistance with the meadow vole experiment. Lastly, we would like to thank Drs. A. Courtney DeVries at Ohio State University and C. Sue Carter at the University of Illinois, Chicago for their pioneering work on CRF in prairie voles and Dr. DeVries’ correspondence on the pharmacological experiments.
Grant support: This research was supported by NIH grants MH65050 to MML, AA13738 to AER, MH58616 to ZXW, MH64692 to LJY, and NSF STC IBN-9876754 and the Yerkes Center Grant RR00165.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aragona BJ, Liu Y, Curtis JT, Stephan FK, Wang Z. A critical role for nucleus accumbens dopamine in partner-preference formation in male prairie voles. J Neurosci. 2003a;23(8):3483–90. doi: 10.1523/JNEUROSCI.23-08-03483.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aragona BJ, Liu Y, Yu YJ, Curtis JT, Detwiler JM, Insel TR, Wang Z. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nat Neurosci. 2006;9(1):133–9. doi: 10.1038/nn1613. [DOI] [PubMed] [Google Scholar]
- Aragona BJ, Liu Y, Yu YJ, Damron A, Perlman G, Wang ZX. Opposite modulation of social attachment by D1- and D2-type dopamine receptor activation in nucleus accumbens shell. Horm Behav. 2003b;44:37. [Google Scholar]
- Bachtell RK, Weitemier AZ, Galvan-Rosas A, Tsivkovskaia NO, Risinger FO, Phillips TJ, Grahame NJ, Ryabinin AE. The Edinger-Westphal-lateral septum urocortin pathway and its relationship to alcohol consumption. J Neurosci. 2003;23(6):2477–87. doi: 10.1523/JNEUROSCI.23-06-02477.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bale TL, Contarino A, Smith GW, Chan R, Gold LH, Sawchenko PE, Koob GF, Vale WW, Lee KF. Mice deficient for corticotropin-releasing hormone receptor-2 display anxiety-like behaviour and are hypersensitive to stress. Nat Genet. 2000;24(4):410–4. doi: 10.1038/74263. [DOI] [PubMed] [Google Scholar]
- Bale TL, Vale WW. Increased depression-like behaviors in corticotropin-releasing factor receptor-2-deficient mice: sexually dichotomous responses. J Neurosci. 2003;23(12):5295–301. doi: 10.1523/JNEUROSCI.23-12-05295.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge KK, Pecine S, Schulkin J. Appetitive effect of CRF in accumbens shell on conditioned approach for sucrose reward. Society For Neuroscience Abstract Viewer/Itinerary Planner. 2004:437.12. [Google Scholar]
- Bielsky IF, Hu SB, Szegda KL, Westphal H, Young LJ. Profound impairment in social recognition and reduction in anxiety-like behavior in vasopressin V1a receptor knockout mice. Neuropsychopharmacology. 2004;29(3):483–93. doi: 10.1038/sj.npp.1300360. [DOI] [PubMed] [Google Scholar]
- Bosch OJ, Nair HP, Neumann ID, Young LJ. Depressive-like behavior following isolation from a female partner is associated with altered brain CRF mRNA and HPA axis activity in the male prairie vole. Society For Neuroscience Abstract Viewer/Itinerary Planner; 2005. (Program No. 420.4) [Google Scholar]
- Carter CS, DeVries AC, Getz LL. Physiological substrates of mammalian monogamy: the prairie vole model. Neurosci Biobehav Rev. 1995;19(2):303–14. doi: 10.1016/0149-7634(94)00070-h. [DOI] [PubMed] [Google Scholar]
- Carter CS, DeVries AC, Taymans SE, Roberts RL, Williams JR, Getz LL. Peptides, steroids, and pair bonding. Ann N Y Acad Sci. 1997;807:260–72. doi: 10.1111/j.1749-6632.1997.tb51925.x. [DOI] [PubMed] [Google Scholar]
- DeVries AC, DeVries MB, Taymans SE, Carter CS. The effects of stress on social preferences are sexually dimorphic in prairie voles. Proc Natl Acad Sci U S A. 1996;93(21):11980–4. doi: 10.1073/pnas.93.21.11980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeVries AC, Guptaa T, Cardillo S, Cho M, Carter CS. Corticotropin-releasing factor induces social preferences in male prairie voles. Psychoneuroendocrinology. 2002;27(6):705–14. doi: 10.1016/s0306-4530(01)00073-7. [DOI] [PubMed] [Google Scholar]
- Ferguson JN, Young LJ, Hearn EF, Matzuk MM, Insel TR, Winslow JT. Social amnesia in mice lacking the oxytocin gene. Nat Genet. 2000;25(3):284–8. doi: 10.1038/77040. [DOI] [PubMed] [Google Scholar]
- Getz LL, Carter CS, Gavish L. The mating system of the prairie vole Microtus ochragaster: Field and laboratory evidence for pair-bonding. Behavioral Ecology and Sociobiology. 1981;8:189–194. [Google Scholar]
- Gruder-Adams S, Getz LL. Comparison of the mating system and paternal behavior in Microtus ochragaster and M. pennsylvanicus. Journal of Mammalogy. 1985;66(1):165–167. [Google Scholar]
- Hotta M, Shibasaki T, Arai K, Demura H. Corticotropin-releasing factor receptor type 1 mediates emotional stress-induced inhibition of food intake and behavioral changes in rats. Brain Res. 1999;823(1–2):221–5. doi: 10.1016/s0006-8993(99)01177-4. [DOI] [PubMed] [Google Scholar]
- Insel TR, Shapiro LE. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proc Natl Acad Sci U S A. 1992;89(13):5981–5. doi: 10.1073/pnas.89.13.5981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel TR, Wang ZX, Ferris CF. Patterns of brain vasopressin receptor distribution associated with social organization in microtine rodents. J Neurosci. 1994;14(9):5381–92. doi: 10.1523/JNEUROSCI.14-09-05381.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jahn O, Eckart K, Brauns O, Tezval H, Spiess J. The binding protein of corticotropin-releasing factor: ligand-binding site and subunit structure. Proc Natl Acad Sci U S A. 2002;99(19):12055–60. doi: 10.1073/pnas.192449299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landgraf R, Gerstberger R, Montkowski A, Probst JC, Wotjak CT, Holsboer F, Engelmann M. V1 vasopressin receptor antisense oligodeoxynucleotide into septum reduces vasopressin binding, social discrimination abilities, and anxiety-related behavior in rats. J Neurosci. 1995;15(6):4250–8. doi: 10.1523/JNEUROSCI.15-06-04250.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liebsch G, Wotjak CT, Landgraf R, Engelmann M. Septal vasopressin modulates anxiety-related behaviour in rats. Neurosci Lett. 1996;217(2–3):101–4. [PubMed] [Google Scholar]
- Lim MM, Murphy AZ, Young LJ. Ventral striatopallidal oxytocin and vasopressin V1a receptors in the monogamous prairie vole (Microtus ochrogaster) J Comp Neurol. 2004a;468(4):555–70. doi: 10.1002/cne.10973. [DOI] [PubMed] [Google Scholar]
- Lim MM, Nair HP, Young LJ. Species and sex differences in brain distribution of corticotropin-releasing factor receptor subtypes 1 and 2 in monogamous and promiscuous vole species. J Comp Neurol. 2005;487(1):75–92. doi: 10.1002/cne.20532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim MM, Tsivkovskaia NO, Bai Y, Young LJ, Ryabinin AE. Distribution of Corticotropin-Releasing Factor and Urocortin 1 in the Vole Brain. Brain Behav Evol. 2006;68(4):229–240. doi: 10.1159/000094360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim MM, Wang Z, Olazabal DE, Ren X, Terwilliger EF, Young LJ. Enhanced partner preference in a promiscuous species by manipulating the expression of a single gene. Nature. 2004b;429(6993):754–7. doi: 10.1038/nature02539. [DOI] [PubMed] [Google Scholar]
- Lim MM, Young LJ. Vasopressin-dependent neural circuits underlying pair bond formation in the monogamous prairie vole. Neuroscience. 2004;125(1):35–45. doi: 10.1016/j.neuroscience.2003.12.008. [DOI] [PubMed] [Google Scholar]
- Liu Y, Wang ZX. Nucleus accumbens oxytocin and dopamine interact to regulate pair bond formation in female prairie voles. Neuroscience. 2003;121(3):537–44. doi: 10.1016/s0306-4522(03)00555-4. [DOI] [PubMed] [Google Scholar]
- Lu L, Liu Z, Huang M, Zhang Z. Dopamine-dependent responses to cocaine depend on corticotropin-releasing factor receptor subtypes. J Neurochem. 2003;84(6):1378–86. doi: 10.1046/j.1471-4159.2003.01635.x. [DOI] [PubMed] [Google Scholar]
- Mantella RC, Vollmer RR, Li X, Amico JA. Female oxytocin-deficient mice display enhanced anxiety-related behavior. Endocrinology. 2003;144(6):2291–6. doi: 10.1210/en.2002-0197. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 4. Academic Press; 1998. [DOI] [PubMed] [Google Scholar]
- Primus RJ, Yevich E, Baltazar C, Gallager DW. Autoradiographic localization of CRF1 and CRF2 binding sites in adult rat brain. Neuropsychopharmacology. 1997;17(5):308–16. doi: 10.1016/S0893-133X(97)00071-7. [DOI] [PubMed] [Google Scholar]
- Ring RH, Malberg JE, Potestio L, Ping J, Boikess S, Luo B, Schechter LE, Rizzo S, Rahman Z, Rosenzweig-Lipson S. Anxiolytic-like activity of oxytocin in male mice: behavioral and autonomic evidence, therapeutic implications. Psychopharmacology (Berl) 2006:1–8. doi: 10.1007/s00213-005-0293-z. [DOI] [PubMed] [Google Scholar]
- Ryabinin AE, Criado JR, Henriksen SJ, Bloom FE, Wilson MC. Differential sensitivity of c-Fos expression in hippocampus and other brain regions to moderate and low doses of alcohol. Mol Psychiatry. 1997;2(1):32–43. doi: 10.1038/sj.mp.4000206. [DOI] [PubMed] [Google Scholar]
- Salo AL, Shapiro LE, Dewsbury DA. Affiliative behavior in different species of voles (Microtus) Psychol Rep. 1993;72(1):316–8. doi: 10.2466/pr0.1993.72.1.316. [DOI] [PubMed] [Google Scholar]
- Shapiro LE, Dewsbury DA. Differences in affiliative behavior, pair bonding, and vaginal cytology in two species of vole (Microtus ochrogaster and M. montanus) J Comp Psychol. 1990;104(3):268–74. doi: 10.1037/0735-7036.104.3.268. [DOI] [PubMed] [Google Scholar]
- Ungless MA, Singh V, Crowder TL, Yaka R, Ron D, Bonci A. Corticotropin-releasing factor requires CRF binding protein to potentiate NMDA receptors via CRF receptor 2 in dopamine neurons. Neuron. 2003;39(3):401–7. doi: 10.1016/s0896-6273(03)00461-6. [DOI] [PubMed] [Google Scholar]
- Weitemier AZ, Tsivkovskaia NO, Ryabinin AE. Urocortin 1 distribution in mouse brain is strain-dependent. Neuroscience. 2005;132(3):729–40. doi: 10.1016/j.neuroscience.2004.12.047. [DOI] [PubMed] [Google Scholar]
- Windle RJ, Shanks N, Lightman SL, Ingram CD. Central oxytocin administration reduces stress-induced corticosterone release and anxiety behavior in rats. Endocrinology. 1997;138(7):2829–34. doi: 10.1210/endo.138.7.5255. [DOI] [PubMed] [Google Scholar]
- Young LJ, Lim MM, Gingrich B, Insel TR. Cellular mechanisms of social attachment. Horm Behav. 2001;40(2):133–8. doi: 10.1006/hbeh.2001.1691. [DOI] [PubMed] [Google Scholar]
- Young LJ, Wang Z. The neurobiology of pair bonding. Nat Neurosci. 2004;7(10):1048–54. doi: 10.1038/nn1327. [DOI] [PubMed] [Google Scholar]